

UV-B Radiation Induced the Changes in the Amount of Amino Acids, Phenolics and Aroma Compounds in Vitis vinifera cv. Pinot Noir Berry under Field Conditions

Abstract
:1. Introduction
2. Materials and Method
2.1. Sites and Materials
2.2. Treatments
2.3. Sample Collection
2.4. Measurement of Chemical Analysis in Grapes
2.4.1. Total Soluble Solids (TSS), pH and Titratable Acidity (TA) in Grape Juice
2.4.2. Amino Acids Analysis
2.4.3. Berry Phenolic Compounds Analysis
- [Tannin]e = tannins concentration in extraction (mg/L epicatechin eq.),
- Ve = final volume of extraction (L), and
- No. = initial number of berry samples.
2.4.4. Volatile Compounds Analysis
2.5. Statistical Analyses
3. Results
3.1. Effects of UV-B Exposure/Exclusion on the Physiology in Pinot Noir Vines
3.2. Effects of UV-B Exposure/Exclusion on Chemical Composition of Pinot Noir Fruit
3.2.1. Amino Acids
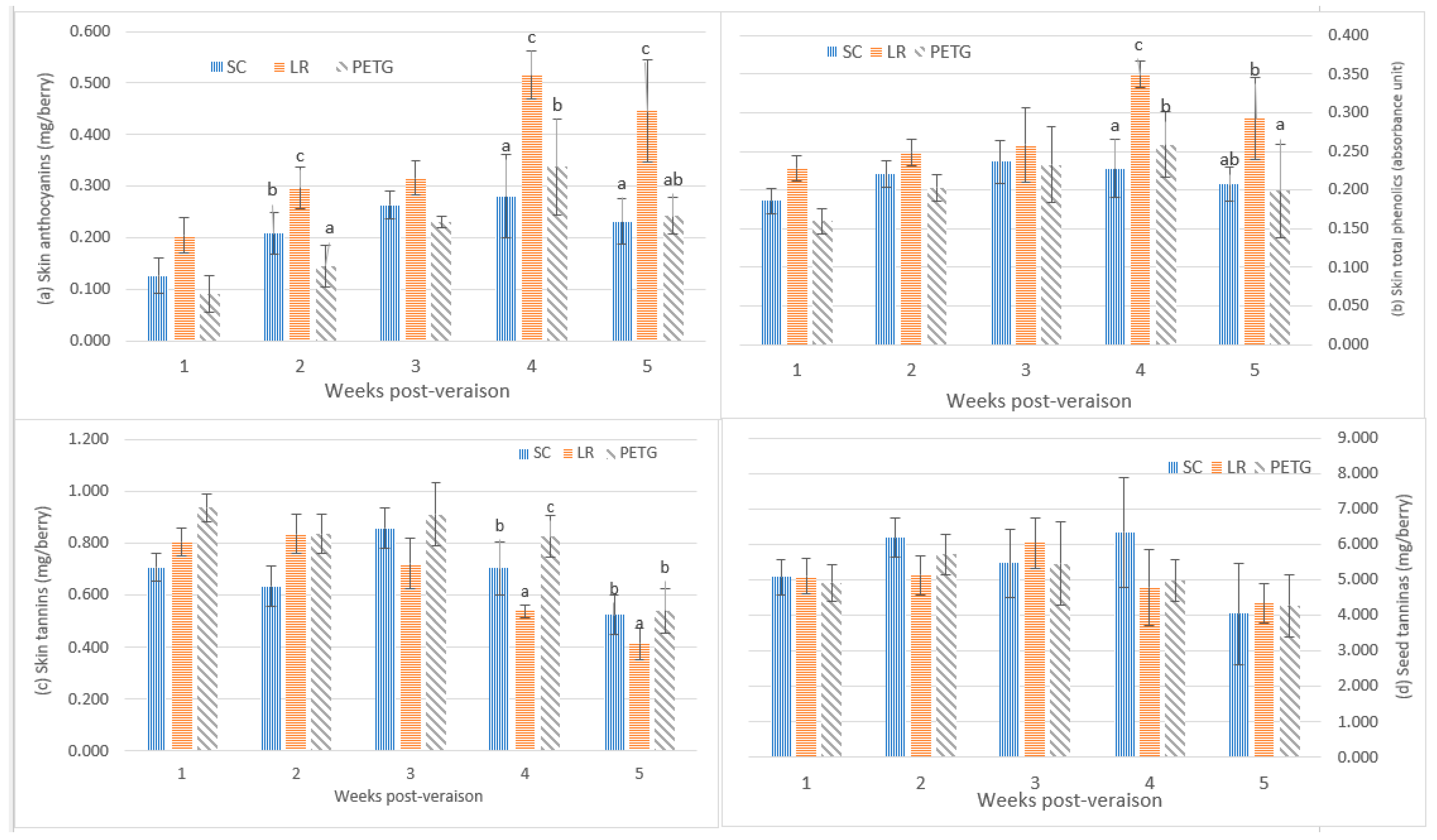
3.2.2. Phenolic Composition
3.2.3. Volatile Composition
4. Discussion
4.1. Alteration of Vine Yield and Pruning Weight as Induced by UV-B Radiation
4.2. The Effects of UV-B Radiation on Amino Acids in Berries
4.3. The Phenolic Composition in Berries in Response to UV-B Radiation
4.4. Effects of UV-B Radiation on Volatile Composition in Berry Juice
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Arias, L.; Berli, F.; Fontana, A.; Bottini, R.; Piccoli, P. Climate Change Effects on Grapevine Physiology and Biochemistry: Benefits and Challenges of High Altitude as an Adaptation Strategy. Front. Plant Sci. 2022, 13, 835425. [Google Scholar] [CrossRef] [PubMed]
- Del-Castillo-Alonso, M.; Monforte, L.; Tomás-Las-Heras, R.; Martínez-Abaigar, J.; Núñez-Olivera, E. To What Extent Are the Effects of UV Radiation on Grapes Conserved in the Resulting Wines? Plants 2021, 10, 1678. [Google Scholar] [CrossRef] [PubMed]
- Hideg, E.; Jansen, M.; Strid, A. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Gregan, S.M.; Winefield, C.; Jordan, B. Comparisons of controlled environment and vineyard experiments in sauvignon blanc grapes reveal similar UV-B signal transduction pathways for flavonol biosynthesis. Plant Sci. 2018, 276, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, M.; Martínez-Abaigar, J.; Glaab, J.; Jansen, M. UV-B induced secondary plant metabolites. Opt. Photonik 2014, 9, 34–37. [Google Scholar] [CrossRef]
- Schreiner, M.; Wiesner-Reinhold, M.; Baldermann, S.; Hanschen, F.S.; Neugart, S. UV-B-induced Changes in Secondary Plant Metabolites. In UV-B Radiation and Plant Life: Molecular Biology to Ecology; CABI: Wallingford, UK, 2017; pp. 39–57. [Google Scholar]
- Jordan, B.R.; Strid, Å.; Wargent, J.J. What role does UVB play in determining photosynthesis? In Handbook of Photosynthesis; CRC Press: Boca Raton, FL, USA, 2016; pp. 333–344. [Google Scholar]
- Whitelam, G.C.; Halliday, K.J. Annual Plant Reviews, Light and Plant Development; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Agasse, A.; Delrot, S.; Gerós, H. Biochemical changes throughout grape berry development and fruit and wine quality. Food 2007, 1, 1–22. [Google Scholar]
- Aleixandre-Tudo, J.L.; Buica, A.; Nieuwoudt, H.; Aleixandre, J.L.; du Toit, W. Spectrophotometric Analysis of Phenolic Compounds in Grapes and Wines. J. Agric. Food Chem. 2017, 65, 4009–4026. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.A.; Saucier, C.; Glories, Y. Grape and wine phenolics: History and perspective. Am. J. Enol. Vitic. 2006, 57, 239–248. [Google Scholar] [CrossRef]
- Silva Ferreira, A.; Monteiro, J.; Oliveira, C.; Guedes de Pinho, P. Study of major aromatic compounds in port wines from carotenoid degradation. Food Chem. 2008, 110, 83–87. [Google Scholar] [CrossRef]
- Edge, R.; Truscott, T.G. Singlet Oxygen and Free Radical Reactions of Retinoids and Carotenoids—A Review. Antioxidants 2018, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Jansen, M.A.; Gaba, V.; Greenberg, B.M. Higher plants and UV-B radiation: Balancing damage, repair and acclimation. Trends Plant Sci. 1998, 3, 131–135. [Google Scholar] [CrossRef]
- González-Barreiro, C.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gándara, J. Wine aroma compounds in grapes: A critical review. Crit. Rev. Food Sci. Nutr. 2015, 55, 202–218. [Google Scholar] [CrossRef] [PubMed]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. The microbiology of wine and vinifications. In Handbook of Enology 1; John Wiley & Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Loscos, N.; Hernández-Orte Pn Cacho, J.; Ferreira, V. Comparison of the suitability of different hydrolytic strategies to predict aroma potential of different grape varieties. J. Agric. Food Chem. 2009, 57, 2468–2480. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development: Sinauer Associates, Incorporated; Sinauer Associates Incorporated: Sunderland, MA, USA, 2015. [Google Scholar]
- Jansen, M.A.; Hectors, K.; O’Brien, N.M.; Guisez, Y.; Potters, G. Plant stress and human health: Do human consumers benefit from UV-B acclimated crops? Plant Sci. 2008, 175, 449–458. [Google Scholar] [CrossRef]
- Bender, D.A. Amino Acid Metabolism; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Nelson, D.L.; Lehninger, A.L.; Cox, M.M. Lehninger Principles of Biochemistry; Macmillan: New York, NY, USA, 2008. [Google Scholar]
- Keller, M. The Science of Grapevines: Anatomy and Physiology; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Mansour, G.; Ghanem, C.; Mercenaro, L.; Nassif, N.; Hassoun, G.; Del Caro, A. Effects of altitude on the chemical composition of grapes and wine: A review. Oeno One 2022, 56, 227–239. [Google Scholar] [CrossRef]
- Stines, A.; Grubb, J.; Gockowiak, H.; Henschke, P.A.; Høj, P.; Heeswijck, R.v. Proline and arginine accumulation in developing berries of Vitis vinifera L. in Australian vineyards: Influence of vine cultivar, berry maturity and tissue type. Aust. J. Grape Wine Res. 2000, 6, 150–158. [Google Scholar] [CrossRef]
- Gregan, S.; Wargent, J.; Liu, L.; Shinkle, J.; Hofmann, R.; Winefield, C.; Trought, M.; Jordan, B. Effects of solar ultraviolet radiation and canopy manipulation on the biochemical composition of Sauvignon Blanc grapes. Aust. J. Grape Wine Res. 2012, 18, 227–238. [Google Scholar] [CrossRef]
- Iland, P.; Ewart, A.; Sitters, J.; Markides, A.; Bruer, N. Techniques for Chemical Analysis and Quality Monitoring during Winemaking; Patrick Iland Wine Promotions: Campbelltown, Australia, 2000. [Google Scholar]
- Bonada, M.; Jeffery, D.; Petrie, P.R.; Moran, M.A.; Sadras, V.O. Impact of elevated temperature and water deficit on the chemical and sensory profiles of Barossa Shiraz grapes and wines. Aust. J. Grape Wine Res. 2015, 21, 240–253. [Google Scholar] [CrossRef]
- Sarneckis, C.J.; Dambergs, R.; Jones, P.; Mercurio, M.; Herderich, M.J.; Smith, P. Quantification of condensed tannins by precipitation with methyl cellulose: Development and validation of an optimised tool for grape and wine analysis. Aust. J. Grape Wine Res. 2006, 12, 39–49. [Google Scholar] [CrossRef]
- Canuti, V.; Conversano, M.; Calzi, M.L.; Heymann, H.; Matthews, M.A.; Ebeler, S.E. Headspace solid-phase microextraction–gas chromatography–mass spectrometry for profiling free volatile compounds in Cabernet Sauvignon grapes and wines. J. Chromatogr. A 2009, 1216, 3012–3022. [Google Scholar] [CrossRef]
- Dennis, E.G.; Keyzers, R.A.; Kalua, C.M.; Maffei, S.M.; Nicholson, E.L.; Boss, P.K. Grape contribution to wine aroma: Production of hexyl acetate, octyl acetate, and benzyl acetate during yeast fermentation is dependent upon precursors in the must. J. Agric. Food Chem. 2012, 60, 2638–2646. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Xu, Y.; Jiang, W.; Li, J. Identification and quantification of impact aroma compounds in 4 nonfloral Vitis vinifera varieties grapes. J. Food Sci. 2010, 75, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Qian, M.C. Development of C6 and other volatile compounds in Pinot Noir grapes determined by Stir Bar Sorptive Extraction-GC-MS. In Flavor Chemistry of Wine and Other Alcoholic Beverages; ACS Publications: Washington, DC, USA, 2012; pp. 81–99. [Google Scholar]
- Yuan, F.; Qian, M.C. Development of C13-norisoprenoids, carotenoids and other volatile compounds in Vitis vinifera L. cv. Pinot noir grapes. Food Chem. 2016, 192, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Jordan, B.; Hofmann, R.; Creasy, G. Effects of interaction of UV-B radiation and water deficit on phenolic compounds in 'Pinot Noir’fruit. Vitis 2022, 61, 27–36. [Google Scholar]
- Howell, G.S. Sustainable grape productivity and the growth-yield relationship: A review. Am. J. Enol. Vitic. 2001, 52, 165–174. [Google Scholar] [CrossRef]
- Kliewer, W.M.; Dokoozlian, N.K. Leaf area/crop weight ratios of grapevines: Influence on fruit composition and wine quality. Am. J. Enol. Vitic. 2005, 56, 170–181. [Google Scholar] [CrossRef]
- Lam, H.-M.; Coschigano, K.; Oliveira, I.; Melo-Oliveira, R.; Coruzzi, G. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef]
- Forde, B.G.; Lea, P.J. Glutamate in plants: Metabolism, regulation, and signalling. J. Exp. Bot. 2007, 58, 2339–2358. [Google Scholar] [CrossRef]
- Bell, S.J.; Henschke, P.A. Implications of nitrogen nutrition for grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Torres, N.; Hilbert, G.; Richard, T.; Sánchez-Díaz, M.; Delrot, S.; Aguirreolea, J.; Pascual, I.; Gomès, E. Ultraviolet-B radiation modifies the quantitative and qualitative profile of flavonoids and amino acids in grape berries. Phytochemistry 2014, 102, 106–114. [Google Scholar] [CrossRef]
- Keller, M.; Torres-Martinez, N. Does UV Radiation Affect Winegrape Composition? Acta Hortic. 2004, 640, 2002. [Google Scholar] [CrossRef]
- Schultz, H.; Linsenmeier, A.; Jahnisch, A.; Moller, M.; Gaubatz, B.; Varadi, G. Is grape composition affected by current levels ofUV-B radiation? Vitis 1998, 37, 191–192. [Google Scholar]
- Zhu, Y.F.; Sun, M.; Harrison, R.; Jordan, B.; Creasy, G.; Hofmann, R. Effects of UV-B and Water Deficit on Aroma Precursors in Grapes and Flavor Release during Wine Micro-Vinification and Consumption. Foods 2022, 11, 1336. [Google Scholar] [CrossRef]
- Garde-Cerdán, T.; Gutiérrez-Gamboa, G.; Fernández-Novales, J.; Pérez-Álvarez, E.; Diago, M. Towards the definition of optimal grape harvest time in Grenache grapevines: Nitrogenous maturity. Sci. Hortic. 2018, 239, 9–16. [Google Scholar] [CrossRef]
- Downton, W.; Loveys, B. Compositional changes during grape berry development in relation to abscisic acid and salinity. Funct. Plant Biol. 1978, 5, 415–423. [Google Scholar] [CrossRef]
- Del-Castillo-Alonso, M.Á.; Diago, M.P.; Tomás-Las-Heras, R.; Monforte, L.; Soriano, G.; Martínez-Abaigar, J.; Núñez-Olivera, E. Effects of ambient solar UV radiation on grapevine leaf physiology and berry phenolic composition along one entire season under Mediterranean field conditions. Plant Physiol. Biochem. 2016, 109, 374–386. [Google Scholar] [CrossRef]
- Carbonell-Bejerano, P.; Diago, M.-P.; Martínez-Abaigar, J.; Martínez-Zapater, J.M.; Tardáguila, J.; Núñez-Olivera, E. Solar ultraviolet radiation is necessary to enhance grapevine fruit ripening transcriptional and phenolic responses. BMC Plant Biol. 2014, 14, 183. [Google Scholar] [CrossRef]
- Cortell, J.M.; Kennedy, J.A. Effect of shading on accumulation of flavonoid compounds in (Vitis vinifera L.) pinot noir fruit and extraction in a model system. J. Agric. Food Chem. 2006, 54, 8510–8520. [Google Scholar] [CrossRef] [PubMed]
- Pastore, C.; Zenoni, S.; Fasoli, M.; Pezzotti, M.; Tornielli, G.B.; Filippetti, I. Selective defoliation affects plant growth, fruit transcriptional ripening program and flavonoid metabolism in grapevine. BMC Plant Biol. 2013, 13, 30. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Lüscher, J.; Sánchez-Díaz, M.; Delrot, S.; Aguirreolea, J.; Pascual, I.; Gomès, E. Ultraviolet-B radiation and water deficit interact to alter flavonol and anthocyanin profiles in grapevine berries through transcriptomic regulation. Plant Cell Physiol. 2014, 55, 1925–1936. [Google Scholar] [CrossRef]
- Falginella, L.; Di Gaspero, G.; Castellarin, S.D. Expression of flavonoid genes in the red grape berry of ‘Alicante Bouschet’ varies with the histological distribution of anthocyanins and their chemical composition. Planta 2012, 236, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.; Eiras-Dias, J.; Castellarin, S.D.; Gerós, H. Berry phenolics of grapevine under challenging environments. Int. J. Mol. Sci. 2013, 14, 18711–18739. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Tian, L.; Liu, H.; Pan, Q.; Zhan, J.; Huang, W. Sugars induce anthocyanin accumulation and flavanone 3-hydroxylase expression in grape berries. Plant Growth Regul. 2009, 58, 251–260. [Google Scholar] [CrossRef]
- Dai, Z.W.; Meddar, M.; Renaud, C.; Merlin, I.; Hilbert, G.; Delrot, S.; Gomès, E. Long-term in vitro culture of grape berries and its application to assess the effects of sugar supply on anthocyanin accumulation. J. Exp. Bot. 2014, 65, 4665–4677. [Google Scholar] [CrossRef] [PubMed]
- Bobeica, N.; Poni, S.; Hilbert, G.; Renaud, C.; Gomès, E.; Delrot, S.; Dai, Z. Differential responses of sugar, organic acids and anthocyanins to source-sink modulation in Cabernet Sauvignon and Sangiovese grapevines. Front. Plant Sci. 2015, 6, 382. [Google Scholar] [CrossRef] [Green Version]
- Solfanelli, C.; Poggi, A.; Loreti, E.; Alpi, A.; Perata, P. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Del-Castillo-Alonso, M.Á.; Monforte, L.; Tomás-Las-Heras, R.; Ranieri, A.; Castagna, A.; Martínez-Abaigar, J.; Núñez-Olivera, E. Secondary metabolites and related genes in Vitis vinifera L. cv. Tempranillo grapes as influenced by ultraviolet radiation and berry development. Physiol. Plant. 2021, 173, 709–724. [Google Scholar] [CrossRef]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. Analysis of tannins in seeds and skins of Shiraz grapes throughout berry development. Aust. J. Grape Wine Res. 2003, 9, 15–27. [Google Scholar] [CrossRef]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. The effect of bunch shading on berry development and flavonoid accumulation in Shiraz grapes. Aust. J. Grape Wine Res. 2004, 10, 55–73. [Google Scholar] [CrossRef]
- Bogs, J.; Downey, M.O.; Harvey, J.S.; Ashton, A.R.; Tanner, G.J.; Robinson, S.P. Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grape berries and grapevine leaves. Plant Physiol. 2005, 139, 652–663. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A.L. Effect of maturity and vine water status on grape skin and wine flavonoids. Am. J. Enol. Vitic. 2002, 53, 268–274. [Google Scholar] [CrossRef]
- Jordan, B.R. The effects of ultraviolet-B radiation on plants: A molecular perspective. Adv. Bot. Res. 1996, 22, 97–162. [Google Scholar]
- Gil, M.; Bottini, R.; Berli, F.; Pontin, M.; Silva, M.F.; Piccoli, P. Volatile organic compounds characterized from grapevine (Vitis vinifera L. cv. Malbec) berries increase at pre-harvest and in response to UV-B radiation. Phytochemistry 2013, 96, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Halitschke, R.; Ziegler, J.; Keinänen, M.; Baldwin, I.T. Silencing of hydroperoxide lyase and allene oxide synthase reveals substrate and defense signaling crosstalk in Nicotiana attenuata. Plant J. 2004, 40, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.; Pontin, M.; Berli, F.; Bottini, R.; Piccoli, P. Metabolism of terpenes in the response of grape (Vitis vinifera L.) leaf tissues to UV-B radiation. Phytochemistry 2012, 77, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Lücker, J.; Bowen, P.; Bohlmann, J. Vitis vinifera terpenoid cyclases: Functional identification of two sesquiterpene synthase cDNAs encoding (+)-valencene synthase and (−)-germacrene D synthase and expression of mono-and sesquiterpene synthases in grapevine flowers and berries. Phytochemistry 2004, 65, 2649–2659. [Google Scholar] [CrossRef]
- Pontin, M.A.; Piccoli, P.N.; Francisco, R.; Bottini, R.; Martinez-Zapater, J.M.; Lijavetzky, D. Transcriptome changes in grapevine (Vitis vinifera L.) cv. Malbec leaves induced by ultraviolet-B radiation. BMC Plant Biol. 2010, 10, 224. [Google Scholar] [CrossRef] [Green Version]
- Berli, F.J.; Moreno, D.; Piccoli, P.; Hespanhol-viana, L.; Silva, M.F.; Bressan-smith, R.; Cavagnaro, J.B.; Bottini, R. Abscisic acid is involved in the response of grape (Vitis vinifera L.) cv. Malbec leaf tissues to ultraviolet-B radiation by enhancing ultraviolet-absorbing compounds, antioxidant enzymes and membrane sterols. Plant Cell Environ. 2010, 33, 1–10. [Google Scholar]
- Grassmann, J.; Hippeli, S.; Spitzenberger, R.; Elstner, E. The monoterpene terpinolene from the oil of Pinus mugo L. in concert with α-tocopherol and β-carotene effectively prevents oxidation of LDL. Phytomedicine 2005, 12, 416–423. [Google Scholar] [CrossRef]
- Lee, S.-J.; Umano, K.; Shibamoto, T.; Lee, K.-G. Identification of volatile components in basil (Ocimum basilicum L.) and thyme leaves (Thymus vulgaris L.) and their antioxidant properties. Food Chem. 2005, 91, 131–137. [Google Scholar] [CrossRef]
- Miao, W.; Luo, J.Q.; Liu, J.D.; Howell, K.; Zhang, P.Z. The Influence of UV on the Production of Free Terpenes in Vitis vinifera cv. Shiraz. Agronomy 2020, 10, 1431. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample (mL) | MCP (mL) | (NH4)2SO4 (mL) | Water (mL) | |
|---|---|---|---|---|
| Treatment | 0.1 | 0.3 | 0.2 | 0.4 |
| Control | 0.1 | 0 | 0.2 | 0.7 |
| Compound | ISTD ID No | RT (min) | Target Ion (m/z) | Qualifier Ions (m/z, % of Target) | Calibration Range ^ (µg/L) | CAS No. |
|---|---|---|---|---|---|---|
| d12 Hexanal | ISTD 1 | 7.78 | 64 | 48 (140.2), 46 (92.6) | - | 1219803-74-3 |
| n-Hexyl d13 Alcohol | ISTD 2 | 10.12 | 64 | 50 (45.2), 46 (44.1) | - | 16416-34-5 |
| d3 Linalool | ISTD 3 | 12.31 | 96 | 124 (25.9), 139 (10.1), 58 (16.8) | - | 1216673-02-7 |
| Hexanal | 1 | 7.85 | 44 | 41 (77.8), 56 (75.2) | 0–1048.6 | 66-25-1 |
| (E)-2-Hexenal | 2 | 9.23 | 41 | 55 (74.4), 39 (59.5) | 0–1517.1 | 6728-26-3 |
| 1-Hexanol | 2 | 10.26 | 56 | 43 (64.5), 55 (51.3) | 0–824.1 | 111-27-3 |
| (E)-3-Hexen-1-ol | 2 | 10.33 | 67 | 82 (58.1), 100 (3.8) | 0–23.4 | 928-97-2 |
| (Z)-3-Hexen-1-ol | 2 | 10.53 | 41 | 67 (78.2), 55 (38.8) | 0–265.4 | 928-96-1 |
| (E)-2-Hexen-1-ol | 2 | 10.70 | 57 | 41 (50), 39 (20.5) | 0–513.3 | 928-95-0 |
| Linalool | 3 | 12.35 | 93 | 12 (28.0), 136 (8.8) | 0–8.6 | 78-70-6 |
| Citronellol | 3 | 15.44 | 138 | 82 (468.2), 95 (397.3), 109 (138.2) | 0–8.2 | 7540-51-4 |
| α-terpineol | 3 | 14.59 | 93 | 121 (75.8), 136 (60.9), 81 (61.36) | 0–6.3 | 10482-56-1 |
| Nerol | 3 | 16.06 | 68 | 123 (28.9), 139 (18.1), 136 (11.4) | 0–7.3 | 106-25-2 |
| Geraniol | 3 | 16.88 | 84 | 93 (122.3), 123 (98.9) | 0–13.3 | 106-24-1 |
| Vintage | Treatment | °Brix | TA (g/L) | pH | Vine Yield (kg) | Pruning Weight (kg) | Ravaz Index |
|---|---|---|---|---|---|---|---|
| 2015–2016 | SC | a 20.5 | 8.6 | 3.53 | 2.58 | 0.82 | 3.38 |
| LR | b 21.6 | 8.6 | 3.58 | 2.47 | 0.66 | 4.28 | |
| PETG | b 21.6 | 8.2 | 3.59 | 2.31 | 0.72 | 3.32 | |
| p-value | 0.028 | n.s | n.s | n.s | n.s | n.s | |
| 2016–2017 | SC | 17.1 | b 11.4 | 3.65 | 3.43 | 0.62 | b 5.55 |
| LR | 18.2 | a 10.4 | 3.76 | 1.60 | 1.21 | a 1.98 | |
| PETG | 16.4 | a 10.5 | 3.62 | 2.97 | 0.71 | b 4.23 | |
| p-value | n.s | 0.036 | n.s | n.s | n.s | 0.020 |
| Rainfall (mm) | Rad (MJm2) | Temperature max (°C) | Temperature min (°C) | Average Temperature (°C) | ||
|---|---|---|---|---|---|---|
| Average values for 1971–2000 | January | 42 | 678.9 | 22.6 | 11.4 | 17 |
| February | 39 | 526.4 | 21.7 | 11 | 16.3 | |
| March | 54 | 437.1 | 20.1 | 9.9 | 15 | |
| April | 54 | 291 | 17.5 | 6.7 | 12.2 | |
| Total | 189 | 1933.4 | / | / | / | |
| 2016 | January | 107 | 578.52 | 22.6 | 11.4 | 17 |
| February | 24 | 600.15 | 21.7 | 11 | 16.3 | |
| March | 34 | 460.04 | 20.1 | 9.9 | 15 | |
| April | 10 | 325.74 | 17.5 | 6.7 | 12.2 | |
| Total | 175 | 1964 | / | / | / | |
| 2017 | January | 42 | 705.5 | 23.2 | 11.3 | 17.2 |
| February | 3 | 550.4 | 23 | 11.4 | 17.2 | |
| March | 73 | 380.2 | 19.2 | 10.6 | 14.9 | |
| April | 123 | 260 | 16.5 | 8.1 | 12.3 | |
| Total | 241 | 1896.1 | / | / | / |
| Amino Acid (µM) | Treatment | p-Value | Treatment | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| SC | LR | PETG | SC | LR | PETG | |||
| α-ketoglutarate | ||||||||
| Pro | a 1827 | b 2474 | ab 2222 | 0.025 | a 11.2% | b 16.1% | ab 15.6% | 0.007 |
| Arg | 6225 | 5949 | 5444 | n.s | 38.2% | 39.5% | 38.1% | n.s |
| Glu | 246 | 211 | 222 | n.s | 1.5% | 1.4% | 1.6% | n.s |
| Gln | 1855 | 856 | 866 | n.s | 11.4% | 5.7% | 6.1% | n.s |
| His | 283 | 217 | 253 | n.s | b 1.7% | a 1.4% | b 1.8% | 0.011 |
| Shikimate (aromatic) | ||||||||
| Phe | 443 | 341 | 315 | n.s | 2.7% | 2.3% | 2.2% | n.s |
| Trp | 144 | 121 | 123 | n.s | 0.9% | 0.8% | 0.9% | n.s |
| Tyr | 29 | 20 | 22 | n.s | 0.2% | 0.1% | 0.2% | n.s |
| Pyruvate | ||||||||
| Leu | 515 | 444 | 451 | n.s | 3.2% | 2.9% | 3.2% | n.s |
| Val | 332 | 289 | 284 | n.s | 2.0% | 1.9% | 2.0% | n.s |
| Ala | 1647 | 1489 | 1482 | n.s | 10.1% | 9.9% | 10.4% | n.s |
| Aspartate | ||||||||
| Asp | 281 | 261 | 252 | n.s | 1.7% | 1.7% | 1.8% | n.s |
| Asn | 101 | 52 | 67 | n.s | 0.6% | 0.3% | 0.5% | n.s |
| Thr | 1157 | 1166 | 1153 | n.s | a 7.1% | ab 7.7% | b 8.1% | 0.033 |
| Ile | 368 | 309 | 301 | n.s | 2.3% | 2.1% | 2.1% | n.s |
| Met | 89 | 73 | 71 | n.s | 0.5% | 0.5% | 0.5% | n.s |
| Lys | 66 | 64 | 63 | n.s | 0.4% | 0.4% | 0.4% | n.s |
| 3-phosphoglycerate | ||||||||
| Cys | N.A | N.A | N.A | N.A | N.A | N.A | N.A | N.A |
| Ser | 656 | 669 | 661 | n.s | 4.0% | 4.4% | 4.6% | n.s |
| Gly | 29 | 32 | 35 | n.s | 0.2% | 0.2% | 0.2% | n.s |
| Total | 16,305 | 15,052 | 14,282 | n.s | ||||
| Amino Acid (µM) | Treatment | p-Value | Treatment | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| SC | LR | PETG | SC | LR | PETG | |||
| α-ketoglutarate | ||||||||
| Pro | 516 | 1332 | 748 | n.s | 4.1% | 8.1% | 6.1% | n.s |
| Arg | 3256 | 2854 | 3349 | n.s | 25.8% | 17.4% | 27.2% | n.s |
| Glu | 276 | 389 | 262 | n.s | 2.2% | 2.4% | 2.1% | n.s |
| Gln | 2180 | 3419 | 2261 | n.s | 17.2% | 20.8% | 18.4% | n.s |
| His | 117 | 192 | 148 | n.s | a 0.9% | b 1.2% | b 1.2% | 0.009 |
| Shikimate (aromatic) | ||||||||
| Phe | 477 | 582 | 465 | n.s | 3.8% | 3.5% | 3.8% | n.s |
| Trp | a 69 | b 155 | ab 114 | 0.033 | 0.5% | 0.9% | 0.9% | n.s |
| Tyr | a 52 | b 77 | a 52 | 0.036 | a 0.4% | b 0.5% | a 0.4% | 0.045 |
| Pyruvate | ||||||||
| Leu | a 397 | b 577 | a 372 | 0.032 | 3.1% | 3.5% | 3.0% | n.s |
| Val | c 776 | b 526 | a 272 | 0.006 | c 6.1% | b 3.2% | a 2.2% | <0.001 |
| Ala | 1895 | 2749 | 1612 | n.s | b 15.0% | b 16.7% | a 13.1% | 0.023 |
| Aspartate | ||||||||
| Asp | 218 | 242 | 242 | n.s | ab 1.7% | a 1.5% | b 2.0% | 0.005 |
| Asn | 67 | 97 | 65 | n.s | 0.5% | 0.6% | 0.5% | n.s |
| Thr | 1061 | 1375 | 1199 | n.s | 8.4% | 8.4% | 9.7% | n.s |
| Ile | a 309 | b 460 | a 304 | 0.029 | 2.4% | 2.8% | 2.5% | n.s |
| Met | 169 | 173 | 40 | n.s | 1.3% | 1.1% | 0.3% | n.s |
| Lys | 31 | 50 | 40 | n.s | a 0.2% | b 0.3% | b 0.3% | 0.042 |
| 3-phosphoglycerate | ||||||||
| Cys | 6 | 10 | 12 | n.s | 0.1% | 0.1% | 0.1% | n.s |
| Ser | a 741 | b 1113 | a 728 | 0.030 | a 5.9% | b 6.8% | a 5.9% | 0.015 |
| Gly | a 30 | b 56 | a 28 | 0.033 | a 0.2% | b 0.3% | a 0.2% | 0.026 |
| Total | 12,646 | 16,429 | 12,314 | n.s | ||||
| Vintage | Treatments | C6 Aldehydes (µg/L) | C6 Alcohols (µg/L) | Free Monoterpenes (µg/L) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hexanal | (E)-2-Hexenal | Hexanol | (E)-3-Hexenol | (Z)-3-Hexenol | (E)-2-Hexenol | Linalool | α-Terpineol | Citronellol | Nerol | Geraniol | ||
| 2015–2016 | SC | 34.7 | 66.4 | 749.7 | 11.9 | 38.4 | 401.3 | 1.7 | 1.6 | 1.2 | 4.7 | 17.6 |
| LR | 44.5 | 67.8 | 671.0 | 11.7 | 43.1 | 404.5 | 1.7 | 1.4 | 1.0 | 4.8 | 16.9 | |
| PETG | 33.8 | 71.1 | 754.3 | 11.6 | 41.5 | 446.2 | 1.7 | 1.6 | 1.0 | 5.1 | 18.2 | |
| p-value | n.s | n.s | n.s | n.s | n.s | n.s | n.s | n.s | n.s | n.s | n.s | |
| 2016–2017 | SC | 35.0 | 35.1 | 1118.2 | 17.4 | b 76.6 | b 148.9 | 1.8 | b 1.4 | 1.0 | a 4.8 | 12.7 |
| LR | 43.7 | 51.9 | 1064.8 | 15.7 | a 45.1 | a 11.7 | 1.8 | b 1.3 | 1.2 | b 8.5 | 13.3 | |
| PETG | 31.4 | 33.9 | 1062.2 | 14.9 | b 89.5 | b 156.1 | 1.7 | a 1.1 | 1.0 | a 4.4 | 13.8 | |
| p-value | n.s | n.s | n.s | n.s | 0.042 | <0.001 | n.s | 0.012 | n.s | <0.001 | n.s | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, M.; Jordan, B.; Creasy, G.; Zhu, Y.-F. UV-B Radiation Induced the Changes in the Amount of Amino Acids, Phenolics and Aroma Compounds in Vitis vinifera cv. Pinot Noir Berry under Field Conditions. Foods 2023, 12, 2350. https://doi.org/10.3390/foods12122350
Sun M, Jordan B, Creasy G, Zhu Y-F. UV-B Radiation Induced the Changes in the Amount of Amino Acids, Phenolics and Aroma Compounds in Vitis vinifera cv. Pinot Noir Berry under Field Conditions. Foods. 2023; 12(12):2350. https://doi.org/10.3390/foods12122350
Chicago/Turabian StyleSun, Meng, Brian Jordan, Glen Creasy, and Yi-Fan Zhu. 2023. "UV-B Radiation Induced the Changes in the Amount of Amino Acids, Phenolics and Aroma Compounds in Vitis vinifera cv. Pinot Noir Berry under Field Conditions" Foods 12, no. 12: 2350. https://doi.org/10.3390/foods12122350