
Phenolic Profiles, Antioxidant, and Hypoglycemic Activities of Ribes meyeri Fruits

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Reagents
2.2. Extraction Procedure
2.3. Purification of Anthocyanin from R. meyeri Fruits
2.4. Qualitative Analysis of Phenolics from R. meyeri Fruits
2.5. Quantitative Analysis of Anthocyanin from R. meyeri Fruits
2.5.1. Determination of Anthocyanin Content
2.5.2. UPLC-MS/MS Analysis
2.6. Determination of Antioxidant Activity
2.7. Antidiabetic Activity Assay
2.8. Statistical Analysis
3. Results
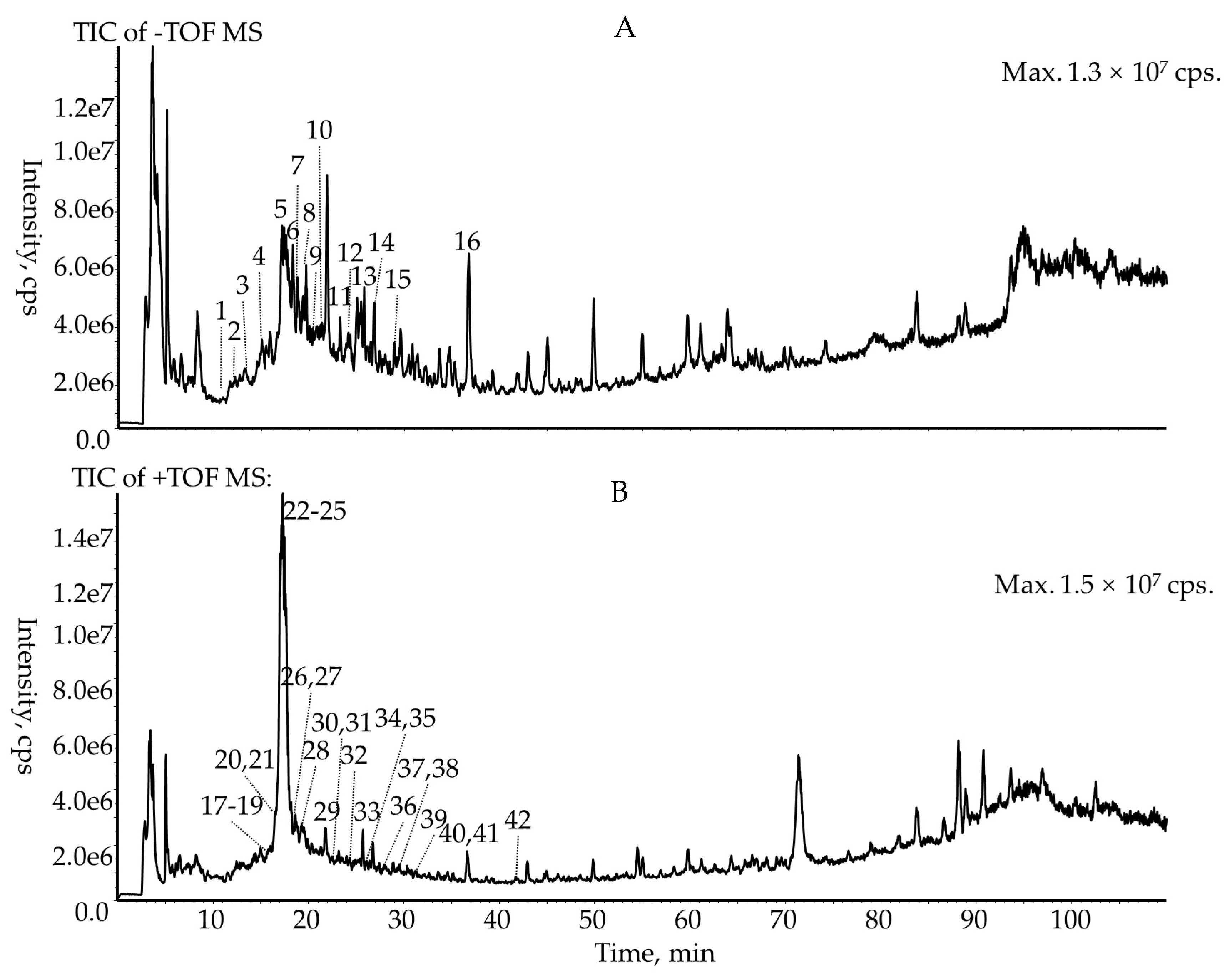
3.1. Qualitative Identification of the Chemical Composition of R. meyeri Fruits
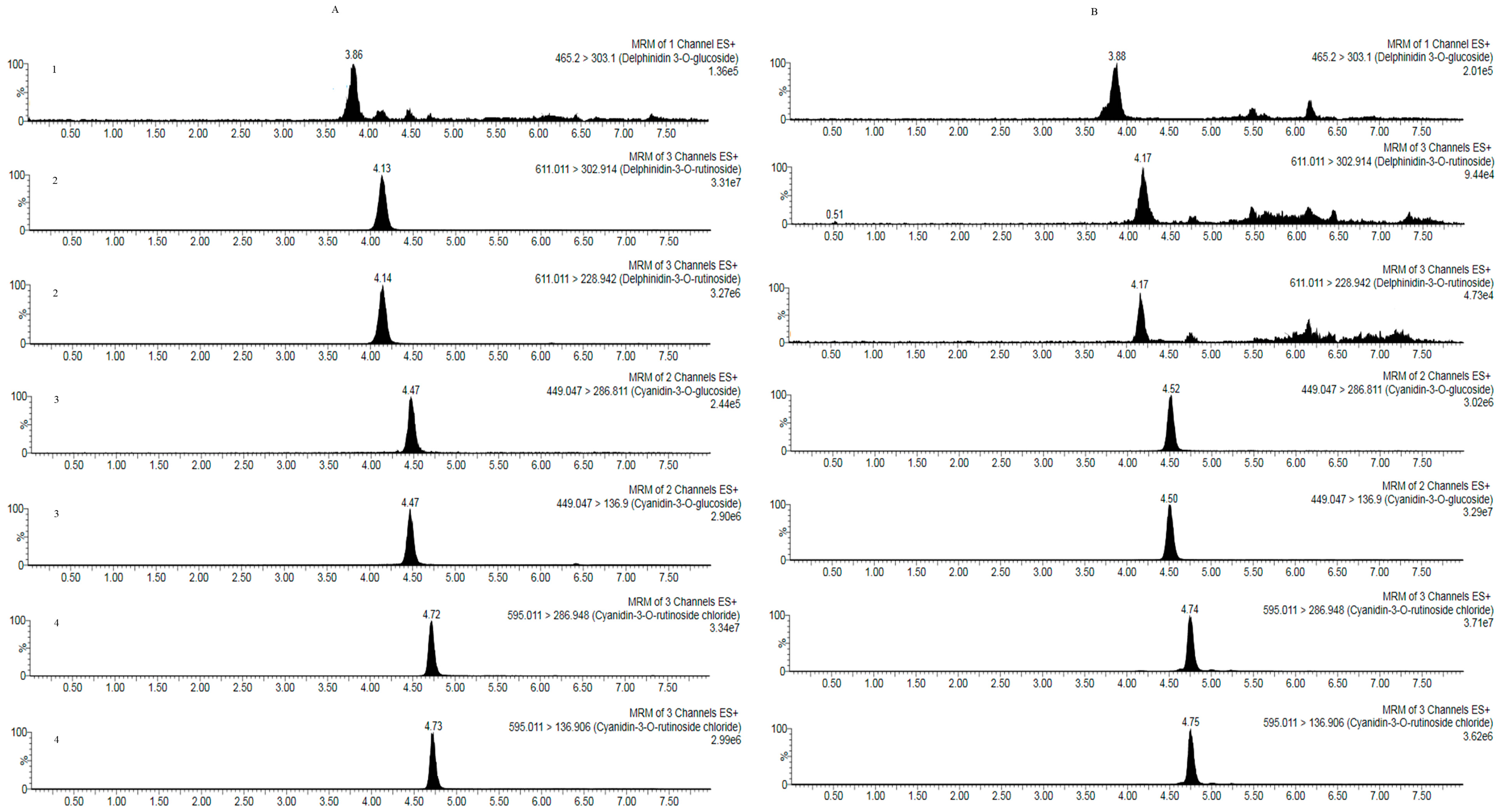
3.2. Quantitative Analysis of the Anthocyanins of R. meyeri Fruits
3.3. Antioxidant Activities
3.4. α-Amylase, α-Glucosidase, and PTP1B Inhibitory Activities
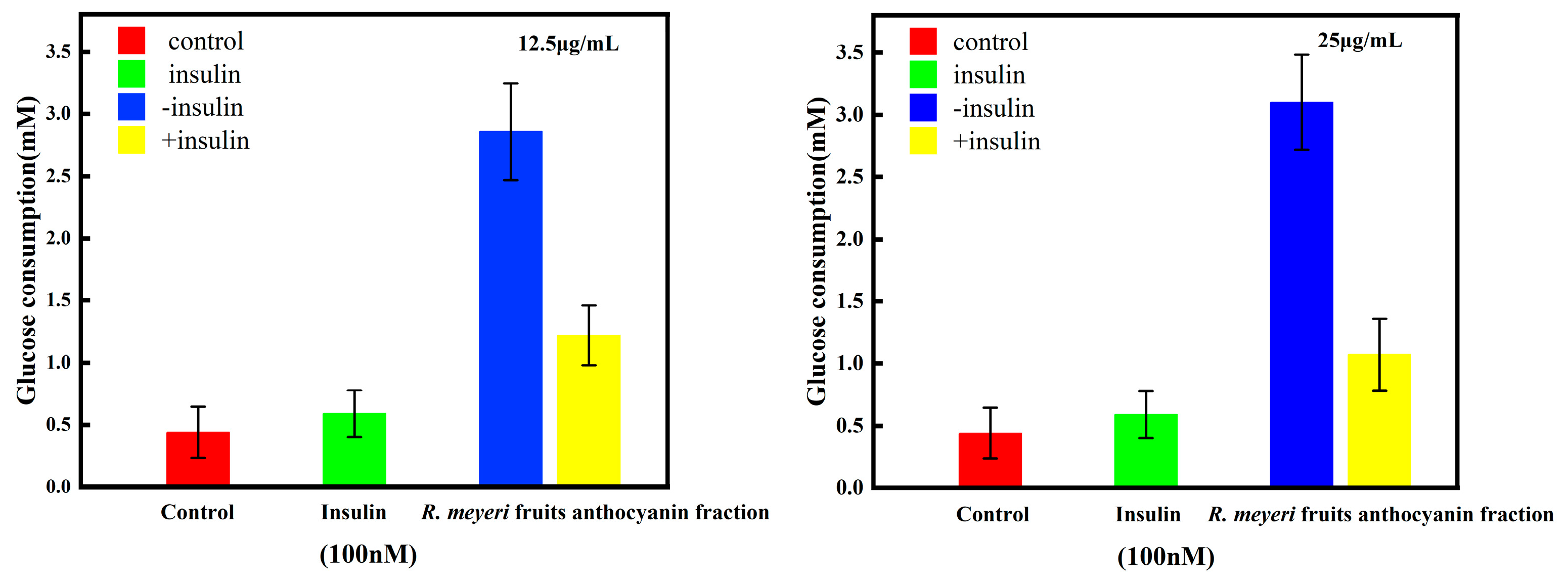
3.5. The Effect of R. meyeri Fruits Anthocyanin on Glucose Consumption of 3T3-L1 Adipocytes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gopalan, A.; Reuben, S.C.; Ahmed, S.; Darvesh, A.S.; Hohmann, J.; Bishayee, A. The health benefits of blackcurrants. Food Funct. 2012, 3, 795–809. [Google Scholar] [CrossRef]
- Pap, N.; Fidelis, M.; Azevedo, L.; do Carmo, M.A.V.; Wang, D.; Mocan, A.; Pereira, E.P.R.; Xavier-Santos, D.; Sant’Ana, A.S.; Yang, B.; et al. Berry polyphenols and human health: Evidence of antioxidant, anti-inflammatory, microbiota modulation, and cell-protecting effects. Curr. Opin. Food Sci. 2021, 42, 167–186. [Google Scholar] [CrossRef]
- Slimestad, R.; Solheim, H. Anthocyanins from black currants (Ribes nigrum L.). J. Agric. Food Chem. 2002, 50, 3228–3231. [Google Scholar] [CrossRef]
- Goiffon, J.P.; Mouly, P.P.; Gaydou, E.M. Anthocyanic pigment determination in red fruit juices, concentrated juices and syrups using liquid chromatography. Anal. Chim. Acta 1999, 382, 39–50. [Google Scholar] [CrossRef]
- Mitic, V.D.; Dimitrijevic, M.V.; Pavlovic, J.; Nikolic, J.; Simonovic, S.R.; Stankov, V.P.J.; Stojanovic, G.S. Comprehensive Evaluation of Antioxidant Activity of Ribes berry Fruit Species: A Chemometric Approach. Anal. Lett. 2018, 51, 908–920. [Google Scholar] [CrossRef]
- Barik, S.K.; Russell, W.R.; Moar, K.M.; Cruickshank, M.; Scobbie, L.; Duncan, G.; Hoggard, N. The anthocyanins in black currants regulate postprandial hyperglycaemia primarily by inhibiting α-glucosidase while other phenolics modulate salivary α-amylase, glucose uptake and sugar transporters. J. Nutr. Biochem. 2020, 78, 108325. [Google Scholar] [CrossRef]
- Paunović, S.M.; Nikolić, M.; Miletić, R.; Mašković, P.; Milinković, M.; Karaklajić-Stajić, Ž. Phytochemical Screening and Biological Activity of Extract Berries of Black Currant (Ribes nigrum L.). Erwerbs Obstbau 2018, 61, 71–78. [Google Scholar] [CrossRef]
- Zorzi, M.; Gai, F.; Medana, C.; Aigotti, R.; Morello, S.; Peiretti, P.G. Bioactive Compounds and Antioxidant Capacity of Small Berries. Foods 2020, 9, 623. [Google Scholar] [CrossRef] [PubMed]
- Koss-Mikolajczyk, I.; Kusznierewicz, B.; Bartoszek, A. The Relationship between Phytochemical Composition and Biological Activities of Differently Pigmented Varieties of Berry Fruits; Comparison between Embedded in Food Matrix and Isolated Anthocyanins. Foods 2019, 8, 646. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, J.M.; Toledo, T. Wild berries and related wild small fruits as traditional healthy foods. Crit. Rev. Food Sci. Nutr. 2022, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Motti, R. Wild Edible Plants: A Challenge for Future Diet and Health. Plants 2022, 11, 344. [Google Scholar] [CrossRef] [PubMed]
- Xinjiang Chinese Herbal Medicine Editorial Committee. Chinese Herbal Medicine in Xinjiang; Xinjiang People’s Press: Urumqi, China, 1976; pp. 349–350. [Google Scholar]
- Li, Z.; Guo, H.; XU, W.; He, D. Antioxidant activities and PTP1B inhibitory effect of different extracts from three Xinjiang plants. Chin. J. Exp. Tradit. Med. 2015, 21, 28–33. [Google Scholar]
- Kashtoh, H.; Baek, K.H. Recent Updates on Phytoconstituent Alpha-Glucosidase Inhibitors: An Approach towards the Treatment of Type Two Diabetes. Plants 2022, 11, 2722. [Google Scholar] [CrossRef] [PubMed]
- Borah, P.K.; Sarkar, A.; Duary, R.K. Water-soluble vitamins for controlling starch digestion: Conformational scrambling and inhibition mechanism of human pancreatic alpha-amylase by ascorbic acid and folic acid. Food Chem. 2019, 288, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Wang, J.; Ao, J.; Hou, Y.; Xi, M.; Cai, Y.; Li, M.; Luo, A. Structure-activity relationships and the underlying mechanism of alpha-amylase inhibition by hyperoside and quercetin: Multi-spectroscopy and molecular docking analyses. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2023, 285, 121797. [Google Scholar] [CrossRef]
- Proenca, C.; Ribeiro, D.; Freitas, M.; Carvalho, F.; Fernandes, E. A comprehensive review on the antidiabetic activity of flavonoids targeting PTP1B and DPP-4: A structure-activity relationship analysis. Crit. Rev. Food Sci. Nutr. 2022, 62, 4095–4151. [Google Scholar] [CrossRef]
- Bai, X.; Fan, W.; Luo, Y.; Liu, Y.; Zhang, Y.; Liao, X. Fast Screening of Protein Tyrosine Phosphatase 1B Inhibitor from Salvia miltiorrhiza Bge by Cell Display-Based Ligand Fishing. Molecules 2022, 27, 7896. [Google Scholar] [CrossRef]
- Xue, Y.; Qing, L.S.; Yong, L.; Xu, X.S.; Hu, B.; Tang, M.Q.; Xie, J. Determination of Flavonoid Glycosides by UPLC-MS to Authenticate Commercial Lemonade. Molecules 2019, 24, 3016. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Lin, Z.; Zhang, H.; Liu, Z.; Xu, Y.; Xu, G.; Li, H.; Ji, R.; Luo, W.; Qiu, Y.; et al. Anthocyanin Accumulation in the Leaves of the Purple Sweet Potato (Ipomoea batatas L.) Cultivars. Molecules 2019, 24, 3743. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Matsumoto, H.; Morifuji, M.; Iida, H.; Takeuchi, Y. Development and validation of a liquid chromatography tandem mass spectrometry method for simultaneous determination of four anthocyanins in human plasma after black currant anthocyanins ingestion. J. Agric. Food Chem. 2010, 58, 1174–1179. [Google Scholar] [CrossRef]
- Zhao, Y.; Lu, H.; Wang, Q.; Liu, H.; Shen, H.; Xu, W.; Ge, J.; He, D. Rapid qualitative profiling and quantitative analysis of phenolics in Ribes meyeri leaves and their antioxidant and antidiabetic activities by HPLC-QTOF-MS/MS and UHPLC-MS/MS. J. Sep. Sci. 2021, 44, 1404–1420. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Cao, J.; Yue, J.; Zhang, X.; Zhao, Y. New triterpenoids from acorns of Quercus liaotungensis and their inhibitory activity against α-glucosidase, α-amylase and protein-tyrosine phosphatase 1B. J. Funct. Foods 2018, 41, 232–239. [Google Scholar] [CrossRef]
- Liu, Y.; Seeram, N.P. Liquid chromatography coupled with time-of-flight tandem mass spectrometry for comprehensive phenolic characterization of pomegranate fruit and flower extracts used as ingredients in botanical dietary supplements. J. Sep. Sci. 2018, 41, 3022–3033. [Google Scholar] [CrossRef]
- Gavrilova, V.; Kajdzanoska, M.; Gjamovski, V.; Stefova, M. Separation, characterization and quantification of phenolic compounds in blueberries and red and black currants by HPLC-DAD-ESI-MSn. J. Agric. Food Chem. 2011, 59, 4009–4018. [Google Scholar] [CrossRef]
- Larson, R.A. The antioxidants of higher plants. Phytochemistry 1988, 27, 969–978. [Google Scholar] [CrossRef]
- Bordonaba, J.G.; Terry, L.A. Biochemical profiling and chemometric analysis of seventeen UK-grown black currant cultivars. J. Agric. Food Chem. 2008, 56, 7422–7430. [Google Scholar] [CrossRef]
- Mota, J.C.; Almeida, P.P.; Freitas, M.Q.; Stockler-Pinto, M.B.; Guimaraes, J.T. Far from being a simple question: The complexity between in vitro and in vivo responses from nutrients and bioactive compounds with antioxidant potential. Food Chem. 2023, 402, 134351. [Google Scholar] [CrossRef]
- Manganaris, G.A.; Goulas, V.; Vicente, A.R.; Terry, L.A. Berry antioxidants: Small fruits providing large benefits. J. Sci. Food Agric. 2014, 94, 825–833. [Google Scholar] [CrossRef] [Green Version]
- Oboh, G.; Ogunsuyi, O.B.; Ogunbadejo, M.D.; Adefegha, S.A. Influence of gallic acid on alpha-amylase and alpha-glucosidase inhibitory properties of acarbose. J. Food Drug Anal. 2016, 24, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, Y.; Miao, M. Inhibition of α-amylase by polyphenolic compounds: Substrate digestion, binding interactions and nutritional intervention. Trends Food Sci. Technol. 2020, 104, 190–207. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, Y.; Zhou, W. In vitro and in silico studies of the inhibition activity of anthocyanins against porcine pancreatic α-amylase. J. Funct. Foods 2016, 21, 50–57. [Google Scholar] [CrossRef]
- Kaeswurm, J.A.H.; Konighofer, L.; Hogg, M.; Scharinger, A.; Buchweitz, M. Impact of B-Ring Substitution and Acylation with Hydroxy Cinnamic Acids on the Inhibition of Porcine alpha-Amylase by Anthocyanin-3-Glycosides. Foods 2020, 9, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Molecular Formula | Relative Molecular Mass | Ion Mode | Retention Time (min) | Parent Ion (m/z) | Daughter Ion (m/z) | Cone Hole Voltage (V) | Collision Energy (eV) |
|---|---|---|---|---|---|---|---|---|
| Delphinidin-3-O-glucoside | C27H31O15 | 465.1 | ESI+ | 3.89 | 465.20 | 303.1 * | 35 | 20 |
| Delphinidin-3-O-rutinoside | C27H31O16 | 611.1 | ESI+ | 4.13 | 611.01 | 302.9 * | 58 | 34 |
| 228.9 | 58 | 68 | ||||||
| Cyanidin-3-O- glucoside | C21H21O12 | 449.1 | ESI+ | 4.47 | 449.04 | 136.9 * | 34 | 50 |
| 286.8 | 34 | 66 | ||||||
| Cyanidin-3-O- rutinoside | C21H21O11 | 595.0 | ESI+ | 4.72 | 595.01 | 286.9 * | 58 | 26 |
| 136.9 | 58 | 70 |
| No. | tR (min) | Molecular Formula | [M-H]− | Major MS2 Ions | Error (ppm) | Identification |
|---|---|---|---|---|---|---|
| 1 | 10.97 | C13H16O10 | 331.0672 | 169, 125 | 3.70 | Galloyl Hex |
| 2 | 12.18 | C15H14O7 | 305.0670 | 261, 219, 179, 167, 125 | 4.66 | Gallocatechin/Epigallocatechin |
| 3 | 13.18 | C13H16O8 | 299.0775 | 239, 179, 137 | 4.27 | Benzoyl Hex |
| 4 | 15.47 | C15H18O8 | 325.0933 | 163, 145, 119 | 4.63 | Hydrocinnamoyl Hex |
| 5 | 17.81 | C27H30O15 | 593.1525 | 285, 284, 255, 227 | 4.05 | Luteolin-Hex-DeHex |
| 6 | 18.11 | C15H14O6 | 289.0720 | 245, 205, 203, 109 | 4.62 | Catechin |
| 7 | 18.91 | C15H18O8 | 325.0934 | 163, 145, 119 | 4.94 | Hydrocinnamoyl Hex |
| 8 | 19.96 | C15H18O8 | 325.0934 | 163, 145, 119 | 4.94 | Hydrocinnamoyl Hex |
| 9 | 20.34 | C9H8O4 | 179.0347 | 135 | 4.55 | Caffeic acid |
| 10 | 21.15 | C15H14O6 | 289.0719 | 245, 205, 203, 109 | 4.62 | Epicatechin |
| 11 | 23.23 | C33H40O20 | 755.2058 | 609, 447, 301, 300, 271, 255, 243, 179, 151 | 3.81 | Quercetin-DeHex-Hex-DeHex |
| 12 | 24.87 | C9H8O3 | 163.0397 | 119 | 4.47 | Coumaric acid |
| 13 | 25.73 | C27H30O16 | 609.1474 | 301, 300, 271, 255, 243, 229, 179, 151 | 3.90 | Quercetin-3-O-Hex-DeHex |
| 14 | 26.80 | C21H20O12 | 463.0891 | 301, 300, 283, 271, 255 | 4.31 | Quercetin-3-O-Hex |
| 15 | 29.69 | C15H12O8 | 319.0463 | 183, 153, 139 | 3.56 | Dihydromyricetin |
| 16 | 36.73 | C34H24O22 | 301.0353 | 271, 255, 229, 179, 151 | 3.39 | Quercetin |
| No. | tR (min) | Molecular Formula | [M+H]− | Major MS2 Ions | Error (ppm) | Identification |
|---|---|---|---|---|---|---|
| 17 | 15.77 | C33H41O20 | 757.2218 | 611, 449, 433, 287, 241, 213 | 4.26 | Cyanidin-Hex-Hex-DeHex |
| 18 | 15.81 | C33H41O20 | 757.2222 | 611, 449, 433, 287, 241, 213 | 4.79 | Cyanidin-Hex-Hex-DeHex |
| 19 | 15.88 | C32H39O20 | 743.2065 | 581, 449, 303, 287, 241, 213 | 4.82 | Cyanidin-Hex-Pen-Hex |
| 20 | 16.47 | C27H31O16 | 611.1633 | 449, 287, 241, 213 | 4.32 | Cyanidin-Hex-Hex |
| 21 | 16.72 | C32H39O19 | 727.2116 | 581, 449, 287, 241, 213 | 4.94 | Cyanidin-Hex-Pen-DeHex |
| 22 | 17.00 | C21H21O11 | 449.1097 | 287, 241, 213 | 4.15 | Cyanidin-3-glucoside |
| 23 | 17.18 | C26H29O15 | 581.1528 | 449, 287, 241, 213 | 4.65 | Cyanidin-Hex-Pen |
| 24 | 17.48 | C32H39O19 | 727.2112 | 581, 449, 287, 241, 213 | 4.39 | Cyanidin-Hex-Pen-DeHex |
| 25 | 17.70 | C27H31O15 | 595.1683 | 449, 287, 241, 213 | 4.29 | Cyanidin-Hex-DeHex |
| 26 | 18.81 | C21H21O10 | 433.1148 | 271, 225, 197 | 4.33 | Pelargonidin-Hex |
| 27 | 18.83 | C33H41O20 | 757.2220 | 611, 449, 433, 287, 241, 213 | 4.53 | Cyanidin-Hex-Hex-DeHex |
| 28 | 19.91 | C20H19O10 | 419.0985 | 287, 241, 213 | 2.93 | Cyanidin-Pen |
| 29 | 22.03 | C21H21O10 | 433.1145 | 287, 241, 213 | 3.64 | Cyanidin-DeHex |
| 30 | 23.13 | C32H39O20 | 743.2069 | 611, 465, 449, 303, 257, 229 | 5.40 | Cyanidin-Hex-Hex-Pen |
| 31 | 23.16 | C33H41O20 | 757.2223 | 611, 465, 303, 257, 229 | 4.92 | Delphinidin-Hex-DeHex-DeHex |
| 32 | 24.30 | C26H29O16 | 597.1478 | 465, 303, 257, 229 | 4.67 | Delphinidin-Hex-Pen |
| 33 | 25.71 | C27H31O16 | 611.1632 | 465, 303, 257, 229 | 4.15 | Delphinidin-Hex-DeHex |
| 34 | 26.30 | C21H21O12 | 465.1045 | 303, 257, 229 | 3.76 | Delphinidin-Hex |
| 35 | 26.70 | C21H21O12 | 465.1046 | 303, 257, 229 | 3.97 | Delphinidin-Hex |
| 36 | 27.69 | C15H11O5 | 271.0613 | 253, 225, 197, 150 | 4.43 | Pelargonidin isomer |
| 37 | 29.33 | C15H11O5 | 271.0608 | 253, 225, 197, 150 | 2.58 | Pelargonidin isomer |
| 38 | 29.70 | C15H11O5 | 271.0611 | 253, 225, 197, 150 | 3.69 | Pelargonidin isomer |
| 39 | 31.30 | C15H11O5 | 271.0611 | 253, 225, 197, 150 | 3.69 | Pelargonidin isomer |
| 40 | 36.54 | C15H11O7 | 303.0511 | 257, 229, 201, 165, 153 | 3.86 | Delphinidin |
| 41 | 36.93 | C15H11O5 | 271.0610 | 253, 225, 197, 150 | 3.32 | Pelargonidin isomer |
| 42 | 41.71 | C15H11O6 | 287.0563 | 241, 213 | 4.48 | Cyanidin |
| Anthocyanins | Regression Equation | R2 | LOQ (ng/mL) | LOD (ng/mL) | Linear Range (ng/mL) |
|---|---|---|---|---|---|
| Delphinidin-3-O-glucoside | y = 26.0x − 322.6 | 0.9941 | 16.98 | 3.01 | 10–1500 |
| Cyanidin-3-O-glucoside | y = 312.6x − 3160.1 | 0.9980 | 7.09 | 2.36 | 10–1500 |
| Delphinidin-3-O-rutinoside | y = 6260.1x − 24904.0 | 0.9969 | 13.86 | 4.55 | 10–1500 |
| Cyanidin-3-O-rutinoside | y = 3216.0x − 8827.9 | 0.9957 | 18.44 | 5.38 | 10–1500 |
| Sample | Delphinidin-3-O-glucoside | Cyanidin-3-O-glucoside | Delphinidin-3-O-rutinoside | Cyanidin-3-O-rutinoside |
|---|---|---|---|---|
| Ribes meyeri fruits methanol extract | 2.27 ± 0.05 | 4.76 ± 0.02 | 0.06 ± 0.00 | 22.01 ± 0.37 |
| Ribes meyeri fruits anthocyanin fraction | 5.99 ± 0.09 | 26.70 ± 0.59 | 0.09 ± 0.00 | 79.36 ± 1.01 |
| Sample | DPPH Radical Scavenging Rate | ABTS Radical Scavenging Rate | α-Glucosidase Inhibitory Activity | α-Amylase Inhibitory Activity | PTP1B Inhibitory Activity | |
|---|---|---|---|---|---|---|
| R. meyeri fruits methanol extract | 111.22 ± 2.28 | 4.06 ± 0.32 | - | 0.35 ± 0.01 | 744.60 ± 6.00 | |
| R. meyeri fruits anthocyanin fraction | 74.22 ± 8.25 | 9.89 ± 0.87 | 56.67 ± 0.56 | 0.30 ± 0.01 | 218.72 ± 0.49 | |
| Positive control | Ascorbic acid | 5.97 ± 0.06 | 2.61 ± 0.12 | |||
| Acarbose | 330.23 ± 1.03 | 3.91 ± 0.05 | ||||
| Sodium vanadate | 1.46 ± 0.40 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Wang, Q.; Zhao, Y.; Ge, J.; He, D. Phenolic Profiles, Antioxidant, and Hypoglycemic Activities of Ribes meyeri Fruits. Foods 2023, 12, 2406. https://doi.org/10.3390/foods12122406
Zhang L, Wang Q, Zhao Y, Ge J, He D. Phenolic Profiles, Antioxidant, and Hypoglycemic Activities of Ribes meyeri Fruits. Foods. 2023; 12(12):2406. https://doi.org/10.3390/foods12122406
Chicago/Turabian StyleZhang, Le, Qiang Wang, Yayun Zhao, Juan Ge, and Dajun He. 2023. "Phenolic Profiles, Antioxidant, and Hypoglycemic Activities of Ribes meyeri Fruits" Foods 12, no. 12: 2406. https://doi.org/10.3390/foods12122406