
Novel Umami Peptides from Hypsizygus marmoreus and Interaction with Umami Receptor T1R1/T1R3


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Preparation of H. marmoreus Hydrolysate
2.3. Separation and Purification of Umami Peptides
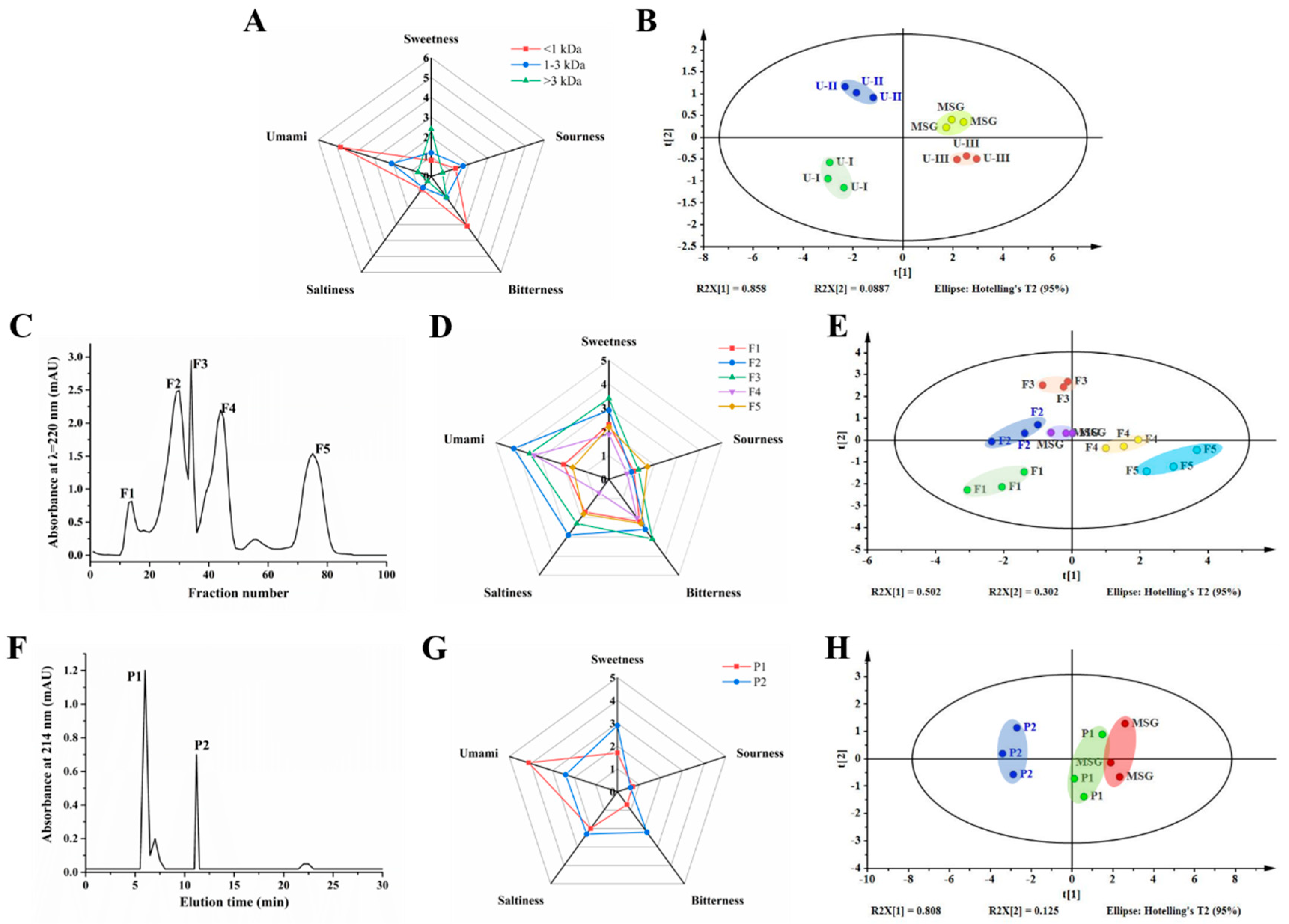
2.3.1. Ultrafiltration of Hydrolysate
2.3.2. Purification of Umami Peptides by Gel Filtration Chromatography (GFC)
2.3.3. Purification of Umami Peptides by RP-HPLC
2.4. Sensory Evaluation
2.5. Electronic Tongue Measurement
2.6. Identification of Umami Peptides Using Nano LC-MS/MS
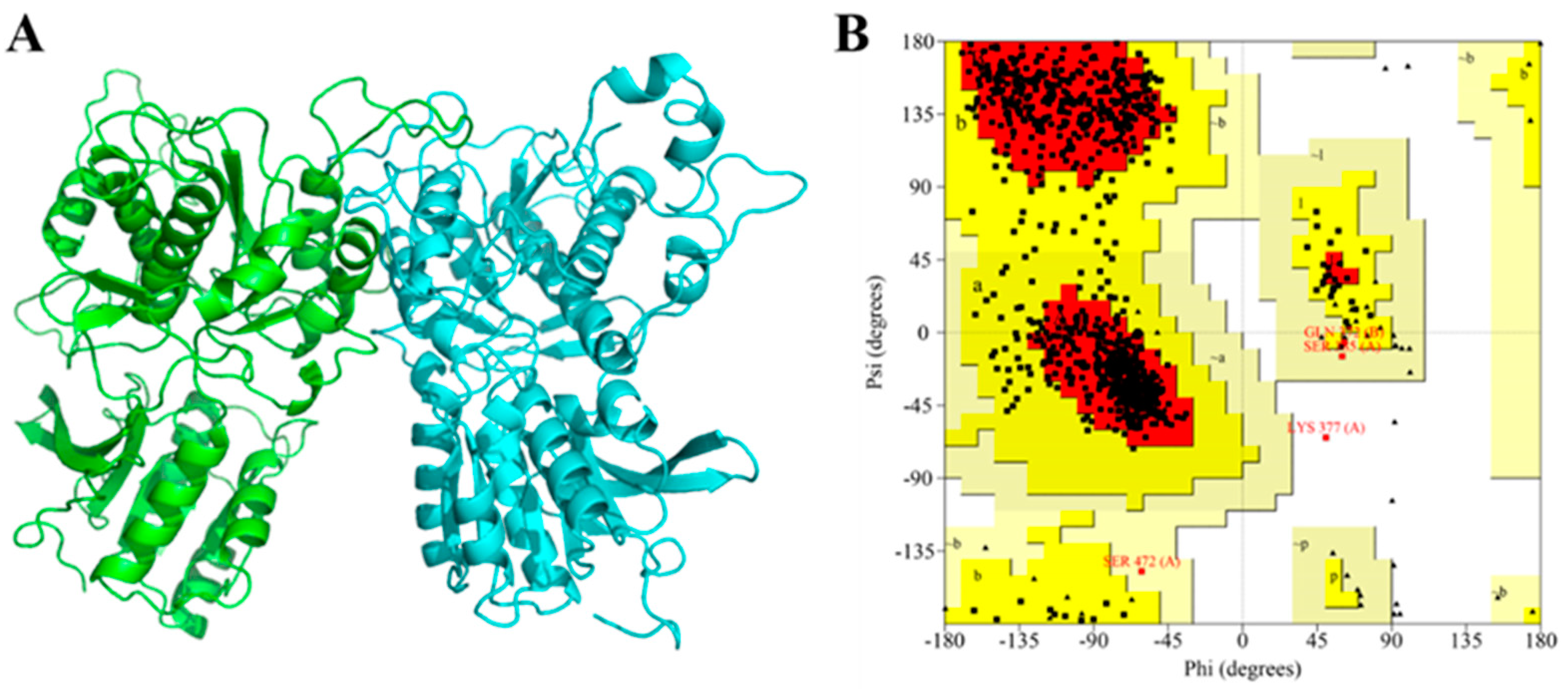
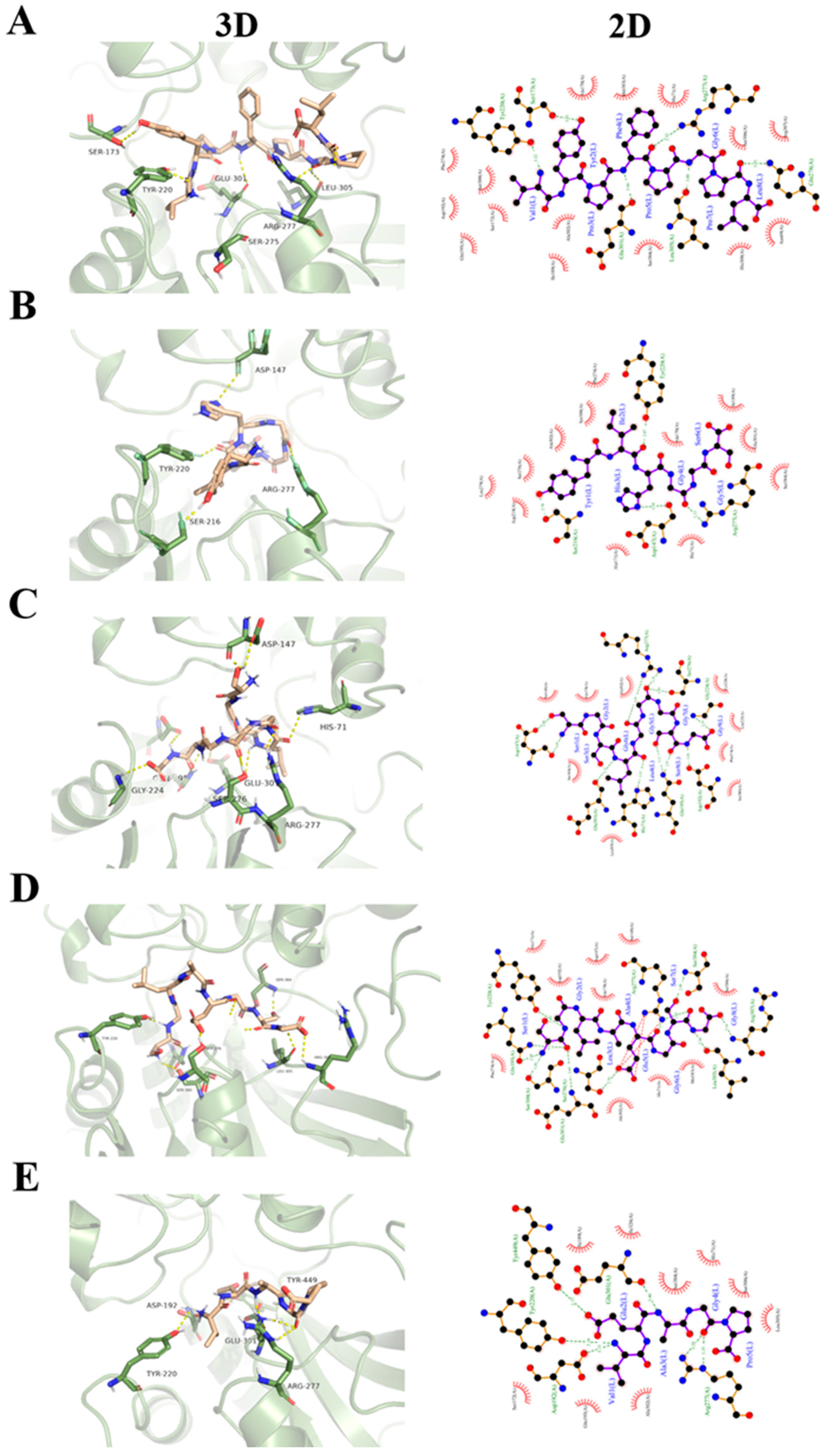
2.7. Homology Modeling and Molecular Docking
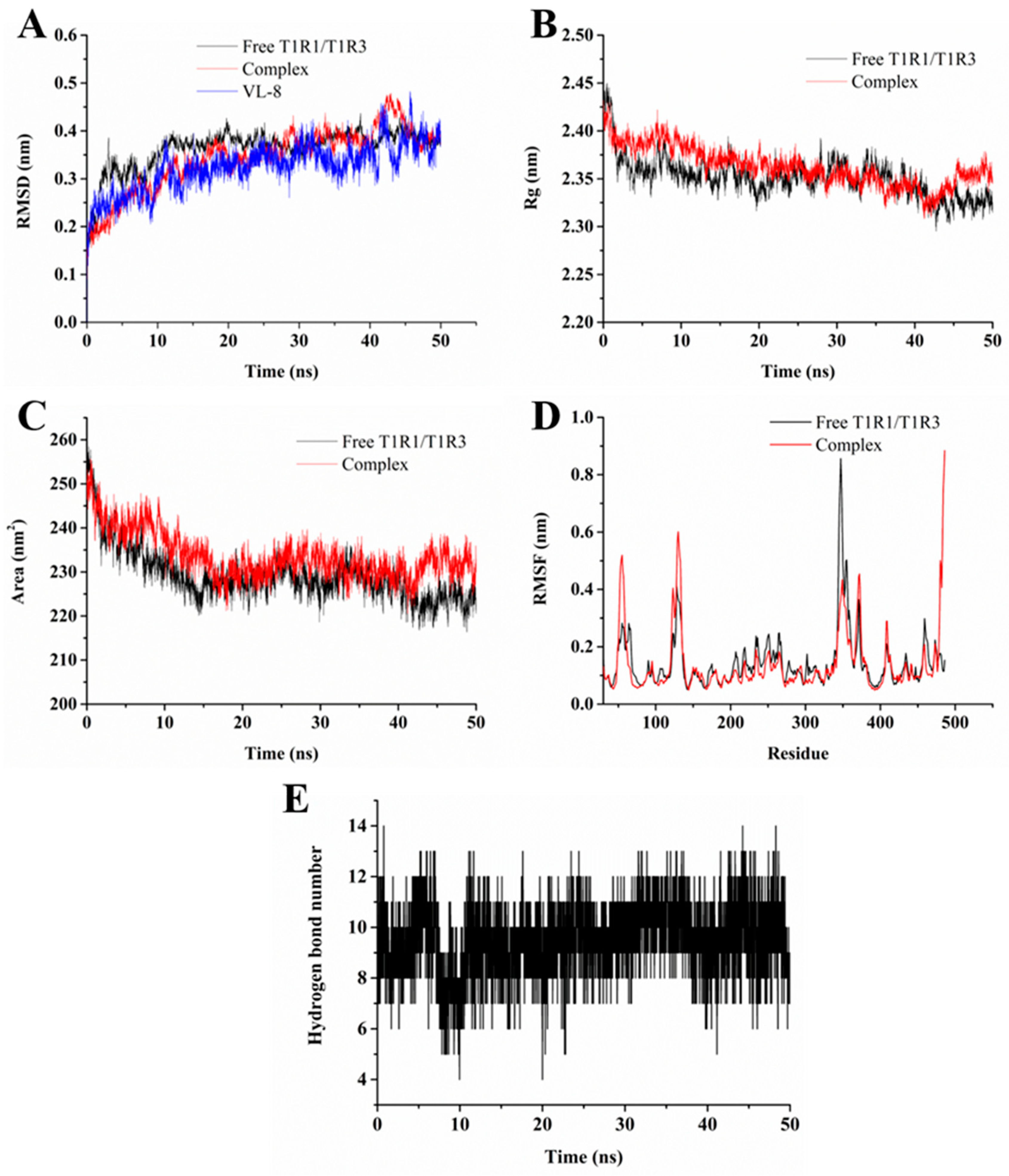
2.8. Molecular Dynamics Simulation Studies
2.9. Statistical Analysis
3. Results and Discussion
3.1. Purification of Umami Peptides
3.2. Identification of Umami Peptides Using LC-MS/MS
3.3. Molecular Docking of Umami Peptides and T1R1/T1R3
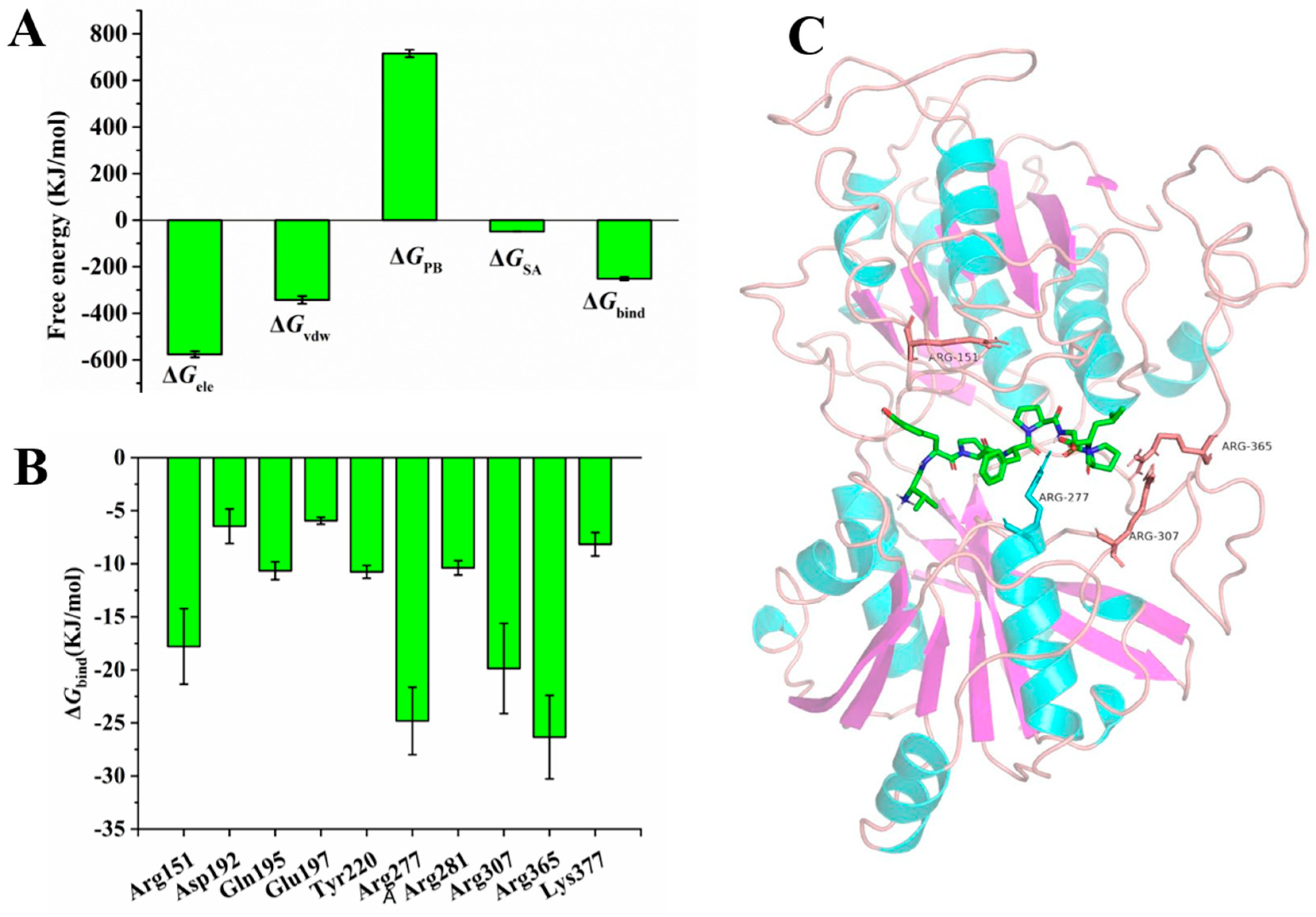
3.4. Molecular Dynamics Simulation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, C.J.; Schieber, A.; Gänzle, M.G. Formation of taste-active amino acids, amino acid derivatives and peptides in food fermentations—A review. Food Res. Int. 2016, 89, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Moerdijk-Poortvliet, T.C.; de Jong, D.L.; Fremouw, R.; de Reu, S.; de Winter, J.M.; Timmermans, K.; Mol, G.; Reuter, N.; Derksen, G.C. Extraction and analysis of free amino acids and 5′-nucleotides, the key contributors to the umami taste of seaweed. Food Chem. 2022, 370, 131352. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Liu, H.; Zhou, X.; Wang, W.; Fan, Y.; Liu, Y. Taste and stability characteristics of two key umami peptides from pufferfish (Takifugu obscurus). Food Chem. 2022, 371, 131124. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gao, H.; Wu, W.; Chen, H.; Fang, X.; Han, Y.; Mu, H. Effects of fermentation with different microbial species on the umami taste of Shiitake mushroom (Lentinus edodes). LWT 2021, 141, 110889. [Google Scholar] [CrossRef]
- Qi, L.; Gao, X.; Pan, D.; Sun, Y.; Cai, Z.; Xiong, Y.; Dang, Y. Research progress in the screening and evaluation of umami peptides. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1462–1490. [Google Scholar] [CrossRef]
- Gao, J.; Fang, D.; Kimatu, B.M.; Chen, X.; Wu, X.; Du, J.; Yang, Q.; Chen, H.; Zheng, H.; An, X.; et al. Analysis of umami taste substances of morel mushroom (Morchella sextelata) hydrolysates derived from different enzymatic systems. Food Chem. 2021, 362, 130192. [Google Scholar] [CrossRef]
- Yang, F.; Lv, S.; Liu, Y.; Bi, S.; Zhang, Y. Determination of umami compounds in edible fungi and evaluation of salty enhancement effect of Antler fungus enzymatic hydrolysate. Food Chem. 2022, 387, 132890. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, Z.; Xin, G.; Sun, B.; Bao, X.; Wei, Y.; Zhao, X.; Xu, H. Advances in umami taste and aroma of edible mushrooms. Trends Food Sci. Technol. 2020, 96, 176–187. [Google Scholar] [CrossRef]
- Shen, D.; Begum, N.; Song, H.; Zhang, Y.; Wang, L.; Zhao, Y.; Zhang, L.; Liu, P. In vitro and in vivo antioxidant activity and umami taste of peptides (<1 kDa) from porcine bone protein extract. Food Biosci. 2021, 40, 100901. [Google Scholar]
- Zhang, Y.; Venkitasamy, C.; Pan, Z.; Liu, W.; Zhao, L. Novel umami ingredients: Umami peptides and their taste. J. Food Sci. 2021, 82, 16–23. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, Y.; Ahmed, Z.; Geng, W.; Tang, W.; Liu, Y.; Jin, H.; Jiang, F.; Wang, J.; Wang, Y. Purification and identification of kokumi-enhancing peptides from chicken protein hydrolysate. Int. J. Food Sci. Technol. 2019, 54, 2151–2158. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, M.; Su, G.; Lin, L. Identification and taste characteristics of novel umami and umami-enhancing peptides separated from peanut protein isolate hydrolysate by consecutive chromatography and UPLC-ESI-QTOF-MS/MS. Food Chem. 2019, 278, 674–682. [Google Scholar] [CrossRef]
- Ruan, S.; Sun, L.; Sun, X.; He, J.; Zhuang, Y. Novel umami peptides from tilapia lower jaw and molecular docking to the taste receptor T1R1/T1R3. Food Chem. 2021, 362, 130249. [Google Scholar]
- Kong, Y.; Zhang, L.; Zhao, J.; Zhang, Y.; Sun, B.; Chen, H. Isolation and identification of the umami peptides from shiitake mushroom by consecutive chromatography and LC-Q-TOF-MS. Food Res. Int. 2019, 121, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.; Chandrashekar, J.; Hoon, M.A.; Feng, L.; Zhao, G.; Ryba, N.J.; Zuker, C.S. An amino-acid taste receptor. Nature 2002, 416, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, F.M.; Vieira, L.B.; Pires, R.G.; Olmo, R.P.; Ferguson, S.S. Metabotropic glutamate receptors and neurodegenerative diseases. Pharmacol. Res. 2017, 115, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun-Waterhouse, D.; Su, G.; Zhao, M. New insight into umami receptor, umami/umami-enhancing peptides and their derivatives: A review. Trends Food Sci. Technol. 2019, 88, 429–438. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Li, D.; Wang, Y.; Li, M.; Chen, C.; Fang, X.; Chen, H.; Zhang, C. Milk protein synthesis is regulated by T1R1/T1R3, a G protein-coupled taste receptor, through the mTOR pathway in the mouse mammary gland. Mol. Nutr. Food Res. 2017, 61, 1601017. [Google Scholar] [CrossRef]
- Huang, A.; Dong, X.; Kang, S.; Chen, D.; Cui, M.; Xu, H. Effects of different protease treatments on the release of umami compounds in the pileus and stipe of Hypsizygus marmoreus. J. Chin. Inst. Food Sci. Technol. 2022, 1–10. Available online: https://kns.cnki.net/kcms/detail/11.4528.TS.20220808.1108.002.html (accessed on 8 August 2022).
- Li, X.; Xie, X.; Wang, J.; Xu, Y.; Yi, S.; Zhu, W.; Mi, H.; Li, T.; Li, J. Identification, taste characteristics and molecular docking study of novel umami peptides derived from the aqueous extract of the clam Meretrix meretrix Linnaeus. Food Chem. 2020, 312, 126053. [Google Scholar] [CrossRef]
- Dang, Y.; Hao, L.; Zhou, T.; Cao, J.; Sun, Y.; Pan, D. Establishment of new assessment method for the synergistic effect between umami peptides and monosodium glutamate using electronic tongue. Food Res. Int. 2019, 121, 20–27. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, X.; Pan, D.; Zhang, Z.; Zhou, T.; Dang, Y. Isolation, characterization and molecular docking of novel umami and umami-enhancing peptides from Ruditapes philippinarum. Food Chem. 2021, 343, 128522. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Da, L.T.; Liu, Y. Understanding the molecular mechanism of umami recognition by T1R1-T1R3 using molecular dynamics simulations. Biochem. Biophys. Res. Commun. 2019, 514, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cui, Z.; Ning, M.; Zhou, T.; Liu, Y. In-silico investigation of umami peptides with receptor T1R1/T1R3 for the discovering potential targets: A combined modeling approach. Biomaterials 2022, 281, 121338. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Shi, Y.; Cao, W.; Shi, J. The inhibition mechanisms of pancreatic lipase by apigenin and its anti-obesity mechanisms revealed by using network pharmacology. Food Biosci. 2022, 45, 101515. [Google Scholar] [CrossRef]
- Ni, M.; Song, X.; Pan, J.; Gong, D.; Zhang, G. Vitexin inhibits protein glycation through structural protection, methylglyoxal trapping, and alteration of glycation site. J. Agric. Food Chem. 2021, 69, 2462–2476. [Google Scholar] [CrossRef]
- Bu, Y.; Liu, Y.; Luan, H.; Zhu, W.; Li, X.; Li, J. Characterization and structure–activity relationship of novel umami peptides isolated from Thai fish sauce. Food Funct. 2021, 12, 5027–5037. [Google Scholar] [CrossRef]
- Wang, K.; Zhuang, H.; Bing, F.; Chen, D.; Feng, T.; Xu, Z. Evaluation of eight kinds of flavor enhancer of umami taste by an electronic tongue. Food Sci. Nutr. 2021, 9, 2095–2104. [Google Scholar] [CrossRef]
- Yu, Z.; Jiang, H.; Guo, R.; Yang, B.; You, G.; Zhao, M.; Liu, X. Taste, umami-enhance effect and amino acid sequence of peptides separated from silkworm pupa hydrolysate. Food Res. Int. 2018, 108, 144–150. [Google Scholar] [CrossRef]
- Rotola-Pukkila, M.; Yang, B.; Hopia, A. The effect of cooking on umami compounds in wild and cultivated mushrooms. Food Chem. 2019, 278, 56–66. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, Y.; Wang, W.; Zhou, X.; Chen, G.; Liu, Y. Seven novel umami peptides from Takifugu rubripes and their taste characteristics. Food Chem. 2020, 330, 127204. [Google Scholar] [CrossRef]
- Yu, Z.; Kang, L.; Zhao, W.; Wu, S.; Ding, L.; Zheng, F.; Liu, J.; Li, J. Identification of novel umami peptides from myosin via homology modeling and molecular docking. Food Chem. 2021, 344, 128728. [Google Scholar] [CrossRef]
- Tao, Z.; Liu, D.; Dai, C.; Liu, Y.; Zhou, G.; Xu, X. Isolation and identification of flavour peptides from Nanjing cooked duck. J. Nanjing Agric. Univ. 2014, 37, 135–142. [Google Scholar]
- Nawaz, K.A.; David, S.M.; Murugesh, E.; Thandeeswaran, M.; Kiran, K.G.; Mahendran, R.; Palaniswamy, M.; Angayarkanni, J. Identification and in silico characterization of a novel peptide inhibitor of angiotensin converting enzyme from pigeon pea (Cajanus cajan). Phytomedicine 2017, 36, 1–7. [Google Scholar] [CrossRef]
- Damak, S.; Rong, M.; Yasumatsu, K.; Kokrashvili, Z.; Varadarajan, V.; Zou, S.; Jiang, P.; Ninomiya, Y.; Margolskee, R.F. Detection of sweet and umami taste in the absence of taste receptor T1r3. Science 2003, 301, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Toda, Y.; Nakagita, T.; Hayakawa, T.; Okada, S.; Narukawa, M.; Imai, H.; Ishimaru, Y.; Misaka, T. Two distinct determinants of ligand specificity in T1R1/T1R3 (the umami taste receptor). J. Biol. Chem. 2013, 288, 36863–36877. [Google Scholar] [CrossRef]
- Vidal-Limon, A.; Aguilar-Toalá, J.E.; Liceaga, A.M. Integration of molecular docking analysis and molecular dynamics simulations for studying food proteins and bioactive peptides. J. Agric. Food Chem. 2022, 70, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhang, H.; Bian, X.; Li, J.; Li, J.; Zhang, H. Insight into the binding of ACE-inhibitory peptides to angiotensin-converting enzyme: A molecular simulation. Mol. Simul. 2019, 45, 215–222. [Google Scholar] [CrossRef]
- Ni, M.; Hu, X.; Gong, D.; Zhang, G. Inhibitory mechanism of vitexin on α-glucosidase and its synergy with acarbose. Food Hydrocoll. 2020, 105, 105824. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Zhu, R.; Yao, W.; Xu, J.; Wang, M.; Ren, J.; Xu, C.; Huang, Z.; Zhang, X.; et al. Computational and experimental approaches to decipher the binding mechanism of general odorant-binding protein 2 from Athetis lepigone to chlorpyrifos and phoxim. J. Agric. Food Chem. 2020, 69, 88–100. [Google Scholar] [CrossRef]
- Lee, Y.; Nakano, A.; Nagasato, Y.; Ichinose, T.; Matsui, T. In Vitro and in Silico Analyses of the Adiponectin Receptor Agonistic Action of Soybean Tripeptides. J. Agric. Food Chem. 2022, 70, 7695–7703. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Peptide Sequence | m/z | Amino Acid Number | MW (Da) |
|---|---|---|---|---|
| VL-8 | VYPFPGPL | 501.79 | 8 | 889.06 |
| YS-6 | YIHGGS | 632.29 | 6 | 632.67 |
| SG-9 | SGSLGGGSG | 677.30 | 9 | 677.67 |
| SG-8 | SGLAEGSG | 676.30 | 8 | 676.68 |
| VP-5 | VEAGP | 471.23 | 5 | 471.51 |
| Umami Peptide | Score (kcal/mol) |
|---|---|
| VL-8 | −9.143 |
| YS-6 | −8.296 |
| SG-9 | −6.959 |
| SG-8 | −7.723 |
| VP-5 | −6.581 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, X.; Wan, C.; Huang, A.; Xu, H.; Lei, H. Novel Umami Peptides from Hypsizygus marmoreus and Interaction with Umami Receptor T1R1/T1R3. Foods 2023, 12, 703. https://doi.org/10.3390/foods12040703
Dong X, Wan C, Huang A, Xu H, Lei H. Novel Umami Peptides from Hypsizygus marmoreus and Interaction with Umami Receptor T1R1/T1R3. Foods. 2023; 12(4):703. https://doi.org/10.3390/foods12040703
Chicago/Turabian StyleDong, Xiaobo, Chao Wan, Aiyun Huang, Huaide Xu, and Hongjie Lei. 2023. "Novel Umami Peptides from Hypsizygus marmoreus and Interaction with Umami Receptor T1R1/T1R3" Foods 12, no. 4: 703. https://doi.org/10.3390/foods12040703