
In Vitro Assessment Methods for Antidiabetic Peptides from Legumes: A Review

Abstract
:1. Introduction
2. Starch to Blood Glucose
3. Antidiabetic Activity of Leguminous Protein Hydrolysates and Peptides
4. In Vitro Studies on Leguminous Antidiabetic Peptides

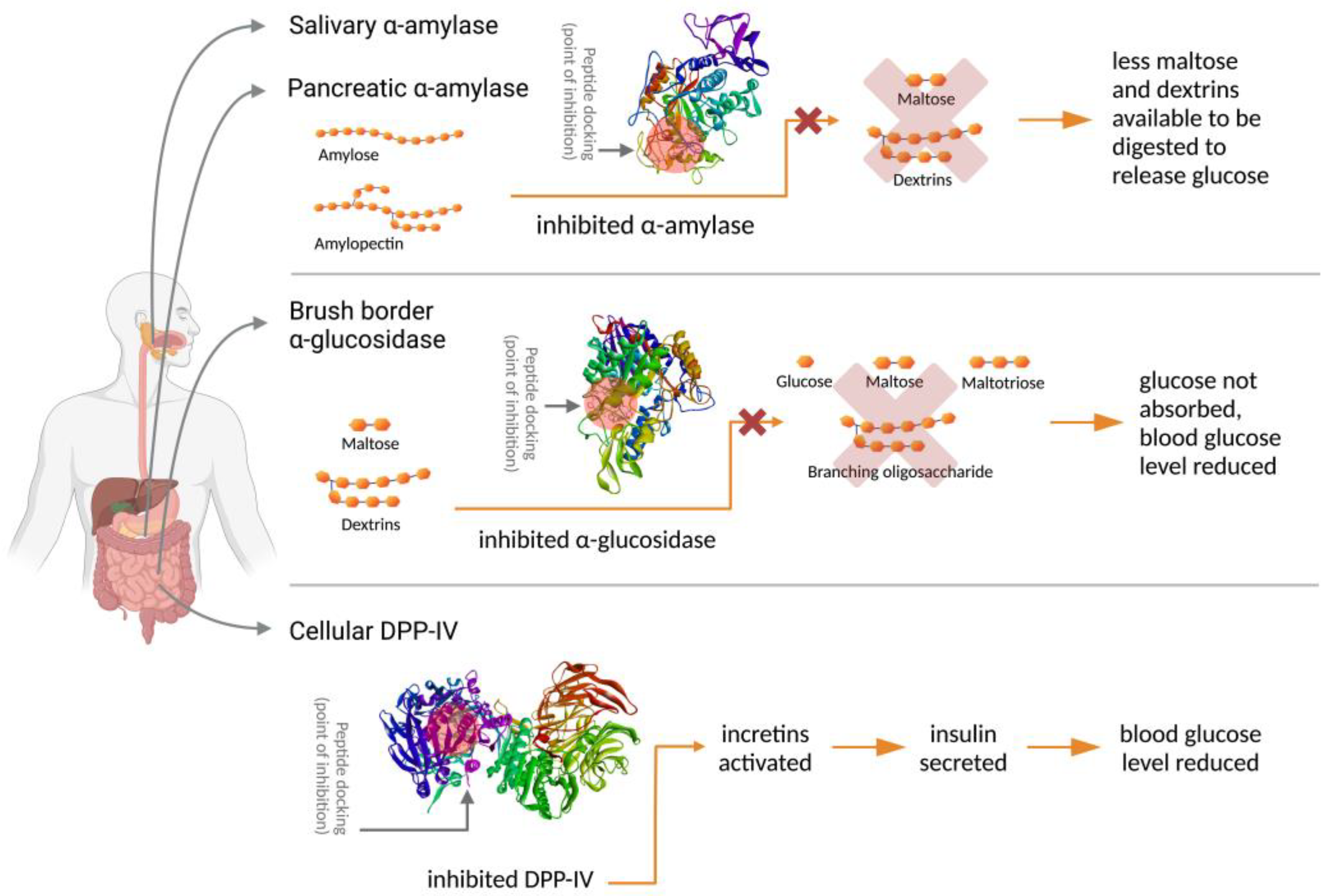{kind=link}
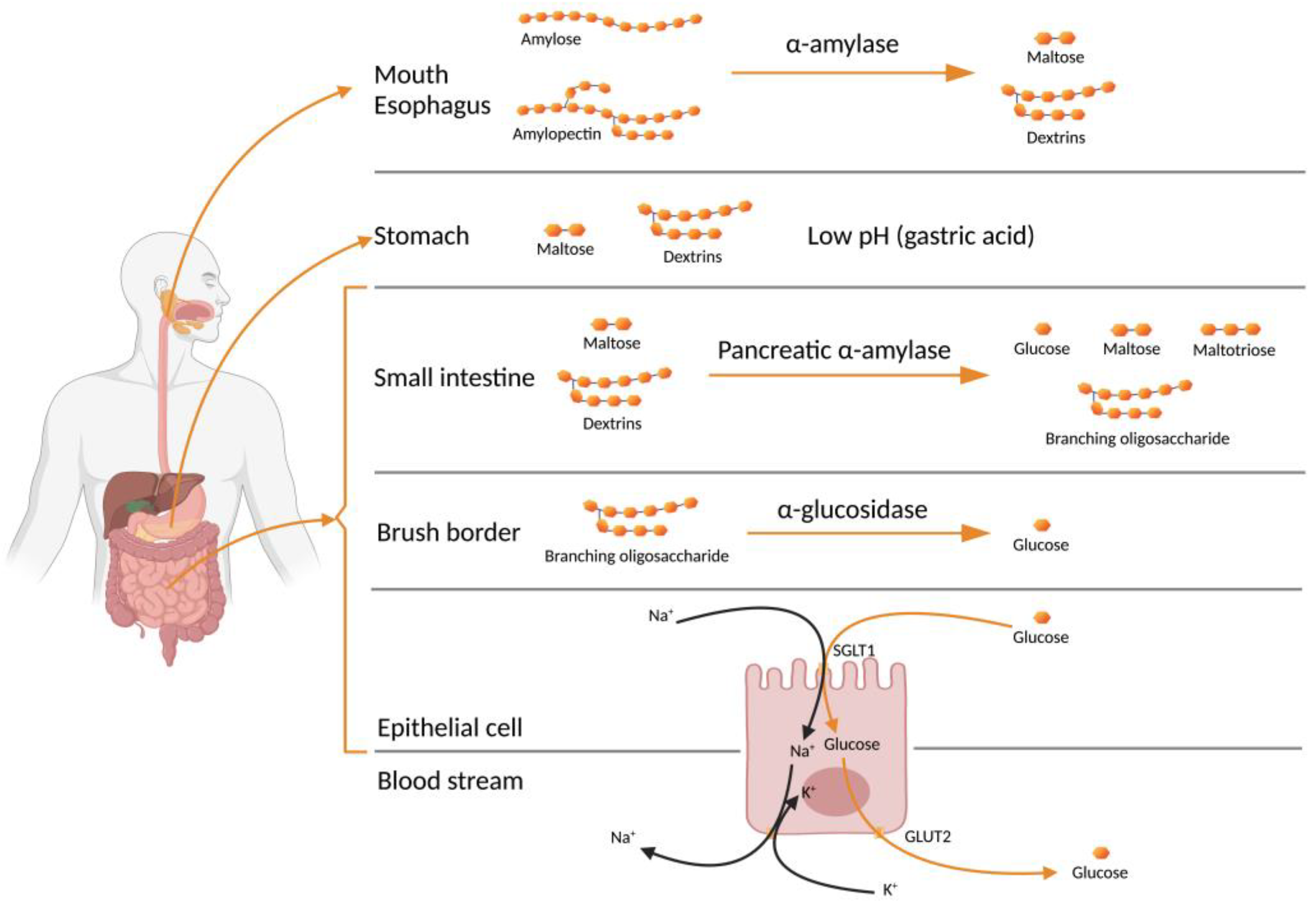{kind=link}
| Legumes | Sample Type | Highest α-Amylase Inhibition * | Highest α-Glucosidase Inhibition * | Highest DPP-IV Inhibition * | References | |||
|---|---|---|---|---|---|---|---|---|
| Value | Sample | Value | Sample | Value | Sample | |||
| Common Bean (Phaseolus vulgaris L.): Black Pinto Red Navy Great Northern | Raw and precooked | 36% inh AC/mg protein [Prot] = not reported [Enz] = 13 U/mL [Ac] = 1 mM | Red beans Raw, whole H: Pepsin-Pancreatin | >40% (~48–67%) inh AC /mg protein (not statistically different) [Prot] = not reported [Enz] = 1 U/mL [Ac] = 1 mM | All beans Raw and precooked, whole H: Pepsin-Pancreatin | 0.093 mg protein/mL [Prot] = 1 mg DW/mL [Enz] = 100 ng/mL [control] = not reported | Navy beans Precooked, whole H: Pepsin-Pancreatin | [59] |
| 0.095 mg protein/mL [Prot] = 1 mg DW/mL [Enz] = 100 ng/mL [control] = not reported | Navy beans Raw, whole H: Pepsin-Pancreatin | |||||||
| Common Bean (Phaseolus vulgaris L.): Mexico, Pinto: Pinto-Bayacora Pinto-Bravo Pinto-Centenario Pinto-Saltillo Mexico, Flores de Mayo and Junio: FMayo-Eugenia FMayo-67 FMayo-199 FMayo-202 FJunio-Leon FJunio-Marcela Mexico, Negros: Negro-Frijozac Negro-Otomi Brazil, Carioca: BRSHorizote BRS-Pontal Perola | Raw | 14.9 ± 1.7% inh AC/mg BPI [Prot] = not reported [Enz] = 13 U/mL [Ac] = 1 mM | Pinto-Bayacora Raw, dehulled H: Pepsin-Pancreatin | [38] | ||||
| 14.9 ± 0.4% inh rel AC/mg BPI [Prot] = not reported [Enz] = 13 U/mL [Ac] = 1 mM | FMayo-67 Raw, dehulled H: Pepsin-Pancreatin | |||||||
| Common Bean (Phaseolus vulgaris L.): Black Otomi BRS-Horizonte BRS-Pontal Perola | Raw | 50.10% inh/mg DW [Prot] = 1 mg DW/mL [Enz] = 1 U/mL [Ac] = 1 mmol/L | BRS-Horizonte Raw, dehulled H: Pepsin-Pancreatin | 0.14 mg DW/mL [Prot] = 1 mg DW/mL [Enz] = 10 ng/mL [control] = not reported | Black Otomi Raw, dehulled H: Pepsin-Pancreatin | [60] | ||
| 49.34% inh/mg DW [Prot] = 1 mg DW/mL [Enz] = 1 U/mL [Ac] = 1 mmol/L | Synthesized peptide: KKSSG | 0.03 mg DW/mL [Prot] = 1 mg DW/mL [Enz] = 10 ng/mL [control] = not reported | Synthesized peptide: KTYGL | |||||
| Black Otomi (Phaseolus vulgaris L.) | Raw | 64.5 ± 2.7% inh/ mg dry matter [Prot] = 1 mg DM/mL [Enz] = 13 U/mL [Ac] = 1 mM | Raw, dehulled H: Flavourzyme, 2 h, 1:20 (E/S) | 75.3 ± 0.7% to 78.4 ± 0.6% inh/mg dry matter (not statistically different) [Prot] = 1 mg DM/mL [Enz] = 1 U/mL [Ac] = 1 mmol/L | Raw, dehulled H: Papain, 2, 3, 4 h, 1:20, 1:30, 1:50 (E/S) | 96.7% inh/ mg dry matter [Prot] = 1 mg DW/mL [Enz] = 10 ng/mL [control] = not reported | Raw, dehulled H: Alcalase, 2 h, 1:20 (E/S) | [61] |
| HTC Common Bean (Phaseolus vulgaris L.): cv Negro 8025 cv. Pinto Durango | Raw | 49.9 ± 1.4% [Prot] = 100 μg/mL [Enz] = 10.8 U/mL [Ac] = 1 mM | Pinto Durango Raw, dehulled H: Bromelain, 2 h F: <1 kDa | 76.4 ± 0.5% [Prot] = not reported [Enz] = 1.0 U/mL [Ac] = 1 mM | Pinto Durango Raw, dehulled H: Alcalase, 2 h F: <1 kDa | 55.3 ± 1.6% [Prot] = 100 μg/mL [Enz] = 100 ng/mL [control] = not reported | Pinto Durango Raw, dehulled H: Alcalase, 2 h F: <1 kDa | [25] |
| Common Bean (Phaseolus vulgaris) | Germinated | 30.88 ± 2.45% AC/mg SP [Prot] = 1 mg/mL [Enz] = 13 U/mL [Ac] = 1 mM | Dehulled, germinated 24 h H: Non-hydrolyzed | 1.2 mg soluble protein/mL [Prot] = 0.1–4.0 mg/mL [Enz] = 100 ng/mL [control] = not reported | Dehulled, non-germinated H: Non-hydrolyzed | [55] | ||
| Pinto Bean (Phaseolus vulgaris cv. Pinto) | Raw | 57.48 ± 2.51% [Prot] = not reported [Enz] = 0.5 mg/mL [Ac] = not used | Raw, whole H: Protamex, pH 6.5, 1 h, 1:10 (E/S) | [42] | ||||
| 62.1 ± 3.49% [Prot] = not reported [Enz] = 0.5 mg/mL [Ac] = not used | Raw, whole H: Protamex, pH 6.5, 1 h, 1:10 (E/S) F: <3 kDa | |||||||
| Pinto Bean (Phaseolus vulgaris cv. Pinto) | Raw | 57.8% inh/100 μg 10.03 ± 0.47 mM [Prot] = 1 mg/mL [Enz] = 0.5 mg/mL [Ac] = not used | Raw, whole H: Protamex, pH 6.5, 1 h, 1:10 (E/S) F: <3 kDa Synthesized pinto bean peptide fraction 5 (PBp5) | [62] | ||||
| Bean (Phaseolus vulgaris L.) | Fermented with L. plantarum 299v | 0.038 µg/mL [Prot] = not reported [Enz] = not reported [Ac] = not used | Dehulled Fermented 22 °C, 3 h H: Amylase-Pepsin-Pancreatin F: Fraction III collected in Sephadex G-10 from 3.5–7 kDa | [63] | ||||
| BRS-Pontal (Phaseolus vulgaris L.) | Raw | 89.1 ± 0.3% [Prot] = 10 mg/mL [Enz] = 10 U/mL [Ac] = 10 mg/mL | Hard-to-cook bean, raw, dehulled H: Non-hydrolyzed F: <3 kDa | 89.2 ± 0.1% [Prot] = 10 mg protein/mL [Enz] = 2 U/mL [Ac] = 10 mg/mL | Easy-to-cook bean, raw, dehulled Non-hydrolyzed F: <3 kDa | [7] | ||
| Carioca Bean (Phaseolus vulgaris L. cv Carioca) | Raw | 101.61 ± 0.78% [Prot] = 1 mg/mL [Enz] = not reported [Ac] = not used | Raw, whole H: Alcalase-Neutrase (1/2:1/2) | 34.73 ± 4.65% [Prot] = 1 mg/mL [Enz] = 0.1 U/mL [Ac] = not used | Raw, whole H: Flavourzyme: Alcalase (1/2:1/2) | [64] | ||
| Cowpea cultivar BRS Novaera (Vigna unguiculata L.) | Germinated | 0.58 mg soluble protein/mL [Prot] = 0.1–4.0 mg/mL [Enz] = 100 ng/mL [control] = not reported | Dehulled, Non-germinated H: Alcalase, 1 h-Pepsin-Pancreatin | [57] | ||||
| Black Cowpea (Vigna unguiculata) | Raw | 96.81% [Prot] = 100 mg/mL [Enz] = 13 U/mL [Ac] = not reported | Raw, whole H: Pepsin-Pancreatin F: <1 kDa | 97.34% [Prot] = 10 mg/mL [Enz] = 2 U/mL [Ac] = not reported | Raw, whole H: Alcalase-Flavourzyme F: >10 kDa | 85% 2.06 mg protein/mL [Prot] = not reported [Enz] = not reported [Stg] = not reported | Raw, whole H: Alcalase-Flavourzyme F: Protein Hydrolysate | [54] |
| Bambara bean (Vigna subterranea) | Raw | 44.253 ± 1.327% [Prot] = 1 mg/mL [Enz] = 0.26 mU/test well [DipA] = not reported | Bambara bean protein isolate H: Alcalase | [13] | ||||
| 29.276 ± 0.878% at 1 mg/mL [Prot] = 1 mg/mL [Enz] = 0.26 mU/test well [DipA] = not reported | Bambara bean protein isolate H: Trypsin-Pepsin-α-chymotrypsin-trypsin-pancreatin | |||||||
| 1.733 mg/mL [Prot] = 1 mg/mL [Enz] = 0.26 mU/test well [DipA] = not reported | Bambara bean protein isolate H: Alcalase H: Thermolysin | |||||||
| Soybean (Glycine max) | Germinated | 1.7 mg/mL [Prot] = 0.2–4 mg/mL [Enz] = 2 U/mL [Ac] = 0.1–1.3 mg/mL | Germinated 6 days, whole H: Pepsin-Pancreatin | As maltase 2.56 mg/mL [Prot] = 1–10 mg/mL [Enz] = 1 U/mL [Ac] = not used | Germinated 6 days, whole H: Pepsin-Pancreatin F: <5 kDa | 0.91 mg/mL [Prot] = 0.08–5 mg/mL [Enz] = 0.26 mU/test well [DipA] = 0.78–50 μM | Germinated 6 days, whole H: Pepsin-Pancreatin F: 5–10 kDa | [16] |
| As sucrase 1.23 mg/mL [Prot] = 1–10 mg/mL [Enz] = 1 U/mL [Ac] = not used | Germinated 6 days, H: Pepsin-Pancreatin F: <5 kDa | |||||||
| ~85% [Prot] = 1 mg/mL [Enz] = 2 U/mL [Ac] = 0.1–1.3 mg/mL | Germinated 6 days, whole H: Pepsin-Pancreatin F1 fraction collected by semi-preparative RP-HPLC from 5–10 kDa | As maltase 32% [Prot] = 1 mg/mL [Enz] = 1 U/mL [Ac] = not used | Germinated 6 days, whole H: Pepsin-Pancreatin F: F4 fraction collected in semi-preparative RP-HPLC from 5–10 kDa | 0.7 mg/mL [Prot] = 0.08–5 mg/mL [Enz] = 0.26 mU/test well [DipA] = 0.78–50 μM | Germinated 6 days, whole H: Pepsin-Pancreatin F: F3 fraction collected by semi-preparative RP-HPLC from 5–10 kDa | |||
| As sucrase 22% [Prot] = 1 mg/mL [Enz] = 1 U/mL [Ac] = not used | Germinated 6 days, whole H: Pepsin-Pancreatin F: F1 fraction collected by semi-preparative RP-HPLC from 5–10 kDa | |||||||
| Soybean | Raw (Isolated soybean protein) | 0.27 mg/mL [Prot] = not reported [Enz] = 0.15 U/mL [Ac] = not used | Protein isolate H: Trypsin F: <5 kDa | [65] | ||||
| 0.049 mg/mL [Prot] = not reported [Enz] = 0.15 U/mL [Ac] = not used | Protein isolate H: Trypsin F: <5 kDa Fraction C-III-2a from RP-HPLC | |||||||
| Soy | Raw (Soy protein powder) | 77.64 ± 1.07% [Prot] = not reported [Enz] = 0.2 U/mL [Ac] = 10 mg/mL | Soy protein isolate H: Alkaline protease-pepsin-pancreatin | 47.94 ± 1.10% [Prot] = not reported [Enz] = 0.02 U/mL [control] = buffer | Soy protein isolate H: Alkaline protease-pepsin-pancreatin | [12] | ||
| 87.10 ± 2.70% [Prot] = not reported [Enz] = 0.2 U/mL [Ac] = 10 mg/mL | Soy protein isolate H: Alkaline protease-pepsin-pancreatin F: H1 fraction from DEAE-52 | |||||||
| 95.35 ± 2.70% [Prot] = not reported [Enz] = 0.2 U/mL [Ac] = 10 mg/mL | Soy protein isolate H: Alkaline protease-pepsin-pancreatin F: H1 fraction from DEAE-52, then H1-2 fraction from Sephadex G-15 | |||||||
| 162.29 ± 0.74 μmol/L [Prot] = not reported [Enz] = 0.2 U/mL [Ac] = 10 mg/mL | Synthesized peptide: WLRL | |||||||
| Yellow field pea (Pisum sativum L.) | Raw (Yellow field pea protein concentrate) | 30.52 ± 0.01% [Prot] = 225 μg/mL [Enz] = 28.57 μg/mL [Ac] = 1.5–3 μg/mL | Yellow field pea protein concentrate H: Chymotrypsin F: 1–3 kDa | 53.35 ± 2.78% [Prot] = 20 mg/mL [Enz] = 8.33 mg/mL [Ac] = 0.00625–0.125 mg/mL | Yellow field pea protein concentrate H: Chymotrypsin F: <1 kDa | [9] | ||
5. Protein Extraction
6. Protein Hydrolysis
7. α-Amylase Inhibitory Assays
8. α-Glucosidase Inhibitory Assays
9. Dipeptidyl Peptidase-IV (DPP-IV) Inhibitory Assays
10. Molecular Mass Profiling
11. Peptide Sequence Identification and Bioactivity Prediction
12. Molecular Docking and Enzyme Kinetics Study
13. Conclusions
14. Future Recommendations
- As legumes are an economical dietary protein source, research to explore antidiabetic peptides from other legume sources is needed;
- Some proteins may cause allergic reactions, and studies on allergenicity and safety of antidiabetic peptides are suggested;
- The mechanism of how antidiabetic peptides are released into the human digestive system, absorbed into the blood stream, and migrated to the target sites is not fully understood. Moreover, at the molecular level, the actual sequence of events and the molecular determinants dictating the inhibition mechanism are far from being understood, requiring an interdisciplinary approach, such as from nutrition and biomolecular science;
- When consumed as peptides, the harsh environment in the human digestion tract may cause changes in the peptides that will affect their bioactivity and bioavailability. Hence, peptide stability, efficacy, and bioavailability studies would assist in determining the dose of peptide intake;
- Low-molecular-weight peptides composed of hydrophobic amino acids are typically bitter, and the application of these peptides in foods without affecting the sensory profile will require a special process such as micro- or nanoparticle encapsulation. Research in this area may offer solutions not only for elucidating sensory aspects, but also bioavailability, bioactivity, and safety;
- Feeding studies using human volunteers will be required prior to the application of the inhibitory peptides as ingredients in the diets, in functional foods, or as nutraceuticals or pharmaceuticals supplements.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Song, X.; Ren, Y.; Shan, P.F. Global, regional, and national burden and trend of diabetes in 195 countries and territories: An analysis from 1990 to 2025. Sci. Rep. Nat. Res. 2020, 10, 14790. [Google Scholar] [CrossRef]
- World Health Organization. Diabetes: Key Facts. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 26 September 2022).
- International Diabetes Federation. IDF Diabetes Atlas. International Diabetes Federation. 2020. Available online: https://idf.org/aboutdiabetes/what-is-diabetes/facts-figures.html (accessed on 3 December 2020).
- NCD Risk Factor Collaboration (NCD-RisC). Worldwide trends in diabetes since 1980: A pooled analysis of 751 population-based studies with 4.4 million participants. Lancet 2016, 387, 1513–1530. [Google Scholar] [CrossRef] [PubMed]
- Korat, A.V.A.; Willett, W.C.; Hu, F.B. Diet, Lifestyle, and Genetic Risk Factors for Type 2 Diabetes: A Review from the Nurses’ Health Study, Nurses’ Health Study 2, and Health Professionals’ Follow-Up Study. Curr. Nutr. Rep. 2014, 3, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.M.; Chen, Y.H.; Yen, P.L.; Chang, S.T. Antihyperglycemic and antioxidant activities of twig extract from Cinnamomum osmophloeum. J. Tradit. Complement. Med. 2016, 6, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Mejía, E.; Batista, K.A.; Fernández, J.J.A.; Fernandes, K.F. Antihyperglycemic and hypoglycemic activity of naturally occurring peptides and protein hydrolysates from easy-to-cook and hard-to-cook beans (Phaseolus vulgaris L.). Food Res. Int. 2019, 121, 238–246. [Google Scholar] [CrossRef]
- Shobana, S.; Sreerama, Y.N.; Malleshi, N.G. Composition and enzyme inhibitory properties of finger millet (Eleusine coracana L.) seed coat phenolics: Mode of inhibition of α-glucosidase and pancreatic amylase. Food Chem. 2009, 115, 1268–1273. [Google Scholar] [CrossRef]
- Awosika, T.O.; Aluko, R.E. Inhibition of the in vitro activities of α-amylase, α-glucosidase and pancreatic lipase by yellow field pea (Pisum sativum L.) protein hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 2021–2034. [Google Scholar] [CrossRef]
- Johnson, M.H.; Lucius, A.; Meyer, T.; Gonzalez De Mejia, E. Cultivar evaluation and effect of fermentation on antioxidant capacity and in vitro inhibition of α-amylase and α-glucosidase by highbush blueberry (Vaccinium corombosum). J. Agric. Food Chem. 2011, 59, 8923–8930. [Google Scholar] [CrossRef]
- Widowati, S.; Astawan, M.; Muchtadi, D.; Wresdiyati, T. Hypoglycemic Activity of Some Indonesian Rice Varieties and Their Physicochemical Properties. Indones. J. Agric. Sci. 2016, 7, 57. [Google Scholar] [CrossRef]
- Wang, R.; Zhao, H.; Pan, X.; Orfila, C.; Lu, W.; Ma, Y. Preparation of bioactive peptides with antidiabetic, antihypertensive, and antioxidant activities and identification of α-glucosidase inhibitory peptides from soy protein. Food Sci. Nutr. 2019, 7, 1848–1856. [Google Scholar] [CrossRef]
- Mune Mune, M.A.; Minka, S.R.; Henle, T. Investigation on antioxidant, angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory activity of Bambara bean protein hydrolysates. Food Chem. 2018, 250, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Hatanaka, T.; Inoue, Y.; Arima, J.; Kumagai, Y.; Usuki, H.; Kawakami, K.; Kimura, M.; Mukaihara, T. Production of dipeptidyl peptidase IV inhibitory peptides from defatted rice bran. Food Chem. 2012, 134, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J. Dipeptidyl Peptidase-4 Inhibition and the Treatment of Type 2 Diabetes. Diabetes Care 2007, 30, 1335–1343. [Google Scholar] [CrossRef]
- González-Montoya, M.; Hernández-Ledesma, B.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Bioactive peptides from germinated soybean with anti-diabetic potential by inhibition of dipeptidyl peptidase-IV, α-amylase, and α-glucosidase enzymes. Int. J. Mol. Sci. 2018, 19, 2883. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, Y.; Dai, Y.; Peng, J. Natural products for the treatment of type 2 diabetes mellitus: Pharmacology and mechanisms. Pharmacol. Res. 2018, 130, 451–465. [Google Scholar] [CrossRef]
- Kumar, P.S.; Saravanan, A.; Sheeba, N.; Uma, S. Structural, functional characterization and physicochemical properties of green banana flour from dessert and plantain bananas (Musa spp.). LWT 2019, 116, 108524. [Google Scholar] [CrossRef]
- Jia, M.; Yu, Q.; Chen, J.; He, Z.; Chen, Y.; Xie, J.; Nie, S.; Xie, M. Physical quality and in vitro starch digestibility of biscuits as affected by addition of soluble dietary fiber from defatted rice bran. Food Hydrocoll. 2020, 99, 105349. [Google Scholar] [CrossRef]
- Ye, J.; Hu, X.; Luo, S.; McClements, D.J.; Liang, L.; Liu, C. Effect of endogenous proteins and lipids on starch digestibility in rice flour. Food Res. Int. 2018, 106, 404–409. [Google Scholar] [CrossRef]
- Oates, C.G. Towards an understanding of starch granule structure and hydrolysis. Trends Food Sci. Technol. 1997, 8, 375–382. [Google Scholar] [CrossRef]
- McDougall, G.J.; Shpiro, F.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Different polyphenolic components of soft fruits inhibit α-amylase and α-glycosidase. J. Agric. Food Chem. 2005, 53, 2760–2766. [Google Scholar] [CrossRef]
- Chi, C.; Li, X.; Zhang, Y.; Chen, L.; Li, L. Understanding the mechanism of starch digestion mitigation by rice protein and its enzymatic hydrolysates. Food Hydrocoll. 2018, 84, 473–480. [Google Scholar] [CrossRef]
- López-Barón, N.; Sagnelli, D.; Blennow, A.; Holse, M.; Gao, J.; Saaby, L.; Müllertz, A.; Jespersen, B.; Vasanthan, T. Hydrolysed pea proteins mitigate in vitro wheat starch digestibility. Food Hydrocoll. 2018, 79, 117–126. [Google Scholar] [CrossRef]
- Oseguera-Toledo, M.E.; Gonzalez de Mejia, E.; Amaya-Llano, S.L. Hard-to-cook bean (Phaseolus vulgaris L.) proteins hydrolyzed by alcalase and bromelain produced bioactive peptide fractions that inhibit targets of type-2 diabetes and oxidative stress. Food Res. Int. 2015, 76, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Oseguera Toledo, M.E.; Gonzalez de Mejia, E.; Sivaguru, M.; Amaya-Llano, S.L. Common bean (Phaseolus vulgaris L.) protein-derived peptides increased insulin secretion, inhibited lipid accumulation, increased glucose uptake and reduced the phosphatase and tensin homologue activation in vitro. J. Funct. Foods 2016, 27, 160–177. [Google Scholar] [CrossRef]
- Claessens, M.; Calame, W.; Siemensma, A.D.; van Baak, M.A.; Saris, W.H.M. The effect of different protein hydrolysate/carbohydrate mixtures on postprandial glucagon and insulin responses in healthy subjects. Eur. J. Clin. Nutr. 2009, 63, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Kansal, R.; Kalia, V.; Tripathi, M.; Gupta, V.K. A kidney bean trypsin inhibitor with an insecticidal potential against Helicoverpa armigera and Spodoptera litura. Acta Physiol. Plant. 2014, 36, 525–539. [Google Scholar] [CrossRef]
- Butterworth, P.J.; Warren, F.J.; Ellis, P.R. Human α-amylase and starch digestion: An interesting marriage. Starch Staerke 2011, 63, 395–405. [Google Scholar] [CrossRef]
- Matsui, T.; Yoshimoto, C.; Osajima, K.; Oki, T.; Osajima, Y. In vitro survey of α–glucosidase inhibitory food components. Biosci. Biotechnol. Biochem. 1996, 60, 2019–2022. [Google Scholar] [CrossRef]
- Wright, E.M.; Sala-Rabanal, M.; Loo, D.D.F.; Hirayama, B.A. Sugar Absorption, 5th ed.; Academic Press-Elsevier: San Diego, CA, USA, 2012. [Google Scholar]
- Wolever, T.M.S.; Jenkins, D.J.A.; Jenkins, A.L.; Josse, R.G. The glycemic index: Methodology and clinical implications. Am. J. Clin. Nutr. 1991, 54, 846–854. [Google Scholar] [CrossRef]
- Litwack, G. Vitamins and Hormones: Incretins and Insulin Secretion, 1st ed.; Academic Press-Elsevier: Oxford, UK, 2010. [Google Scholar]
- Lopez, M.J.; Mohiuddin, S.S. Biochemistry, Essential Amino Acids; StatPearls: Tampa, FL, USA, 2021. [Google Scholar]
- Petsko, G.; Ringe, D. Protein Structure and Function; New Science Press Ltd.: Singapore, 2004. [Google Scholar]
- López-Barrios, L.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bioactive Peptides and Hydrolysates from Pulses and Their Potential Use as Functional Ingredients. J. Food Sci. 2014, 79, R273–R283. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Mojica, L.; de Mejía, E.G. Characterization and Comparison of Protein and Peptide Profiles and their Biological Activities of Improved Common Bean Cultivars (Phaseolus vulgaris L.) from Mexico and Brazil. Plant Foods Hum. Nutr. 2015, 70, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Malaguti, M.; Dinelli, G.; Leoncini, E.; Bregola, V.; Bosi, S.; Cicero, A.F.G.; Hrelia, S. Bioactive peptides in cereals and legumes: Agronomical, biochemical and clinical aspects. Int. J. Mol. Sci. 2014, 15, 21120–21135. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Marya Khan, H.; Nabavi, S.M.; Habtemariam, S. Anti-diabetic potential of peptides: Future prospects as therapeutic agents. Life Sci. 2018, 193, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Ngoh, Y.Y.; Gan, C.Y. Enzyme-assisted extraction and identification of antioxidative and α-amylase inhibitory peptides from Pinto beans (Phaseolus vulgaris cv. Pinto). Food Chem. 2016, 190, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef]
- Lemes, A.C.; Sala, L.; Ores, J.D.C.; Braga, A.R.C.; Egea, M.B.; Fernandes, K.F. A review of the latest advances in encrypted bioactive peptides from protein-richwaste. Int. J. Mol. Sci. 2016, 17, 950. [Google Scholar] [CrossRef]
- Venâncio, T.M.; Oliveira, A.E.A.; Silva, L.B.; Machado, O.L.T.; Fernandes, K.V.S.; Xavier-Filho, J. A protein with amino acid sequence.pdf. Braz. J. Med. Biol. Res. 2003, 36, 1167–1173. [Google Scholar] [CrossRef]
- Xavier-Filho, J.; Oliveira, A.E.A.; da Silva, L.B.; Azevedo, C.R.; Venâncio, T.M.; Machado, O.L.T.; Oliva, M.L.; Fernandes, K.V.S.; Xavier-Neto, J. Plant insulin or glucokinin: A conflicting issue. Braz. J. Plant Physiol. 2003, 15, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Kehinde, B.A.; Sharma, P. Recently isolated antidiabetic hydrolysates and peptides from multiple food sources: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 322–340. [Google Scholar] [CrossRef] [PubMed]
- Oseguera-Toledo, M.E.; González de Mejía, E.; Reynoso-Camacho, R.; Cardador-Martínez, A.; Amaya-Llano, S.L. Proteins and bioactive peptides. Nutrafoods 2014, 13, 147–157. [Google Scholar] [CrossRef]
- Patil, S.P.; Goswami, A.; Kalia, K.; Kate, A.S. Plant-Derived Bioactive Peptides: A Treatment to Cure Diabetes. Int. J. Pept. Res. Ther. 2020, 26, 955–968. [Google Scholar] [CrossRef]
- Yan, J.; Zhao, J.; Yang, R.; Zhao, W. Bioactive peptides with antidiabetic properties: A review. Int. J. Food Sci. Technol. 2019, 54, 1909–1919. [Google Scholar] [CrossRef]
- Moreno-Valdespino, C.A.; Luna-Vital, D.; Camacho-Ruiz, R.M.; Mojica, L. Bioactive proteins and phytochemicals from legumes: Mechanisms of action preventing obesity and type-2 diabetes. Food Res. Int. 2020, 130, 108905. [Google Scholar] [CrossRef] [PubMed]
- Khattab, R.Y.; Arntfield, S.D.; Nyachoti, C.M. Nutritional quality of legume seeds as affected by some physical treatments, Part 1: Protein quality evaluation. LWT-Food Sci. Technol. 2009, 42, 1107–1112. [Google Scholar] [CrossRef]
- Mojica, L.; Luna-Vital, D.A.; González de Mejía, E. Characterization of peptides from common bean protein isolates and their potential to inhibit markers of type-2 diabetes, hypertension and oxidative stress. J. Sci. Food Agric. 2017, 97, 2401–2410. [Google Scholar] [CrossRef]
- Castañeda-Pérez, E.; Jiménez-Morales, K.; Quintal-Novelo, C.; Moo-Puc, R.; Chel-Guerrero, L.; Betancur-Ancona, D. Enzymatic protein hydrolysates and ultrafiltered peptide fractions from Cowpea Vigna unguiculata L bean with in vitro antidiabetic potential. J. Iran. Chem. Soc. 2019, 16, 1773–1781. [Google Scholar] [CrossRef]
- de Souza Rocha, T.; Hernandez, L.M.R.; Mojica, L.; Johnson, M.H.; Chang, Y.K.; González de Mejía, E. Germination of Phaseolus vulgaris and alcalase hydrolysis of its proteins produced bioactive peptides capable of improving markers related to type-2 diabetes in vitro. Food Res. Int. 2015, 76, 150–159. [Google Scholar] [CrossRef]
- Tharanathan, R.N.; Mahadevamma, S. Grain legumes—A boon to human nutrition. Trends Food Sci. Technol. 2003, 14, 507–518. [Google Scholar] [CrossRef]
- de Souza Rocha, T.; Hernandez, L.M.R.; Chang, Y.K.; de Mejía, E.G. Impact of germination and enzymatic hydrolysis of cowpea bean (Vigna unguiculata) on the generation of peptides capable of inhibiting dipeptidyl peptidase IV. Food Res. Int. 2014, 64, 799–809. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Strategies for the discovery and identification of food protein-derived biologically active peptides. Trends Food Sci. Technol. 2017, 69, 289–305. [Google Scholar] [CrossRef]
- Mojica, L.; Chen, K.; de Mejía, E.G. Impact of Commercial Precooking of Common Bean (Phaseolus vulgaris) on the Generation of Peptides, After Pepsin-Pancreatin Hydrolysis, Capable to Inhibit Dipeptidyl Peptidase-IV. J. Food Sci. 2015, 80, H188–H198. [Google Scholar] [CrossRef] [PubMed]
- Mojica, L.; Gonzalez De Mejia Elvira Menjivar, M.; Granados-Silvestre, M.Á. Antidiabetic Effect of Black Bean Peptides through Reduction of Glucose Absorption and Modulation of SGLT1, GLUT2 and DPP-IV in in vitro and in vivo Models. FASEB J. 2016, 30, 125–126. [Google Scholar]
- Mojica, L.; de Mejía, E.G. Optimization of enzymatic production of anti-diabetic peptides from black bean (Phaseolus vulgaris L.) proteins, their characterization and biological potential. Food Funct. 2016, 7, 713–727. [Google Scholar] [CrossRef]
- Ngoh, Y.Y.; Gan, C.Y. Identification of Pinto bean peptides with inhibitory effects on α-amylase and angiotensin converting enzyme (ACE) activities using an integrated bioinformatics-assisted approach. Food Chem. 2018, 267, 124–131. [Google Scholar] [CrossRef]
- Jakubczyk, A.; Karaś, M.; Złotek, U.; Szymanowska, U. Identification of potential inhibitory peptides of enzymes involved in the metabolic syndrome obtained by simulated gastrointestinal digestion of fermented bean (Phaseolus vulgaris L.) seeds. Food Res. Int. 2017, 100, 489–496. [Google Scholar] [CrossRef]
- Ohara, A.; Cason, V.G.; Nishide, T.G.; Miranda de Matos, F.; de Castro, R.J.S. Improving the antioxidant and antidiabetic properties of common bean proteins by enzymatic hydrolysis using a blend of proteases. Biocatal. Biotransform. 2020, 39, 100–108. [Google Scholar] [CrossRef]
- Jiang, M.; Yan, H.; He, R.; Ma, Y. Purification and a molecular docking study of α-glucosidase-inhibitory peptides from a soybean protein hydrolysate with ultrasonic pretreatment. Eur. Food Res. Technol. 2018, 244, 1995–2005. [Google Scholar] [CrossRef]
- Nilsson, B.L.; Soellner, M.B.; Raines, R.T. Chemical Synthesis of Proteins. Annu. Rev. Biophys. Biomol. Struct. 2005, 34, 91–118. [Google Scholar] [CrossRef]
- Kent, S.B.H. Novel protein science enabled by total chemical synthesis. Protein Sci. 2019, 28, 313–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oseguera-Toledo, M.E.; De Mejia, E.G.; Dia, V.P.; Amaya-Llano, S.L. Common bean (Phaseolus vulgaris L.) hydrolysates inhibit inflammation in LPS-induced macrophages through suppression of NF-κB pathways. Food Chem. 2011, 127, 1175–1185. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.S.; Ying-Yuan, N.; Gan, C.Y. A comparative study of physicochemical characteristics and functionalities of pinto bean protein isolate (PBPI) against the soybean protein isolate (SPI) after the extraction optimisation. Food Chem. 2014, 152, 447–455. [Google Scholar] [CrossRef] [PubMed]
- de Castro, R.J.S.; Cason, V.G.; Sato, H.H. Binary mixture of proteases increases the antioxidant properties of white bean (Phaseolus vulgaris L.) protein-derived peptides obtained by enzymatic hydrolysis. Biocatal. Agric. Biotechnol. 2017, 10, 291–297. [Google Scholar] [CrossRef]
- Chel-Guerrero, L.; Pérez-Flores, V.; Betancur-Ancona, D.; Dávila-Ortiz, G. Functional Properties of Flours and Protein Isolates from Phaseolus lunatus and Canavalia ensiformis Seeds. J. Agric. Food Chem. 2001, 50, 584–591. [Google Scholar] [CrossRef]
- Mune Mune, M.A. Optimizing Functional Properties of Bambara Bean Protein Concentrate by Enzymatic Hydrolysis Using Pancreatin. J. Food Process. Preserv. 2015, 39, 2572–2580. [Google Scholar] [CrossRef]
- Mora-Escobedo, R.; Robles-Ramírez, M.D.C.; Ramón-Gallegos, E.; Reza-Alemán, R. Effect of protein hydrolysates from germinated soybean on cancerous cells of the human cervix: An In Vitro study. Plant Foods Hum. Nutr. 2009, 64, 271–278. [Google Scholar] [CrossRef]
- Los, F.G.B.; Zielinski, A.A.F.; Wojeicchowski, J.P.; Nogueira, A.; Demiate, I.M. Beans (Phaseolus vulgaris L.): Whole seeds with complex chemical composition. Curr. Opin. Food Sci. 2018, 19, 63–71. [Google Scholar] [CrossRef]
- Bautista-Expósito, S.; Peñas, E.; Dueñas, M.; Silván, J.M.; Frias, J.; Martínez-Villaluenga, C. Individual contributions of Savinase and Lactobacillus plantarum to lentil functionalization during alkaline pH-controlled fermentation. Food Chem. 2018, 257, 341–349. [Google Scholar] [CrossRef]
- Carbonaro, M.; Vecchini, P.; Carnovale, E. Protein Solubility of Raw and Cooked Beans (Phaseolus vulgaris): Role of the Basic Residues. J. Agric. Food Chem. 1993, 41, 1169–1175. [Google Scholar] [CrossRef]
- Megías, C.; Yust, M.D.M.; Pedroche, J.; Lquari, H.; Girón-Calle, J.; Alaiz, M.; Millán, F.; Vioque, J. Purification of an ACE Inhibitory Peptide after Hydrolysis of Sunflower (Helianthus annuus L.) Protein Isolates. J. Agric. Food Chem. 2004, 52, 1928–1932. [Google Scholar] [CrossRef] [PubMed]
- Jakubczyk, A.; Karaś, M.; Baraniak, B.; Pietrzak, M. The impact of fermentation and in vitro digestion on formation angiotensin converting enzyme (ACE) inhibitory peptides from pea proteins. Food Chem. 2013, 141, 3774–3780. [Google Scholar] [CrossRef]
- Karaś, M.; Baraniak, B.; Rybczyńska, K.; Gmiński, J.; Gaweł-Bęben, K.; Jakubczyk, A. The influence of heat treatment of chickpea seeds on antioxidant and fibroblast growth-stimulating activity of peptide fractions obtained from proteins digested under simulated gastrointestinal conditions. Int. J. Food Sci. Technol. 2015, 50, 2097–2103. [Google Scholar] [CrossRef]
- Megías, C.; Pedroche, J.; Yust, M.M.; Girón-Calle, J.; Alaiz, M.; Millán, F.; Vioque, J. Production of copper-chelating peptides after hydrolysis of sunflower proteins with pepsin and pancreatin. LWT-Food Sci. Technol. 2008, 41, 1973–1977. [Google Scholar] [CrossRef]
- Campos, M.R.S.; Guerrero, L.A.C.; Ancona, D.A.B. Angiotensin-I converting enzyme inhibitory and antioxidant activities of peptide fractions extracted by ultrafiltration of cowpea Vigna unguiculata hydrolysates. J. Sci. Food Agric. 2010, 90, 2512–2518. [Google Scholar] [CrossRef] [PubMed]
- Lunow, D.; Kaiser, S.; Brückner, S.; Gotsch, A.; Henle, T. Selective release of ACE-inhibiting tryptophan-containing dipeptides from food proteins by enzymatic hydrolysis. Eur. Food Res. Technol. 2013, 237, 27–37. [Google Scholar] [CrossRef]
- Vaštag, Ž.; Popović, L.; Popović, S.; Peričin-Starčević, I.; Krimer-Malešević, V. In vitro study on digestion of pumpkin oil cake protein hydrolysate: Evaluation of impact on bioactive properties. Int. J. Food Sci. Nutr. 2013, 64, 452–460. [Google Scholar] [CrossRef]
- Marcela, G.M.; Eva, R.G.; del Carmen, R.R.M.; Rosalva, M.E. Evaluation of the Antioxidant and Antiproliferative Effects of Three Peptide Fractions of Germinated Soybeans on Breast and Cervical Cancer Cell Lines. Plant Foods Hum. Nutr. 2016, 71, 368–374. [Google Scholar] [CrossRef]
- Yuan, X.; Gu, X.; Tang, J. Purification and characterisation of a hypoglycemic peptide from Momordica Charantia L. Var. abbreviata Ser. Food Chem. 2008, 111, 415–420. [Google Scholar] [CrossRef]
- Pan, S.; Wang, S.; Jing, L.; Yao, D. Purification and characterisation of a novel angiotensin-I converting enzyme (ACE)-inhibitory peptide derived from the enzymatic hydrolysate of Enteromorpha clathrata protein. Food Chem. 2016, 211, 423–430. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, J.; Tian, J.; Chen, S.; Ye, X.; Hu, Y.; Chen, J. Inhibitory mechanism of novel allosteric inhibitor, Chinese bayberry (Myrica rubra Sieb. et Zucc.) leaves proanthocyanidins against α-glucosidase. J. Funct. Foods 2019, 56, 286–294. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Worthington, V. Alpha amylase. In Worthington Enzyme Manual; Worthington Biochemical Corp.: Freehold, NJ, USA, 1993; pp. 36–41. [Google Scholar]
- Świeca, M.; Baraniak, B.; Gawlik-Dziki, U. In vitro digestibility and starch content, predicted glycemic index and potential in vitro antidiabetic effect of lentil sprouts obtained by different germination techniques. Food Chem. 2013, 138, 1414–1420. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Apostolidis, E.; Kwon, Y.I.; Ghaedian, R.; Shetty, K. Fermentation of Milk and Soymilk by Lactobacillus bulgaricus and Lactobacillus acidophilus Enhances Functionality for Potential Dietary Management of Hyperglycemia and Hypertension. Food Biotechnol. 2007, 21, 217–236. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Hernández-Ledesma, B. Release of dipeptidyl peptidase IV, α-amylase and α-glucosidase inhibitory peptides from quinoa (Chenopodium quinoa Willd.) during in vitro simulated gastrointestinal digestion. J. Funct. Foods 2017, 35, 531–539. [Google Scholar] [CrossRef]
- Siow, H.L.; Lim, T.S.; Gan, C.Y. Development of a workflow for screening and identification of α-amylase inhibitory peptides from food source using an integrated Bioinformatics-phage display approach: Case study—Cumin seed. Food Chem. 2017, 214, 67–76. [Google Scholar] [CrossRef]
- Hao, X.; Li, J.; Shi, Q.; Zhang, J.; He, X.; Ma, H. Characterization of a novel legumin α-amylase inhibitor from chickpea (Cicer arietinum L.) seeds. Biosci. Biotechnol. Biochem. 2009, 73, 1200–1202. [Google Scholar] [CrossRef]
- Guang, C.; Phillips, R.D. Plant food-derived angiotensin i converting enzyme inhibitory peptides. J. Agric. Food Chem. 2009, 57, 5113–5120. [Google Scholar] [CrossRef]
- Rudolph, S.; Lunow, D.; Kaiser, S.; Henle, T. Identification and quantification of ACE-inhibiting peptides in enzymatic hydrolysates of plant proteins. Food Chem. 2017, 224, 19–25. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, H.S.; Choi, J.W.; Ra, K.S.; Kim, J.M.; Suh, H.J. Novel tripeptides with α-glucosidase inhibitory activity isolated from silk cocoon hydrolysate. J. Agric. Food Chem. 2011, 59, 11522–11525. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Wang, M.H.; Rhee, H.I. A novel α-glucosidase inhibitor from pine bark. Carbohydr. Res. 2004, 339, 715–717. [Google Scholar] [CrossRef] [PubMed]
- Ranilla, L.G.; Kwon, Y.I.; Apostolidis, E.; Shetty, K. Phenolic compounds, antioxidant activity and in vitro inhibitory potential against key enzymes relevant for hyperglycemia and hypertension of commonly used medicinal plants, herbs and spices in Latin America. Bioresour. Technol. 2010, 101, 4676–4689. [Google Scholar] [CrossRef]
- Uraipong, C.; Zhao, J. Rice bran protein hydrolysates exhibit strong in vitro α-amylase, β-glucosidase and ACE-inhibition activities. J. Sci. Food Agric. 2016, 96, 1101–1110. [Google Scholar] [CrossRef]
- Ruiz-Ruiz, J.; Martínez-Ayala, A.; Drago, S.; González, R.; Betancur-Ancona, D.; Chel-Guerrero, L. Extrusion of a hard-to-cook bean (Phaseolus vulgaris L.) and quality protein maize (Zea mays L.) flour blend. LWT-Food Sci. Technol. 2008, 41, 1799–1807. [Google Scholar] [CrossRef]
- Velarde-Salcedo, A.J.; Barrera-Pacheco, A.; Lara-González, S.; Montero-Morán, G.M.; Díaz-Gois, A.; de Mejia, E.G.; de la Rosa, A.P.B. In vitro inhibition of dipeptidyl peptidase IV by peptides derived from the hydrolysis of amaranth (Amaranthus hypochondriacus L.) proteins. Food Chem. 2013, 136, 758–764. [Google Scholar] [CrossRef]
- Tulipano, G.; Sibilia, V.; Caroli, A.M.; Cocchi, D. Whey proteins as source of dipeptidyl dipeptidase IV (dipeptidyl peptidase-4) inhibitors. Peptides 2011, 32, 835–838. [Google Scholar] [CrossRef]
- Silveira, S.T.; Martínez-Maqueda, D.; Recio, I.; Hernández-Ledesma, B. Dipeptidyl peptidase-IV inhibitory peptides generated by tryptic hydrolysis of a whey protein concentrate rich in β-lactoglobulin. Food Chem. 2013, 141, 1072–1077. [Google Scholar] [CrossRef] [PubMed]
- Harnedy, P.A.; O’Keeffe, M.B.; Fitzgerald, R.J. Purification and identification of dipeptidyl peptidase (DPP) IV inhibitory peptides from the macroalga Palmaria palmata. Food Chem. 2015, 172, 400–406. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef]
- Dia, V.P.; Torres, S.; De Lumen, B.O.; Erdman, J.W.; De Mejia, E.G. Presence of lunasin in plasma of men after soy protein consumption. J. Agric. Food Chem. 2009, 57, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.; Chaves, I.; Carrilho, D.; Veloso, M.; Ricardo, C.P. Detection of novel trypsin inhibitors in the cotyledons of Phaseolus vulgaris seeds. J. Plant Physiol. 2010, 167, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.S.; Zhang, Y.; Ng, T.B. Brown kidney bean bowman-birk trypsin inhibitor is heat and pH stable and exhibits anti-proliferative activity. Appl. Biochem. Biotechnol. 2013, 169, 1306–1314. [Google Scholar] [CrossRef]
- Rui, X.; Boye, J.I.; Ribereau, S.; Simpson, B.K.; Prasher, S.O. Comparative study of the composition and thermal properties of protein isolates prepared from nine Phaseolus vulgaris legume varieties. Food Res. Int. 2011, 44, 2497–2504. [Google Scholar] [CrossRef]
- Montoya, C.A.; Lallès, J.P.; Beebe, S.; Leterme, P. Phaseolin diversity as a possible strategy to improve the nutritional value of common beans (Phaseolus vulgaris). Food Res. Int. 2010, 43, 443–449. [Google Scholar] [CrossRef]
- Carrasco-Castilla, J.; Hernández-Álvarez, A.J.; Jiménez-Martínez, C.; Jacinto-Hernández, C.; Alaiz, M.; Girón-Calle, J.; Vioque, J.; Dávila-Ortiz, G. Antioxidant and metal chelating activities of Phaseolus vulgaris L. var. Jamapa protein isolates, phaseolin and lectin hydrolysates. Food Chem. 2012, 131, 1157–1164. [Google Scholar] [CrossRef]
- Montoya, C.A.; Leterme, P.; Victoria, N.F.; Toro, O.; Souffrant, W.B.; Beebe, S.; Lallès, J.-P. Susceptibility of phaseolin to in vitro proteolysis is highly variable across common bean varieties (Phaseolus vulgaris). J. Agric. Food Chem. 2008, 56, 2183–2191. [Google Scholar] [CrossRef]
- Vasconcelos, I.M.; Maia, F.M.M.; Farias, D.F.; Campello, C.C.; Carvalho, A.F.U.; Moreira, R.D.A.; de Oliveira, J.T.A. Protein fractions, amino acid composition and antinutritional constituents of high-yielding cowpea cultivars. J. Food Compos. Anal. 2010, 23, 54–60. [Google Scholar] [CrossRef]
- López-Pedrouso, M.; Alonso, J.; Zapata, C. Evidence for phosphorylation of the major seed storage protein of the common bean and its phosphorylation-dependent degradation during germination. Plant Mol. Biol. 2014, 84, 415–428. [Google Scholar] [CrossRef]
- Sangronis, E.; Rodríguez, M.; Cava, R.; Torres, A. Protein quality of germinated Phaseolus vulgaris. Eur. Food Res. Technol. 2006, 222, 144–148. [Google Scholar] [CrossRef]
- Peñas, E.; Préstamo, G.; Gomez, R. High pressure and the enzymatic hydrolysis of soybean whey proteins. Food Chem. 2004, 85, 641–648. [Google Scholar] [CrossRef]
- Montoya, C.A.; Leterme, P.; Beebe, S.; Souffrant, W.B.; Mollé, D.; Lallès, J.P. Phaseolin type and heat treatment influence the biochemistry of protein digestion in the rat intestine. Br. J. Nutr. 2008, 99, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Unzón, H.Y.; Ortega-Delgado, M.L. Denaturation by heat, sodium dodecyl sulphate and dithiothreitol of globulins and phaseolin from dry bean (Phaseolus vulgaris L.). Plant Foods Hum. Nutr. 1988, 38, 211–223. [Google Scholar] [CrossRef]
- Orona-Tamayo, D.; Valverde, M.E.; Paredes-López, O. Bioactive peptides from selected latin american food crops—A nutraceutical and molecular approach. Crit. Rev. Food Sci. Nutr. 2019, 59, 1949–1975. [Google Scholar] [CrossRef] [PubMed]
- Mojica, L.; Luna-Vital, D.A.; Gonzalez de Mejia, E. Black bean peptides inhibit glucose uptake in Caco-2 adenocarcinoma cells by blocking the expression and translocation pathway of glucose transporters. Toxicol. Rep. 2018, 5, 552–560. [Google Scholar] [CrossRef]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Mojica, L.; Gonzalez de Mejia, E.; Granados-Silvestre, M.Á.; Menjivar, M. Evaluation of the hypoglycemic potential of a black bean hydrolyzed protein isolate and its pure peptides using in silico, in vitro and in vivo approaches. J. Funct. Foods 2017, 31, 274–286. [Google Scholar] [CrossRef]
- Liu, R.; Cheng, J.; Wu, H. Discovery of food-derived dipeptidyl peptidase IV inhibitory peptides: A review. Int. J. Mol. Sci. 2019, 20, 463. [Google Scholar] [CrossRef]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Peptides Derived from Soy and Lupin Protein as Dipeptidyl-Peptidase IV Inhibitors: In Vitro Biochemical Screening and in Silico Molecular Modeling Study. J. Agric. Food Chem. 2016, 64, 9601–9606. [Google Scholar] [CrossRef] [PubMed]
- Brayer, G.; Luo, Y.; Withers, S. The structure of human pancreatic a-amylase at 1.8 Å resolution and comparisons with related enzymes. Protein Sci. 1995, 4, 1730–1742. [Google Scholar] [CrossRef]
- Juillerat-Jeanneret, L. Dipeptidyl peptidase IV and its inhibitors: Therapeutics for type 2 diabetes and what else? J. Med. Chem. 2014, 57, 2197–2212. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.D.; Ghate, M.D. Recent approaches to medicinal chemistry and therapeutic potential of dipeptidyl peptidase-4 (DPP-4) inhibitors. Eur. J. Med. Chem. 2014, 74, 574–605. [Google Scholar] [CrossRef] [PubMed]
- Obiro, W.C.; Zhang, T.; Jiang, B. The nutraceutical role of the Phaseolus vulgaris α-amylase inhibitor. Br. J. Nutr. 2008, 100, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.H.; Gaspar, A.R.M. Structural properties of bioactive peptides with α-glucosidase inhibitory activity. Chem. Biol. Drug Des. 2018, 91, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. Features of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from dietary proteins. J. Food Biochem. 2019, 43, e12451. [Google Scholar] [CrossRef]
- Power, O.; Nongonierma, A.B.; Jakeman, P.; Fitzgerald, R.J. Food protein hydrolysates as a source of dipeptidyl peptidase IV inhibitory peptides for the management of type 2 diabetes. Proc. Nutr. Soc. 2014, 73, 34–46. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Dellafiora, L.; Paolella, S.; Galaverna, G.; Cozzini, P.; FitzGerald, R.J. In silico approaches applied to the study of peptide analogs of Ile-Pro-Ile in relation to their dipeptidyl peptidase IV inhibitory properties. Front. Endocrinol. 2018, 9, 329. [Google Scholar] [CrossRef] [Green Version]


| Legumes | Authors | Type of Processing | Protein Extraction Method | Sample Preparation | Sample: Water Ratio | Extraction | Precipitation | Method Reference | ||
|---|---|---|---|---|---|---|---|---|---|---|
| pH, T, t | Separation Technique | pH | Separation Technique | |||||||
| Common bean (Phaseolus vulgaris L.) | [59] | Raw and precooked | AE-IP |
| 1:10 (beans ground in a commercial blender) | pH 8 (0.1 M NaOH) 35 °C 1 h stirring |
| pH 4.3 (HCl) |
| - |
| Common bean (Phaseolus vulgaris L.) | [38] | Raw | AE-IP |
| 1:10 (in blender) | pH 8 (0.2 M NaOH) 35 °C 1 h agitation |
| pH 4.3 (HCl) |
| [68] |
| Common bean (Phaseolus vulgaris L.) | [53] | Raw | AE-IP |
| 1:10 (in blender) | pH 8 (0.2 M NaOH) 35 °C 1 h agitation |
| pH 4.3 (HCl) |
| [68] |
| Black bean (Phaseolus vulgaris L.) | [61] | Raw | AE-IP |
| 1:10 (in blender) | pH 8 (0.1 M NaOH) 35 °C 1 h stirring |
| pH 4.3 (0.1 M HCl) |
| - |
| Hard-to-cook bean (Phaseolus vulgaris L.) | [25] | Raw | AE-IP |
| 1:10 | pH 8.0 (0.2 N NaOH) 35 °C 1 h agitation |
| pH 4.3 (0.1 M HCl) |
| - |
| Common bean (Phaseolus vulgaris) | [55] | Germinated | AE-IP |
| NR | pH 8.0 (0.2 M NaOH) 35 °C 1 h agitation |
| pH 4.3 (1.0 M HCl) |
| [68] |
| Pinto bean (Phaseolus vulgaris cv. Pinto) | [42] | Raw | PBE |
| 1:20 (w/v) | pH 8 ± 0.1 (Phosphate buffer) 25 °C 1 h (250 rpm) |
| NA | NA | [69] |
| Pinto bean (Phaseolus vulgaris cv. Pinto) | [62] | Raw | PBE |
| 1:20 (w/v) | pH 8 ± 0.1 (Phosphate buffer) 25 °C 1 h (250 rpm) |
| NA | NA | [69] |
| Bean (Phaseolus vulgaris L. var. Eureka) | [63] | Fermented with L. plantarum 299v | WE |
| NR | NA |
| NA | NA | - |
| Easy-to-cook bean and Hard-to-cook bean (Phaseolus vulgaris L.) | [7] | Raw | AE-IP |
| 10 g flour in 50 mL water | pH 8.0 (0.1 mol/L NaOH) 35 °C 1 h stirring |
| pH 4.3 (diluted HCl) |
| [68] |
| Common bean/Carioca bean (Phaseolus vulgaris L. cv Carioca) | [64] | Raw | AE-IP |
| NR | pH 9.0 (0.1 mol/L NaOH) 1 h stirring |
| pH 4.3 (1 mol/L HCl) |
| [68,70] |
| Cowpea bean (Vigna unguiculata) | [57] | Germinated | AE-IP |
| NR | pH 8.0 (0.2 M NaOH) 35 °C 1 h agitation |
| pH 4.3 (1.0 M HCl) |
| [68] |
| Cowpea bean (Vigna unguiculata L.) | [54] | Raw | AE-IP | Milling | 1:6 (w/v) | pH 11 (T and t NR) |
| pH 4.5 |
| [71] |
| Bambara bean (Vigna subterranean) | [13] | Raw | AE-IP |
| 10 g flour in 100 mL 0.17 M NaCl | pH 8.9 4 °C 30 min stirring |
| pH 4.5 washed 2× 15 min |
| [72] |
| Soybean (Glycine max) | [16] | Germinated | AE-IP |
| NR | pH 9.0 (T and t NR) | NR | pH 4.5 | Isoflavones, phenolic, and saponin compounds extracted using 70% ethanol, 1 h agitation, Troom, until phenolic compounds not detected in alcoholic extract | [73] |
| Legumes | Authors | Protein Isolate Solution | Enzymes | Hydrolysis Condition | Hydrolysate Isolation | Cl/Fr/Se/Pf/Sy | Method Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Name | Specification | Ratio | pH | T (°C) | t (min) | Cessation Step | ||||||
| Common bean (Phaseolus vulgaris L.) | [59] | NR | Pepsin | Porcine | 1:20 (E/S, w/v) | 2.0 | 37 | 120 | Continued to pancreatin | Continued to pancreatin | [77] | |
| From pepsin | Pancreatin | 8 × USP | 1:20 (E/S, w/w) | 7.5 | 37 | 120 | 75 °C 20 min | C 20,000× g 15 min 4 °C S: FD | Clarification: 500 Da (salt elimination) | |||
| Common bean (Phaseolus vulgaris L.) | [38] | NR | Pepsin | Porcine 420 U/mg solid | 1:20 (E/S, w/v) | 2.0 | 37 | 120 | Continued to pancreatin | Continued to pancreatin | [77] | |
| From pepsin | Pancreatin | 8 × USP | 1:20 (E/S, w/w) | 7.5 | 37 | 120 | 75 °C 20 min | NR | Clarification: 0.45 µm Separation: In-gel tryptic digestion protein analysis | |||
| Common bean (Phaseolus vulgaris L.) | [53] | 10% (w/v) | Pepsin | Porcine | 1:20 (E/S, w/w) | 2 | 37 | 180 | Continued to pancreatin | Continued to pancreatin | [77] | |
| From pepsin | Pancreatin | 8 × USP | 1:20 (E/S, w/w) | 7.5 | 37 | 180 | 80 °C 20 min | NR | Clarification: 0.45 µm | |||
| Black bean (Phaseolus vulgaris L.) | [61] | 1:20 (w/v) protein in water. Autoclaved at 121 °C, 5 min | Protease K | NR | 1:20 1:30 1:50 ESR | 7.5 | 37 | 2 × 603 × 604 × 60 | 75 °C 20 min | C 20,000× g 15 min 4 °C S: FD | Clarification: 500 Da (salt elimination) | - |
| Pepsin | NR | 2.0 | 37 | |||||||||
| Trypsin | NR | 7.5 | 37 | |||||||||
| Papain | NR | 6.5 | 60 | |||||||||
| Flavourzyme | NR | 8.0 | 50 | |||||||||
| Thermolysin | NR | 8 | 50 | |||||||||
| Chymotrypsin | NR | 7.5 | 37 | |||||||||
| Alcalase | NR | 7.0 | 50 | |||||||||
| 1:20 (w/v) Alcalase hydrolysate in water | Pepsin | NR | 1:20 (E/S, w/w) | 2.0 | 37 | 2 × 60 | Continued to pancreatin | Continued to pancreatin | ||||
| From pepsin | Pancreatin | NR | 1:20 (E/S, w/w) | 7.5 | 37 | 2 × 60 | 75 °C 20 min | C 20,000× g 15 min 4 °C S: FD | Clarification: 500 Da (salt elimination) | |||
| Hard-to-cook bean (Phaseolus vulgaris L.) | [25] | 1:10 (w/v) Protein in water. Incubated at pH 7.0, 70 °C, 15 min | Alcalase | Bacillus licheniformis | 1:17 ESR | 8 (0.5 N NaOH) | 50 | NR | 75 °C 20 min Continued to pepsin | C 14,000× g 30 min 4 °C S: FD | Clarification: 3 kDa (salt removal) | [68,77] |
| Bromelain | Pineapple | 1:17 ESR | 7 (0.5 N NaOH) | 45 | NR | 75 °C 20 min Continued to Pepsin | C 14,000× g 30 min 4 °C S: FD | |||||
| From Alcalase and bromelain | Pepsin | 662 units/mg | 1:20 ESR | 2.0 | 37 | 3 × 60 | Continued to pancreatin | Continued to pancreatin | ||||
| From pepsin | Pancreatin | 8 × USP | 1:20 ESR | 7.5 | 37 | 3 × 60 | 75 °C 20 min | C 20,000× g 15 min 4 °C | Fractionation: 1, 3, 5, and 10 kDa | |||
| Common bean (Phaseolus vulgaris) | [55] | 8:100 (w/v) Incubated at pH 8.0 (0.5 M NaOH), 50 °C, 10 min | Alcalase | Bacillus licheniformis 2.4 AU/g | 0.75 AU/g of protein | 8.0 (0.5 M NaOH) | NR | 0 1 × 60 2 × 60 3 × 60 4 × 60 | 0.1 M HCl 1.2 mL Continued to pepsin | C 14,000× g 30 min 10 °C S: FD | Clarification: 0.8 kDa (salt elimination) | - |
| From Alcalase | Pepsin | Porcine | 1:20 (E/S, w/w) | 2.0 | 37 | 120 | Continued to pancreatin | Continued to pancreatin | [77] | |||
| From pepsin | Pancreatin | NR | 1:20 (E/S, w/w) | 7.5 | 37 | 120 | 75 °C 20 min | C 20,000× g 15 min 4 °C S: FD | ||||
| Pinto bean (Phaseolus vulgaris cv. Pinto) | [42] | 1:10 (w/v) Protein in phosphate buffer | Protamex | NR | 1:10 1:30 1:50 (E/S, w/v) | 6.5 7.5 8.5 | 50 | 30 60 90 | 95 °C 30 min | C 15 min 4 °C S stored at −20 °C | Fractionation: 3, 10, 30, and 50, 100 kDa | - |
| Pinto bean (Phaseolus vulgaris cv. Pinto) | [62] | 1:10 (w/v) Protein in phosphate buffer | Protamex | NR | 1:10 1:30 1:50 (E/S, w/v) | 6.5 7.5 8.5 | 50 | 30 60 90 | 95 °C 30 min | C 15 min 4 °C S stored at −20 °C | Fractionation: 3, 10, 30, 50, and 100 kDa Synthesis: Mimotopes, Clayton, VIC, Australia | - |
| Bean (Phaseolus vulgaris L. var. Eureka) | [63] | 4% (w/v) protein in salt solution (7 mM NaHCO3 and 0.35 mM NaCl) Incubated at 37 °C, 5 min | α-Amylase | Hog pancreas 50 U/mg | 1:10 ESR | NR | 37 | 10 | Continued to pepsin | Continued to pepsin | [78,79] | |
| From α-amylase | Pepsin | Porcine gastric mucosa 250 units/mg | 1:100 ESR | 2.5 (1 M HCl) | 37 | 120 | Continued to pancreatin | Continued to pancreatin | ||||
| From pepsin | Pancreatin (0.7%) and bile extract (2.5%) | Porcine pancreas | 1:2.5 ESR | Neutral (1 M NaOH) | 37 | 60 | 100 °C 5 min | NR | Fractionation: 3.5 and 7.0 kDa Separation: Sephadex G10 | |||
| Easy-to-cook bean and hard-to-cook bean (Phaseolus vulgaris L.) | [7] | NR | Pepsin | NR | 1:20 (E/S, w/w) | 2.0 | 37 | 120 | Continued to pancreatin | Continued to pancreatin | [80] | |
| From pepsin | Pancreatin | NR | 1:20 (E/S, w/w) | 7.5 | 37 | 120 | 80 °C 20 min | C 5000× g 10 min 4 °C S: FD | Clarification: 0.45 µm Fractionation: 3 and 10 kDa | |||
| Common bean/Carioca bean (Phaseolus vulgaris L. cv Carioca) | [64] | 100 mg/mL | Simplex centroid mixture design of: Flavourzyme-Alcalase-Neutrase | FlavourzymeTM 500 L Aspergillus oryzae AlcalaseTM 2.4 L Bacillus licheniformis NeutraseTM 0.8 L Bacillus amyloliquefaciens | 50 U/mL | 7 | 50 | 120 | 100 °C 20 min | C 10,000× g 20 min 5 °C S: FD | - | |
| Cowpea bean (Vigna unguiculata) | [57] | 8:100 (w/v) Incubated at pH 8.0 (0.5 M NaOH), 50 °C, 10 min | Alcalase | Bacillus licheniformis 2.4 AU/g | 0.75 AU/g of protein | 8.0 (maintained by 0.5 M NaOH) | NR | 0 1 × 60 2 × 60 3 × 60 4 × 60 | 0.1 M HCl 1.2 mL Continued to pepsin | C 14,000× g 30 min 10 °C S: FD | Clarification: 0.8 kDa (salt elimination) | - |
| From Alcalase | Pepsin | Porcine | 1:20 (E/S, w/w) | 2.0 | 37 | 120 | Continued to pancreatin | Continued to pancreatin | [77] | |||
| From pepsin | Pancreatin | NR | 1:20 (E/S, w/w) | 7.5 | 37 | 120 | 75 °C 20 min | C 20,000× g 15 min 4 °C S: FD | ||||
| Cowpea bean (Vigna unguiculata L.) | [54] | 4% (w/v) | Alcalase | Alcalasa® Bacillus licheniformis | 0.3 AU/g protein isolate | 8.0 | 50 | 45 | Continued to pepsin | Continued to pepsin | [81] | |
| 4% (w/v) | Flavourzima | Flavourzima® Aspergillus oryzae | 50 UAPL/g protein isolate | 7.0 | 50 | 45 | Continued to pepsin | Continued to pepsin | ||||
| From Alcalase and Flavourzima | Pepsin | Sigma P70007 Porcine gastric mucosa | 1:10 ESR | 2 | 37 | 45 | Continued to pancreatin | Continued to pancreatin | ||||
| From pepsin | Pancreatin | Sigma 1750 Pig pancreas | 1:10 ESR | 7.5 | 37 | 45 | NR | NR | Fractionation: 1, 3, 5, and 10 kDa | |||
| Bambara bean (Vigna subterranean) | [13] | 5% (w/v) protein in double-distilled water | Alcalase | Bacillus licheniformis ≥2.4 U/g protein | 4% ESR | 7 (adjusted every 30 min, 0.5 M NaOH) | NR | 24 × 60 | 95 °C 5 min | C 8000× g 10 min 4 °C S: FD | [82] | |
| Trypsin | Porcine pancreas 1.5 U/g protein | 1% ESR | 7 (adjusted every 30 min, 0.5 M NaOH) | 55 | 24 × 60 | 95 °C 5 min | C 8000× g 10 min 4 °C S: FD | |||||
| Thermolysin | Geobacillus stearothermophilus 0.03–0.17 U/g protein | 1% ESR | 8 (adjusted every 30 min, 0.5 M NaOH) | 70 | 24 × 60 | 95 °C 5 min | C 8000× g 10 min 4 °C S: FD | Separation: RP-HPLC | ||||
| 20 mg hydrolysate/mL in 0.01 M HCl | Pepsin | Porcine gastric mucosa 3200–4500 U/mg protein | 0.04 mg/mg ESR | 2.10 | 37 | 30 | Continued to α-chymotrypsin and trypsin | [83] | ||||
| From pepsin | α-Chymotrypsin | Bovine pancreas ≥0.04 U/g protein | 0.02 mg/mg ESR | 7.5 | Continued to pancreatin | |||||||
| Trypsin | Porcine pancreas 1.5 U/g protein | 0.08 mg/mg ESR | 7.5 | |||||||||
| From α-chymotrypsin and trypsin | Pancreatin | NR | NR | 7.5 | 37 | 90 | 100 °C 5 min | C 8000× g 10 min 4 °C S: FD | ||||
| Soybean (Glycine max) | [16] | 5% (w/v) protein in distilled water | Pepsin | Porcine gastric mucosa 250 units/mg solid | 4% (w/v, protein basis) | 2.0 (1N HCl) | 37 | 60 | Continued to pancreatin | Continued to pancreatin | [84] | |
| From pepsin | Pancreatin | Porcine pancreas 8 × USP | 4% (w/v, protein basis) | 7.5 (1 N NaOH) | 37 | 120 | 100 °C 10 min | C 16,000× g 10 min S: FD |
| |||
| Soybean | [65] | 6:10 (w/v) protein in distilled water Ultrasonication (65 kHz, 245 W), Troom, 20 min Cooled to 37 °C. Diluted to 1.0% pH adjusted to 8.0 (1 M NaOH), incubated at 37 °C for 5 min | Trypsin | 6400 U/g | NR | 8.0 (1 M NaOH) | 37 | 22 | 90 °C 10 min | C 8000× g 25 min |
| [85,86] |
| Soy | [87] | 2% (w/v) protein in distilled water Heated to 100 °C for 10 min. Let cool | Alkaline Protease | 200 U/mg | 6000 U/g protein content in slurry | 9 | 50 | Until DH stable | 100 °C 10 min Continued to pepsin | C 4.436× g 10 min Stored at −20 °C | - | |
| Papain | 800 U/mg | 6.5 | 60 | Until DH stable | - | |||||||
| Trypsin | NR | 7 | 37 | Until DH stable | - | |||||||
| 10 mg/mL hydrolysate in deionized water | Pepsin | Porcine gastric mucosa 500/mg | 2.5 U/mg substrate | 2 (1 M HCl) | 37 (stirring waterbath) | 2 × 60 | Continued to pancreatin | Continued to pancreatin | [88] | |||
| From Pepsin | Pancreatin | Porcine pancreas | 4:100 ESR | 7.2 (1 M NaOH) | 37 | 2 × 60 | 95 °C 20 min | C 8000 rpm 10 min | Isolation: DEAE-52, Sephadex G-15 | |||
| Yellow field pea (Pisum sativum L.) | [9] | 5% protein in double-distilled water | Alcalase | NR | 4:100 ESR | NR | NR | 4 × 60 | pH 5.0 (2 M NaOH, 2 M HCl) 95 °C 15 min | C 10,000× g 15 min 4 °C S: FD | Fractionation: 1, 3, 5, and 10 kDa | - |
| Chymotripsin | NR | NR | NR | |||||||||
| Pepsin | NR | NR | NR | |||||||||
| Trypsin | NR | NR | NR | |||||||||
| Legumes | Authors | α-Amylase Type | Volume, Activity, Concentration, pH | Results Range | Method Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Enzyme | Inhibitor | Control (Acarbose) | Substrate (Starch) | Buffer | % Inhibition | IC50 | ||||
| Common bean (Phaseolus vulgaris L.) | [59] | Porcine pancreas type VI-B | 500 µL 13 U/mL buffer | 500 µL | 500 µL 1 mM | 500 µL 1% soluble starch in buffer | 0.02 M Na-phosphate buffer, pH 6.9 | Hydrolysates: ~4–36% rel ac/mg protein | - | - |
| Common bean (Phaseolus vulgaris L.) | [38] | Porcine pancreas type VI-B | 500 µL 13 U/mL buffer | 500 µL | 500 µL 1 mM | 500 µL 1% soluble starch in buffer | 0.02 M Na-phosphate buffer, pH 6.9 | Hydrolysates: 2.5–14.9% rel ac/mg BPI | - | [59] |
| Black bean (Phaseolus vulgaris L.) | [61] | Porcine pancreas type VI-B | 500 µL 13 U/mL buffer | 500 µL 1 mg DM/mL buffer | 500 µL 1 mM | 500 µL 1% soluble starch in buffer | 0.02 M Na-phosphate buffer, pH 6.9 | Hydrolysates: 13.0–61.8% inhibition/mg dry hydrolysate | - | [10] |
| Hard-to-cook bean (Phaseolus vulgaris L.) | [25] | B. subtilis | 500 µL 10.8 U/mL | 500 µL 100 µg/mL | 500 µL 1 mM | 500 µL 1% starch in buffer | 0.02 mM phosphate buffer with 6 mM NaCl, pH 6.9 | Hydrolysates: ~15–50% rel ac Peptide fractions: ~5–51% rel ac | - | - |
| Common bean (Phaseolus vulgaris) | [55] | Porcine pancreas type VI-B | 500 µL 13 U/mL buffer | 500 µL 1 mg/mL | 500 µL 1 mM | 500 µL 1% soluble starch in buffer | 0.02 M Na-phosphate buffer, pH 6.9 | 7.61–30.88% rel ac | - | - |
| Pinto bean (Phaseolus vulgaris cv. Pinto) | [42] | NR | 500 µL 0.5 mg/mL buffer | 100 µL | Control: enzyme, starch, without inhibitor Blank: starch, inhibitor, without enzyme | 500 µL 1% starch in buffer | 0.02 M Na-phosphate buffer in 6 mM NaCl, pH 6.9 | Hydrolysates: 15.78–57.48% Peptide fractions: 0.17–62.1% | - | [89] |
| Pinto bean (Phaseolus vulgaris cv. Pinto) | [62] | Human saliva | 500 µL 0.5 mg/mL buffer | 100 µL 1 mg/mL | Control: enzyme, starch, without inhibitor Blank: starch, inhibitor, without enzyme | 500 µL 1% (w/v) starch in buffer | 0.02 M, Na-phosphate buffer in 6 mM NaCl, pH 6.9 | Peptide fractions: ~5–58% inhibition/100 µg pinto bean peptides | Synthetic peptide fractions: 23.33–10.03 mM | [89] |
| Bean (Phaseolus vulgaris L. var. Eureka) | [63] | Hog pancreas (50 U/mg) | 0.25 mL | 0.25 mL | Enzyme, starch, without inhibitor | 0.5 mL 1% (w/v) soluble starch in buffer | 100 mM phosphate buffer containing 6 mM NaCl, pH 7.0 | - | Peptide fractions: 2.12– 0.038 µg/mL | [90] |
| Easy-to-cook bean and hard-to-cook bean (Phaseolus vulgaris L.) | [7] | Porcine pancreas | 500 µL 10 U/mL buffer | 500 µL 10 mg/mL | 500 µL 10 mg/mL | 500 µL 1% starch in buffer | 50 mmol/L Na-phosphate buffer, pH 6.9 | Naturally occurring peptide fractions: 3.0–89.1% Peptide fractions in hydrolysates: 1.3–53.4% Acarbose 32.8% | - | [59,91] |
| Common bean/Carioca bean (Phaseolus vulgaris L. cv Carioca) | [64] | B. licheniformis (Termamyl® 2X) | 0.5 mL in buffer | 0.5 mL 1 mg/mL | Enzyme, starch, distilled water | 0.5 mL 1% (w/v) starch in buffer | 50 mmol/L phosphate buffer, pH 7 | Non hydrolysed protein: 4.67% Hydrolysates: 30.05–101.61% | - | [91,92] |
| Cowpea bean (Vigna unguiculata L.) | [54] | Porcine pancreas type VI-B | 200 µL 13 U/mL buffer | 200 µL For PP: 100 mg/mL water For AF-PP: 50 mg/mL water | Details not reported | 200 µL 1% starch in buffer | 20 mM Na-phosphate buffer, pH 6.9 | Hydrolysates and peptide fractions: 24.39–96.81% Acarbose: 98.41% | - | [59] |
| Soybean (Glycine max) | [16] | Porcine pancreas | 100 µL 2 U/mL buffer | 50 µL Hydrolysate: 0.2–4 mg/mL Peptide fractions: 1 mg/mL | 50 µL 0.1–1.3 mg/mL Negative control: 50 µL distilled water | 100 µL 1% potato soluble starch in buffer | 0.02 M Na-phosphate buffer, pH 6.9 | Peptide fractions: ~84% not detected, relative to negative control | Hydrolysate and peptide fractions: >10.00– 1.7 mg peptide/mL Acarbose: 0.16 mg acarbose /mL | [93] |
| Yellow field pea (Pisum sativum L.) | [9] | Porcine pancreas | 100 µL 28.57 µg/mL buffer | 100 µL 50–225 µg/mL buffer | 100 µL 1.5–3 µg/mL buffer | 100 µL 1 g/100 mL starch in buffer | 0.02 M Na-phosphate buffer, pH 6.9 | Hydrolysates: ~7–30% Peptide fractions: ~2–30% Acarbose: ~45–74% | - | [94] |
| Legumes | Authors | α-Glucosidase Type | Volume, Activity, Concentration, pH | Results Range | Method Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Enzyme | Inhibitor | Control (Acarbose) | Substrate | Buffer | % Inhibition | IC50 | ||||
| Common bean (Phaseolus vulgaris L.) | [59] | S. cerevisiae | 100 µL 1 U/mL buffer | 50 µL | 50 µL 1 mM | 50 µL PNPG 5 mM in buffer | 0.1 M Na-phosphate buffer, pH 6.9 | Hydrolysates: ~48–68% inh AC/mg protein | - | - |
| Common bean (Phaseolus vulgaris L.) | [53] | S. cerevisiae | 100 µL 1 U/mL buffer | 50 µL 1 mg DW/mL | 50 µL Acarbose 1 mmol/L | 50 µL PNPG 5 mmol/L in buffer | 0.1 mol/L Na-phosphate buffer, pH 6.9 | Hydrolysates: 46.90–50.10% Peptides: 36.30–49.34% | - | - |
| Black bean (Phaseolus vulgaris L.) | [61] | S. cerevisiae | 100 µL 1 U/mL buffer | 50 µL 1 mg DM/mL buffer | 50 µL 1 mmol/L | 50 µL PNPG 1 mM in buffer | 0.1 M Na-phosphate buffer, pH 6.9 | Hydrolysates: 3.6–78.4% inhibition/mg dry hydrolysate | - | [10] |
| Hard-to-cook bean (Phaseolus vulgaris L.) | [25] | S. cerevisiae | 100 µL 1.0 U/mL buffer | 50 µL | 50 µL 1 mM | 50 µL PNPG 5 mM in buffer | 0.1 M phosphate buffer, pH 6.9 | Hydrolysates: ~37–65% rel ac Peptide fractions: ~50–74% rel ac | - | [92] |
| Easy-to-cook bean and hard-to-cook bean (Phaseolus vulgaris L.) | [7] | NR | 60 µL 2 U/mL | 60 µL 10 mg protein/mL | 60 µL 10 mg/mL | 60 µL PNPG 0.1 mol/L | 0.5 mol/L phosphate buffer, pH 6.8 | Naturally occurring peptide fractions: 35.9–89.2% Peptide fractions in hydrolysates: 22.9–53.5% Acarbose 76.0% | - | [59] |
| Common bean/Carioca bean (Phaseolus vulgaris L. cv Carioca) | [64] | S. cerevisiae | 100 µL 0.1 U/mL | 50 µL 1 mg/mL buffer | Distilled water | 50 µL PNPG 5 mmol/L | 50 mmol/L phosphate buffer, pH 7 | Non-hydrolysed protein: 19.23% Hydrolysates: 8.67–34.73% | - | [92] |
| Cowpea bean (Vigna unguiculata L.) | [54] | S. cerevisiae (SLBP0778V) | 20 µL 2 U/mL buffer | 20 µL For PP: 200 mg protein/mL water For AF-PP: 10 mg protein/mL water | Details not reported | 20 µL PNPG 1 mM | 50 mM K-phosphate buffer, pH 6.8 | Hydrolysates and peptide fractions: 4.069–97.34% Acarbose: 90.18% | - | [59] |
| Soybean (Glycine max) | [16] | Rat intestine | 50 µL 1 U/mL buffer | 100 µL 1–10 mg/mL | 100 µL Negative control: distilled water | 50 µL Maltose 2 mM | 0.1 M maleat buffer, pH 6.9 | Peptide fractions (substrate maltose): ~20–32% | Hydrolysate and peptide fractions (substrate maltose): >10.00–2.56 mg/mL Acarbose: 0.07 mg/mL | [93] |
| 50 µL 1 U/mL buffer | 100 µL 1–10 mg/mL | 100 µL Negative control: distilled water | 50 µL Sucrose 20 mM | 0.1 M maleat buffer, pH 6.9 | Peptide fractions (substrate sucrose): ~8–22% | Hydrolysate and peptide fractions (substrate sucrose): 5.27–1.23 mg/mL Acarbose: 0.03 mg/mL | - | |||
| Soybean | [65] | NR | 0.2 mL 0.15 U/mL | 0.1 mL | Control 1:bBuffer, substrate, enzyme Control 2: buffer, enzyme Control 3: enzyme, inhibitor | 0.2 mL PNPG 50 mM in buffer | 0.1 M K-phosphate buffer, pH 6.8 | - | Hydrolysate: 1.93 mg/mL Peptides < 5 kDa: 0.27 mg/mL Peptides > 5 kDa: 3.31 mg/mL Glu-Ser-Arg: 20.4 μM Glu-ala-Lys: 520.2 μM | [98] |
| Soy | [12] | S. cerevisiae | 10 µL 0.2 U/mL buffer | 50 µL | 10 mg/mL buffer | 50 µL PNPG 1 mg/mL buffer | 0.2 M Na-phosphate buffer | Hydrolysates: ~30–80% Alkaline protease H1 Fraction from DEAE-52: 87.10 ± 2.70% Alkaline protease H1 fraction from DEAE-52 then H-12 fraction from Sephadex G-15: 95.35 ± 2.70% | Alkaline protease hydrolysate: 4.94 ± 0.07 mg/mL Acarbose: 0.52 ± 0.05 mg/mL | [99] |
| Yellow field pea (Pisum sativum L.) | [9] | Rat intestinal acetone powder | 50 µL 8.33 mg/mL buffer | 50 µL 5–20 mg/mL buffer | 50 µL 0.00625–0.125 mg/mL buffer | 100 µL PNPG 5 mM in buffer | 0.1 M Na-phosphate buffer, pH 6.9 | Hydrolysates: ~8–47% Peptide fractions: ~6–53% Acarbose: ~45–67% | - | [8,100,101] |
| Legumes | Authors | DPP-IV Inhibitor Kit | DPP-IV Type | Volume, Activity, Concentration, pH | Results Range | Method Reference | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Enzyme | Inhibitor | Control | Blank | Substrate | Assay Buffer | % Inhibition | IC50 | |||||
| Common bean (Phaseolus vulgaris L.) | [59] | DPP-IV Glo Protease assay (Promega, G8351) | Purified DPP-IV, human enzyme | 10 µL 100 ng/mL | 40 µL 1 mg DW/mL | 40 µL Enzyme control | 50 µL Assay buffer | 50 µL DPP-IV Glo reagent | 100 mM Tris pH 8.0 200 mM NaCl 1 mM EDTA | - | Hydrolysates: ~1–0.1 mg protein/mL | - |
| Common bean (Phaseolus vulgaris L.) | [53] | DPP-IV Glo Protease assay (Promega, G8351) | Purified DPP-IV, human enzyme (D4943) | 10 µL 10 ng/mL | 40 µL 1 mg DW/mL | 40 µL Enzyme control | 50 µL Assay buffer | 50 µL DPP-IV Glo reagent | 100 mmol/L Tris pH 8.0 200 mmol/L NaCl 1 mmol/L EDTA | - | Hydrolysates: 0.33–0.14 mg DW/mL Peptides: 0.87–0.03 mg DW/mL Diprotin A: 0.02 mg DW/mL | - |
| Black bean (Phaseolus vulgaris L.) | [61] | DPP-IV Glo Protease assay (Promega, G8351) | Purified DPP-IV, human enzyme | 10 µL 10 ng/mL | 40 µL 1 mg DW/mL | 40 µL Enzyme control | 50 µL Assay buffer | 50 µL DPP-IV Glo reagent | 100 mM Tris pH 8.0 200 mM NaCl 1 mM EDTA | Hydrolysates: 13.9–96.7% | - | - |
| Hard-to-cook bean (Phaseolus vulgaris L.) | [25] | DPP-IV (Sigma Aldrich, protocol SSGPNA01) | DPP-IV, porcine kidney | 100 ng/mL | 100 µg/mL | NR | NR | 500 µM Gly-Pro-4-Nitroanilide | 100 mM Tris pH 8.0 | Hydrolysates: ~5–55% | - | [103] |
| Common bean (Phaseolus vulgaris) | [55] | DPP-IV Glo Protease assay (Promega, G8351) | Purified DPP-IV, human enzyme (≥1.0 U/vial) | 10 µL 100 ng/mL | 40 µL 0.1–4.0 mg/mL buffer | Enzyme control: 40 µL | 50 µL Assay buffer | 50 µL DPP-IV Glo reagent | 100 mM Tris pH 8.0 200 mM naCl 1 mM EDTA | - | Hydrolysates: ~3.3–0.75 mg hydrolysate/mL | - |
| Cowpea bean (Vigna unguiculata) | [57] | DPP-IV Glo Protease assay (Promega, G8351) | Purified DPP-IV, human enzyme | 10 µL 100 ng/mL | 40 µL 0.1–4.0 mg/mL buffer | Enzyme control: 40 µL | 50 µL Assay buffer | 50 µL DPP-IV Glo reagent | 100 mM Tris pH 8.0 200 mM NaCl 1 mM EDTA | - | Hydrolysates: ~3.0–0.5 mg hydrolysate/mL | |
| Cowpea bean (Vigna unguiculata L.) | [54] | DPP-IV (Sigma-Aldrich, MAK203) | NR | NR | NR | Sitagliptin | NR | NR | NR | Hydrolysates and peptide fractions: 67.65–85% Sitagliptin: 97.77% | - | - |
| Bambara bean (Vigna subterranean) | [13] | DPP-IV Drug Discovery Kit (Enzo Life Sciences) | Recombinant-soluble human DPP-IV | 15 µL 0.26 mU/test well | 50 µL/well 1 mg/mL | Diprotin A | NR | 50 µL 100 µM H-Gly-Pro-p-nitroaniline in assay buffer | NR | Hydrolysates: 7.981 ± 0.240–44.253 ± 1.327% at 1 mg/mL Simulated GI digesta: 8.996 ± 0.043–29.276 ± 0.878% at 1 mg/mL | Hydrolysates: 1.733→2.5 mg/mL | [104] |
| Soybean (Glycine max) | [16] | DPP-IV Drug Discovery Kit (Enzo Life Sciences) | Recombinant-soluble human DPP-IV | 15 µL 0.26 mU/test well | 50 µL/well 0.08–5 mg/mL | Positive control: 50 µL/well Diprotin A 0.78–50 µM | NR | 50 µL 100 µM H-Gly-Pro-p-nitroaniline in assay buffer | NR | - | Hydrolysate and peptide fractions: 2.21–0.91 mg/mL Diprotin A: 0.003 mg/mL | [105] |
| Soy | [12] | - | DPP-IV from human | 50 µL 0.02 U/mL buffer | 25 µL | Buffer to replace enzyme | Buffer to replace inhibitor and enzyme | 25 µL 12 mM Gly-Pro-p-nitroanilide | Tris buffer pH 8.0 | Hydrolysates: ~40–47% | Alkaline protease hydrolysate: 2.73 ± 0.08 mg/mL | [106] |
| Name of Online Tools | Link | Function | References |
|---|---|---|---|
| RSCB PDB | http://www.rcsb.org/pdb/home/home.do (accessed on 5 August 2020) | To retrieve the 3D crystal structure of enzymes. | [25,53,55,57,61,62,65,122] |
| UniProt | http://expasy.org/ (accessed on 5 August 2020) | To provide access to protein databases and software tools. | [122,123] |
| BLAST | https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 5 August 2020) | To confirm regions of similarity between biological sequences. | [25,38,53,55,57,59,61] |
| PeptideDB | http://www.peptides.be (accessed on 5 August 2020) | To validate the novelty of peptides. | [62] |
| PepDraw | http://www.tulane.edu/~biochem/WW/PepDraw/ (accessed on 5 August 2020) | To predict peptide structures and physicochemical properties. | [16,38,53,55,57,59,122] |
| I-TASSER | https://zhanglab.dcmb.med.umich.edu/I-TASSER/ (accessed on 5 August 2020) | To predict protein structure and structure-based functions. | [122,124] |
| BIOPEP | http://www.uwm.edu.pl/biochemia/index.php/pl/biopep (accessed on 5 August 2020) | To predict potential biological activities of peptides. | [16,25,38,53,55,57,59,61,62,122] |
| PeptideRanker | http://distilldeep.ucd.ie/PeptideRanker/ (accessed on 5 August 2020) | To predict peptide bioactivity potential. | [42,62] |
| Pepsite2 | http://pepsite2.russelllab.org/ (accessed on 5 August 2020) | To predict the binding sites and analyze the binding mechanisms of bioactive peptides. | [62] |
| GRAMM-X | http://vakser.compbio.ku.edu/resources/gramm/grammx/ (accessed on 5 August 2020) | To simulate molecular docking. | [25,55] |
| Rosetta FlexPepDock | http://flexpepdock.furmanlab.cs.huji.ac.il/ (accessed on 5 August 2020) | To refine the peptide–protein docking complex models. | [25,55] |
| Name of Software | Function | References |
|---|---|---|
| ChemBio3D Ultra | To convert 2D peptides to 3D | [65] |
| Instant MarvinSketch | To design peptides | [53] |
| Maestro | To model protein structure | [122,124] |
| VEGA suites | To build a canonical α-helix of peptides | [126] |
| AutoDock Vina |
| [65,122] |
| AutoDock Tools |
| [53,61,65,122,124] |
| AutoGrid | To generate affinity maps and spacing | [53] |
| DockingServer | To perform docking calculation | [53,61,124] |
| PyMol | To visually analyze results from AutoDock Vina | [65] |
| PLANTS | To simulate docking | [126] |
| Discovery Studio (Accelrys Software) |
| [25,55,57,61,122,124] |
| Discovery Studio Client (Dassault Systèmes Biovia Corp ®) |
| [122] |
| Protein Source | Most Important Peptides | Protein Hydrolysate Fraction | Peptide Sequence | Inhibitory Activities | Authors | |||
|---|---|---|---|---|---|---|---|---|
| α-Amylase | α-Glucosidase | DPP-IV | Others | |||||
| Soybean (Glycine max) | Potential α-amylase, α-glucosidase, DPP-IV inhibitor peptides: subfractions F1, F2, and F3 collected by RP-HPLC from 5–10 kDa fraction obtained from 6-day germinated soybean protein digest | 5–10 kDa, F1 | NNDDRDS, VVNPDNNEN, LSSTEAQQS, NAENNQRN, IKSQSES, EEPQQPQQ, GQSSRPQD, LAGNQEQE, NLKSQQA, QEPQESQQ, SQRPQDRHQ, QQQQQGGSQSQ, QQQQQGGSQSQKG, PETMQQQQQQ, SDESTESETEQA | 85% | Maltase: 28% Sucrase: 22% | IC50: 0.8 mg/mL | NR | [16] |
| 5–10 kDa, F2 | RNLQGENEEEDSGA, VTRGQGKV, KKGVIT, IMSDESTESETEQA | 20% | Maltase: 21% Sucrase: 21% | IC50: 0.75 mg/mL | NR | |||
| 5–10 kDa, F3 | NALKPDNRIESEGG, SSPDIYNPQAGSVT, RQNIGQNSSPDIYNPQAG, VVAEQAGEQGFE HKNKNPF | 5% | Maltase: 30% Sucrase: 8% | IC50: 0.6 mg/mL | NR | |||
| Soybean | Potential α-glucosidase inhibitor peptides: GSR EAK | Hydrolysate <5 kDa separated by ion exchange chromatography, Fraction C-III isolated and purified by Sephadex G-25, Fraction C-III-2 separated by RP-HPLC, Fraction C-III-2a collected. | GSR EAK | NR | IC50: 20.4 μM IC50: 520.2 μM α-glucosidase (5NN8): - GSR and EAK bind differently from Acarbose; they bind close to active site, mainly through van der Waals contacts, anion-π interactions, and hydrogen bonds. | NR | NR | [65] |
| Soybean | Most frequently occurring peptide in soybean proteins with DPP-IV inhibitor activity: GA, GP, and PG | NR | GA, GP, PG | NR | NR | Most frequently occurring peptides in soybean proteins having DPP-IV inhibitor activity | NR | [123] |
| Soy and Lupin | Potential DPP-IV inhibitor peptides: Soy 1: IAVPTGVA Lup 1: LTFPGSAED | NA (Peptides were synthesized) | Soy 1: IAVPTGVA Lup 1: LTFPGSAED | NR | NR | IC50 106 μM 228 μM DPP-IV (4PNZ): Soy 1 N-terminus and C-terminus matched the binding of omarigliptin. Other interactions include salt bridge, ionic network, π-π stacking, electrostatic interaction, and ionic interactions. | NR | [126] |
| Common bean (Phaseolus vulgaris) | Potential DPP-IV inhibitor peptides detected in unfractionated protein hydrolysate: LAPPG, KLLLRRLQ, REYLLVAQ, LRENNKLMLLELK, RLLLKLRQ | Unfractionated protein hydrolysate | LAPPG, KLLLRRLQ, REYLLVAQ, LRENNKLMLLELK, RLLLKLRQ | NR | NR | Potential activity predicted by BIOPEP database. | NR | [59] |
| Common bean (Phaseolus vulgaris L.) | Potential DPP-IV and α-glucosidase inhibitor peptides: KTYGL, KKSSG, CPGNK, and GGGLHK | Unfractionated protein hydrolysate | KTYGL KKSSG CPGNK GGGLHK | NR | % inh/mg 36.30 ± 8.80 49.34 ± 6.50 37.60 ± 6.80 46.10 ± 8.30 α-glucosidase (3AJ7): binds outside the active site, mainly through polar interactions, hydrophobic.interactions, and hydrogen bonds. | IC50 (mg/mL) 0.03 ± 0.00 0.64 ± 0.16 0.87 ± 0.02 0.61 ± 0.10 Diprotin A: 0.02 ± 0.00 DPP-IV (1RWQ): bind to catalytic site, mainly through hydrogen, hydrophobic, polar, and cation π bonds. | NR | [53] |
| Black bean (Black-Otomi) | Potential GLUT2 and SGLT1 inhibitor peptides: AKSPLF, ATNPLF, FEELN, and LSKSVL | Unfractionated protein hydrolysate | AKSPLF, ATNPLF, FEELN, LSKSVL | NR | NR | NR | GLUT2 (P12336): binds outside catalytic site, mainly through hydrophobic, polar, cation-π, π-π interactions. SGLT1 (3DH4): binds outside catalytic site, mainly through polar and hydrophobic interactions. | [124] |
| Black bean | Potential SGLT1, GLUT2, PKC, AMPK inhibitor peptides: AKSPLF, ATNPLF, FEELN, and LSVSVL | Unfractionated protein hydrolysate | AKSPLF, ATNPLF, FEELN, LSVSVL | NR | NR | NR | GLUT2 (P12336): most potent FEELN. SGLT1 (2XQ2): most potent ATNPLF. PKC (4RA5): most potent ATNPLF, binds to catalytic site through hydrogen bond. AMPK (4QFG): most potent ATNPLF, binds to catalytic site through hydrogen bond. | [122] |
| Common bean (Phaseolus vulgaris L.) | Potential DPP-IV inhibitor peptides: SGAM, DSSG, LLAH, YVAT, EPTE and KPKL | Unfractionated protein hydrolysate | SGAM, DSSG, LLAH, YVAT, EPTE, KPKL | NR | NR | Potential activity predicted by BIOPEP database | NR | [38] |
| Black bean (Phaseolus vulgaris L.) | Potential α-amylase, α-glucosidase, DPP-IV inhibitor peptides: AKSPLF, QTPF, FEELN, LSKSVL, and EGLELLLLLLAG | Unfractionated protein hydrolysate | AKSPLF, QTPF, FEELN, LSKSVL, EGLELLLLLLAG | α-amylase (1B2Y): Good potential AKSPLF, FEELN, QTPF, LSKSVL, interacted with the catalytic site (TYR151, HIS201, ILE235), mainly through hydrophobic interactions, polar interactions, and hydrogen bonds. | α-glucosidase (3AJ7): Good potential AKSPLF, FEELN, QTPF, and LSKSVL, interacted with the catalytic site (ASP34, THR83, and ASN32), mainly through hydrogen bonds and polar interactions; only one hydrophobic interaction. | DPP-IV (3W2T): Highest inhibition potential EGLELLLLLLAG, AKSPLF, FEELN, interacted with the catalytic site (ASP192, GLU191, ARG253), mainly through hydrogen bonds, electrostatic or polar interactions, and hydrophobic interactions. | NR | [61] |
| Pinto bean (Phaseolus vulgaris cv. Pinto) | Potential α-amylase inhibitor peptides: PPHMLP, PPMHLP, PLPWGAGF, GDAACCGLPLLP, PPHMGGP, PLPPHDLL, and FNPFPSPHTP | <3 kDa | PPHMLP, PPMHLP, PLPWGAGF, GDAACCGLPLLP, PPHMGGP, PLPPHDLL, FNPFPSPHTP | Peptide sequence detected in protein hydrolysate fraction that has the highest α-amylase inhibitor activity. | NR | NR | NR | [42] |
| Pinto bean (Phaseolus vulgaris cv. Pinto) | Novel potential dual functional (α-amylase and angiotensin converting enzyme) inhibitory peptides: PBp1: PPHMLP PBp2: PLPWGAGF PBp3: PPHMGGP PBp4: PLPLHMLP PBp5: LSSLEMGSLGALFVCM | NA (Peptides were synthesized) | PBp1: PPHMLP PBp2: PLPWGAGF PBp3: PPHMGGP PBp4: PLPLHMLP PBp5: LSSLEMGSLGALFVCM | IC50 (mM) 23.33 ± 0.15 15.73 ± 0.06 19.83 ± 0.12 15.80 ± 0.17 10.03 ± 0.47 | NR | NR | NR | [62] |
| Hard-to-cook bean (Phaseolus vulgaris L.) | Potential α-amylase and DPP-IV inhibitor peptides: FFL, LLSL, QQEG, and NEGEAH | Unfractionated hydrolysate and <1 kDa | FFL, LLSL, QQEG, NEGEAH | α-amylase (1HNY): potential peptides FFL and NEGEAH interacted with the active site (ASP197, GLU233, and ASP300), mainly through van der Waals contacts, hydrogen bonds, electrostatic, charged, and π interactions. | NR | DPP-IV (1X70): Potential peptides LLSL and QQEG, interacted with S1 and S2 pockets of three pockets of active site, mainly through van der Waals contacts, hydrogen bonds, electrostatic, charged, and π interactions. | NR | [25] |
| Cowpea bean (Vigna unguiculata) | Potential DPP-IV inhibitors: KVSVVAL and TTAGLLE | Unfractionated protein hydrolysate | KVSVVAL, TTAGLLE | NR | NR | DPP-IV (1X70): most potent KVSVVAL, low total energy score and several interactions with catalytic region. TTAGLLE binds with active site at S2 and S3 pockets, mainly through van der Waals and electrostatic interactions. | NR | [57] |
| Common bean (Phaseolus vulgaris) | Potential DPP-IV inhibitor peptide: RGPLVNPDPKPFL | Unfractionated protein hydrolysate | RGPLVNPDPKPFL | NR | NR | DPP-IV (1X70): Dock around S3 pocket, mainly through van der Waals and electrostatic interactions. | NR | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahmi, A.; Arcot, J. In Vitro Assessment Methods for Antidiabetic Peptides from Legumes: A Review. Foods 2023, 12, 631. https://doi.org/10.3390/foods12030631
Rahmi A, Arcot J. In Vitro Assessment Methods for Antidiabetic Peptides from Legumes: A Review. Foods. 2023; 12(3):631. https://doi.org/10.3390/foods12030631
Chicago/Turabian StyleRahmi, Alia, and Jayashree Arcot. 2023. "In Vitro Assessment Methods for Antidiabetic Peptides from Legumes: A Review" Foods 12, no. 3: 631. https://doi.org/10.3390/foods12030631