

Fresh/High-Zinc Maize: A Promising Solution for Alleviating Zinc Deficiency through Significant Micronutrient Accumulation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Iron and Zinc Concentration in Maize Kernels
2.3. Estimated Average Requirement
2.4. Statistical Analysis
3. Results
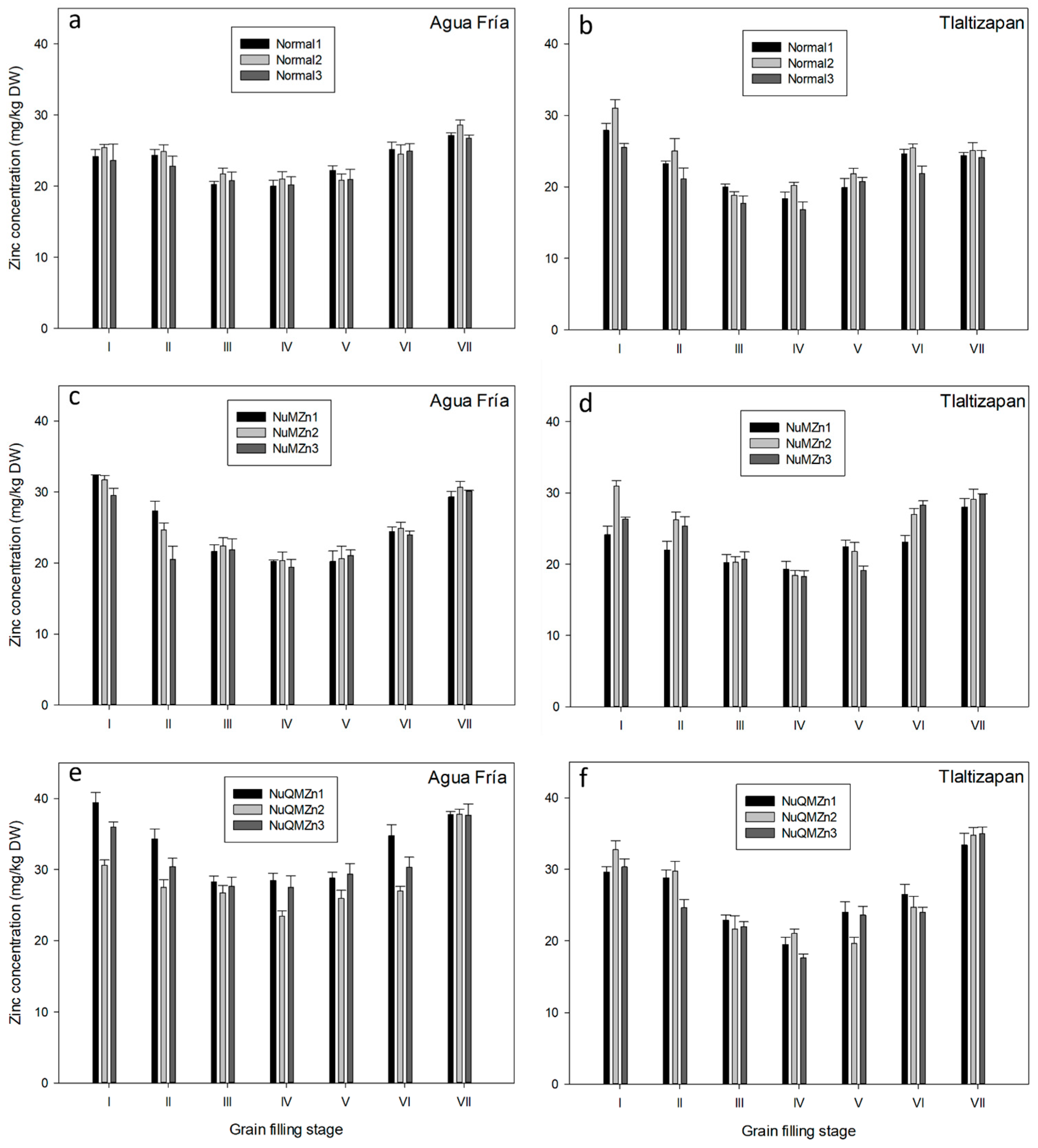
3.1. Zinc Accumulation during Kernel Development
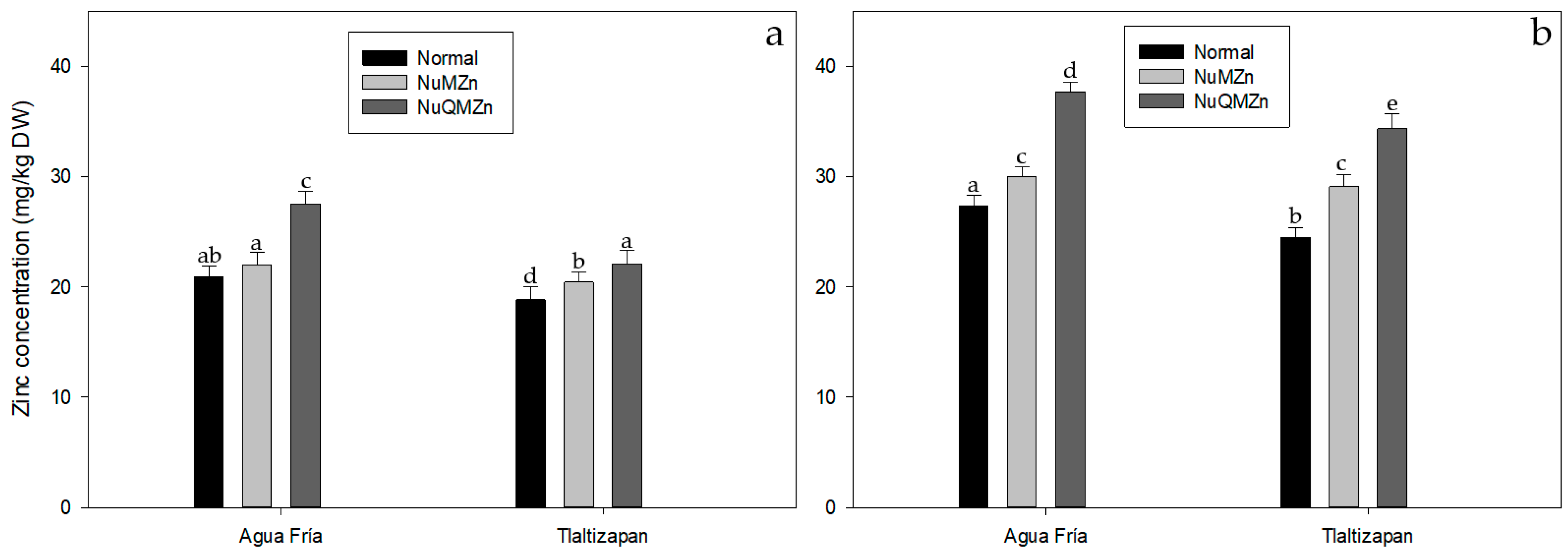
3.2. Zinc Content at Stage III (Fresh Maize) and Stage VII (Harvest Maturity)
3.3. Iron Content in Stage III and VII
3.4. Contribution of Fresh High-Zinc Maize to the Estimated Average Requirement (EAR) of Zinc and Iron
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Genotype | Iron in Fresh Maize (mg kg−1 DW) | Pre-School Children (1–3 Years Old) | Children (4–8 Years Old) | Children (9–13 Years Old) | Pregnant Women (19–50 Years Old) |
|---|---|---|---|---|---|
| % EAR | |||||
| Normal1-AF | 12.77 | 29.81 | 43.62 | 60.62 | 15.55 |
| Normal2-AF | 13.01 | 30.36 | 44.42 | 61.74 | 15.84 |
| Normal3-AF | 13.52 | 31.54 | 46.16 | 64.16 | 16.46 |
| NuMZn1-TL | 11.87 | 27.69 | 40.52 | 56.32 | 14.45 |
| NuMZn2-TL | 11.37 | 26.54 | 38.84 | 53.98 | 13.85 |
| NuMZn3-TL | 11.92 | 27.82 | 40.70 | 56.57 | 14.51 |
| NuMZn1-AF | 14.26 | 33.28 | 48.71 | 67.69 | 17.36 |
| NuMZn2-AF | 13.94 | 32.53 | 47.60 | 66.15 | 16.97 |
| NuMZn3-AF | 13.92 | 32.48 | 47.54 | 66.07 | 16.95 |
| Normal1-TL | 14.14 | 33.00 | 48.29 | 67.12 | 17.22 |
| Normal2-TL | 13.76 | 32.10 | 46.98 | 65.29 | 16.75 |
| Normal3-TL | 13.92 | 32.47 | 47.52 | 66.04 | 16.94 |
| NuQMZn1-AF | 13.17 | 30.73 | 44.97 | 62.51 | 16.03 |
| NuQMZn2-AF | 12.61 | 29.43 | 43.07 | 59.86 | 15.36 |
| NuQMZn3-AF | 12.72 | 29.68 | 43.44 | 60.37 | 15.49 |
| NuQMZn1-TL | 12.40 | 28.93 | 42.33 | 58.84 | 15.09 |
| NuQMZn2-TL | 12.88 | 30.06 | 43.99 | 61.13 | 15.68 |
| NuQMZn3-TL | 12.12 | 28.27 | 41.37 | 57.50 | 14.75 |
References
- Welch, R.M. The Impact of Mineral Nutrients in Food Crops on Global Human Health. Plant Soil 2002, 247, 83–90. [Google Scholar] [CrossRef]
- Bulletin, N. Assessment of the Risk of Zinc Deficiency in Populations. Food Nutr. Bull. 2004, 25, S130–S162. [Google Scholar] [CrossRef]
- Xue, Y.F.; Yue, S.C.; Liu, D.Y.; Zhang, W.; Chen, X.P.; Zou, C.Q. Dynamic Zinc Accumulation and Contributions of Pre- and/or Post-Silking Zinc Uptake to Grain Zinc of Maize as Affected by Nitrogen Supply. Front. Plant Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Golabi, S.; Adelipour, M.; Mobarak, S.; Piri, M.; Seyedtabib, M.; Bagheri, R.; Suzuki, K.; Ashtary-Larky, D.; Maghsoudi, F.; Naghashpour, M. The Association between Vitamin d and Zinc Status and the Progression of Clinical Symptoms among Outpatients Infected with SARS-CoV-2 and Potentially Non-Infected Participants: A Cross-Sectional Study. Nutrients 2021, 13, 3368. [Google Scholar] [CrossRef] [PubMed]
- Ekpa, O.; Palacios-Rojas, N.; Kruseman, G.; Fogliano, V.; Linnemann, A.R. Sub-Saharan African Maize-Based Foods: Technological Perspectives to Increase the Food and Nutrition Security Impacts of Maize Breeding Programmes. Glob. Food Sec. 2018, 17, 48–56. [Google Scholar] [CrossRef]
- Chomba, E.; Westcott, C.M.; Westcott, J.E.; Mpabalwani, E.M.; Krebs, N.F.; Patinkin, Z.W.; Palacios, N.; Hambidge, K.M. Zinc Absorption from Biofortified Maize Meets the Requirements of Young Rural Zambian Children. J. Nutr. 2015, 145, 514–519. [Google Scholar] [CrossRef] [Green Version]
- Bouis, H.E.; Welch, R.M. Biofortification—A Sustainable Agricultural Strategy for Reducing Micronutrient Malnutrition in the Global South. Crop Sci. 2010, 50, S-20–S-32. [Google Scholar] [CrossRef] [Green Version]
- Welch, R.M.; Graham, R.D. Breeding for Micronutrients in Staple Food Crops from a Human Nutrition Perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Woods, B.J.; Gallego-Castillo, S.; Talsma, E.F.; Álvarez, D. The Acceptance of Zinc Biofortified Rice in Latin America: A Consumer Sensory Study and Grain Quality Characterization. PLoS ONE 2020, 15, e0242202. [Google Scholar] [CrossRef]
- Ramírez-Jaspeado, R.; Palacios-Rojas, N.; Funes, J.; Pérez, S.; Donnet, M.L. Identification of Potential Areas in Mexico for Intervention with Biofortified High-Zinc Maize. Rev. Fitotec. Mex. 2018, 41, 327–337. [Google Scholar]
- Palacios-Rojas, N.; McCulley, L.; Kaeppler, M.; Titcomb, T.J.; Gunaratna, N.S.; Lopez-Ridaura, S.; Tanumihardjo, S.A. Mining Maize Diversity and Improving Its Nutritional Aspects within Agro-Food Systems. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1809–1834. [Google Scholar] [CrossRef] [PubMed]
- Virk, P.S.; Andersson, M.S.; Arcos, J.; Govindaraj, M.; Pfeiffer, W.H. Transition From Targeted Breeding to Mainstreaming of Biofortification Traits in Crop Improvement Programs. Front. Plant Sci. 2021, 12, 703990. [Google Scholar] [CrossRef] [PubMed]
- Hindu, V.; Palacios-Rojas, N.; Babu, R.; Suwarno, W.B.; Rashid, Z.; Usha, R.; Saykhedkar, G.R.; Nair, S.K. Identification and Validation of Genomic Regions Influencing Kernel Zinc and Iron in Maize. Theor. Appl. Genet. 2018, 131, 1443–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasanna, B.M.; Palacios-Rojas, N.; Hossain, F.; Muthusamy, V.; Menkir, A.; Dhliwayo, T.; Ndhlela, T.; San Vicente, F.; Nair, S.K.; Vivek, B.S. Molecular Breeding for Nutritionally Enriched Maize: Status and Prospects. Front. Genet. 2020, 10, 1392. [Google Scholar] [CrossRef] [Green Version]
- Harvest Plus Biofortified Crops around the World. Available online: https://www.harvestplus.org/home/global-reach/#map (accessed on 14 June 2023).
- Cheah, Z.X.; Kopittke, P.M.; Harper, S.M.; O’hare, T.J.; Wang, P.; Paterson, D.J.; De Jonge, M.D.; Bell, M.J. In Situ Analyses of Inorganic Nutrient Distribution in Sweetcorn and Maize Kernels Using Synchrotron-Based x-Ray Fluorescence Microscopy. Ann. Bot. 2019, 123, 543–556. [Google Scholar] [CrossRef]
- Gallego-Castillo, S.; Taleon, V.; Talsma, E.F.; Rosales-Nolasco, A.; Palacios-Rojas, N. Effect of Maize Processing Methods on the Retention of Minerals, Phytic Acid and Amino Acids When Using High Kernel-Zinc Maize. Curr. Res. Food Sci. 2021, 4, 279–286. [Google Scholar] [CrossRef]
- Chakraborti, M.; Prasanna, B.M.; Hossain, F.; Mazumdar, S.; Singh, A.M.; Guleria, S.; Gupta, H.S. Identification of Kernel Iron- and Zinc-Rich Maize Inbreds and Analysis of Genetic Diversity Using Microsatellite Markers. J. Plant Biochem. Biotechnol. 2011, 20, 224–233. [Google Scholar] [CrossRef]
- Keigler, J.I.; Wiesinger, J.A.; Flint-Garcia, S.A.; Glahn, R.P. Iron Bioavailability of Maize (Zea mays L.) after Removing the Germ Fraction. Front. Plant Sci. 2023, 14, 1114760. [Google Scholar] [CrossRef]
- Maqbool, M.A.; Beshir, A.R. Zinc Biofortification of Maize (Zea mays L.): Status and Challenges. Plant Breed. 2019, 138, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Cabrera-Soto, L.; Pixley, K.V.; Rosales-Nolasco, A.; Galicia-Flores, L.A.; Palacios-Rojas, N. Carotenoid and Tocochromanol Profiles during Kernel Development Make Consumption of Biofortified “Fresh” Maize an Option to Improve Micronutrient Nutrition. J. Agric. Food Chem. 2018, 66, 9391–9398. [Google Scholar] [CrossRef]
- Osorio-Saenz, A.; Santacruz-Varela, A.; Córdova-Téllez, L.; González-Hernández, V.A.; Mascorro-Gallardo, J.O.; Conde-Martínez, F.V.; Carrillo-Castañeda, G. Mexican Maize Landraces for Corn on the Cob Production at the Central Highlands. Maydica 2019, 64, 1–8. [Google Scholar]
- Baseggio, M.; Murray, M.; Wu, D.; Ziegler, G.; Kaczmar, N.; Chamness, J.; Hamilton, J.P.; Robin Buell, C.; Vatamaniuk, O.K.; Buckler, E.S.; et al. Genome-Wide Association Study Suggests an Independent Genetic Basis of Zinc and Cadmium Concentrations in Fresh Sweet Corn Kernels. G3 Genes Genomes Genet. 2021, 11, jkab186. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, J.S.; Min, W.H.; Zheng, M.Z.; Li, H. Changes of Moisture Distribution and Migration in Fresh Ear Corn during Storage. J. Integr. Agric. 2019, 18, 2644–2651. [Google Scholar] [CrossRef]
- Queiroz, O.C.M.; Arriola, K.G.; Daniel, J.L.P.; Adesogan, A.T. Effects of 8 Chemical and Bacterial Additives on the Quality of Corn Silage. J. Dairy Sci. 2013, 96, 5836–5843. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.R. Grain Fill Stages in Corn. Available online: http://www.king.corn/news/timeless/GrainFill.html (accessed on 22 April 2022).
- Palacios-Rojas, N. Calidad Nutricional e Industrial de Maíz. Protocolos; CIMMYT: Texcoco, Mexico, 2018; ISBN 9786078263783. [Google Scholar]
- Institute of Medicine (US), Panel on Micronutrients. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Noulas, C.; Tziouvalekas, M.; Karyotis, T. Zinc in Soils, Water and Food Crops. J. Trace Elem. Med. Biol. 2018, 49, 252–260. [Google Scholar] [CrossRef]
- De Groote, H.; Tessema, M.; Gameda, S.; Gunaratna, N.S. Soil Zinc, Serum Zinc, and the Potential for Agronomic Biofortification to Reduce Human Zinc Deficiency in Ethiopia. Sci. Rep. 2021, 11, 8770. [Google Scholar] [CrossRef]
- Liu, D.Y.; Zhang, W.; Liu, Y.M.; Chen, X.P.; Zou, C.Q. Soil Application of Zinc Fertilizer Increases Maize Yield by Enhancing the Kernel Number and Kernel Weight of Inferior Grains. Front. Plant Sci. 2020, 11, 188. [Google Scholar] [CrossRef]
- Mageto, E.K.; Lee, M.; Dhliwayo, T.; Palacios-Rojas, N.; Vicente, F.S.; Burgueño, J.; Hallauer, A.R. An Evaluation of Kernel Zinc in Hybrids of Elite Quality Protein Maize (QPM) and Non-QPM Inbred Lines Adapted to the Tropics Based on a Mating Design. Agronomy 2020, 10, 695. [Google Scholar] [CrossRef]
- Cheah, Z.X.; Kopittke, P.M.; Scheckel, K.G.; Noerpel, M.R.; Bell, M.J. Comparison of Zn Accumulation and Speciation in Kernels of Sweetcorn and Maize Differing in Maturity. Ann. Bot. 2020, 125, 185–193. [Google Scholar] [CrossRef]
- Glahn, R.; Tako, E.; Gore, M.A. The Germ Fraction Inhibits Iron Bioavailability of Maize: Identification of an Approach to Enhance Maize Nutritional Quality via Processing and Breeding. Nutrients 2019, 11, 833. [Google Scholar] [CrossRef] [Green Version]
- Milani, P.; Torres-Aguilar, P.; Hamaker, B.; Manary, M.; Abushamma, S.; Laar, A.; Steiner, R.; Ehsani, M.; de la Parra, J.; Skaven-Ruben, D.; et al. The Whole Grain Manifesto: From Green Revolution to Grain Evolution. Glob. Food Sec. 2022, 34, 100649. [Google Scholar] [CrossRef]


| Trait | AF | TL |
|---|---|---|
| pH | 8.05 | 8.23 |
| Bulk density (g cm3) | 1.18 | 1.07 |
| Organic matter concentration (%) | 3.60 | 2.25 |
| p-Olsen (mg kg−1) | 28.85 | 25.48 |
| Salinity (dS m−1) | 0.84 | 0.98 |
| Total carbonates (%) | 7.07 | 28.98 |
| K (mg kg−1) | 398.25 | 414.17 |
| Ca (mg kg−1) | 6437.25 | 5155.67 |
| Mg (mg kg−1) | 251.00 | 629.17 |
| Na (mg kg−1) | 35.13 | 36.27 |
| Fe (mg kg−1) | 18.00 | 3.21 |
| Zn (mg kg−1) | 0.84 | 0.76 |
| Mn (mg kg−1) | 2.98 | 3.97 |
| Cu (mg kg−1) | 0.98 | 0.83 |
| B (mg kg−1) | 0.85 | 0.71 |
| S (mg kg−1) | 5.39 | 13.91 |
| N-NO3 (mg kg−1) | 30.08 | 13.62 |
| Genotype | Zinc in Fresh Corn (mg kg−1 DW) | Pre-School Children (1–3 Years Old) | Children (4–8 Years Old) | Children (9–13 Years Old) | Pregnant Women (19–50 Years Old) |
|---|---|---|---|---|---|
| % EAR a | |||||
| Normal1-AF | 20.23 | 56.63 | 70.79 | 80.91 | 56.63 |
| Normal2-AF | 21.72 | 60.80 | 76.00 | 86.86 | 60.80 |
| Normal3-AF | 20.75 | 58.09 | 72.61 | 82.98 | 58.09 |
| Normal1-TL | 19.96 | 55.90 | 69.88 | 79.86 | 55.90 |
| Normal2-TL | 18.86 | 52.79 | 65.99 | 75.42 | 52.79 |
| Normal3-TL | 17.67 | 49.48 | 61.85 | 70.68 | 49.48 |
| NuMZn1-AF | 21.64 | 60.58 | 75.73 | 86.55 | 60.58 |
| NuMZn2-AF | 22.42 | 62.78 | 78.47 | 89.68 | 62.78 |
| NuMZn3-AF | 21.87 | 61.24 | 76.55 | 87.49 | 61.24 |
| NuMZn1-TL | 20.23 | 56.65 | 70.81 | 80.92 | 56.65 |
| NuMZn2-TL | 20.28 | 56.78 | 70.97 | 81.11 | 56.78 |
| NuMZn3-TL | 20.72 | 58.01 | 72.52 | 82.88 | 58.01 |
| NuQMZn1-AF | 28.23 | 79.04 | 98.80 | 112.91 | 79.04 |
| NuQMZn2-AF | 26.73 | 74.85 | 93.56 | 106.92 | 74.85 |
| NuQMZn3-AF | 27.63 | 77.37 | 96.71 | 110.53 | 77.37 |
| NuQMZn1-TL | 22.88 | 64.06 | 80.08 | 91.51 | 64.06 |
| NuQMZn2-TL | 21.65 | 60.63 | 75.79 | 86.61 | 60.63 |
| NuQMZn3-TL | 21.97 | 61.52 | 76.90 | 87.88 | 61.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosales, A.; Molina-Macedo, A.; Leyva, M.; San Vicente, F.; Palacios-Rojas, N. Fresh/High-Zinc Maize: A Promising Solution for Alleviating Zinc Deficiency through Significant Micronutrient Accumulation. Foods 2023, 12, 2757. https://doi.org/10.3390/foods12142757
Rosales A, Molina-Macedo A, Leyva M, San Vicente F, Palacios-Rojas N. Fresh/High-Zinc Maize: A Promising Solution for Alleviating Zinc Deficiency through Significant Micronutrient Accumulation. Foods. 2023; 12(14):2757. https://doi.org/10.3390/foods12142757
Chicago/Turabian StyleRosales, Aldo, Aide Molina-Macedo, Mayolo Leyva, Félix San Vicente, and Natalia Palacios-Rojas. 2023. "Fresh/High-Zinc Maize: A Promising Solution for Alleviating Zinc Deficiency through Significant Micronutrient Accumulation" Foods 12, no. 14: 2757. https://doi.org/10.3390/foods12142757