1. Introduction
As a probiotic, lactic acid bacteria (LAB) are extensively found in nature [
1,
2]. People in many regions consume foods containing LAB [
3], and LAB have been proven to have important roles in human health. However, in recent years, postbiotics have been widely studied since they are safer and easier to commercialize than probiotics [
4,
5]. Postbiotics have many functions, such as enhancing the epithelial barrier function [
6,
7], favorable modulation of the gut microbiota [
8], immune responses [
9,
10], and systemic metabolism [
11]. Moreover, studies on postbiotics focus on cell-free supernatants, exopolysaccharides (EPS), enzymes, cell wall fragments, and bacterial lysates.
EPS is a long-chain, high molecular weight polysaccharide with diverse structures and physicochemical properties [
12]. These polysaccharides are secondary metabolites produced by different microorganisms, such as Lactobacillus and Bacillus, and are secreted to the extracellular space [
13,
14]. EPS has many advantages when compared with other postbiotics: a. cost-effectiveness, on the one hand, the medium required to produce EPS is cheap, and on the other hand, the extraction process of EPS is not complicated because it is secreted to the extracellular space; b. the abundance of species, the structure of EPS depends on the kinds of microorganisms and the composition of the culture medium [
15]. In theory, a strain in different culture media could produce EPSs with different structures. This provides a quantitative basis for screening EPS with advantageous functions. For example, the hydrophobic group attached to the hydrophilic chain of EPS makes it amphiphilic and able to act as a natural emulsifier. The addition of EPS to low-fat mayonnaise could improve the rheological properties and stability of the product [
16], and the emulsification activity of some EPSs was comparable to that of xanthan gum [
17]. In addition, EPS could be used as a potential antioxidant in food processing because of its excellent antioxidant capacity [
18]. Therefore, these properties of EPS have great potential to be used in food additives and functional food raw materials.
However, there are two problems before we further exploit EPS. The first is the yield of EPS. EPS production is influenced by numerous factors, such as the species of the microorganisms, the medium components (e.g., carbon source, nitrogen source), and different nutrients preferred by different microorganisms for EPS production [
13,
19]. In addition, the culture conditions (e.g., temperature, pH) also influence the production of EPS [
20,
21]. Moreover, the optimum conditions for EPS production are not always consistent with the growth of microorganisms. Even some EPS syntheses are performed by microorganisms only under stress conditions [
22]. Whereas the optimal temperature for
Lactobacillus paracasei growth is 37 °C, the temperature suitable for EPS production is 20 °C [
20]. Therefore, systematic optimization of these factors is crucial to maximizing the EPS yield. The other problem is the biological activity of the EPS. The yield of EPS determines whether it is industrially viable, while the function determines the direction of research and application. Due to the diversity of EPS structures, the functions of EPS are also extensive, such as antioxidant, anti-inflammatory, emulsification activity, antimicrobial, anti-tumor, immunomodulatory, and cholesterol-lowering activities [
23,
24]. Therefore, uncovering the potential biological activities of EPS is crucial for further study and its application in the food industry.
This study also addresses both of these problems. The first part was to optimize the level of EPS production by L. plantarum R301, which was preserved in our laboratory. Single-factor optimization and response surface optimization were used for optimizing media composition and culture conditions to obtain the highest EPS yield while investigating the effect of different nutrients and culture conditions on EPS production. The other part is to investigate the potential biological activity of EPS for its subsequent research and application. Specifically, this work reported the antioxidant activity of the EPS in vitro, including DPPH, ABTS, hydroxyl radical-scavenging capacity, and reducing power. Moreover, the anti-inflammatory activity of EPS was investigated using an LPS-induced RAW 264.7 inflammation model.
4. Discussion
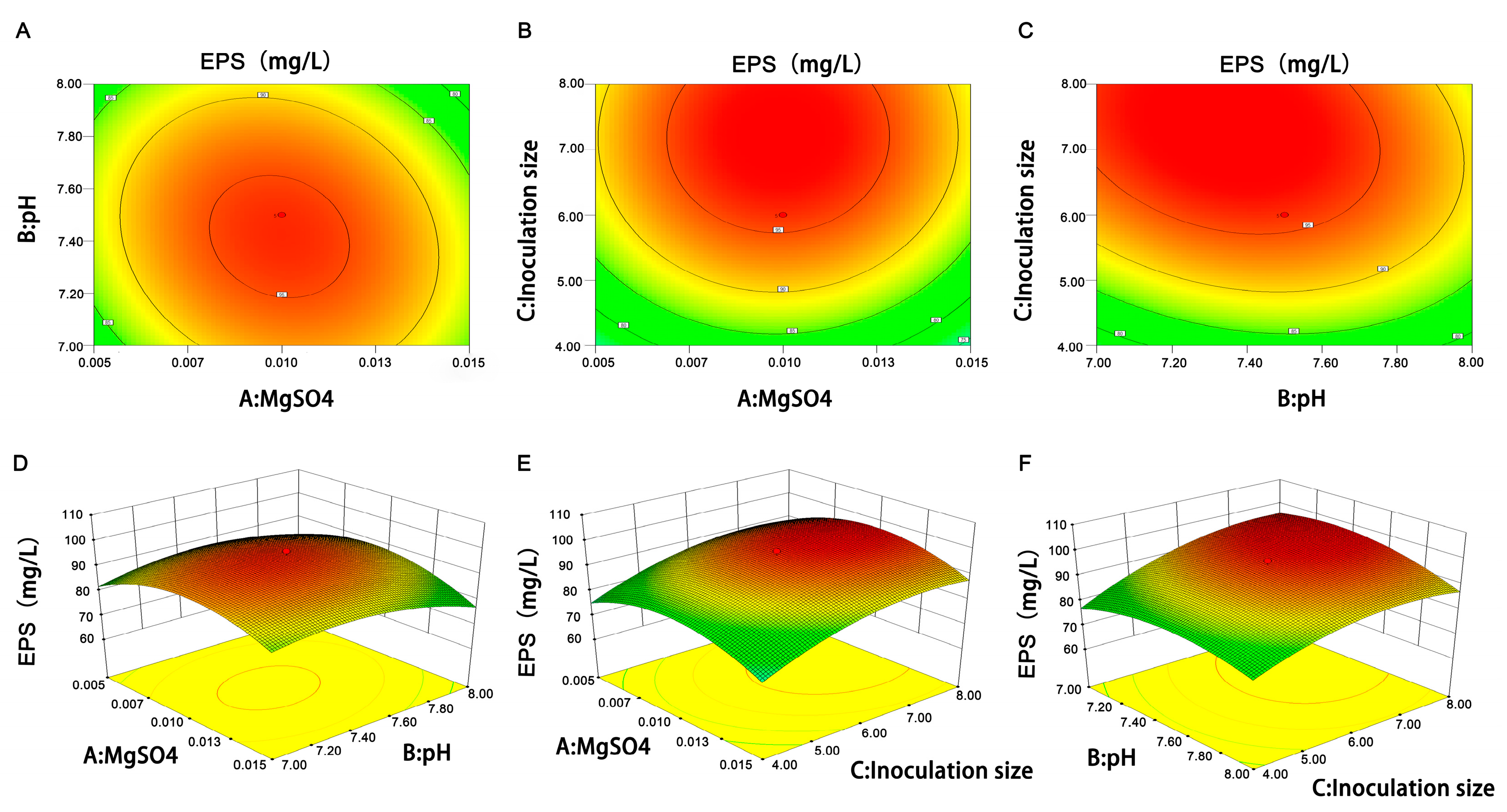
EPS production is affected by many factors, such as genetics, the culture medium, and fermentation conditions. The genetics factor mostly depends on the strain types, such as
Pseudoalteromonas agarivorans (2783.6 mg/L) [
28],
Lactobacillus rhamnosus (210.28 mg/L) [
29], and
L. plantarum (280.105 mg/L) [
30], while the culture medium and fermentation conditions need to be optimized for the specific strain. The present study optimized EPS production by
L. plantarum R301 using a single-factor experiment and RSM. The single-factor experiment preliminarily obtained the conditions for EPS production: 4% glucose, 2% small peptide, 0.06% MnSO
4, 0.02% MgSO
4, fermentation temperature of 27 °C, 3% inoculum size, initial pH of 7.0, and the fermentation time of 24 h (
Figure 1). The RSM experiment further optimized three factors (MgSO
4 concentration, initial pH, and inoculation size) to obtain the maximum EPS yield (
Table 4,
Table 5,
Table 6 and
Table 7). The carbon and nitrogen sources were identified as the primary influential factors in EPS production, and different strains prefer different nutrient sources. This study shows that glucose is the most favorable carbon source for EPS production. However, choosing the optimum nitrogen source was not an easy job, because most of the nitrogen sources contained polysaccharides that could be detected via the phenol–sulfate method, which would contaminate the results of EPS yield optimization. The small peptide that contained a variety of short peptides was processed in our laboratory. It was used as the nitrogen source because it contained fewer polysaccharides and could better promote the growth of
L. plantarum R301. The impact of metal ions on EPS production is mainly reflected by effects on the growth of bacteria. In this study, excessive or undersized MgSO
4 was not suitable for the growth of
L. plantarum R301, which directly led to the reduction of EPS production (
Figure 1F). In addition, the MgSO
4 concentration was the significant factor that affected EPS production in the RSM experiment, which means that it might have some other functions for EPS production. Some metal elements are important for bacterial growth and metabolism [
31]. Whether the MgSO
4 concentration promotes the production of EPS by affecting the metabolism of
L. plantarum R301 needs further study. The optimum fermentation temperature of 27 °C was interesting because the optimum growth temperature of
L. plantarum R301 was 37 °C. This was not the same as most of the relevant studies, which had the same temperature [
21,
29]. One theory is that microorganisms produce EPS to protect themselves under stress [
32,
33]. For this study,
L. plantarum R301 growth restriction may be due to the low temperature as a stressor, thus improving EPS production to protect itself. The research of Bengoa et al. [
20] and Magdalena et al. [
22] also supported our hypothesis in that the optimum temperatures were 20 °C and 25 °C, respectively. Here, we optimized the conditions of EPS production in
L. plantarum R301, and the EPS production increased from 53.34 mg/L to 97.85 mg/L, which increased by 84.70%.
As we discussed, finding EPS with good functional activity is the basis for further studies. In this research, we mainly studied the functions of EPS in two aspects: antioxidant and anti-inflammatory. Reactive oxygen species (ROS) have significant functions in various cellular processes, including apoptosis, cell signaling, genetic regulation, and the transportation of ions [
34]. Although reactive oxygen species (ROS) have important physiological functions, an excess of ROS can cause various harmful effects on the body. These include damaging DNA, promoting the development of cancer, and accelerating cellular degeneration, which can lead to several diseases, such as inflammation, lung injury, and other disorders [
35,
36,
37]. Certain antioxidants, such as butylated hydroxyanisole (BHT) and butylated hydroxytoluene (BHA) have been demonstrated to effectively scavenge excessive free radicals. However, it is important to note that these antioxidants may also have harmful effects on the body [
34,
38]. For this reason, it is necessary to find novel green and safe antioxidants. In this study, we assessed three kinds of free radicals: DPPH, ABTS, and ·OH (
Figure 3A–C). The scavenging rate of EPS for these three free radicals could reach 100% at different concentrations. To some extent, the EPS of
L. plantarum R301 had strong antioxidant activity, because most known EPS does not have such strong scavenging activity. For example, the EPS of
L. plantarum KX041 could reach approximately 80% of the scavenging rate of the three free radicals at a concentration of 8 mg/mL [
21]. At a concentration of 8 mg/mL, the EPS of
Bacillus velezensis SN-1 exhibited a scavenging rate of approximately 60% for the three free radicals [
13]. In addition, the EPS of
L. plantarum R301 also had a certain reducing power (
Figure 3D), which indicated that the EPS had different antioxidant mechanisms [
39]. However, the free radical-scavenging activity can only be used for a preliminary evaluation. In vitro and in vivo experiments do not always produce the same effects [
40]. For instance, antioxidant peptides derived from ovotransferrin, which were identified using the oxygen-radical absorbance capacity assay, were found to lack antioxidant activity in endothelial cells [
41]. For this reason, more comprehensive studies on the antioxidant activity of EPS in vivo are needed in the future.
Lipopolysaccharide (LPS) is the most extensively studied type of pathogen-associated molecular pattern (PAMP). When present, LPS can bind to and activate toll-like receptor-4 (TLR-4), which is a type of pattern recognition receptor present on the surface of specific immune cells, such as macrophages, neutrophils, and dendritic cells. The binding of LPS to TLR-4 triggers a signaling pathway that activates multiple transcription factors and ultimately induces an inflammatory response. The immune response triggered by this process involves the synthesis of pro-inflammatory cytokines, which play a vital role in the immune system’s capacity to combat invading pathogens. This inflammatory response includes the production of pro-inflammatory cytokines, which play a crucial role in the immune system’s ability to eliminate invading pathogens [
42]. However, excessive inflammation can disturb the immunomodulatory balance, such as the cytokine storms triggered by an overactive inflammatory response, which can further aggravate the disease and even cause death. Therefore, controlling inflammation is an effective way to prevent further deterioration of the disease. Here, we used the LPS-induced macrophage inflammation model to evaluate the anti-inflammatory activity of EPS. We found that the EPS could decrease production of the pro-inflammatory cytokine Il-6 (
Figure 5E). In addition, the expression of Cox-2, which is known as a major mediator of inflammation, was reduced by EPS treatment (
Figure 5D). These results suggested that the EPS had anti-inflammatory effects, but the exact mechanism was unclear. There are several underlying anti-inflammatory mechanisms of EPS, such as EPS being able to suppress the expression of TLR4 [
27]. The previous study showed that EPS could reduce NO production (
Figure 5A), which was one of the ROS in LPS-induced macrophages. It was easy to relate this result to the excellent antioxidant activity of EPS [
43]. However, whether there is a relationship between antioxidant activity and the anti-inflammatory activity of EPS and how they interact need further study.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}