

Amphibian Skin and Skin Secretion: An Exotic Source of Bioactive Peptides and Its Application
 ,
,  and
and 
Abstract
:1. Introduction
2. Amphibians Production and Nutritional Composition
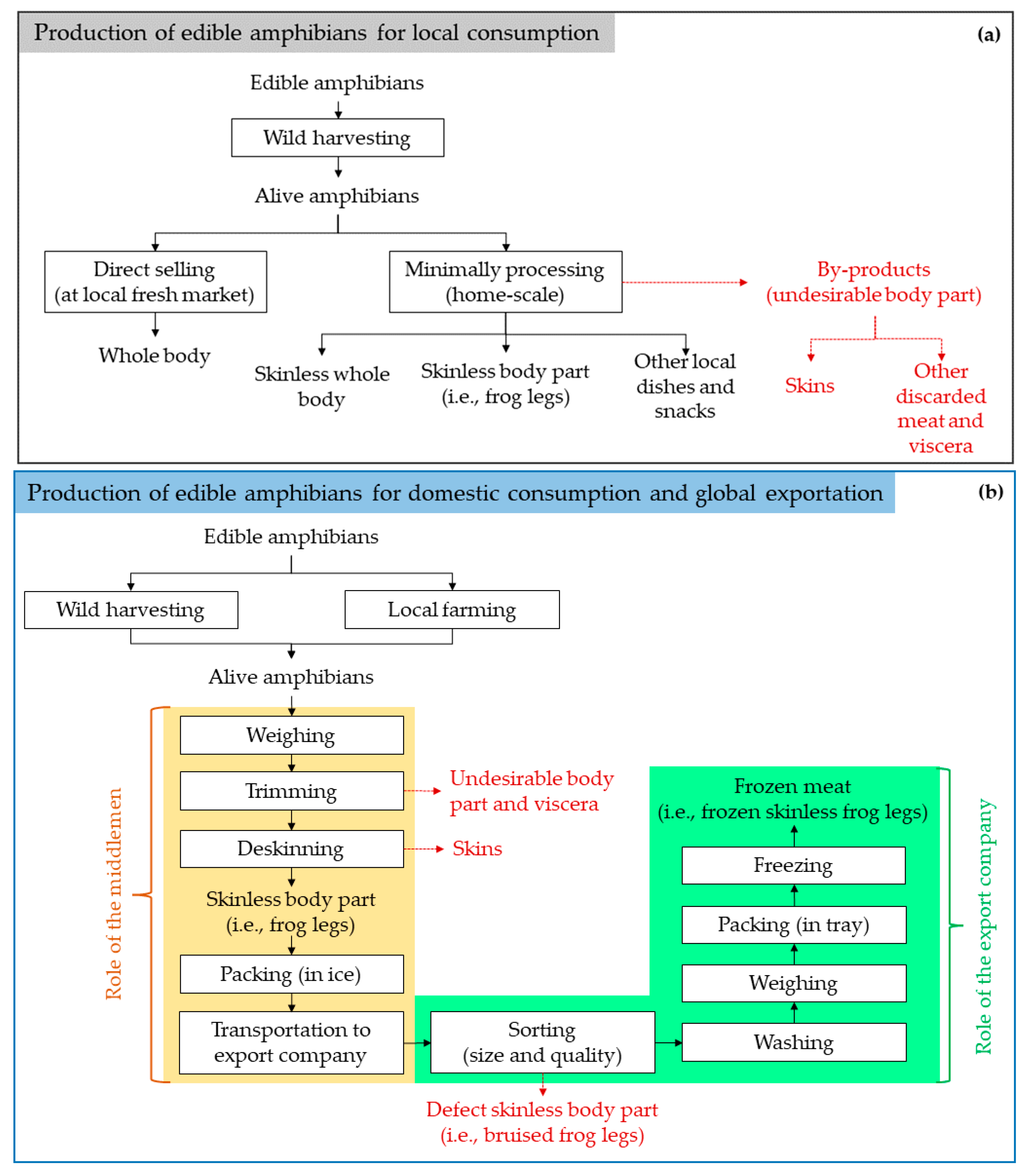
2.1. Production Status
2.2. Nutritional Composition
3. Bioactivities of Peptides Derived from Amphibian Skin
3.1. Antioxidant Activity
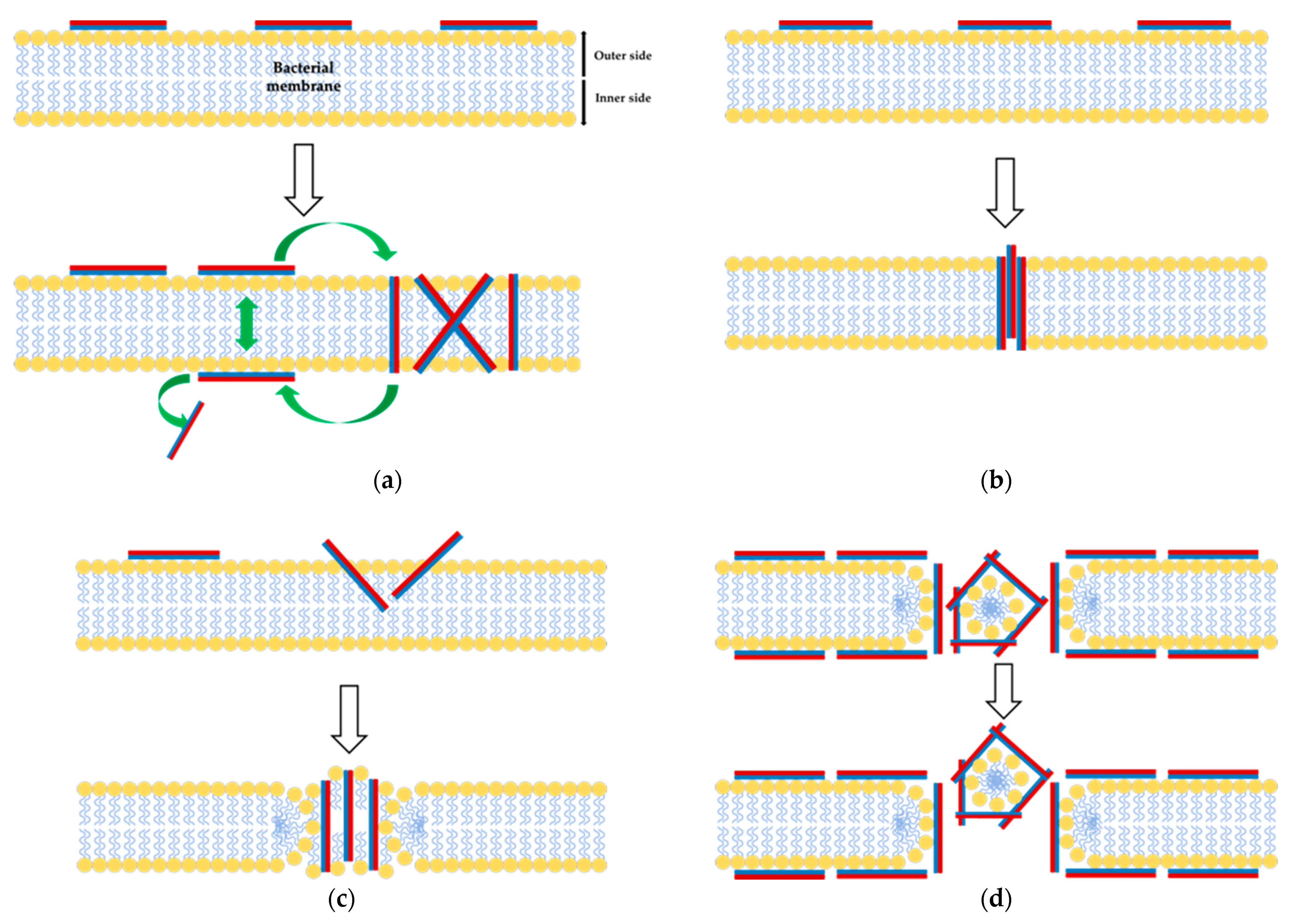
3.2. Antimicrobial Activity
3.3. Anticancer Activity

{kind=link}
{kind=link}
{kind=link}
| Peptides | Species (Common name) | Assay(s): Cell Lines | Amino Acid Sequence | Reference |
|---|---|---|---|---|
| Brevinin-2R | Rana ridibunda (Marsh frog) | MTT assay (in vitro): Jurkat T-cell leukemia, BJAB B-cell lymphoma, MCF-7 breast adenocarcinoma, L929 fibrosarcoma, A549 lung carcinoma. | KLKNFAKGVAQSLLNKASCKLSGQC | [50] |
| Brevinin-1Da | Rana dalmatina (European frog) | – | IIPLLLGKVVCAITKKC | [49] |
| Brevinin-1RL1 | Rana limnocharis (Asian rice frog) | MTS assay (in vitro): HCT116 colorectal adenocarcinoma (LD50: 5.9 ± 0.2 μM), MDA-MB-231 breast adenocarcinoma (LD50: 5.4 ± 0.3 μM), SW480 colorectal adenocarcinoma (LD50: 10.4 ± 0.4 μM), A549 lung adenocarcinoma (LD50: 5.8 ± 0.2 μM), SMMC-7721 hepatocellular carcinoma (LD50: 6.9 ± 0.5 μM), B16-F10 melanomas (LD50: 6.6 ± 0.3 μM), NCM460 colon mucosal epithelial (LD50: 16.8 ± 0.6 μM), BEAS-2B bronchial epithelial (LD50: 16.6 ± 0.3 μM), HaCaT keratinocyte cell (LD50: 28.7 ± 0.4 μM). | FFPLIAGLAARFLPKIFCSITKRC | [92] |
| Dermaseptin-B2 | Phyllomedusa bicolor (South American tree frog) | Proliferation and angiogenesis assays (in vitro): PC-3 human prostatic adenocarcinoma (EC50: 2 μM), NIH-3T3 non-tumor mouse (na.). | GLWSKIKEVGKEAAKAAAKAAGKAAGAVSEAV-CONH2 | [93] |
| Dermaseptin-B3 | Phyllomedusa bicolor (South American tree frog) | Proliferation and angiogenesis assays (in vitro): PC-3 human prostatic adenocarcinoma (EC50: 3 μM), NIH-3T3 non-tumor mouse (EC50: 0.2–1 µM). | ALWKNMLKGIGKLAGQAALGAVK TLVG-COOH | [93] |
| Dermaseptin-PP | Phyllomedusa palliata | MTT assay (in vitro): H157 human non-small cell lung cancer (IC50: 1.6 μM), MCF-7 human breast adenocarcinoma (IC50: 2.9 μM), PC-3 human prostate carcinoma (IC50: 4.2 μM), U251 MG human neuronal glioblastoma (IC50: 2.5 μM). MTT assay (in vivo): H157 tumor in nude mice (at 10−4 M induced ~80% LDH release). | ALWKDMLKGIGKLAGKAALGAVKTLV-NH2 | [89] |
| Hymenochirin-1B | Hymennochirus boettgeri (Congo dwarf clawed frog) | MTT assay (in vitro): HepG2 and PLC human hepatocellular carcinoma cells; NCIH1299, A549 and H460 human lung cancer cells. Cytotoxicity assay (in vitro): A549 lung adenocarcinoma (LD50: 2.5 ± 0.2 μM), MDA-MB-231 breast adenocarcinoma (LD50: 9.0 ± 0.3 μM), HT-29 colorectal adenocarcinoma (LD50: 9.7 ± 0.2 μM), Hep-G2 hepatocarcinoma (LD50: 22.5 ± 1.4 μM). | IKLSPETKDNLKKVLKGAIKGAIAVAKMV | [94,95] |
| Pseudhymenochirin-2Pa | Pseudhymenochirus merlini (Merlin’s dwarf gray frog) | Cytotoxicity assay (in vitro): A549 lung adenocarcinoma (LD50: 6.0 ± 0.6 μM), MDA-MB-231 breast adenocarcinoma (LD50: 6.2 ± 0.6 μM), HT-29 colorectal adenocarcinoma (LD50: 11.5 ± 2.6 μM), HUVEC colorectal adenocarcinoma (LD50: 68 ± 2 μM). | GIFPIFAKLLGKVIKVASSLISKGRTE | [12] |
| Pseudhymenochirin-1Pb | Pseudhymenochirus merlini (Merlin’s dwarf gray frog) | Cytotoxicity assay (in vitro): A549 lung adenocarcinoma (LD50: 2.5 ± 0.2 μM), MDA-MB-231 breast adenocarcinoma (LD50: 6.6 ± 0.3 μM), HT-29 colorectal adenocarcinoma (LD50: 9.5 ± 1.3 μM), HUVEC colorectal adenocarcinoma (LD50: 5.6 ± 0.9 μM). | IKIPSFFRNILKKVGKEAVSLIAGALKQS | [12] |
| Temporin-1CEa | Rana chensinensis (Chinese brown frog) | MTT assay (in vitro): SMMC-7721 human hepatocellular carcinoma (LD50: 44.9–50.3 μM), BEL-7402 human hepatocellular carcinoma (LD50: 36.9–40.2 μM), Bcap-37 human breast carcinoma (LD50: 37.3–39.4 μM), MDA-MB-231 human breast Caucasian adenocarcinoma (LD50: 55.0–63.3 μM), MCF-7 human breast carcinoma (LD50: 27.8–34.5 μM), LK-2 human squamous cell carcinoma (LD50: 58.3–63.4 μM), A-549 human lung adenocarcinoma (LD50: 52.1–53.3 μM), NCIH446 human small cell lung carcinoma (LD50: 59.4–67.7 μM), BGC-823 human gastric carcinoma (LD50: 55.2–63.2 μM), Hela human cervical carcinoma (LD50: 36.3–43.9), HO-8910 human ovarian carcinoma (LD50: 62.8–66.7 μM), HT-29 human colon carcinoma (LD50: >88.91 μM). Hemolysis assay (in vitro): healthy human blood cell (LD50: 99.08 μM). LDH assay (in vitro): MCF-7 breast cancer (LD50: 40.2–49.1 μM). | FVDLKKIANIINSIF-NH2 | [90] |
| XLAsp-P1 | Xenopus laevis (African clawed frog) | MTT assay (in vitro): MCF-7 breast cancer (LD50: <5 μg/mL). | DEDDD | [91] |
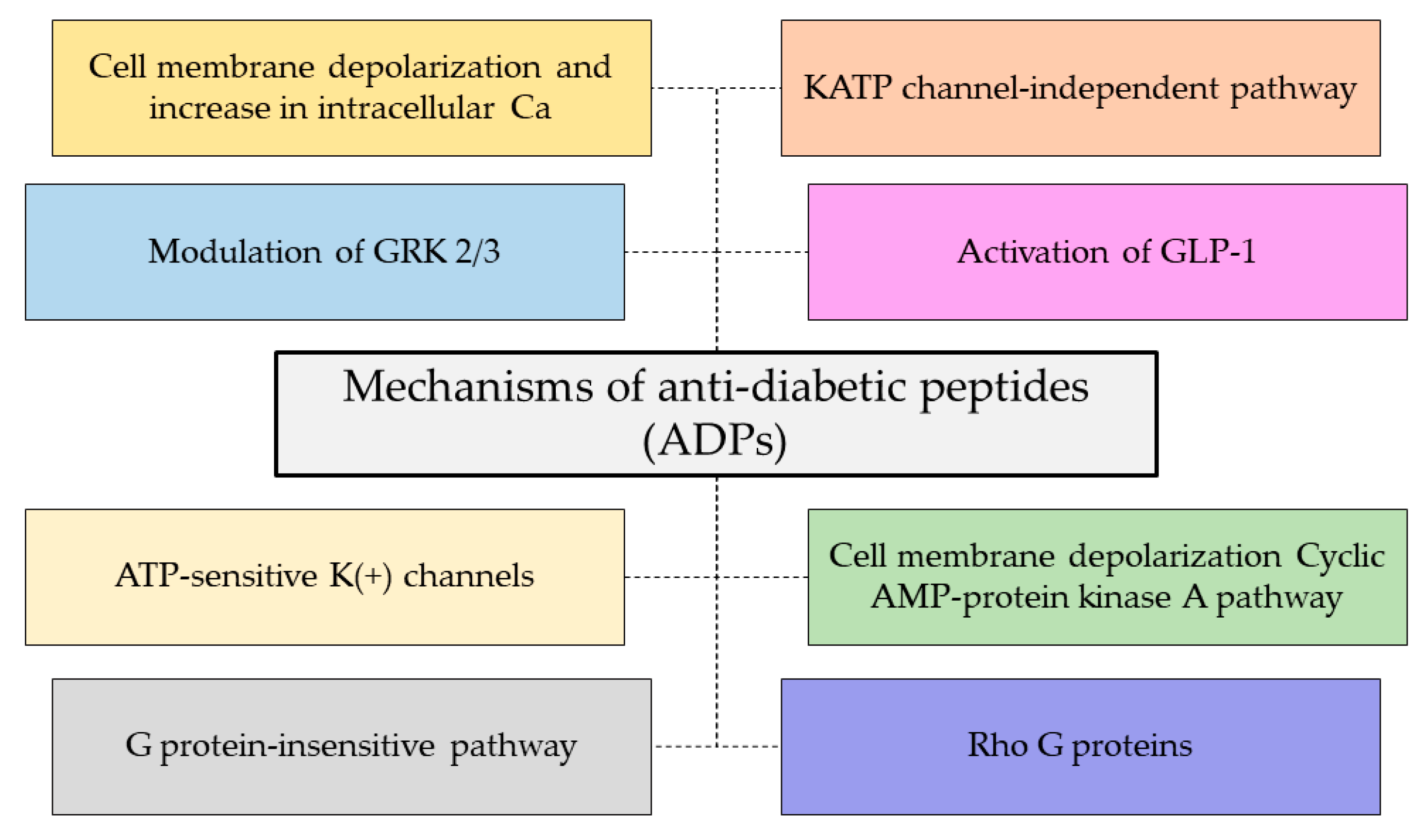
3.4. Antidiabetic Activity
3.5. Other Bioactivities
4. Multiple Constraints toward Amphibian Skin Derivatives
5. Prospective Application in Food and Biomedical Industries
5.1. Application in Food Industry
5.2. Application in Biomedical Industry
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adil, S.; Altaf, M.; Hussain, T.; Umair, M.; Ni, J.; Abbasi, A.M.; Bussmann, R.W.; Ashraf, S. Cultural and medicinal use of amphibians and reptiles by indigenous people in Punjab, Pakistan with comments on conservation implications for herpetofauna. Animals 2022, 12, 2062. [Google Scholar] [CrossRef]
- Bazaz, M.R.; Mashreghi, M.; Shahri, N.M.; Mashreghi, M.; Asoodeh, A.; Rassouli, M.B.; Golmohammadzadeh, S. Pharmaceutical application of frog skin on full-thickness skin wound healing in mice. Pharm. Biol. 2013, 51, 1600–1606. [Google Scholar] [CrossRef] [Green Version]
- Burggren, W.W.; Warburton, S. Amphibians as animal models for laboratory research in physiology. ILAR J. 2007, 48, 260–269. [Google Scholar] [CrossRef]
- Turvey, S.T.; Chen, S.; Tapley, B.; Liang, Z.; Wei, G.; Yang, J.; Wang, J.; Wu, M.; Redbond, J.; Brown, T.; et al. From dirty to delicacy? Changing exploitation in China threatens the world’s largest amphibians. People Nat. 2021, 3, 446–456. [Google Scholar] [CrossRef]
- Sai, K.P.; Jagannadham, M.V.; Vairamani, M.; Raju, N.P.; Devi, A.S.; Nagaraj, R.; Sitaram, N. Tigerinins: Novel antimicrobial peptides from the Indian frog Rana tigerina. J. Biol. Chem. 2001, 276, 2701–2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coad, L.; Fa, J.E.; Abernethy, K.; Van Vliet, N.S.C.; Wilkie, D.; El Bizri, H.R.; Ingram, D.J.; Cawthorn, D.-M.; Nasi, R. Towards a Sustainable, Participatory and Inclusive Wild Meat Sector; Center for International Forestry Research (CIFOR): Bogor, Indonesia, 2019. [Google Scholar]
- He, D.; Zhu, W.; Zeng, W.; Lin, J.; Ji, Y.; Wang, Y.; Zhang, C.; Lu, Y.; Zhao, D.; Su, N.; et al. Nutritional and medicinal characteristics of Chinese giant salamander (Andrias davidianus) for applications in healthcare industry by artificial cultivation: A review. Food Sci. Hum. Wellness 2018, 7, 1–10. [Google Scholar] [CrossRef]
- Drakulić, S.; Feldhaar, H.; Lisičić, D.; Mioč, M.; Cizelj, I.; Seiler, M.; Spatz, T.; Rödel, M. Population-specific effects of developmental temperature on body condition and jumping performance of a widespread European frog. Ecol. Evol. 2016, 6, 3115–3128. [Google Scholar] [CrossRef] [Green Version]
- Indriani, S.; Karnjanapratum, S.; Nalinanon, S. Production of collagen hydrolysates from Asian bullfrog (Rana tigerina) skin using papain. In Proceedings of the the 6th International Conference on Food and Applied Bioscience 2022, Chiang Mai, Thailand, 14–15 February 2022; pp. 259–268. [Google Scholar]
- Qian, Z.-J.; Jung, W.-K.; Kim, S.-K. Free radical scavenging activity of a novel antioxidative peptide purified from hydrolysate of bullfrog skin, Rana catesbeiana Shaw. Bioresour. Technol. 2008, 99, 1690–1698. [Google Scholar] [CrossRef]
- Clark, D.; Durell, S.; Maloy, W.; Zasloff, M. Ranalexin. A novel antimicrobial peptide from bullfrog (Rana catesbeiana) skin, structurally related to the bacterial antibiotic, polymyxin. J. Biol. Chem. 1994, 269, 10849–10855. [Google Scholar] [CrossRef]
- Mechkarska, M.; Attoub, S.; Sulaiman, S.; Pantic, J.; Lukic, M.L.; Conlon, J.M. Anti-cancer, immunoregulatory, and antimicrobial activities of the frog skin host-defense peptides pseudhymenochirin-1Pb and pseudhymenochirin-2Pa. Regul. Pept. 2014, 194–195, 69–76. [Google Scholar] [CrossRef]
- Liu, H.; Mu, L.; Tang, J.; Shen, C.; Gao, C.; Rong, M.; Zhang, Z.; Liu, J.; Wu, X.; Yu, H.; et al. A potential wound healing-promoting peptide from frog skin. Int. J. Biochem. Cell Biol. 2014, 49, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Ojo, O.; Conlon, J.; Flatt, P.; Abdel-Wahab, Y. Frog skin peptides (tigerinin-1R, magainin-AM1, -AM2, CPF-AM1, and PGla-AM1) stimulate secretion of glucagon-like peptide 1 (GLP-1) by GLUTag cells. Biochem. Biophys. Res. Commun. 2013, 431, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, D.; Li, S.; Liu, B.; Gao, L. Sustained release of BSA from a novel drug delivery matrix – bullfrog skin collagen film. Macromol. Biosci. 2004, 4, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M. Chapter 45—Bradykinin Peptides. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 321–325. [Google Scholar]
- Spindel, E.R. Chapter 46—Bombesin Peptides. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 326–330. [Google Scholar]
- Luca, V.; Barra, D.; Mangoni, M.L. Chapter 47—Bombinins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 331–337. [Google Scholar]
- Bowie, J.H. Chapter 48—Caeruleins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 338–341. [Google Scholar]
- Montero-Hadjadje, M.; Delestre, C.; Courel, M.; Anouar, Y. Chapter 49—Chromogranins/Secretogranins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 342–349. [Google Scholar]
- Nicolas, P.; Ladram, A. Chapter 50—Dermaseptins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 350–363. [Google Scholar]
- Severini, C.; Improta, G. Chapter 55—Tachykinins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 391–399. [Google Scholar]
- Rinaldi, A.C.; Conlon, J.M. Chapter 56—Temporins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 400–406. [Google Scholar]
- Ojo, O.O.; Srinivasan, D.K.; Owolabi, B.O.; Flatt, P.R.; Abdel-Wahab, Y.H. Beneficial effects of tigerinin-1R on glucose homeostasis and beta cell function in mice with diet-induced obesity-diabetes. Biochimie 2015, 109, 18–26. [Google Scholar] [CrossRef]
- Karnjanapratum, S.; Benjakul, S. Asian bullfrog (Rana tigerina) skin gelatin extracted by ultrasound-assisted process: Characteristics and in-vitro cytotoxicity. Int. J. Biol. Macromol. 2020, 148, 391–400. [Google Scholar] [CrossRef]
- Chen, X.; Liu, S.; Fang, J.; Zheng, S.; Wang, Z.; Jiao, Y.; Xia, P.; Wu, H.; Ma, Z.; Hao, L. Peptides isolated from amphibian skin secretions with emphasis on antimicrobial peptides. Toxins 2022, 14, 722. [Google Scholar] [CrossRef]
- Plácido, A.; Bueno, J.; Barbosa, E.A.; Moreira, D.C.; Dias, J.D.N.; Cabral, W.F.; Albuquerque, P.; Bessa, L.J.; Freitas, J.; Kuckelhaus, S.A.S.; et al. The antioxidant peptide salamandrin-I: First bioactive peptide identified from skin secretion of Salamandra genus (Salamandra salamandra). Biomolecules 2020, 10, 512. [Google Scholar] [CrossRef] [Green Version]
- Schloegel, L.M.; Picco, A.M.; Kilpatrick, A.M.; Davies, A.J.; Hyatt, A.D.; Daszak, P. Magnitude of the US trade in amphibians and presence of Batrachochytrium dendrobatidis and ranavirus infection in imported North American bullfrogs (Rana catesbeiana). Biol. Conserv. 2009, 142, 1420–1426. [Google Scholar] [CrossRef]
- Efenakpo, O. Frog consumption pattern in Ibadan, Nigeria. JSMaP 2015, 1, 522–531. [Google Scholar]
- Cunningham, A.A.; Turvey, S.T.; Zhou, F.; Meredith, H.M.R.; Guan, W.; Liu, X.; Sun, C.; Wang, Z.; Wu, M. Development of the Chinese giant salamander Andrias davidianus farming industry in Shaanxi province, China: Conservation threats and opportunities. Oryx 2015, 50, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Diesmos, A.; Brown, R.; Gee, G. Preliminary report on the amphibians and reptiles of Balbalasang-Balbalan National Park, Luzon Island, Philippines. Sylvatrop 2004, 13, 63–80. [Google Scholar]
- Pariyanonth, P.; Daorerk, V. Frog farming in Thailand. In Proceedings of the INFOFISH-AQUATECH ´94 International Conference on Aquaculture, Colombo, Sri Lanka, 29–31 August 1994; pp. 126–130. [Google Scholar]
- Mohneke, M. (Un)Sustainable Use of Frogs in West Africa and Resulting Consequences for the Ecosystem; Humboldt University: Berlin, Germany, 2011. [Google Scholar]
- Kusrini, M.D.; Alford, R.A. Indonesia’s exports of frogs’ legs. TRAFFIC Bull. 2006, 21, 13–24. [Google Scholar]
- Gratwicke, B.; Evans, M.J.; Jenkins, P.T.; Kusrini, M.D.; Moore, R.D.; Sevin, J.; Wildt, D.E. Is the international frog legs trade a potential vector for deadly amphibian pathogens? Front. Ecol. Environ. 2010, 8, 438–442. [Google Scholar] [CrossRef]
- Onadeko, A.B.; Egonmwan, R.I.; Saliu, J.K. Edible amphibian species: Local knowledge of their consumption in southwest Nigeria and their nutritional value. West Afr. J. Appl. Ecol. 2011, 19, 67–76. [Google Scholar]
- Şimşek, E.; Alkaya, A.; Şereflişan, H.; Ozyilmaz, A. Comparisons of biochemical compositions in marsh frog (Pelophylax ridibundus) (Anura; Ranidae) grown in different conditions; wild, semicultured and cultured ones. Turk. J. Zool. 2022, 46, 261–269. [Google Scholar] [CrossRef]
- Mathew, J.T.; Ndamito, M.M.; Shaba, E.Y.; Mohammed, S.S.; Salihu, A.B.; Abu, Y. Determination of the nutritive and anti-nutritive values of Pelophylax esculentus (edible frog) found in Hanyan Gwari, Minna Niger state, Nigeria. Adv. Res. 2015, 4, 412–420. [Google Scholar] [CrossRef]
- Schairer, M.L.; Dierenfeld, E.S.; Fitzpatrick, M.P. Nutrient composition of whole green frogs, Rana clamitans and Southern toads, Bufo terrestris. Bull. Assoc. Reptil. Amphib. Veter 1998, 8, 17–20. [Google Scholar] [CrossRef]
- Oyibo, S.O.; Akani, G.C.; Amuzie, C.C. Nutritional and serum biochemistry of the edible frog Hoplobatrachus occipitalis in rivers state, Nigeria. Asian J. Res. Zool. 2020, 3, 35–41. [Google Scholar] [CrossRef]
- Daniel, U.I.; Vincent-Akpu, I.F.; Umesi, N.; Togi, P.D. Comparative study of the proximate composition of Pyxicephalus adspersus and Oreochromis niloticus from Nigerian wetland. Int. J. Curr. Res. 2016, 8, 42680–42685. [Google Scholar]
- Zhu, Y.; Bao, M.; Chen, C.; Yang, X.; Yan, W.; Ren, F.; Wang, P.; Wen, P. Comparison of the nutritional composition of bullfrog meat from different parts of the animal. Korean J. Food Sci. Anim. Resour. 2021, 41, 1049–1059. [Google Scholar] [CrossRef]
- Özogul, F.; Özogul, Y.; Olgunoglu, A.I.; Boga, E.K. Comparison of fatty acid, mineral and proximate composition of body and legs of edible frog (Rana esculenta). Int. J. Food Sci. Nutr. 2008, 59, 558–565. [Google Scholar] [CrossRef]
- Zhang, J.; Duan, R. Characterisation of acid-soluble and pepsin-solubilised collagen from frog (Rana nigromaculata) skin. Int. J. Biol. Macromol. 2017, 101, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.-G.; Pei, J.; Du, Y.-N.; Pan, J.; Gao, R.; Chen, D.-J.; Wu, H.-T.; Zhu, B.-W. Characterization and functional properties of gelatin extracted from Chinese giant salamander (Andrias Davidianus) Skin. J. Aquat. Food Prod. Technol. 2019, 28, 861–876. [Google Scholar] [CrossRef]
- Wang, X.; Ren, S.; Guo, C.; Zhang, W.; Zhang, X.; Zhang, B.; Li, S.; Ren, J.; Hu, Y.; Wang, H. Identification and functional analyses of novel antioxidant peptides and antimicrobial peptides from skin secretions of four East Asian frog species. Acta Biochim. Et Biophys. Sin. 2017, 49, 550–559. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Wang, Y.; Wu, C.; Li, X.; Fu, Z.; Yang, M.; Bian, W.; Wang, S.; Song, Y.; Tang, J.; et al. Cathelicidin-OA1, a novel antioxidant peptide identified from an amphibian, accelerates skin wound healing. Sci. Rep. 2018, 8, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: Biodiversity and therapeutic promises. Front. Biosci. 2016, 21, 1341–1371. [Google Scholar] [CrossRef] [Green Version]
- Conlon, J.; Seidel, B.; Nielsen, P.F. An atypical member of the brevinin-1 family of antimicrobial peptides isolated from the skin of the European frog Rana dalmatina. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2004, 137, 191–196. [Google Scholar] [CrossRef]
- Ghavami, S.; Asoodeh, A.; Klonisch, T.; Halayko, A.J.; Kadkhoda, K.; Kroczak, T.J.; Gibson, S.B.; Booy, E.P.; Naderi-Manesh, H.; Los, M. Brevinin-2R(1) semi-selectively kills cancer cells by a distinct mechanism, which involves the lysosomal-mitochondrial death pathway. J. Cell. Mol. Med. 2008, 12, 1005–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musale, V.; Moffett, R.C.; Owolabi, B.; Conlon, J.M.; Flatt, P.R.; Abdel-Wahab, Y.H.A. Mechanisms of action of the antidiabetic peptide [S4K]CPF-AM1 in db/db mice. J. Mol. Endocrinol. 2021, 66, 115–128. [Google Scholar] [CrossRef]
- Indriani, S.; Sae-Leaw, T.; Benjakul, S.; Quan, T.H.; Karnjanapratum, S.; Nalinanon, S. Impact of different ultrasound-assisted processes for preparation of collagen hydrolysates from Asian bullfrog skin on characteristics and antioxidative properties. Ultrason. Sonochemistry 2022, 89, 106163. [Google Scholar] [CrossRef] [PubMed]
- Indriani, S.; Benjakul, S.; Kishimura, H.; Karnjanapratum, S.; Nalinanon, S. Impact of extraction condition on the yield and molecular characteristics of collagen from Asian bullfrog (Rana tigerina) skin. LWT 2022, 162, 113439. [Google Scholar] [CrossRef]
- Karnjanapratum, S.; Sinthusamran, S.; Sae-Leaw, T.; Benjakul, S.; Kishimura, H. Characteristics and gel properties of gelatin from skin of Asian bullfrog (Rana tigerina). Food Biophys. 2017, 12, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Zahari, M.S.A.; Susanti, D.; Hashi, A.A. Ethical dimension of consuming frog components for medical and research purposes. Rev. Sci. 2017, 7, 15–20. [Google Scholar]
- Di Nunzio, M.; Loffi, C.; Montalbano, S.; Chiarello, E.; Dellafiora, L.; Picone, G.; Antonelli, G.; Tedeschi, T.; Buschini, A.; Capozzi, F.; et al. Cleaning the label of cured meat: Effect of the replacement of nitrates/nitrites on nutrients bioaccessibility, peptides formation, and cellular toxicity of in vitro digested salami. Int. J. Mol. Sci. 2022, 23, 12555. [Google Scholar] [CrossRef]
- Di Nunzio, M.; Loffi, C.; Chiarello, E.; Dellafiora, L.; Picone, G.; Antonelli, G.; Di Gregorio, C.; Capozzi, F.; Tedeschi, T.; Galaverna, G.; et al. Impact of a shorter brine soaking time on nutrient bioaccessibility and peptide formation in 30-months-ripened Parmigiano Reggiano cheese. Molecules 2022, 27, 664. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The Chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Rajapakse, N.; Mendis, E.; Byun, H.G.; Kim, S.K. Purification and in vitro antioxidative effects of giant squid muscle peptides on free radical-mediated oxidative systems. J. Nutr. Biochem. 2005, 9, 562–569. [Google Scholar] [CrossRef]
- Zhou, D.-Y.; Tang, Y.; Zhu, B.-W.; Qin, L.; Li, D.-M.; Yang, J.-F.; Lei, K.; Murata, Y. Antioxidant activity of hydrolysates obtained from scallop (Patinopecten yessoensis) and abalone (Haliotis discus hannai Ino) muscle. Food Chem. 2011, 132, 815–822. [Google Scholar] [CrossRef]
- Ketnawa, S.; Martínez-Alvarez, O.; Benjakul, S.; Rawdkuen, S. Gelatin hydrolysates from farmed Giant catfish skin using alkaline proteases and its antioxidative function of simulated gastro-intestinal digestion. Food Chem. 2016, 192, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Thiansilakul, Y.; Benjakul, S.; Shahidi, F. Antioxidative activity of protein hydrolysate from round scad muscle using alcalase and flavourzyme. J. Food Biochem. 2007, 31, 266–287. [Google Scholar] [CrossRef]
- Wu, H.-C.; Chen, H.-M.; Shiau, C.-Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Peng, X.; Xiong, Y.L.; Kong, B. Antioxidant activity of peptide fractions from whey protein hydrolysates as measured by electron spin resonance. Food Chem. 2009, 113, 196–201. [Google Scholar] [CrossRef]
- Liu, Q.; Kong, B.; Xiong, Y.L.; Xia, X. Antioxidant activity and functional properties of porcine plasma protein hydrolysate as influenced by the degree of hydrolysis. Food Chem. 2010, 118, 403–410. [Google Scholar] [CrossRef]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef]
- Yin, S.; Wang, Y.; Liu, N.; Yang, M.; Hu, Y.; Li, X.; Fu, Y.; Luo, M.; Sun, J.; Yang, X. Potential skin protective effects after UVB irradiation afforded by an antioxidant peptide from Odorrana andersonii. Biomed. Pharmacother. 2019, 120, 109535. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, X.; Liu, X.; Wu, J.; Liu, C.; Gong, W.; Zhao, Z.; Hong, J.; Lin, D.; Wang, Y.; et al. Antioxidant peptidomics reveals novel skin antioxidant system. Mol. Cell. Proteom. 2009, 8, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Amerikova, M.; El-Tibi, I.P.; Maslarska, V.; Bozhanov, S.; Tachkov, K. Antimicrobial activity, mechanism of action, and methods for stabilisation of defensins as new therapeutic agents. Biotechnol. Biotechnol. Equip. 2019, 33, 671–682. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [Green Version]
- Hajfathalian, M.; Ghelichi, S.; García-Moreno, P.J.; Moltke Sørensen, A.-D.; Jacobsen, C. Peptides: Production, bioactivity, functionality, and applications. Crit. Rev. Food Sci. Nutr. 2018, 58, 3097–3129. [Google Scholar] [CrossRef] [Green Version]
- Sila, A.; Nedjar-Arroume, N.; Hedhili, K.; Chataigné, G.; Balti, R.; Nasri, M.; Dhulster, P.; Bougatef, A. Antibacterial peptides from barbel muscle protein hydrolysates: Activity against some pathogenic bacteria. LWT 2013, 55, 183–188. [Google Scholar] [CrossRef]
- Abuine, R.; Rathnayake, A.U.; Byun, H.-G. Biological activity of peptides purified from fish skin hydrolysates. Fish. Aquat. Sci. 2019, 22, 10. [Google Scholar] [CrossRef] [Green Version]
- Goraya, J.; Knoop, F.C.; Conlon, J.M. Ranatuerin 1T: An antimicrobial peptide isolated from the skin of the frog, Rana temporaria. Peptides 1999, 20, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Dorica-Mirela, S.; Ionel, J. Biologically active natural peptides. J. Agroaliment. Process. Technol. 2009, 15, 484–499. [Google Scholar]
- Savelyeva, A.; Ghavami, S.; Davoodpour, P.; Asoodeh, A.; Los, M.J. An overview of brevinin superfamily: Structure, function and clinical perspectives. Adv. Exp. Med. Biol. 2014, 818, 197–212. [Google Scholar]
- Park, S.; Park, S.H.; Ahn, H.C.; Kim, S.; Kim, S.S.; Lee, B.J.; Lee, B.J. Structural study of novel antimicrobial peptides, nigrocins, isolated from Rana nigromaculata. FEBS Lett. 2001, 507, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.B.; Yi, K.-S.; Matsuzaki, K.; Kim, M.S.; Kim, S.C. Structure-activity analysis of buforin II, a histone H2A-derived antimicrobial peptide: The proline hinge is responsible for the cell-penetrating ability of buforin II. Proc. Natl. Acad. Sci. USA 2000, 97, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Savoia, D.; Guerrini, R.; Marzola, E.; Salvadori, S. Synthesis and antimicrobial activity of dermaseptin S1 analogues. Bioorganic Med. Chem. 2008, 16, 8205–8209. [Google Scholar] [CrossRef] [Green Version]
- Conlon, J.M.; Leprince, J.; Vaudry, H.; Jiansheng, H.; Nielsen, P.F. A family of antimicrobial peptides related to japonicin-2 isolated from the skin of the Chaochiao brown frog Rana chaochiaoensis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 144, 101–105. [Google Scholar] [CrossRef]
- Dennison, S.R.; Morton, L.H.; Harris, F.; Phoenix, D.A. Low pH enhances the action of maximin H5 against Staphylococcus aureus and helps mediate lysylated phosphatidylglycerol-induced resistance. Biochemistry 2016, 55, 3735–3751. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Lai, R. The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Papo, N.; Mignogna, G.; Andreu, D.; Shai, Y.; Barra, D.; Simmaco, M. Ranacyclins, a new family of short cyclic antimicrobial peptides: Biological function, mode of action, and parameters involved in target specificity. Biochemistry 2003, 42, 14023–14035. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jiang, Y.; Chen, X.; Wang, L.; Ma, C.; Xi, X.; Zhang, Y.; Chen, T.; Shaw, C.; Zhou, M. Ranacyclin-NF, a novel Bowman–Birk type protease inhibitor from the skin secretion of the East Asian frog, Pelophylax nigromaculatus. Biology 2020, 9, 149. [Google Scholar] [CrossRef] [PubMed]
- Wade, D.; Silberring, J.; Soliymani, R.; Heikkinen, S.; Kilpeläinen, I.; Lankinen, H.; Kuusela, P. Antibacterial activities of temporin A analogs. FEBS Lett. 2000, 479, 6–9. [Google Scholar] [CrossRef]
- Li, A.; Zhang, Y.; Wang, C.; Wu, G.; Wang, Z. Purification, molecular cloning, and antimicrobial activity of peptides from the skin secretion of the black-spotted frog, Rana nigromaculata. World J. Microbiol. Biotechnol. 2013, 29, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Maryam, A.; Zhang, H.; Mehmood, T.; Ma, T. Killing cancer with platycodin D through multiple mechanisms. J. Cell. Mol. Med. 2015, 20, 389–402. [Google Scholar] [CrossRef]
- Jiang, D.; Rasul, A.; Batool, R.; Sarfraz, I.; Hussain, G.; Tahir, M.M.; Qin, T.; Selamoglu, Z.; Ali, M.; Li, J.; et al. Potential anticancer properties and mechanisms of action of formononetin. BioMed Res. Int. 2019, 2019, 5854315. [Google Scholar] [CrossRef]
- Dong, Z.; Hu, H.; Yu, X.; Tan, L.; Ma, C.; Xi, X.; Li, L.; Wang, L.; Zhou, M.; Chen, T.; et al. Novel frog skin-derived peptide dermaseptin-PP for lung cancer treatment: In vitro/vivo evaluation and anti-tumor mechanisms study. Front. Chem. 2020, 8, 476. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, H.-B.; Li, S.; Tian, L.-L.; Shang, D.-J. Antitumor effects and cell selectivity of temporin-1CEa, an antimicrobial peptide from the skin secretions of the Chinese brown frog (Rana chensinensis). Biochimie 2012, 94, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Hao, L.; Bao, W.; Zhang, P.; Su, D.; Cheng, Y.; Nie, L.; Wang, G.; Hou, F.; Yang, Y. A novel short anionic antibacterial peptide isolated from the skin of Xenopus laevis with broad antibacterial activity and inhibitory activity against breast cancer cell. Arch. Microbiol. 2016, 198, 473–482. [Google Scholar] [CrossRef]
- Ju, X.; Fan, D.; Kong, L.; Yang, Q.; Zhu, Y.; Zhang, S.; Su, G.; Li, Y. Antimicrobial peptide brevinin-1RL1 from frog skin secretion induces apoptosis and necrosis of tumor cells. Molecules 2021, 26, 2059. [Google Scholar] [CrossRef]
- Van Zoggel, H.; Hamma-Kourbali, Y.; Galanth, C.; Ladram, A.; Nicolas, P.; Courty, J.; Amiche, M.; Delbé, J. Antitumor and angiostatic peptides from frog skin secretions. Amino Acids 2010, 42, 385–395. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, C.; Xiao, G.; Gu, Y. Host defense peptide hymenochirin-1B induces lung cancer cell apoptosis and cell cycle arrest through the mitochondrial pathway. Biochem. Biophys. Res. Commun. 2019, 512, 269–275. [Google Scholar] [CrossRef]
- Attoub, S.; Arafat, H.; Mechkarska, M.; Conlon, J.M. Anti-tumor activities of the host-defense peptide hymenochirin-1B. Regul. Pept. 2013, 187, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Rehani, P.R.; Iftikhar, H.; Nakajima, M.; Tanaka, T.; Jabbar, Z.; Rehani, R.N. Safety and mode of action of diabetes medications in comparison with 5-Aminolevulinic acid (5-ALA). J. Diabetes Res. 2019, 2019, 4267357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, J.M.; Mechkarska, M.; Abdel-Wahab, Y.H.; Flatt, P.R. Peptides from frog skin with potential for development into agents for type 2 diabetes therapy. Peptides 2018, 100, 275–281. [Google Scholar] [CrossRef]
- Soltaninejad, H.; Zare-Zardini, H.; Ordooei, M.; Ghelmani, Y.; Ghadiri-Anari, A.; Mojahedi, S.; Hamidieh, A.A. Antimicrobial peptides from amphibian innate immune system as potent antidiabetic agents: A literature review and bioinformatics analysis. J. Diabetes Res. 2021, 2021, 2894722. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Ren, G.; Zhang, B.; Ma, F.; Fan, J.; Qiu, Z. Screening and identification of a novel antidiabetic peptide from collagen hydrolysates of Chinese giant salamander skin: Network pharmacology, inhibition kinetics and protection of IR-HepG2 cells. Food Funct. 2022, 13, 3329–3342. [Google Scholar] [CrossRef]
- Ojo, O.O.; Abdel-Wahab, Y.H.A.; Flatt, P.R.; Conlon, J.M. Insulinotropic actions of the frog skin host-defense peptide alyteserin-2a: A structure-activity study. Chem. Biol. Drug Des. 2013, 82, 196–204. [Google Scholar] [CrossRef]
- Mechkarska, M.; Ojo, O.O.; Meetani, M.A.; Coquet, L.; Jouenne, T.; Abdel-Wahab, Y.H.; Flatt, P.R.; King, J.D.; Conlon, J.M. Peptidomic analysis of skin secretions from the bullfrog Lithobates catesbeianus (Ranidae) identifies multiple peptides with potent insulin-releasing activity. Peptides 2011, 32, 203–208. [Google Scholar] [CrossRef]
- Conlon, J.M.; Power, G.J.; Abdel-Wahab, Y.H.; Flatt, P.R.; Jiansheng, H.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H. A potent, non-toxic insulin-releasing peptide isolated from an extract of the skin of the Asian frog, Hylarana guntheri (Anura:Ranidae). Regul. Pept. 2008, 151, 153–159. [Google Scholar] [CrossRef]
- Zahid, O.K.; Mechkarska, M.; Ojo, O.O.; Abdel-Wahab, Y.H.; Flatt, P.R.; Meetani, M.A.; Conlon, J.M. Caerulein- and xenopsin-related peptides with insulin-releasing activities from skin secretions of the clawed frogs, Xenopus borealis and Xenopus amieti (Pipidae). Gen. Comp. Endocrinol. 2011, 172, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Marenah, L.; Flatt, P.; Orr, D.F.; Shaw, C.; Abdel-Wahab, Y. Skin secretions of Rana saharica frogs reveal antimicrobial peptides esculentins-1 and -1B and brevinins-1E and -2EC with novel insulin releasing activity. J. Endocrinol. 2006, 188, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ojo, O.O.; Srinivasan, D.K.; Owolabi, B.O.; Vasu, S.; Conlon, J.M.; Flatt, P.R.; Abdel-Wahab, Y.H.A. Esculentin-2CHa-related peptides modulate islet cell function and improve glucose tolerance in mice with diet-induced obesity and insulin resistance. PLoS ONE 2015, 10, e0141549. [Google Scholar] [CrossRef] [PubMed]
- Owolabi, B.O.; Ojo, O.O.; Srinivasan, D.K.; Conlon, J.M.; Flatt, P.R.; Abdel-Wahab, Y.H.A. In vitro and in vivo insulinotropic properties of the multifunctional frog skin peptide hymenochirin-1B: A structure–activity study. Amino Acids 2016, 48, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Abdel-Wahab, Y.H.; Flatt, P.; Leprince, J.; Vaudry, H.; Jouenne, T.; Condamine, E. A glycine-leucine-rich peptide structurally related to the plasticins from skin secretions of the frog Leptodactylus laticeps (Leptodactylidae). Peptides 2009, 30, 888–892. [Google Scholar] [CrossRef]
- Abdel-Wahab, Y.H.; Power, G.J.; Flatt, P.R.; Woodhams, D.C.; Rollins-Smith, L.A.; Conlon, J.M. A peptide of the phylloseptin family from the skin of the frog Hylomantis lemur (Phyllomedusinae) with potent in vitro and in vivo insulin-releasing activity. Peptides 2008, 29, 2136–2143. [Google Scholar] [CrossRef] [PubMed]
- Manzo, G.; Scorciapino, M.A.; Srinivasan, D.; Attoub, S.; Mangoni, M.L.; Rinaldi, A.C.; Casu, M.; Flatt, P.R.; Conlon, J.M. Conformational analysis of the host-defense peptides pseudhymenochirin-1Pb and -2Pa and design of analogues with insulin-releasing activities and reduced toxicities. J. Nat. Prod. 2015, 78, 3041–3048. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, Y.H.; Power, G.J.; Ng, M.T.; Flatt, P.R.; Conlon, J.M. Insulin-releasing properties of the frog skin peptide pseudin-2 and its [Lys18]-substituted analogue. Biol. Chem. 2008, 389, 143–148. [Google Scholar] [CrossRef]
- Abdel-Wahab, Y.H.; Marenah, L.; Orr, D.F.; Shaw, C.; Flatt, P.R. Isolation and structural characterisation of a novel 13-amino acid insulin-releasing peptide from the skin secretion of Agalychnis calcarifer. Biol. Chem. 2005, 386, 581–587. [Google Scholar] [CrossRef]
- Yasser, H.A.A.-W.; Lamin, M.; Peter, R.F.; Conlon, J.M. Insulin releasing properties of the temporin family of antimicrobial peptides. Protein Pept. Lett. 2007, 14, 702–707. [Google Scholar] [CrossRef]
- Pantic, J.M.; Mechkarska, M.; Lukic, M.L.; Conlon, J.M. Effects of tigerinin peptides on cytokine production by mouse peritoneal macrophages and spleen cells and by human peripheral blood mononuclear cells. Biochimie 2014, 101, 83–92. [Google Scholar] [CrossRef]
- Velnar, T.; Bailey, T.; Smrkolj, V. The wound healing process: An overview of the cellular and molecular mechanisms. J. Int. Med. Res. 2009, 37, 1528–1542. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, P.; Nathan, P.; Vasanthan, P.; Musa, S.; Govindasamy, V. Stem cells conditioned medium: A new approach to skin wound healing management. Cell Biol. Int. 2013, 37, 1122–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Jin, W.; Chen, D.; Dong, M.; Xin, X.; Li, C.; Xu, Z. Collagens made from giant salamander (Andrias davidianus) skin and their odorants. Food Chem. 2021, 361, 130061. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.V.S.S.; Sai, K.P.; Babu, M. Application of frog (Rana tigerina Daudin) skin collagen as a novel substrate in cell culture. J. Biomed. Mater. Res. 2002, 61, 197–202. [Google Scholar] [CrossRef]
- Li, H.; Liu, B.; Chen, H.; Gao, Z. A new kind of biomaterials bullfrog skin collagen. Chin. Chem. Lett. 2003, 14, 1146–1149. [Google Scholar]
- Li, H.; Liu, B.; Gao, L.; Chen, H. Studies on bullfrog skin collagen. Food Chem. 2004, 84, 65–69. [Google Scholar] [CrossRef]
- Nan, J.; Zou, M.; Wang, H.; Xu, C.; Zhang, J.; Wei, B.; He, L.; Xu, Y. Effect of ultra-high pressure on molecular structure and properties of bullfrog skin collagen. Int. J. Biol. Macromol. 2018, 111, 200–207. [Google Scholar] [CrossRef]
- Qian, Z.-J.; Jung, W.-K.; Ngo, N.D.; Lee, S.-H.; Kim, S.-K. Isolation and characterization of collagen from skin of bullfrog, Rana catesbeiana Shaw. Fish. Aquat. Sci. 2007, 10, 53–59. [Google Scholar] [CrossRef]
- Wang, T.; Lin, S.; Shen, Y.; Liu, S.; Wang-McCall, T.; Chin, M.; Lin, T.; Yang, C.; Wu, W.; Yang, C. Type I collagen from bullfrog (Rana catesbeiana) fallopian tube. Afr. J. Biotechnol. 2011, 10, 8414–8420. [Google Scholar]
- Jansen, W.; Müller, A.; Grabowski, N.T.; Kehrenberg, C.; Muylkens, B.; Al Dahouk, S. Foodborne diseases do not respect borders: Zoonotic pathogens and antimicrobial resistant bacteria in food products of animal origin illegally imported into the European Union. Veter- J. 2018, 244, 75–82. [Google Scholar] [CrossRef]
- Hampton, J.; Hyndman, T.; Allen, B.; Fischer, B. Animal harms and food production: Informing ethical choices. Animals 2021, 11, 1225. [Google Scholar] [CrossRef]
- Smith, B.P.C.; Tyler, M.J.; Williams, B.D.; Hayasaka, Y. Chemical and olfactory characterization of odorous compounds and their precursors in the parotoid gland secretion of the green tree frog, Litoria caerulea. J. Chem. Ecol. 2003, 29, 2085–2100. [Google Scholar] [CrossRef]
- Frynta, D.; Peléšková, Š.; Rádlová, S.; Janovcová, M.; Landová, E. Human evaluation of amphibian species: A comparison of disgust and beauty. Sci. Nat. 2019, 106, 41. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Gao, J.; Yao, L.; Zeng, L.; Liu, Q.; Zhou, Q.; Zhang, H.; Lu, D.; Fu, J.; Liu, Q.S.; et al. Evidence of foodborne transmission of the coronavirus (COVID-19) through the animal products food supply chain. Environ. Sci. Technol. 2021, 55, 2713–2716. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.W. The chemistry of poisons in amphibian skin. In Chemical Ecology: The Chemistry of Biotic Interaction; National Academies of Sciences, Ed.; The National Academies Press: Washington, DC, USA, 1995; pp. 17–28. [Google Scholar]
- Mohneke, M.; Rödel, M.O. Declining amphibian populations and possible ecological consequences—A review. Salamandra 2009, 45, 203–210. [Google Scholar]
- West, J. Importance of Amphibians: A Synthesis of Their Environmental Functions, Benefits to Humans, and Need for Conservation; Bridgewater State University: Bridgewater, MA, USA, 2018. [Google Scholar]
- Tkaczewska, J. Peptides and protein hydrolysates as food preservatives and bioactive components of edible films and coatings—A review. Trends Food Sci. Technol. 2020, 106, 298–311. [Google Scholar] [CrossRef]
- Etemadian, Y.; Ghaemi, V.; Shaviklo, A.R.; Pourashouri, P.; Mahoonak, A.R.S.; Rafipour, F. Development of animal/ plant-based protein hydrolysate and its application in food, feed and nutraceutical industries: State of the art. J. Clean. Prod. 2020, 278, 123219. [Google Scholar] [CrossRef]
- Chotphruethipong, L.; Aluko, R.E.; Benjakul, S. Hydrolyzed collagen from porcine lipase-defatted seabass skin: Antioxidant, fibroblast cell proliferation, and collagen production activities. J. Food Biochem. 2019, 43, e12825. [Google Scholar] [CrossRef]
- Chen, J.; Gao, K.; Liu, S.; Wang, S.; Elango, J.; Bao, B.; Dong, J.; Liu, N.; Wu, W. Fish collagen surgical compress repairing characteristics on wound healing process in vivo. Mar. Drugs 2019, 17, 33. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.K.; Yeo, K.P.; Chun, Y.Y.; Tan, T.T.Y.; Tan, N.S.; Angeli, V.; Choong, C. Fish scale-derived collagen patch promotes growth of blood and lymphatic vessels in vivo. Acta Biomater. 2017, 63, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Raman, M.; Gopakumar, K. Fish collagen and its applications in food and pharmaceutical industry: A review. EC Nutr. 2018, 13, 752–767. [Google Scholar]
- Feng, X.; Zhang, X.; Li, S.; Zheng, Y.; Shi, X.; Li, F.; Guo, S.; Yang, J. Preparation of aminated fish scale collagen and oxidized sodium alginate hybrid hydrogel for enhanced full-thickness wound healing. Int. J. Biol. Macromol. 2020, 164, 626–637. [Google Scholar] [CrossRef] [PubMed]
- Jana, P.; Mitra, T.; Selvaraj, T.K.R.; Gnanamani, A.; Kundu, P. Preparation of guar gum scaffold film grafted with ethylenediamine and fish scale collagen, cross-linked with ceftazidime for wound healing application. Carbohydr. Polym. 2016, 153, 573–581. [Google Scholar] [CrossRef] [PubMed]



| Scientific Name (Common Name) | Body Part | Moisture (g/100 g) | Protein (g/100 g) | Fat (g/100 g) | Carbohydrate (g/100 g) | Ash (g/100 g) | Reference |
|---|---|---|---|---|---|---|---|
| Frogs | |||||||
| Bufo terrestris (Southern toads) | Whole body | 68.9–74.8 | 16.8–17.5 | 2.8–5.1 | nd. | 3.2–3.8 | [39] |
| Hoplobatrachus occipitalis (Crowned bullfrog) | Meat | 71.7–77.8 | 16.9–19.5 | 1.1–1.8 | 4.6 | 5.0 | [40] |
| Pelophylax esculentus (European frog) | Meat and bones | 77.7 | 7.0 | 3.6 | 6.5 | 2.0 | [38] |
| Pyxicephalus adspersus (Giant African bullfrog) | Thigh and calf | 71.9–78.5 | 6.4–12.0 | 1.7–3.7 | 0.5–0.6 | 4.0–6.0 | [41] |
| Rana catesbiana (American bullfrog) | Fore-chest | 83.4 | 15.3 | 0.6 | nd. | 0.6 | |
| Thigh | 76.9 | 21.2 | 1.4 | nd. | 0.8 | [42] | |
| Calf | 78.4 | 17.7 | 1.0 | nd. | 0.8 | ||
| Rana clamitans/Lithobates clamitans (Green frog/wood frog) | Whole body | 76.2–78.8 | 15.3–16.7 | 1.5–3.2 | nd. | 2.7–3.3 | [39] |
| Rana esculenta (Green frog) | Fore-chest | 79.5 | 18.9 | 1.2 | nd. | 0.6 | [43] |
| Thigh and calf | 79.7 | 19.2 | 0.7 | nd. | 0.6 | ||
| Rana nigromaculata (Black-spotted frog) | Skin | 74.0 | 6.3 | 0.1 | 19.1 | 0.5 | [44] |
| Rana ridibunda (Marsh frog) | Meat | 79.8–82.8 | 10.5–15.7 | 0.7–1.5 | 0.9–1.3 | nd. | [37] |
| Salamanders | |||||||
| Andrias davidianus (Chinese giant alamander) | Meat | 79.0–82.3 | 14.0–16.4 | 2.5–3.5 | nd. | 0.7–1.1 | [7] |
| Skin | 67.6 | 29.1 | 1.21 | nd. | 0.5 | [45] |
| Peptides | Species (Common Name) | Assay(s) | Amino Acid Sequence | Reference |
|---|---|---|---|---|
| Andersonin-C1 | Odorrana margaretae (Green odorous frog) | – | TSRCIFYRRKKCS | [26] |
| Andersonin-G1 | Odorrana andersonii (Golden crossband frog) | – | KEKLKLKAKAPKCYNDKLACT | [26] |
| Andersonin-H3 | Odorrana margaretae (Green odorous frog) | – | VAIYGRDDRSDVCRQVQHNWLVCDTY | [26] |
| Antioxidin-RP1 | Rana pleuraden (Yunnan pond frog) | ABTS (at 80 μg/mL of peptide): 95.9 ± 4.7% *, DPPH (at 80 μg/mL of peptide): 96.9 ± 5.1% *, NO (at 80 μg/mL of peptide): 66.4 ± 9.8% *, FRAP (at 80 μg/mL of peptide, 700 nm): 0.41 ± 0.007. | AMRLTYNKPCLYGT | [68] |
| APBSP | Rana catesbeiana Shaw (American bullfrog) | DPPH (at 1.5 mg/mL of peptide): 16.6–58.6% *, Hydroxyl radical (at 1.5 mg/mL of peptide): 32.8–75.2% *, Superoxide radical (at 1.5 mg/mL of peptide): 10.9–51.5% *, Peroxyl radical (at 1.5 mg/mL of peptide): 36.0–59.7% *. | LEELEEELEGCE | [10] |
| Cathelicid-OA1 | Odorran andersonii (Golden crossband frog) | ABTS (at 4–32 μM of peptide): ~12.5–90% *, DPPH (at 128 μM of peptide): ~90% *. | IGRDPTWSHLAASCLKCIFDDLPKTHN | [47] |
| Nigroain-B-MS1 | Hylarana maosuoensis (Maoson frog) | ABTS (at 50 μM of peptide): 68.3% *, DPPH (at 50 μM of peptide): 99.7% *. | CVVSSGWKWNYKIRCKLTGNC | [46] |
| Nigroain-C-MS1 | Hylarana maosuoensis (Maoson frog) | ABTS (at 50 μM of peptide): 58.3% *, DPPH (at 50 μM of peptide): 99.8% *. | FKTWKNRPILSSCSGIIKG | [46] |
| OA-VII2 | Odorrana andersonii (Golden crossband frog) | Index of oxidative damage in vivo (HaCaT cells exposed with UVB irradiation mouse skin, at 0.5–10 μM of peptide): increase of CAT (23.9–54.5%), reduction of LDH (73.5–100.7%), increase of SOD (79.4 ± 2.0%), increase of GSH (67.4 ± 10.1%). | VIPFLACRPLGL | [26,67] |
| OA-GL21 | Odorrana andersonii (Golden crossband frog) | ABTS (at 500 µM of peptide): ~70% *, DPPH (at 500 µM of peptide): <10% *. | GLLSGHYGRVVSTQSGHYGRG | [26] |
| OM-LV20 | Odorrana margaretae (Green odorous frog) | ABTS: na., DPPH: na., NO (at 1 mM of peptide): 40.1 ± 5.8% *. | LVGKLLKGAVGDVCGLLPIC | [26] |
| OS-LL11 | Odorrana schmackeri (Schmacker’s frog) | ABTS (at 1 μM of peptide): 20–30% *, DPPH (at 1 μM of peptide): <10% *. | LLPPWLCPRNK | [26] |
| Pleurain-A1 | Rana pleuraden (Yunnan pond frog) | ABTS (at 80 μg/mL of peptide): 74.8 ± 2.2% *, DPPH (at 80 μg/mL of peptide): na., NO (at 80 μg/mL of peptide): 21.1 ± 5.2% *, FCA (at 80 μg/mL of peptide, 700 nm): 0.15 ± 0.04. | SIITMTKEAKLPQLWKQIACRLYNTC | [26,68] |
| Pleurain-D4 | Rana pleuraden (Yunnan pond frog) | ABTS (at 80 μg/mL of peptide): na., DPPH (at 80 μg/mL of peptide): 30.3 ± 12.7% *, NO (at 80 μg/mL of peptide): 35.7 ± 3.6% *, FRAP (at 80 μg/mL of peptide, 700 nm): 0.21 ± 0.02. | FLSGILKLAFKIPSVLCAVLKNC | [26,68] |
| Pleurain-E1 | Rana pleuraden (Yunnan pond frog) | ABTS (at 80 μg/mL of peptide): 6.3 ± 8.5% *, DPPH (at 80 μg/mL of peptide): 69.1 ± 9.6% *, NO (at 80 μg/mL of peptide): 11.5 ± 2.9% *, FRAP (at 80 μg/mL of peptide, 700 nm): 0.29 ± 0.02. | AKAWGIPPHVIPQIVPVRIRPLCGNV | [26,68] |
| Ranacyclin-HB1 | Pelophylax hubeiensis (Hubei gold-striped pond frog) | ABTS (at 50 μM of peptide): 49.3 ± 3.8% *, DPPH (at 50 μM of peptide): 11.6 ± 2.4% *. | GAPKGCWTKSYPPQPCFGKK | [46] |
| Salamandrin | Salamandra salamandra (Fire salamander) | ABTS: 0.3 ± 0.0 Trolox equivalent/mg of peptide, DPPH: 0.1 ± 0.0 Trolox equivalent/mg of peptide). | FAVWGCADYRGY | [27] |
| Temporin-MS1 | Hylarana maosuoensis (Maoson frog) | ABTS (at 50 μM of peptide): 21.4 ± 2.2% *, DPPH: na. | FLTGLIGGLMKALGK | [46] |
| Peptides | Species (Common Name) | MIC against Microorganisms | Amino Acid Sequence | Reference |
|---|---|---|---|---|
| Brevinin-1 | Rana brevipoda (Daruma pond frog) | Staphylococcus aureus (8–137.5 μM), Enterococcus faecalis (37.5 μM), Nocardia asteroids (37.5 μM), Escherichia coli (34–37.5 μM). | FLPVLAGIAAKVVPALFCKITKKC | [5,46] |
| Brevinin-1Da | Rana dalmatina (European frog) | S. aureus (7 μM), E. coli (30 μM). | IIPLLLGKVVCAITKKC | [49] |
| Brevinin-1E | Rana esculenta (Green frog) | S. aureus (0.6 μM), E. coli (1.8 μM). | FLPLLAGLAANFLPKIFC1KITRKC1 | [75,76] |
| Brevinin-1T | Rana temporaria (Common frog) | – | VNPIILGVLPKFVCLITKKC | [49] |
| Brevinin-2RNa | Rana nigromaculata (Black-spotted frog) | – | GLFDVVKGVLKGVGKNVAGSLLEQLKCKLSGGC | [77] |
| Buforin-2 | Bufo bufo garagrizans (Asian toad) | Bacillus subtilis (2 μg/mL), S. aureus (4 μg/mL), Streptococcus mutans (2 μg/mL), Streptococcus pneumoniae (4 μg/mL), E. coli (4 μg/mL), Salmonella typhimurium (1 μg/mL), Serratia marcescens (4 μg/mL), Pseudomonas putida (2 μg/mL), Candida albicans (1 μg/mL), Cryptococcus neoformans (1 μg/mL), S. cerevisiae (1 μg/mL). | TRSSRAGLQFPVGRVHRLLRK | [78] |
| Dermaseptin-S1 | Phylomedusa sauvagil (Hylinae tree frog) | S. aureus (12 μM), E. coli (12 μM), Pseudomonas aeruginosa (>24 μM), C. albicans (>24 μM). | ALWKTMLKKLGTMALHAGKAALGAAADTISQGTQ | [5,21,79] |
| Esculentin-1 | Rana esculenta (Green frog) | E. coli (0.2 μM), Bacillus meganterium (0.1 μM), S. aureus (0.4 μM), P. aeruginosa (0.7 μM), C. albican (0.5 μM), S. cerevisiae (0.9 μM). | GIFSKLGRKKIKNLLISGLKNVGKEVGMDVVRTGIDAGOKIKGEC | [5,26] |
| Gaegurin-5 | Rana rugosa (Wrinkled frog) | – | FLGALFKVASKVLPSVKCAImTKKC | [5] |
| Japonicin-2 | Rana chaochiaoensis Liu (Chinese brown frog) | S. aureus (20 μM), E. coli (12 μM). | FGLPMLSILPKALCILLKRKC | [80] |
| Magainin-2 | Xenopus laevis (African clawed frog) | C. albicans, Trichosporon beigelii, Aspergillus flavus, Fusarium oxyspovrum (ns.). | GIGKFLHSAKKFGKAFVGEIMNS | [5,75] |
| Maximin-H5 | Bombina maxima (Chinese red belly toad) | S. aureus (90 µM). | ILGPVLGLVSDTLDOVLGIL-NH2 | [81] |
| Nigrocin-1 | Rana nigromaculata (Black-spotted frog) | Micrococcus luteus (2.5 μg/mL), Shigella dysenteriae (10 μg/mL), Klebsiella pneumoniae (10 μg/mL), P. aeruginosa (75 μg/mL), S. typhimurium (22.5 μg/mL), Proteus mirabilis (>200 μg/mL), S. marcescens (>200 μg/mL), C. albicans (100 μg/mL). | GLLDSIKGMAISAGKGALQNLLKVASC1KLDKTC2 | [77] |
| Nigrocin-2 | Rana nigromaculata (Black-spotted frog) | M. luteus (2.5 μg/mL), S. dysenteriae (10 μg/mL), K. pneumoniae (10 μg/mL), P. aeruginosa (100 μg/mL), S. typhimurium (22.5 μg/mL), P. mirabilis (>200 μg/mL), S. marcescens (>200 μg/mL), C. albicans (150 μg/mL). | GLLSKVLGVGKKVLC1GVSGLC2 | [77] |
| Pseudhymenochirin-1Pb | Pseudhymenochirus merlini (Merlin’s dwarf gray/clawed frog) | E. coli (10 μM), K. pneumoniae (20 μM), P. aeruginosa (20 μM), C. albicans (80 μM). | IKIPSFFRNILKKVGKEAVSLIAGALKQS | [12] |
| Pseudhymenochirin-2Pa | Pseudhymenochirus merlini (Merlin’s dwarf gray/clawed frog) | E. coli (>80 μM), K. pneumoniae (>80 μM), P. aeruginosa (>80 μM), C. albicans (80 μM). | GIFPIFAKLLGKVIKVASSLISKGRTE | [12] |
| Ranacyclin-B-RN1 | Hylarana Nigrovittata (Black-striped frog) | S. aureus (6 μM). | SALVGCWTKSYPPKPCFGR | [82] |
| Ranacyclin-B-RN1 | Hylarana Nigrovittata (Black-striped frog) | S. aureus (12.7 μM). | SALVGCGTKSYPPKPCFGR | [82] |
| Ranacyclin-E | Rana temporaria (Common frog) | Lethal concentration: E. coli (na., >100 μM), Yersinia pseudotuberculosis (9 μM), Pseudomonas syringae pv tabaci (80 μM), B. megaterium (3 μM), Staphylococcus lentus (7 μM), M. luteus (5 μM), C. albicans (na., >100 μM), Candida tropicalis (7.4 μM), Candida guillier-mondii (3.4 μM). MIC: Phytophthora nicotianae spores (32 μM). | SAPRGCWTKSYPPKPCK | [83] |
| Ranacyclin-T | Rana esculenta (Green frog) | Lethal concentration: E. coli (30 μM), Y. pseudotuberculosis (5 μM), Ps. syringae pv tabaci (16 μM), B. megaterium (3 μM), S. lentus (10 μM), M. luteus (8 μM), C. albicans (22 μM), C. tropicalis (14 μM), C. guillier-mondii (1.0 μM). MIC: P. nicotianae spores (16 μM). | GALRGCWTKSYPPKPCK | [83] |
| Ranacyclin-NF | Pelophylax nigromaculatus (Black-spotted frog) | S. aureus (512 μM), E. faecalis (>512 μM), E. coli (>512 μM), P. aeruginosa (>512 μM), P. pneumoniae (>512 μM), C. albicans (>512 μM). | GAPRGCWTKSYPPQPCF | [84] |
| Ranacyclin-NF3L | Pelophylax nigromaculatus (Black-spotted frog) | S. aureus (>512 μM), E. faecalis (>512 μM), E. coli (>512 μM), P. aeruginosa (>512 μM), P. pneumoniae (>512 μM), C. albicans (>512 μM). | GALRGCWTKSYPPQPCF | [84] |
| Ranalexin | Rana catesbeiana (American bullfrog) | S. aureus (4 μg/mL), E. coli (32 μg/mL), P. aeruginosa (128 μg/mL). | FLGLIKIVPAMIC1AVTKKC1 | [11,75] |
| Ranateurin-4 | Rana temporaria (Common frog) | – | FLPFIARLAAKVFPSIICSVTKKC | [5] |
| Ranateurin-T | Rana temporaria (Common frog) | S. aureus (120 μM), E. coli (40 μM), C. albicans (na., >150 μM). | GLLSGLKKVGKHVAKNVAVSLMDSLKCKISGDC | [74] |
| Temporin-A | Rana temporaria (Common frog) | S. aureus (2.6–5.2 μM), E. faecalis (20.9 μM), E. faecium (10.5 μM). | FLPLIGRVLSGIL-Am | [5,85] |
| Temporin-B | Rana temporaria (Common frog) | K. pneumoniae (128 μg/mL), A. baumanii (32–64 μg/mL), P. aeruginosa (>128 μg/mL), E. coli (64 μg/mL), S. aureus (16–32 μg/mL), E. faecalis (64 μg/mL), C. albicans (32 μg/mL). | LLPIVGNLLKSLL-Am | [75] |
| Temporin-L | Rana temporaria (Common frog) | K. pneumoniae (16 μg/mL), A. baumanii (4 μg/mL), P. aeruginosa (16–64 μg/mL), E. coli (4 μg/mL), S. aureus (2–4 μg/mL), E. faecalis (4–8 μg/mL), C. albicans (8 μg/mL). | FVQWFSKFLGRIL | [75] |
| Temporin-1RNa | Rana nigromaculata (Black-spotted frog) | E. coli (25 μM), P. aeruginosa (25 μM), S. aureus (12.5 μM), B. cereus (6.25 μM), Streptococcus lactis (6.25 μM), C. albicans (12.5 μM). | ILPIRSLIKKLL-NH2 | [44,86] |
| Temporin-1RNb | Rana nigromaculata (Black-spotted frog) | E. coli (12.5 μM), P. aeruginosa (12.5 μM), S. aureus (3.13 μM), B. cereus (3.13 μM), S. lactis (3.13 μM), C. albicans (6.25 μM). | FLPLKKLRFGLL-NH2 | [78,86] |
| Tigerinin-1 | Rana tigerina (Asian bullfrog) | B. subtilis (30 μg/mL), S. aureus (30 μg/mL), E.coli (40 μg/mL), P. putida (40 μg/mL), M. luteus (20 μg/mL), S. cerevisiae (80 μg/mL). | FC1TMIPIPRC2Y-Am | [5] |
| Tigerinin-2 | Rana tigerina (Asian bullfrog) | B. subtilis (20 μg/mL), S. aureus (40 μg/mL), E.coli (50 μg/mL), P. putida (50 μg/mL), M. luteus (20 μg/mL), S. cerevisiae (100 μg/mL). | RVCFAIPLPICH-Am | [5] |
| Tigerinin-3 | Rana tigerina (Asian bullfrog) | B. subtilis (30 μg/mL), S. aureus (30 μg/mL), E.coli (40 μg/mL), P. putida (40 μg/mL), M. luteus (30 μg/mL), S. cerevisiae (80 μg/mL). | RVCYAIPLPICY-Am | [71] |
| Peptides | Species (Common Name) | Assay(s) | Amino Acid Sequence | Reference |
|---|---|---|---|---|
| Alyteserin-2a | Alytes obstetricans (Midwife toad) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (TC: 30 nM, Basal release of insulin at 3 μM: 296 ± 26%). In vivo studies: ns. | ILGKLLSTAAGLLSNL | [100] |
| Amolopin | Amolops loloensis (Loloku sucker frog) | Insulin release (in vitro): INS-1 (significantly increased insulin release from 40 to 50 mLU/L at 12.5–50 μg/mL). | FLPIVGKSLSGLSGKL | [98] |
| Bombesin (protein fractions) | Bombina variegate (Yellow-bellied toad) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (insulin secretion: 1.5–3.5 ng/106 cells/20 min). | EQRLGHQWAVGHLM | [98] |
| Bombesin-related peptide | Bombina variegate (Yellow-bellied toad) | – | EDSFGNQWARGHFM | [98] |
| Brevinin-1CBb | Lithobates septentrionalis (Mink frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.0–3.0 ng/106 cells/20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (101.5–144.3% of control). | FLPFIARLAAKVFPSIICSVTKKC | [98,101] |
| Brevinin-1Pa | Rana pipiens (Northern leopard frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells. | FLPIIAGVAAKVFPKIFCAISKKC | [98] |
| Brevinin-1E | Rana esculenta (Green frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells. | FLPAIFRMAAKVVPTIICSITKKC | [98] |
| Brevinin-2GUb | Hylarana guentheri (Günther’s frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (0.7–1.8 ng/106 cells/20 min). | GVIIDTLKGAAKTVAAELLRKAHCKLTNSC | [98,102] |
| Brevinin-2EC | Pelophylax esculentus (European black-spotted frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells. | GILLDKLKNFAKTAGKGVLQSLLNTASCKLSGQC | [98] |
| Brevinin-2-related peptide (B2RP) | Lithobates septentrionalis (Mink frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells. | GIWDTIKSMGKVFAGKILQNL | [98] |
| Lithobates septentrionalis (Mink frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells. | GMASKAGSVLGKVAKVALKAAL | ||
| Caerulein-B1 | Xenopus borealis (Marsabit clawed frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.4 ± 0.1 ng/106 cells/20 min, stimulated rate 1.8 ± 0.0 ng/106 cells/20 min). | EQDY(SO3)GTGWMDF | [98,103] |
| CPF-AM1 | Xenopus amieti (Volcano clawed frog) | GLP-1-releasing activity by GLUTag cells (in vitro): 3.2-fold greater than basal rate at 3 μM concentration). | GLGSVL GKALKIGANLL-NH2 | [14] |
| Dermaseptin B4 | Phyllomedusa trinitatis (Trinidadian monkey frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells. | ALWKDILKNVGKAAGKAVLNTVTDMVNQ | [98] |
| Dermaseptin-LI1 | Agalychnis litodryas (Red-eyed treefrog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells. | AVWKDFLKNIGKAAGKAVLNSVTDMVNE | [98] |
| Esculentin-1 | Rana esculenta (Green frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (3.0 ± 0.3 ng/106 cells/20 min). | GIFSKLGRKKIKNLLISGLKNVGKEVGMDVVRTGIDIAGCKIKGEC | [98,104] |
| Esculentin-1b | Rana esculenta (Green frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (2.9 ± 0.2 ng/106 cells/20 min). | GIFSKLAGKKLKNLLISGLKNVGKEVGMDVVRTGIDIAGCKIKGEC | [98,104] |
| Esculentin-2Cha | Lithobates chiricahuensis (Chiricahua leopard frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (TC: 0.3 nM). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (102.7 ± 1.1% of control). Insulin-releasing activity in high fat fed mice (in vivo): Plasma insulin (AUC: 165.0 ± 16. 9 ng/mL/min). | GFSSIFRGVAKFASKGLGKDLAKLGVDLVACKISKQC | [105] |
| GM-14 | Bombina variegate (Yellow-bellied toad) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells. | GKPFYPPPIYPEDM | [98] |
| GPPGPA | Andrias davidianus (Chinese giant salamander) | α-glucosidase inhibitory activity (in vitro): IC50 (0.3 ± 0.1 mg/mL). | Unknown. | [99] |
| Hymenochirin-1B | Hymenochirus boettgeri (Zaire dwarf clawed frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (TC: 1.0 nM, Insulin release at 1 μM: 13.4 ± 0.1% of total insulin content, Basal release of insulin at 1 nM: 304.4 ± 19.4%). | IKLSPETKDNLKKVLKGAIKGAIAVAKMV | [98,106] |
| IN-21 | Bombina variegate (Yellow-bellied toad) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells. | IYNAICPCKHCNKCKPGLLAN | [98] |
| Magainin-AM1 | Xenopus amieti (Volcano clawed frog) | GLP-1-releasing activity (in vitro): GLUTag cells (1.1–2.0 pg/106 cells/h). | GIKEFAHSLGKFG KAFVGGILNQ | [14] |
| Magainin-AM2 | Xenopus amieti (Volcano clawed frog) | GLP-1-releasing activity (in vitro): GLUTag cells (1.1–2.7 pg/106 cells/h). | GVSKILHSAGKFGKAFLGEIMKS | [14] |
| Ocellatin-L2 | Leptodactylus laticeps (Santa Fe white-lipped frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (181% of basal rate at 3 μM). | GVVDILKGAAKDLAGHLATKVMDKL | [98,107] |
| Palustrin-1c | Lithobates palustris (Pickerel frog) | – | ALSILRGLEKLAKMGIALTNCKATKKC | [98] |
| Palustrin-2CBa | Rana catesbeiana (American bullfrog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.1–2.4 ng/106 cells/20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (100.3–104.4% of control). | GFLDIIKDTGKEFAVKILNNLKCKLAGGCPP | [98,101] |
| PGLa-AM1 | Xenopus amieti (Volcano clawed frog) | GLP-1-releasing activity (in vitro): GLUTag cells (1.1–2.6 pg/106 cells/h). | GMASKAGSVLGKVAKVALKAAL-NH2 | [14] |
| Phylloseptin-L2 | Hylomantis lemur (Lemur tree frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (2.2 ± 0.2 ng/106 cells/20 min). Insulin-releasing activity in mice (in vivo): Plasma insulin (AUC: 59 ng/mL/min). | FLSLIPHVISALSSL | [98,108] |
| Plasticin-L1 | Leptodactylus laticeps (Santa Fe white-lipped frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.0–1.7 ng/106 cells/20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (101.1–113.7% of basal release). | GLVNGLLSSVLGGGQGGGGLLGGIL | [98,107] |
| Pseudhymenochirin-1Pb | Pseudhymenochirus merlini (Merlin’s dwarf gray frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.0–5.0 ng/106 cells/20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (460% of control at 3 μM). | GIFPIFAKLLGKVIKVASSLISKGRTE | [98,109] |
| Pseudhymenochirin-2Pa | Pseudhymenochirus merlini (Merlin’s dwarf gray frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.0–7.0 ng/106 cells/20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (<1500% of control at 3 μM). | IKIPSFFRNILKKVGKEAVSLIAGALKQS | [98,109] |
| Pseudin-2 | Pseudis paradoxa (Shrinking frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.0–2.0 ng/106 cells/20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (100–150% of control). | GLNALKKVFQGIHEAIKLINNHVQ | [98,110] |
| Ranatuerin-1CBa | Rana catesbeiana (American bullfrog) | – | SMLSVLKNLGKVGLGFVACKVNKQC | [98] |
| Ranatuerin-2CBc | Rana catesbeiana (American bullfrog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.0–2.8 ng/106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (98.8–103.7% of control). | GFLDIIKNLGKTFAGHMLDKIKCTIGTCPPSP | [98,101] |
| Ranatuerin-2CBd | Rana catesbeiana (American bullfrog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.2–2.5 ng/106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (99.2–103.8% of control). | GFLDIIKNLGKTFAGHMLDKIRCTIGTCPPSP | [98,101] |
| RK-13 | Agalychnis calcarifer (Costa Rican flying tree frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (0.8–1.5 ng/106 cells × 20 min). | RRKPLFPLIPRPK | [98,111] |
| Temporin-CBa | Rana catesbeiana (American bullfrog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (0.9–2.1 ng/106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (101.2–104.8 % of control). | FLPIASLLGKYL | [98,101] |
| Temporin-CBf | Rana catesbeiana (American bullfrog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.0–2.4 ng/106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (101.2–103.9% of control). | FLPIASMLGKYL | [98,101] |
| Temporin-DRa | Rana draytonii (California red-legged frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (0.6–0.9 ng/106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (3.5–6.4% of total cell content). | HFLGTLVNLAKKIL | [98,112] |
| Temporin-DRb | Rana draytonii (California red-legged frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (0.6–1.5 ng/106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (3.7–5.7% of total cell content). | NFLGTLVNLAKKIL | [98,112] |
| Temporin-Oe | Rana ornativentris (Montane brown frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (0.7–2.3 ng/106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (6.2–9.2% of total cell content). | ILPLLGNLLNGLL | [98,112] |
| Temporin-TGb | Rana tagoi (Tago’s brown frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (0.6–1.8 ng/106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (4.0–5.6% of total cell content). | AVDLAKIANKVLSSLF | [98,112] |
| Temporin-Va | Lithobates virgatipes (Carpenter frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (0.8–1.5 ng/(106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (7.4–13.6% of total cell content). | FLSSIGKLIGNLL | [98,112] |
| Temporin-Vb | Lithobates virgatipes (Carpenter frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (0.8–1.7 ng/106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (7.4–9.8% of total cell content). | FLSIIAKVLGSLF | [98,112] |
| Temporin-Vc | Lithobates virgatipes (Carpenter frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (0.8–2.6 ng/106 cells × 20 min). LDH-releasing activity (in vitro): BRIN-BD11 clonal β-cells (7.4–10.9% of total cell content). | FLPLVTMLLGKLF | [98,112] |
| Tigerinin-1R | Rana tigerina (Asian bullfrog) | GLUTag cell culture (in vitro), glucose homeostasis and beta cell function in mice with diet-induced obesity-diabetes (in vivo). | RVCSAIPLPICH | [14,24] |
| Xenopsin | Xenopus amieti (Volcano clawed frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.5–2.1 ng/106 cells/20 min). | EGKRPWIL | [98,103] |
| Xenopsin-AM2 | Xenopus amieti (Volcano clawed frog) | Insulin-releasing activity (in vitro): BRIN-BD11 clonal β-cells (1.0–2.0 ng/106 cells/20 min). | EGRRPWIL | [98,103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Indriani, S.; Karnjanapratum, S.; Nirmal, N.P.; Nalinanon, S. Amphibian Skin and Skin Secretion: An Exotic Source of Bioactive Peptides and Its Application. Foods 2023, 12, 1282. https://doi.org/10.3390/foods12061282
Indriani S, Karnjanapratum S, Nirmal NP, Nalinanon S. Amphibian Skin and Skin Secretion: An Exotic Source of Bioactive Peptides and Its Application. Foods. 2023; 12(6):1282. https://doi.org/10.3390/foods12061282
Chicago/Turabian StyleIndriani, Sylvia, Supatra Karnjanapratum, Nilesh Prakash Nirmal, and Sitthipong Nalinanon. 2023. "Amphibian Skin and Skin Secretion: An Exotic Source of Bioactive Peptides and Its Application" Foods 12, no. 6: 1282. https://doi.org/10.3390/foods12061282