

Functional Quality Characteristics of the Meat from a Dual-Purpose Poultry Crossbreed Suitable for Backyard Rearing in Comparison to Commercial Broilers

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Birds and Meat Sample Collection
2.3. Analytical Methods
2.3.1. Preparation of Hydrolysate
2.3.2. Estimation of Protein, Antioxidant Activity, and Antiglycation Capacity
2.3.3. Estimation of Carnosine, Anserine, and Creatine
2.3.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.4. Statistical Analyses
3. Results
3.1. Antioxidant Capacity Assays
3.2. The Concentration of Selected Functional Molecules
3.3. Expression of the Carnosine-Related Genes
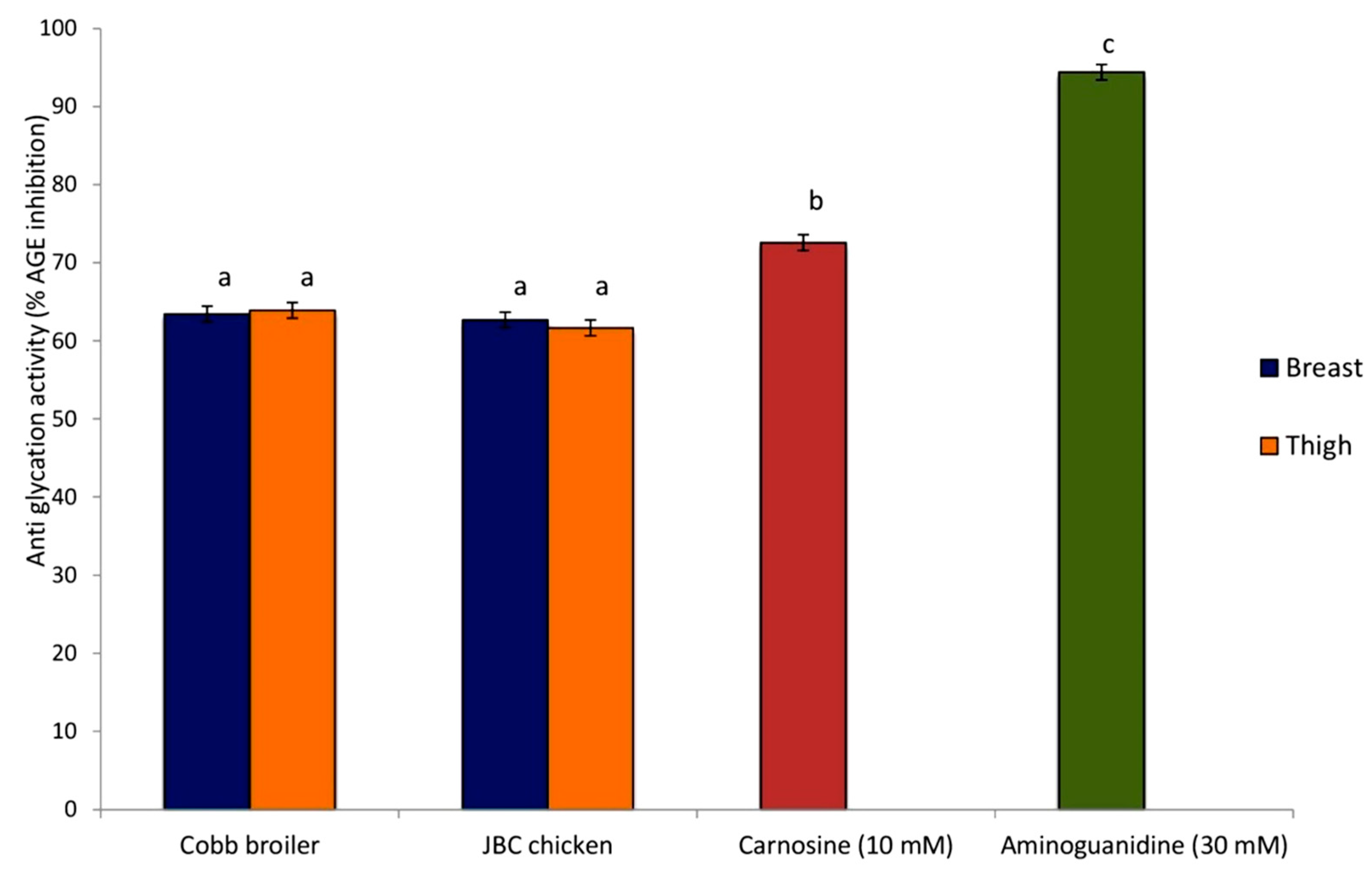
3.4. The Antiglycation Potential of Meat Extract
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pellattiero, E.; Tasoniero, G.; Cullere, M.; Gleeson, E.; Baldan, G.; Contiero, B.; DalleZotte, A. Are meat quality traits and sensory attributes in favor of slow-growing chickens? Animals 2020, 10, 960. [Google Scholar] [CrossRef]
- Marangoni, F.; Corsello, G.; Cricelli, C.; Ferrara, N.; Ghiselli, A.; Lucchin, L.; Poli, A. Role of poultry meat in a balanced diet aimed at maintaining health and wellbeing: An Italian consensus document. Food Nutr. Res. 2015, 9, 27606. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, D.C.F.; de Arruda, A.M.V.; Gonçalves, A.A. Quality characteristics of broiler chicken meat from free-range and industrial poultry system for the consumers. J. Food Sci. Technol. 2017, 54, 1818–1826. [Google Scholar] [CrossRef]
- Singh, M.; Mollier, R.T.; Paton, R.N.; Pongener, N.; Yadav, R.; Singh, V.; Katiyar, R.; Kumar, R.; Sonia, C.; Bhatt, M.; et al. Backyard poultry farming with improved germplasm: Sustainable food production and nutritional security in fragile ecosystem. Front. Sustain. Food Syst. 2022, 6, 962268. [Google Scholar] [CrossRef]
- Samanta, I.; Joardar, S.N.; Das, P.K. Biosecurity strategies for backyard poultry: A controlled way for safe food production. Food Control. Bio. 2018, 16, 481–517. [Google Scholar] [CrossRef]
- Chaiban, C.; Robinson, T.P.; Fevre, E.M.; Ogola, J.; Akoko, J.; Gilbert, M.; Vanwambeke, S.O. Early intensification of backyard poultry systems in the tropics: A case study. Animal 2020, 14, 2387–2396. [Google Scholar] [CrossRef]
- Jayasena, D.D.; Jung, S.; Bae, Y.S.; Kim, S.H.; Lee, S.K.; Lee, J.H.; Jo, C. Changes in endogenous bioactive compounds of Korean native chicken meat at different ages and during cooking. Poult. Sci. 2014, 93, 1842–1849. [Google Scholar] [CrossRef]
- FAO; IFAD. Farmer Field Schools for Family Poultry Producers—A Practical Manual for Facilitators; FAO: Rome, Italy, IFAD: Rome, Italy, 2022. [Google Scholar]
- Rajkumar, U.; Rama Rao, S.V.; Raju, M.V.L.N.; Chatterjee, R.N. Backyard poultry farming for sustained production and enhanced nutritional and livelihood security with special reference to India: A review. Trop. Anim. Health Prod. 2021, 53, 176. [Google Scholar] [CrossRef]
- Padhi, M.K. Importance of indigenous breeds of chicken for rural economy and their improvements for higher production performance. Scientifica 2016, 2016, 2604685. [Google Scholar] [CrossRef] [Green Version]
- Kumaresan, A.; Bujarbaruah, K.M.; Pathak, K.A.; Chhetri, B.; Ahmed, S.K.; Haunshi, S. Analysis of a village chicken production system and performance of improved dual purpose chickens under a subtropical hill agro-ecosystem in India. Trop. Anim. Health Prod. 2008, 40, 395–402. [Google Scholar] [CrossRef]
- Haunshi, S.; Devatkal, S.; Prince, L.L.L.; Ullengala, R.; Ramasamy, K.; Chatterjee, R. Carcass characteristics, meat quality and nutritional composition of Kadaknath, a native chicken breed of India. Foods 2022, 11, 3603. [Google Scholar] [CrossRef]
- Pal, S.; Prakash, B.; Kumar, A.; Singh, Y. Review on backyard poultry farming: Resource utilization for better livelihood of the rural population. Int. J. Curr. Microbiol. Appl. Sci. 2020, 5, 2361–2371. [Google Scholar] [CrossRef]
- Padwar, P.; Thakur, M.S. Effect of melatonin gene on growth and production traits in Kadaknath and Jabalpur colour birds. J. Entomol. Zool. Stud. 2019, 7, 786–788. [Google Scholar]
- Hipkiss, A.R.; Gaunitz, F. Inhibition of tumour cell growth by carnosine: Some possible mechanisms. Amino Acids 2014, 46, 327–337. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Derave, W.; De Courten, B.; Baba, S.P. An update on carnosine and anserine research. Amino Acids 2019, 51, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Serpen, A.; Gökmen, V.; Fogliano, V. Total antioxidant capacities of raw and cooked meats. Meat Sci. 2012, 90, 60–65. [Google Scholar] [CrossRef]
- Abplanalp, W.; Haberzettl, P.; Bhatnagar, A.; Conklin, D.J.; O’Toole, T.E. Carnosine supplementation mitigates the deleterious effects of particulate matter exposure in mice. J. Am. Heart Assoc. 2019, 8, e013041. [Google Scholar] [CrossRef] [Green Version]
- Avgerinos, K.I.; Spyrou, N.; Bougioukas, K.I.; Kapogiannis, D. Effects of creatine supplementation on cognitive function of healthy individuals. Exp. Gerontol. 2018, 108, 166–173. [Google Scholar] [CrossRef]
- Seidel, U.; Huebbe, P.; Rimbach, G. Taurine: A regulator of cellular redox-homeostasis and skeletal muscle function. Mol. Nutr. Food Res. 2019, 63, e1800569. [Google Scholar] [CrossRef]
- Wu, Z.L.; Hou, Y.Q.; Dai, Z.; Hu, C.A. Metabolism, nutrition and redox signaling of hydroxyproline. Antioxid. Redox. Signal. 2019, 30, 674–682. [Google Scholar] [CrossRef]
- Devatkal, S.K.; Vishnuraj, M.R.; Kulkarni, V.V.; Kotaiaha, T. Carcass and meat quality characterization of indigenous and improved variety of chicken genotypes. Poult. Sci. 2018, 97, 2947–2956. [Google Scholar] [CrossRef]
- Sharma, R.; Sehrawat, R.; Ahlawat, S.; Sharma, V.; Parmar, A.; Thakur, M.S.; Mishra, A.K.; Tantia, M.S. An attempt to valorize the only black meat chicken breed of India by delineating superior functional attributes of its meat. Sci. Rep. 2022, 12, 3555. [Google Scholar] [CrossRef]
- Lengkidworraphiphat, P.; Wongpoomchai, R.; Taya, S.; Jaturasitha, S. Effect of genotypes on macronutrients and antioxidant capacity of chicken breast. Asian-Australas. J. Anim. Sci. 2020, 33, 1817–1823. [Google Scholar] [CrossRef]
- Sehrawat, R.; Sharma, R.; Ahlawat, S.; Sharma, V.; Thakur, M.S.; Kaur, M.; Tantia, M.S. First report on better functional property of black chicken meat from India. Indian J. Anim. Res. 2021, 55, 727–733. [Google Scholar] [CrossRef]
- ICAR. Nutrient Requirements of Poultry; Indian Council of Agricultural Research: New Delhi, India, 2013. Available online: http://krishi.icar.gov.in/jspui/handle/123456789/9857 (accessed on 28 March 2023).
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Dinis, T.C.P.; Madeira, V.M.C.; Almeida, L.M. Action of phenolic derivatives (acetoaminophen, salycilate and 5aminosalycilate) as inhibitors of membrane lipid peroxidation and its peroxyl radical scavengers. Arch. Biochem. Biophys 1994, 315, 161–169. [Google Scholar] [CrossRef]
- Abdelkader, H.; Alany, R.G.; Pierscionek, B. Age-related cataract and drug therapy: Opportunities and challenges for topical antioxidant delivery to the lens. J. Pharm. Pharmacol. 2016, 67, 537–550. [Google Scholar] [CrossRef]
- Mora, L.; Sentandreu, M.A.; Toldra, F. Hydrophilic chromatographic determination of carnosine, anserine, balenine, creatine, and creatinine. J. Agric. Food Chem. 2007, 55, 4664–4669. [Google Scholar] [CrossRef]
- Qi, B.; Wang, J.; Ma, Y.B.; Wu, S.G.; Qi, G.H.; Zhang, H.J. Effect of dietary β-alanine supplementation on growth performance, meat quality, carnosine content, and gene expression of carnosine-related enzymes in broilers. Poult. Sci. 2018, 97, 1220–1228. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sacchetti, G.; Di Mattia, C.; Pittia, P.; Martino, G. Application of a radical scavenging activity test to measure the total antioxidant activity of poultry meat. Meat Sci. 2008, 80, 1085. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [Green Version]
- Anraku, M.; Gebicki, J.M.; Iohara, D.; Tomida, H.; Uekama, K.; Maruyama, T.; Hirayama, F.; Otagiri, M. Antioxidant activities of chitosans and its derivatives in in vitro and in vivo studies. Carbohydr. Polym. 2018, 199, 149. [Google Scholar] [CrossRef]
- Buldurun, K.; Turan, N.; Bursal, E.; Mantarcı, A.; Turkan, F.; Taslimi, P.; Gülçin, İ. Synthesis, spectroscopic properties, crystal structures, antioxidant activities and enzyme inhibition determination of Co(II) and Fe(II) complexes of Schiff base. Res. Chem. Intermed. 2020, 46, 283–297. [Google Scholar] [CrossRef]
- Bibi Sadeer, N.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The versatility of antioxidant assays in food science and safety—Chemistry, applications, strengths, and limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular -antioxidant-rich US foods. J. Food Comp. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Apak, R.; Ozyurek, M.; Güçlü, K.; Capanoğlu, E. Antioxidant activity/ capacity measurement. Classification, physicochemical principles, mechanisms, and electron transfer (ET)-based assays. J. Agric. Food Chem. 2016, 64, 997–1027. [Google Scholar] [CrossRef] [Green Version]
- Prior, R.L.; Wu, X.L.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food. Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Miguel, M.G. Antioxidant activity of medicinal and aromatic plants. A review. Flavour Fragr. J. 2010, 25, 291–312. [Google Scholar] [CrossRef]
- Haunshi, S.; Prince, L.L.L. Kadaknath: A popular native chicken breed of India with unique black color characteristics. Poult. Sci. J. 2021, 77, 427–440. [Google Scholar] [CrossRef]
- Kohen, R.; Yamamoto, Y.; Cundy, K.C.; Ames, B.N. Antioxidant activity of carnosine, homocarnosine, and anserine present in muscle and brain. Proc. Natl. Acad. Sci. USA 1988, 85, 3175–3179. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.S.; Kim, D.; Nyberg, S.; Poma, A.; Cecchin, D.; Jain, S.A.; Kim, K.A.; Shin, Y.J.; Kim, E.H.; Kim, M.; et al. LRP-1 functionalized polymersomes enhance the efficacy of carnosine in experimental stroke. Sci. Rep. 2020, 10, 699. [Google Scholar] [CrossRef] [Green Version]
- Wu, G. Amino Acids: Biochemistry and Nutrition; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Dolan, E.; Saunders, B.; Dantas, W.; Murai, I.; Roschel, H.; Artioli, G.; Harris, R.; Bicudo, J.; Sale, C.; Gualano, B.A. Comparative study of humming birds and chickens provides mechanistic insight on the histidine containing dipeptide role in skeletal muscle metabolism. Sci. Rep. 2018, 8, 14788. [Google Scholar] [CrossRef]
- Wu, G. Important roles of dietary taurine, creatine, carnosine, anserine and 4-hydroxyproline in human nutrition and health. Amino Acids 2020, 52, 329–360. [Google Scholar] [CrossRef] [Green Version]
- Intarapichet, K.O.; Maikhunthod, B. Genotype and gender differences in carnosine extracts and antioxidant activities of chicken breast and thigh meats. Meat Sci. 2005, 71, 634–642. [Google Scholar] [CrossRef]
- Mohamed, S. Functional foods against metabolic syndrome (obesity, diabetes, hypertension and dyslipidemia) and cardiovascular disease. Trends Food Sci. Technol. 2014, 35, 114–128. [Google Scholar] [CrossRef]
- Beseni, B.K.; Bagla, V.P.; Njanje, I.; Matsebatlela, T.M.; Mampuru, L.; Mokgotho, M.P. Antioxidant, antiglycation, and hypoglycaemic effect of Seriphiumplumosum crude plant extracts. Evid. Based Complement Alternat. Med. 2017, 2017, 6453567. [Google Scholar] [CrossRef] [Green Version]
- Starowicz, M.; Zieliński, H. Inhibition of advanced glycation end-product formation by high antioxidant-leveled spices commonly used in European cuisine. Antioxidants 2019, 8, 100. [Google Scholar] [CrossRef] [Green Version]
- Freund, M.A.; Chen, B.; Decker, E.A. The inhibition of advanced glycation end products by carnosine and other natural dipeptides to reduce diabetic and age-related complications. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1367–1378. [Google Scholar] [CrossRef] [Green Version]
- Szwergold, B. Carnosine and anserine act as effective transglycating agents in decomposition of aldose-derived Schiff bases. Biochem. Biophys. Res. Commun. 2005, 1, 36–41. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef]
- Bassinello, D.; de Salles Painelli, V.; Dolan, E.; Lixandrão, M.; Cajueiro, M.; de Capitani, M.; Saunders, B.; Sale, C.; Artioli, G.G.; Gualano, B.; et al. Beta-alanine supplementation improves isometric, but not isotonic or isokinetic strength endurance in recreationally strength-trained young men. Amino Acids 2019, 51, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, R.P.; de Oliveira, L.F.; Saunders, B.; de Andrade Kratz, C.; de SallesPainelli, V.; da Eira Silva, V.; Marins, J.C.B.; Franchini, E.; Gualano, B.; Artioli, G.G. Effects of β-alanine and sodium bicarbonate supplementation on the estimated energy system contribution during high-intensity intermittent exercise. Amino Acids 2019, 51, 83–96. [Google Scholar] [CrossRef]
- Varanoske, A.N.; Hoffman, J.R.; Church, D.D.; Coker, N.A.; Baker, K.M.; Dodd, S.J.; Harris, R.C.; Oliveira, L.P.; Dawson, V.L.; Wang, R.; et al. Comparison of sustained-release and rapid-release β-alanine formulations on changes in skeletal muscle carnosine and histidine content and isometric performance following a muscle-damaging protocol. Amino Acids. 2019, 51, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Antonini, F.M.; Petruzzi, E.; Pinzani, P.; Orlando, C.; Poggesi, M.; Serio, M.; Pazzagli, M.; Masotti, G. The meat in the diet of aged subjects and the antioxidant effects of carnosine. Arch. Gerontol. Geriatr. Suppl. 2002, 8, 7–14. [Google Scholar] [CrossRef]
- Bauchart, C.; Savary-Auzelous, I.; Mirand, P.; Thomas, E.; Morzel, M.; Remond, D. Carnosine concentration of ingested meat affects carnosine net release into the portal vein of mini pigs. J. Nutr. 2007, 137, 589–593. [Google Scholar] [CrossRef] [Green Version]
- Peiretti, P.; Medana, C.; Visentin, S.; Bello, F.; Meineri, G. Effect of cooking method on carnosine and its homoglogues, pentosidine and thiobarbituric acid-reactive substance contents in beef and turkey meat. Food Chem. 2012, 132, 80–85. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Brosnan, M.E. Creatine: Endogenous metabolite, dietary, and therapeutic supplement. Annu. Rev. Nutr. 2007, 27, 241–261. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.N.; Agharkar, A.S.; Gonzales, E.B. A review of creatine supplementation in age-related diseases: More than a supplement for athletes. F1000Research 2014, 3, 222. [Google Scholar] [CrossRef] [Green Version]
- De Benedetto, F.; Pastorelli, R.; Ferrario, M.; de Blasio, F.; Marinari, S.; Brunelli, L.; Wouters, E.F.M.; Polverino, F.; Celli, B.R. Supplementation with Qter® and Creatine improves functional performance in COPD patients on long term oxygen therapy. Respir. Med. 2018, 142, 86–93. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sr No | Antioxidant Capacity Assays | Unit of Measurement | Genetic Group | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cobb Broiler | Jabalpur Color Chicken | ||||||||||
| Mean | SE | Minimum | Maximum | Mean | SE | Minimum | Maximum | ||||
| 1 | ABTS (2,2′-azinobis-3- ethylbenzothiazoline-6-sulfonic acid) radical scavenging assay | % Inhibition | 43.78 | 1.47 | 33.33 | 57.58 | 52.12 | 1.36 | 43.94 | 63.64 | * |
| TEABTS (Trolox equivalent antioxidative capacity) | Trolox equivalent (TE) µM/g of tissue | 6062.5 | 257.31 | 3350 | 8350 | 7375 | 210.65 | 6100 | 9350 | * | |
| 2 | DPPH (1,1-diphenyl-2-picrylhydrazyl) radical scavenging assay | % Inhibition | 70.06 | 0.59 | 60.51 | 74.30 | 73.92 | 0.44 | 68.80 | 76.63 | * |
| 3 | FRAP (ferric reducing antioxidant power) | mM Fe2+/g tissue | 15.24 | 0.40 | 11.6 | 18.5 | 22.84 | 0.25 | 20.7 | 24.1 | * |
| 4 | Cupric reducing antioxidative capacity (CUPRAC) assay | Trolox equivalent (TE) mM/g of tissue | 9 | 0.24 | 7.219 | 10.438 | 12.71 | 0.32 | 10.75 | 16.32 | * |
| 5 | ORAC (oxygen radical absorption capacity) assay | Trolox equivalent (TE) µM/g of tissue | 748.56 | 7.48 | 713.02 | 776.0 | 765.82 | 9.48 | 730.02 | 803.07 | NS |
| 6 | MCA (metal chelation activity) | % Inhibition | 53.63 | 1.79 | 40.22 | 65.71 | 46.3 | 2.36 | 30.5 | 63.41 | * |
| EDTA equivalent activity— EEA µM/g of tissue | 2819.29 | 85.84 | 2170.02 | 3400.03 | 2468.22 | 112.81 | 1712.03 | 3290.30 | * | ||
| Carnosine equivalent activity—CEA mM/g of tissue | 148.63 | 4.34 | 116.03 | 178.24 | 130.89 | 5.70 | 92.75 | 172.62 | * | ||
| 7 | Superoxide dismutase activity | % Inhibition | 93.16 | 1.07 | 83.25 | 99.62 | 95.84 | 0.93 | 88.12 | 99.87 | NS |
| Sr No | Antioxidant Capacity Assays | Unit of Measurement | Genetic Group | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cobb Broiler | Jabalpur Color Chicken | ||||||||||
| Mean | SE | Minimum | Maximum | Mean | SE | Minimum | Maximum | ||||
| 1 | ABTS (2,2′-azinobis-3- ethylbenzothiazoline-6-sulfonic acid) radical scavenging assay | % Inhibition | 29.62 | 1.27 | 21.21 | 37.88 | 28.48 | 1.06 | 21.21 | 36.36 | NS |
| TEABTS (Trolox equivalent antioxidative capacity) | Trolox equivalent (TE) µM/g of tissue | 3737.50 | 210.32 | 2350.00 | 5100.00 | 3550.00 | 175.47 | 2350.00 | 4850.00 | NS | |
| 2 | DPPH (1,1-diphenyl-2-picrylhydrazyl) radical scavenging assay | % Inhibition | 63.46 | 0.56 | 60.17 | 70.78 | 67.27 | 0.63 | 60.32 | 71.28 | * |
| 3 | FRAP (ferric reducing antioxidant power) | mM Fe2+/g tissue | 19.20 | 0.31 | 16.80 | 21.60 | 26.83 | 0.36 | 24.10 | 29.80 | * |
| 4 | Cupric reducing antioxidative capacity (CUPRAC) assay | Trolox equivalent (TE) mM/g of tissue | 7.16 | 0.25 | 5.08 | 9.44 | 7.49 | 0.30 | 5.45 | 10.50 | NS |
| 5 | ORAC (oxygen radical absorption capacity) assay | Trolox equivalent (TE) µM/g of tissue | 762.82 | 9.19 | 717.41 | 795.06 | 785.95 | 6.40 | 751.26 | 822.93 | * |
| 6 | MCA (metal chelation activity) | % Inhibition | 80.75 | 0.95 | 71.25 | 86.61 | 63.13 | 1.87 | 47.32 | 81.25 | * |
| EDTA equivalent activity— EEA µM/g of tissue | 4117.96 | 45.60 | 3662.98 | 4398.47 | 3273.85 | 89.61 | 2516.97 | 4141.91 | * | ||
| Carnosine equivalent activity—CEA mM/g of tissue | 214.26 | 2.30 | 191.27 | 228.43 | 171.60 | 4.53 | 133.36 | 215.47 | * | ||
| 7 | Superoxide dismutase activity | % Inhibition | 89.50 | 1.34 | 75.50 | 99.88 | 93.69 | 1.32 | 77.00 | 98.50 | * |
| Sr No | Metabolite | Genotype | Breast | Thigh | ||
|---|---|---|---|---|---|---|
| Mean ± SE | Range | Mean ± SE | Range | |||
| 1 | Carnosine (mg/g of tissue) | Cobb broiler | 2.73 ± 0.10 | 2.11–3.39 | 0.98 ± 0.03 # | 0.78–1.20 |
| JBC chicken | 2.66 ± 0.09 | 2.15–3.39 | 1.11 ± 0.04 # | 0.74–1.36 | ||
| 2 | Anserine (mg/g of tissue) | Cobb broiler | 4.85 ± 0.22 | 3.51–6.33 | 2.27 ± 0.14 # | 1.67–3.14 |
| JBC chicken | 5.11 ± 0.12 | 4.27–6.10 | 2.01 ± 0.16 # | 1.07–2.92 | ||
| 3 | Creatine (mg/g of tissue) | Cobb broiler | 3.06 ± 0.12 | 2.39–3.86 | 2.56 ± 0.10 # | 2.0–3.25 |
| JBC chicken | 2.80 ± 0.12 | 1.39–2.85 | 2.46 ± 0.22 | 1.58–3.40 | ||
| 4 | Protein (g/100 g of tissue) | Cobb broiler | 21.81 ± 0.10 | 18.48–24.22 | 18.31 ± 0.03 # | 16.65–20.14 |
| JBC chicken | 25.65 ± 0.39 * | 22.06–28.63 | 19.04 ± 0.23 *,# | 17.48–20.89 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, R.; Sehrawat, R.; Ahlawat, S.; Sharma, V.; Thakur, M.S.; Mishra, A.K.; Arora, R.; Tantia, M.S. Functional Quality Characteristics of the Meat from a Dual-Purpose Poultry Crossbreed Suitable for Backyard Rearing in Comparison to Commercial Broilers. Foods 2023, 12, 2434. https://doi.org/10.3390/foods12132434
Sharma R, Sehrawat R, Ahlawat S, Sharma V, Thakur MS, Mishra AK, Arora R, Tantia MS. Functional Quality Characteristics of the Meat from a Dual-Purpose Poultry Crossbreed Suitable for Backyard Rearing in Comparison to Commercial Broilers. Foods. 2023; 12(13):2434. https://doi.org/10.3390/foods12132434
Chicago/Turabian StyleSharma, Rekha, Renuka Sehrawat, Sonika Ahlawat, Vivek Sharma, Mohan Singh Thakur, A. K. Mishra, Reena Arora, and M. S. Tantia. 2023. "Functional Quality Characteristics of the Meat from a Dual-Purpose Poultry Crossbreed Suitable for Backyard Rearing in Comparison to Commercial Broilers" Foods 12, no. 13: 2434. https://doi.org/10.3390/foods12132434