
Food Allergens of Plant Origin

Abstract
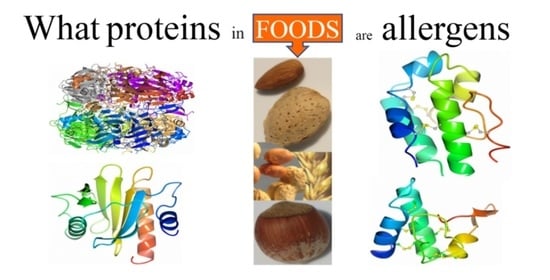
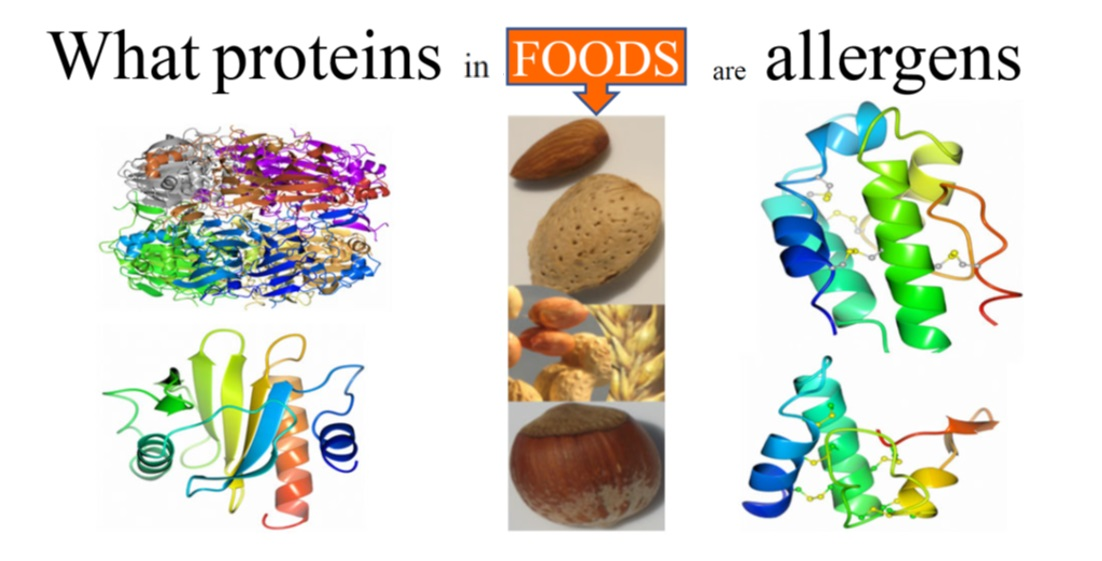
:
1. Introduction
2. Food Allergens
3. IgE Epitopes
4. Thermostability
5. Resistance to Digest
6. Cross-Reactivity
7. Summary and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sicherer, S.H.; Sampson, H.A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 2018, 141, 41–58. [Google Scholar] [CrossRef]
- Gupta, R.S.; Springston, E.E.; Warrier, M.R.; Smith, B.; Kumar, R.; Pongracic, J.; Holl, J.L. The prevalence, severity, and distribution of childhood food allergy in the United States. Pediatrics 2011, 128, e9–e17. [Google Scholar] [CrossRef] [PubMed]
- Spolidoro, G.C.I.; Amera, Y.T.; Ali, M.M.; Nyassi, S.; Lisik, D.; Ioannidou, A.; Rovner, G.; Khaleva, E.; Venter, C.; van Ree, R.; et al. FREQUENCY of food allergy in Europe: An updated systematic review and meta-analysis. Allergy 2023, 78, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.J.; Thalayasingam, M.; Lee, B.W. Food allergy in Asia: How does it compare? Asia Pac. Allergy 2013, 3, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Motosue, M.S.; Bellolio, M.F.; Van Houten, H.K.; Shah, N.D.; Campbell, R.L. National trends in emergency department visits and hospitalizations for food-induced anaphylaxis in US children. Pediatr. Allergy Immunol. 2018, 29, 538–544. [Google Scholar] [CrossRef]
- Gupta, R.S.; Warren, C.M.; Smith, B.M.; Jiang, J.L.; Blumenstock, J.A.; Davis, M.M.; Schleimer, R.P.; Nadeau, K.C. Prevalence and severity of food allergies among US adults. JAMA Netw. Open 2019, 2, e185630. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Munoz-Furlong, A.; Sampson, H.A. Prevalence of peanut and tree nut allergy in the United States determined by means of a random digit dial telephone survey: A 5-year follow-up study. J. Allergy Clin. Immunol. 2003, 112, 1203–1207. [Google Scholar] [CrossRef]
- Kumfer, A.M.; Commins, S.P. Primary prevention of food allergy. Curr. Allergy Asthma Rep. 2019, 19, 7. [Google Scholar] [CrossRef]
- Bhalla, P.L.; Singh, M.B. Biotechnology-based allergy diagnosis and vaccination. Trends Biotechnol. 2008, 26, 153–161. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef]
- Fattakhova, G.; Masilamani, M.; Borrego, F.; Gilfillan, A.M.; Metcalfe, D.D.; Coligan, J.E. The high-affinity immunoglobulin-E receptor (FcepsilonRI) is endocytosed by an AP-2/clathrin-independent, dynamin-dependent mechanism. Traffic 2006, 7, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Krempski, J.W.; Dant, C.; Nadeau, K.C. The origins of allergy from a systems approach. Ann. Allergy Asthma Immunol. 2020, 125, 507–516. [Google Scholar] [CrossRef]
- IUIS/WHO-Allergen-Nomenclature-Subcommittee. Allergen nomenclature. Bull. World Health Organ. 1994, 72, 797–806. [Google Scholar]
- Osborne, T.B. The Vegetable Proteins, 2nd ed.; Green and Co.: London, UK, 1924; p. 440. [Google Scholar]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, K.; Job, C.; Groot, S.P.; Puype, M.; Demol, H.; Vandekerckhove, J.; Job, D. Proteomic analysis of arabidopsis seed germination and priming. Plant Physiol. 2001, 126, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Mooney, B.P.; Krishnan, H.B.; Thelen, J.J. High-throughput peptide mass fingerprinting of soybean seed proteins: Automated workflow and utility of UniGene expressed sequence tag databases for protein identification. Phytochemistry 2004, 65, 1733–1744. [Google Scholar] [CrossRef]
- Salminen, T.A.; Blomqvist, K.; Edqvist, J. Lipid transfer proteins: Classification, nomenclature, structure, and function. Planta 2016, 244, 971–997. [Google Scholar] [CrossRef]
- Kader, J.C.; Julienne, M.; Vergnolle, C. Purification and characterization of a spinach-leaf protein capable of transferring phospholipids from liposomes to mitochondria or chloroplasts. Eur. J Biochem. 1984, 139, 411–416. [Google Scholar] [CrossRef]
- Sun, J.Y.; Gaudet, D.A.; Lu, Z.X.; Frick, M.; Puchalski, B.; Laroche, A. Characterization and antifungal properties of wheat nonspecific lipid transfer proteins. Mol. Plant Microbe Interact. 2008, 21, 346–360. [Google Scholar] [CrossRef]
- Odintsova, T.I.; Slezina, M.P.; Istomina, E.A.; Korostyleva, T.V.; Kovtun, A.S.; Kasianov, A.S.; Shcherbakova, L.A.; Kudryavtsev, A.M. Non-specific lipid transfer proteins in Triticum kiharae Dorof. et Migush.: Identification, characterization and expression profiling in response to pathogens and resistance inducers. Pathogens 2019, 8, 221. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Wu, J.H.; Ng, T.B.; Ye, X.Y.; Rao, P.F. A non-specific lipid transfer protein with antifungal and antibacterial activities from the mung bean. Peptides 2004, 25, 1235–1242. [Google Scholar] [CrossRef]
- Wang, N.J.; Lee, C.C.; Cheng, C.S.; Lo, W.C.; Yang, Y.F.; Chen, M.N.; Lyu, P.C. Construction and analysis of a plant non-specific lipid transfer protein database (nsLTPDB). BMC Genom. 2012, 13 (Suppl. S1), S9. [Google Scholar] [CrossRef]
- Krause, S.; Reese, G.; Randow, S.; Zennaro, D.; Quaratino, D.; Palazzo, P.; Ciardiello, M.A.; Petersen, A.; Becker, W.M.; Mari, A. Lipid transfer protein (Ara h 9) as a new peanut allergen relevant for a Mediterranean allergic population. J. Allergy Clin. Immunol. 2009, 124, 771–778.e775. [Google Scholar] [CrossRef]
- WHO/IUIS-Allergen-Nomenclature-Sub-Committee. Allergen Nomenclature. Available online: http://www.allergen.org/ (accessed on 14 April 2023).
- Sanchez-Monge, R.; Blanco, C.; Lopez-Torrejon, G.; Cumplido, J.; Recas, M.; Figueroa, J.; Carrillo, T.; Salcedo, G. Differential allergen sensitization patterns in chestnut allergy with or without associated latex-fruit syndrome. J. Allergy Clin. Immunol. 2006, 118, 705–710. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Vieths, S.; Pravettoni, V.; Farioli, L.; Trambaioli, C.; Fortunato, D.; Luttkopf, D.; Calamari, M.; Ansaloni, R.; Scibilia, J.; et al. Identification of hazelnut major allergens in sensitive patients with positive double-blind, placebo-controlled food challenge results. J. Allergy Clin. Immunol. 2002, 109, 563–570. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Farioli, L.; Pravettoni, V.; Robino, A.M.; Scibilia, J.; Fortunato, D.; Conti, A.; Borgonovo, L.; Bengtsson, A.; Ortolani, C. Lipid transfer protein and vicilin are important walnut allergens in patients not allergic to pollen. J. Allergy Clin. Immunol. 2004, 114, 908–914. [Google Scholar] [CrossRef]
- Sander, I.; Rozynek, P.; Rihs, H.P.; van Kampen, V.; Chew, F.T.; Lee, W.S.; Kotschy-Lang, N.; Merget, R.; Bruning, T.; Raulf-Heimsoth, M. Multiple wheat flour allergens and cross-reactive carbohydrate determinants bind IgE in baker’s asthma. Allergy 2011, 66, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Offermann, L.R.; Bublin, M.; Perdue, M.L.; Pfeifer, S.; Dubiela, P.; Borowski, T.; Chruszcz, M.; Hoffmann-Sommergruber, K. Structural and functional characterization of the hazelnut allergen Cor a 8. J. Agric. Food Chem. 2015, 63, 9150–9158. [Google Scholar] [CrossRef] [PubMed]
- Charvolin, D.; Douliez, J.P.; Marion, D.; Cohen-Addad, C.; Pebay-Peyroula, E. The crystal structure of a wheat nonspecific lipid transfer protein (ns-LTP1) complexed with two molecules of phospholipid at 2.1 A resolution. Eur. J. Biochem. 1999, 264, 562–568. [Google Scholar] [CrossRef]
- Tassin-Moindrot, S.; Caille, A.; Douliez, J.P.; Marion, D.; Vovelle, F. The wide binding properties of a wheat nonspecific lipid transfer protein. Solution structure of a complex with prostaglandin B2. Eur. J. Biochem. 2000, 267, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- McNicholas, S.; Potterton, E.; Wilson, K.S.; Noble, M.E. Presenting your structures: The CCP4mg molecular-graphics software. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 386–394. [Google Scholar] [CrossRef]
- Sy, D.; Le Gravier, Y.; Goodfellow, J.; Vovelle, F. Protein stability and plasticity of the hydrophobic cavity in wheat ns-LTP. J. Biomol. Struct. Dyn. 2003, 21, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Samuel, D.; Liu, Y.J.; Cheng, C.S.; Lyu, P.C. Solution structure of plant nonspecific lipid transfer protein-2 from rice (Oryza sativa). J. Biol. Chem. 2002, 277, 35267–35273. [Google Scholar] [CrossRef]
- Youle, R.J.; Huang, A.H.C. Occurrence of low-molecular weight and high cysteine containing albumin storage proteins in oilseeds of diverse species. Am. J. Bot. 1981, 68, 44–48. [Google Scholar] [CrossRef]
- Utsumi, S. Plant food protein engineering. Adv. Food Nutr. Res. 1992, 36, 89–208. [Google Scholar]
- Shewry, P.R. Plant storage proteins. Biol. Rev. Camb. Philos. Soc. 1995, 70, 375–426. [Google Scholar] [CrossRef]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar]
- Candido Ede, S.; Pinto, M.F.; Pelegrini, P.B.; Lima, T.B.; Silva, O.N.; Pogue, R.; Grossi-de-Sa, M.F.; Franco, O.L. Plant storage proteins with antimicrobial activity: Novel insights into plant defense mechanisms. FASEB J. 2011, 25, 3290–3305. [Google Scholar] [CrossRef]
- Odintsova, T.I.; Rogozhin, E.A.; Sklyar, I.V.; Musolyamov, A.K.; Kudryavtsev, A.M.; Pukhalsky, V.A.; Smirnov, A.N.; Grishin, E.V.; Egorov, T.A. Antifungal activity of storage 2S albumins from seeds of the invasive weed dandelion Taraxacum officinale Wigg. Protein Pept. Lett. 2010, 17, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Pelegrini, P.B.; Noronha, E.F.; Muniz, M.A.; Vasconcelos, I.M.; Chiarello, M.D.; Oliveira, J.T.; Franco, O.L. An antifungal peptide from passion fruit (Passiflora edulis) seeds with similarities to 2S albumin proteins. Biochim. Biophys. Acta 2006, 1764, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Kleber-Janke, T.; Crameri, R.; Appenzeller, U.; Schlaak, M.; Becker, W.M. Selective cloning of peanut allergens, including profilin and 2S albumins, by phage display technology. Int. Arch. Allergy Immunol. 1999, 119, 265–274. [Google Scholar] [CrossRef]
- Klemans, R.J.; Knol, E.F.; Michelsen-Huisman, A.; Pasmans, S.G.; de Kruijf-Broekman, W.; Bruijnzeel-Koomen, C.A.; van Hoffen, E.; Knulst, A.C. Components in soy allergy diagnostics: Gly m 2S albumin has the best diagnostic value in adults. Allergy 2013, 68, 1396–1402. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Farioli, L.; Pravettoni, V.; Ispano, M.; Conti, A.; Ansaloni, R.; Rotondo, F.; Incorvaia, C.; Bengtsson, A.; Rivolta, F.; et al. Sensitization to the major allergen of Brazil nut is correlated with the clinical expression of allergy. J. Allergy Clin. Immunol. 1998, 102, 1021–1027. [Google Scholar] [CrossRef]
- Robotham, J.M.; Wang, F.; Seamon, V.; Teuber, S.S.; Sathe, S.K.; Sampson, H.A.; Beyer, K.; Seavy, M.; Roux, K.H. Ana o 3, an important cashew nut (Anacardium occidentale L.) allergen of the 2S albumin family. J. Allergy Clin. Immunol. 2005, 115, 1284–1290. [Google Scholar] [CrossRef] [PubMed]
- Garino, C.; Zuidmeer, L.; Marsh, J.; Lovegrove, A.; Morati, M.; Versteeg, S.; Schilte, P.; Shewry, P.; Arlorio, M.; van Ree, R. Isolation, cloning, and characterization of the 2S albumin: A new allergen from hazelnut. Mol. Nutr. Food Res. 2010, 54, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.M.; Irsigler, A.; Dhanarajan, P.; Ayuso, R.; Bardina, L.; Sampson, H.A.; Roux, K.H.; Sathe, S.K. Cloning and characterization of 2S albumin, Car i 1, a major allergen in pecan. J. Agric. Food Chem. 2011, 59, 4130–4139. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K.; Bardina, L.; Grishina, G.; Beyer, K.; Sampson, H.A. Identification of two pistachio allergens, Pis v 1 and Pis v 2, belonging to the 2S albumin and 11S globulin family. Clin. Exp. Allergy 2009, 39, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B.; Crespo, J.F.; Maleki, S.J.; Rodriguez, J.; Novak, N. Pin p 1 is a major allergen in pine nut and the first food allergen described in the plant group of gymnosperms. Food Chem. 2016, 210, 70–77. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Varin, E.; Farioli, L.; Pravettoni, V.; Ortolani, C.; Trambaioli, C.; Fortunato, D.; Giuffrida, M.G.; Rivolta, F.; Robino, A.; et al. The major allergen of sesame seeds (Sesamum indicum) is a 2S albumin. J. Chromatogr. B Biomed. Sci. Appl. 2001, 756, 85–93. [Google Scholar] [CrossRef]
- Beyer, K.; Bardina, L.; Grishina, G.; Sampson, H.A. Identification of sesame seed allergens by 2-dimensional proteomics and Edman sequencing: Seed storage proteins as common food allergens. J. Allergy Clin. Immunol. 2002, 110, 154–159. [Google Scholar] [CrossRef]
- Teuber, S.S.; Dandekar, A.M.; Peterson, W.R.; Sellers, C.L. Cloning and sequencing of a gene encoding a 2S albumin seed storage protein precursor from English walnut (Juglans regia), a major food allergen. J. Allergy Clin. Immunol. 1998, 101, 807–814. [Google Scholar] [CrossRef]
- Pantoja-Uceda, D.; Bruix, M.; Gimenez-Gallego, G.; Rico, M.; Santoro, J. Solution structure of RicC3, a 2S albumin storage protein from Ricinus communis. Biochemistry 2003, 42, 13839–13847. [Google Scholar] [CrossRef] [PubMed]
- Pantoja-Uceda, D.; Palomares, O.; Bruix, M.; Villalba, M.; Rodriguez, R.; Rico, M.; Santoro, J. Solution structure and stability against digestion of rproBnIb, a recombinant 2S albumin from rapeseed: Relationship to its allergenic properties. Biochemistry 2004, 43, 16036–16045. [Google Scholar] [CrossRef] [PubMed]
- Rundqvist, L.; Tengel, T.; Zdunek, J.; Bjorn, E.; Schleucher, J.; Alcocer, M.J.; Larsson, G. Solution structure, copper binding and backbone dynamics of recombinant Ber e 1-the major allergen from Brazil nut. PLoS ONE 2012, 7, e46435. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, K.; Schweimer, K.; Reese, G.; Randow, S.; Suhr, M.; Becker, W.M.; Vieths, S.; Rosch, P. Structure and stability of 2S albumin-type peanut allergens: Implications for the severity of peanut allergic reactions. Biochem. J. 2006, 395, 463–472. [Google Scholar] [CrossRef]
- Mueller, G.A.; Gosavi, R.A.; Pomes, A.; Wunschmann, S.; Moon, A.F.; London, R.E.; Pedersen, L.C. Ara h 2: Crystal structure and IgE binding distinguish two subpopulations of peanut allergic patients by epitope diversity. Allergy 2011, 66, 878–885. [Google Scholar] [CrossRef]
- Lane, B.G.; Bernier, F.; Dratewka-Kos, E.; Shafai, R.; Kennedy, T.D.; Pyne, C.; Munro, J.R.; Vaughan, T.; Walters, D.; Altomare, F. Homologies between members of the germin gene family in hexaploid wheat and similarities between these wheat germins and certain Physarum spherulins. J. Biol. Chem. 1991, 266, 10461–10469. [Google Scholar] [CrossRef]
- Ko, T.P.; Ng, J.D.; McPherson, A. The three-dimensional structure of canavalin from jack bean (Canavalia ensiformis). Plant Physiol. 1993, 101, 729–744. [Google Scholar] [CrossRef]
- Lawrence, M.C.; Izard, T.; Beuchat, M.; Blagrove, R.J.; Colman, P.M. Structure of phaseolin at 2.2 A resolution. Implications for a common vicilin/legumin structure and the genetic engineering of seed storage proteins. J. Mol. Biol. 1994, 238, 748–776. [Google Scholar] [CrossRef]
- Dunwell, J.M. Cupins: A new superfamily of functionally diverse proteins that include germins and plant storage proteins. Biotechnol. Genet. Eng. Rev. 1998, 15, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Dunwell, J.M.; Purvis, A.; Khuri, S. Cupins: The most functionally diverse protein superfamily? Phytochemistry 2004, 65, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.; Schlichting, B.; Felgendreher, M.; Schonheit, P. Cupin-type phosphoglucose isomerases (Cupin-PGIs) constitute a novel metal-dependent PGI family representing a convergent line of PGI evolution. J. Bacteriol. 2005, 187, 1621–1631. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Wang, Y.; Chen, Y.W.; Fu, T.J.; Kothary, M.H.; McHugh, T.H.; Zhang, Y. Crystal structure of the Korean pine (Pinus koraiensis) 7S seed storage protein with copper ligands. J. Agric. Food Chem. 2014, 62, 222–228. [Google Scholar] [CrossRef]
- Mills, E.N.; Jenkins, J.; Marigheto, N.; Belton, P.S.; Gunning, A.P.; Morris, V.J. Allergens of the cupin superfamily. Biochem. Soc. Trans. 2002, 30, 925–929. [Google Scholar] [CrossRef]
- Rabjohn, P.; Helm, E.M.; Stanley, J.S.; West, C.M.; Sampson, H.A.; Burks, A.W.; Bannon, G.A. Molecular cloning and epitope analysis of the peanut allergen Ara h 3. J. Clin. Investig. 1999, 103, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Holzhauser, T.; Wackermann, O.; Ballmer-Weber, B.K.; Bindslev-Jensen, C.; Scibilia, J.; Perono-Garoffo, L.; Utsumi, S.; Poulsen, L.K.; Vieths, S. Soybean (Glycine max) allergy in Europe: Gly m 5 (beta-conglycinin) and Gly m 6 (glycinin) are potential diagnostic markers for severe allergic reactions to soy. J. Allergy Clin. Immunol. 2009, 123, 452–458. [Google Scholar] [CrossRef]
- Willison, L.N.; Tripathi, P.; Sharma, G.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Cloning, expression and patient IgE reactivity of recombinant Pru du 6, an 11S globulin from almond. Int. Arch. Allergy Immunol. 2011, 156, 267–281. [Google Scholar] [CrossRef]
- Wang, F.; Robotham, J.M.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Ana o 2, a major cashew (Anacardium occidentale L.) nut allergen of the legumin family. Int. Arch. Allergy Immunol. 2003, 132, 27–39. [Google Scholar] [CrossRef]
- Beyer, K.; Grishina, G.; Bardina, L.; Grishin, A.; Sampson, H.A. Identification of an 11S globulin as a major hazelnut food allergen in hazelnut-induced systemic reactions. J. Allergy Clin. Immunol. 2002, 110, 517–523. [Google Scholar] [CrossRef]
- Kabasser, S.; Pratap, K.; Kamath, S.; Taki, A.C.; Dang, T.; Koplin, J.; Perrett, K.; Hummel, K.; Radauer, C.; Breiteneder, H.; et al. Identification of vicilin, legumin and antimicrobial peptide 2a as macadamia nut allergens. Food Chem. 2022, 370, 131028. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bhardwaj, S.R.; Lyu, S.C.; Chinthrajah, S.; Nadeau, K.C.; Li, C. Expression, purification, characterization, and patient IgE reactivity of new macadamia nut iso-allergen. Protein Expr. Purif. 2022, 203, 106211. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.M.; Irsigler, A.; Dhanarajan, P.; Ayuso, R.; Bardina, L.; Sampson, H.A.; Roux, K.H.; Sathe, S.K. Cloning and characterization of an 11S legumin, Car i 4, a major allergen in pecan. J. Agric. Food Chem. 2011, 59, 9542–9552. [Google Scholar] [CrossRef]
- Beyer, K.; Grishina, G.; Bardina, L.; Sampson, H.A. Identification of 2 new sesame seed allergens: Ses i 6 and Ses i 7. J. Allergy Clin. Immunol. 2007, 119, 1554–1556. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Du, W.X.; Fan, Y.; Yi, J.; Lyu, S.C.; Nadeau, K.C.; Thomas, A.L.; McHugh, T. Purification and characterization of a black walnut (Juglans nigra) allergen, Jug n 4. J. Agric. Food Chem. 2017, 65, 454–462. [Google Scholar] [CrossRef]
- Wallowitz, M.; Peterson, W.R.; Uratsu, S.; Comstock, S.S.; Dandekar, A.M.; Teuber, S.S. Jug r 4, a legumin group food allergen from walnut (Juglans regia Cv. Chandler). J. Agric. Food Chem. 2006, 54, 8369–8375. [Google Scholar] [CrossRef] [PubMed]
- Adachi, M.; Kanamori, J.; Masuda, T.; Yagasaki, K.; Kitamura, K.; Mikami, B.; Utsumi, S. Crystal structure of soybean 11S globulin: Glycinin A3B4 homohexamer. Proc. Natl. Acad. Sci. USA 2003, 100, 7395–7400. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.-S.; Lin, X.-D.; Zhang, Y.-S.; Wang, L.; Wu, K.; Huang, S.-Z. Isolation of peanut genes encoding arachins and conglutins by expressed sequence tags. Plant Sci. 2005, 169, 439–445. [Google Scholar] [CrossRef]
- Jin, T.; Howard, A.; Zhang, Y.Z. Purification, crystallization and initial crystallographic characterization of peanut major allergen Ara h 3. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2007, 63, 848–851. [Google Scholar] [CrossRef]
- Jin, T.; Guo, F.; Chen, Y.-W.; Howard, A.; Zhang, Y.-Z. Crystal structure of Ara h 3, a major allergen in peanut. Mol. Immunol. 2009, 46, 1796–1804. [Google Scholar] [CrossRef]
- Jin, T.; Albillos, S.M.; Guo, F.; Howard, A.; Fu, T.J.; Kothary, M.H.; Zhang, Y.Z. Crystal structure of prunin-1, a major component of the almond (Prunus dulcis) allergen amandin. J. Agric. Food Chem. 2009, 57, 8643–8651. [Google Scholar] [CrossRef]
- Vajravijayan, S.; Nandhagopal, N.; Gunasekaran, K. Crystal structure determination and analysis of 11S coconut allergen: Cocosin. Mol. Immunol. 2017, 92, 132–135. [Google Scholar] [CrossRef]
- Jin, T.; Wang, C.; Zhang, C.; Wang, Y.; Chen, Y.W.; Guo, F.; Howard, A.; Cao, M.J.; Fu, T.J.; McHugh, T.H.; et al. Crystal structure of cocosin, a potential food allergen from coconut (Cocos nucifera). J. Agric. Food Chem. 2017, 65, 7560–7568. [Google Scholar] [CrossRef] [PubMed]
- Gatehouse, J.A.; Lycett, G.W.; Croy, R.R.; Boulter, D. The post-translational proteolysis of the subunits of vicilin from pea (Pisum sativum L.). Biochem. J. 1982, 207, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Torrejon, G.; Salcedo, G.; Martin-Esteban, M.; Diaz-Perales, A.; Pascual, C.Y.; Sanchez-Monge, R. Len c 1, a major allergen and vicilin from lentil seeds: Protein isolation and cDNA cloning. J. Allergy Clin. Immunol. 2003, 112, 1208–1215. [Google Scholar] [CrossRef] [PubMed]
- Burks, A.W.; Cockrell, G.; Stanley, J.S.; Helm, R.M.; Bannon, G.A. Recombinant peanut allergen Ara h I expression and IgE binding in patients with peanut hypersensitivity. J. Clin. Investig. 1995, 96, 1715–1721. [Google Scholar] [CrossRef]
- Che, H.L.; Zhang, Y.Z.; Lyu, S.C.; Nadeau, K.C.; McHugh, T. Identification of almond (Prunus dulcis) vicilin as a food allergen. J. Agric. Food Chem. 2019, 67, 425–432. [Google Scholar] [CrossRef]
- Wang, F.; Robotham, J.M.; Teuber, S.S.; Tawde, P.; Sathe, S.K.; Roux, K.H. Ana o 1, a cashew (Anacardium occidental) allergen of the vicilin seed storage protein family. J. Allergy Clin. Immunol. 2002, 110, 160–166. [Google Scholar] [CrossRef]
- Lauer, I.; Foetisch, K.; Kolarich, D.; Ballmer-Weber, B.K.; Conti, A.; Altmann, F.; Vieths, S.; Scheurer, S. Hazelnut (Corylus avellana) vicilin Cor a 11: Molecular characterization of a glycoprotein and its allergenic activity. Biochem. J. 2004, 383, 327–334. [Google Scholar] [CrossRef]
- Mattsson, L.; Holmqvist, M.; Porsch, H.; Larsson, H.; Pontoppidan, B.; Valcour, A.; Lidholm, J. A new vicilin-like allergen in hazelnut giving rise to a spectrum of IgE-binding low-molecular-weight N-terminal fragments. Clin. Exp. Allergy 2022, 52, 1208–1212. [Google Scholar] [CrossRef]
- Zhang, Y.; Lee, B.; Du, W.X.; Lyu, S.C.; Nadeau, K.C.; Grauke, L.J.; Zhang, Y.; Wang, S.; Fan, Y.; Yi, J.; et al. Identification and characterization of a new pecan [Carya illinoinensis (Wangenh.) K. Koch] allergen, Car i 2. J. Agric. Food Chem. 2016, 64, 4146–4151. [Google Scholar] [CrossRef] [PubMed]
- Willison, L.N.; Tawde, P.; Robotham, J.M.; Penney, R.M.t.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Pistachio vicilin, Pis v 3, is immunoglobulin E-reactive and cross-reacts with the homologous cashew allergen, Ana o 1. Clin. Exp. Allergy 2008, 38, 1229–1238. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Du, W.X.; Fan, Y.T.; Yi, J.; Lyu, S.C.; Nadeau, K.C.; McHugh, T.H. Identification, characterization, and initial epitope mapping of pine nut allergen Pin k 2. Food Res. Int. 2016, 90, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Albillos, S.M.; Chen, Y.W.; Kothary, M.H.; Fu, T.J.; Zhang, Y.Z. Purification and characterization of the 7S vicilin from Korean pine (Pinus koraiensis). J. Agric. Food Chem. 2008, 56, 8159–8165. [Google Scholar] [CrossRef] [PubMed]
- Teuber, S.S.; Jarvis, K.C.; Dandekar, A.M.; Peterson, W.R.; Ansari, A.A. Identification and cloning of a complementary DNA encoding a vicilin-like proprotein, jug r 2, from english walnut kernel (Juglans regia), a major food allergen. J. Allergy Clin. Immunol. 1999, 104, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Monge, R.; Lopez-Torrejon, G.; Pascual, C.Y.; Varela, J.; Martin-Esteban, M.; Salcedo, G. Vicilin and convicilin are potential major allergens from pea. Clin. Exp. Allergy 2004, 34, 1747–1753. [Google Scholar] [CrossRef]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gislason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Wichers, H.J.; De Beijer, T.; Savelkoul, H.F.; Van Amerongen, A. The major peanut allergen Ara h 1 and its cleaved-off N-terminal peptide; possible implications for peanut allergen detection. J. Agric. Food Chem. 2004, 52, 4903–4907. [Google Scholar] [CrossRef]
- Dubiela, P.; Kabasser, S.; Smargiasso, N.; Geiselhart, S.; Bublin, M.; Hafner, C.; Mazzucchelli, G.; Hoffmann-Sommergruber, K. Jug r 6 is the allergenic vicilin present in walnut responsible for IgE cross-reactivities to other tree nuts and seeds. Sci. Rep. 2018, 8, 11366. [Google Scholar] [CrossRef]
- Goggin, D.E.; Mir, G.; Smith, W.B.; Stuckey, M.; Smith, P.M. Proteomic analysis of lupin seed proteins to identify conglutin Beta as an allergen, Lup an 1. J. Agric. Food Chem. 2008, 56, 6370–6377. [Google Scholar] [CrossRef]
- Marcus, J.P.; Green, J.L.; Goulter, K.C.; Manners, J.M. A family of antimicrobial peptides is produced by processing of a 7S globulin protein in Macadamia integrifolia kernels. Plant J. 1999, 19, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Foo, A.C.Y.; Nesbit, J.B.; Gipson, S.A.Y.; Cheng, H.; Bushel, P.; DeRose, E.F.; Schein, C.H.; Teuber, S.S.; Hurlburt, B.K.; Maleki, S.J.; et al. Structure, immunogenicity, and IgE cross-reactivity among walnut and peanut vicilin-buried peptides. J. Agric. Food Chem. 2022, 70, 2389–2400. [Google Scholar] [CrossRef] [PubMed]
- Foo, A.C.Y.; Nesbit, J.B.; Gipson, S.A.Y.; DeRose, E.F.; Cheng, H.; Hurlburt, B.K.; Kulis, M.D.; Kim, E.H.; Dreskin, S.C.; Mustafa, S.; et al. Structure and IgE cross-reactivity among cashew, pistachio, walnut, and peanut vicilin-buried peptides. J. Agric. Food Chem. 2023, 71, 2990–2998. [Google Scholar] [CrossRef]
- Cabanos, C.; Urabe, H.; Tandang-Silvas, M.R.; Utsumi, S.; Mikami, B.; Maruyama, N. Crystal structure of the major peanut allergen Ara h 1. Mol. Immunol. 2011, 49, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Chruszcz, M.; Maleki, S.J.; Majorek, K.A.; Demas, M.; Bublin, M.; Solberg, R.; Hurlburt, B.K.; Ruan, S.; Mattison, C.P.; Breiteneder, H.; et al. Structural and immunologic characterization of Ara h 1, a major peanut allergen. J. Biol. Chem. 2011, 286, 39318–39327. [Google Scholar] [CrossRef]
- Shikhi, M.; Jain, A.; Salunke, D.M. Comparative study of 7S globulin from Corylus avellana and Solanum lycopersicum revealed importance of salicylic acid and Cu-binding loop in modulating their function. Biochem. Biophys. Res. Commun. 2020, 522, 127–132. [Google Scholar] [CrossRef]
- Maruyama, N.; Adachi, M.; Takahashi, K.; Yagasaki, K.; Kohno, M.; Takenaka, Y.; Okuda, E.; Nakagawa, S.; Mikami, B.; Utsumi, S. Crystal structures of recombinant and native soybean beta-conglycinin beta homotrimers. Eur. J. Biochem. 2001, 268, 3595–3604. [Google Scholar] [CrossRef]
- Che, H.; Zhang, Y.; Jiang, S.; Jin, T.; Lyu, S.C.; Nadeau, K.C.; McHugh, T. Almond (Prunus dulcis) allergen Pru du 8, the first member of a new family of food allergens. J. Agric. Food Chem. 2019, 67, 8626–8631. [Google Scholar] [CrossRef]
- Zhang, Y.; Jin, T. Almond allergens: Update and perspective on identification and characterization. J. Sci. Food Agric. 2020, 100, 4657–4663. [Google Scholar] [CrossRef]
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial peptides from plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef]
- Nolde, S.B.; Vassilevski, A.A.; Rogozhin, E.A.; Barinov, N.A.; Balashova, T.A.; Samsonova, O.V.; Baranov, Y.V.; Feofanov, A.V.; Egorov, T.A.; Arseniev, A.S.; et al. Disulfide-stabilized helical hairpin structure and activity of a novel antifungal peptide EcAMP1 from seeds of barnyard grass (Echinochloa crus-galli). J. Biol. Chem. 2011, 286, 25145–25153. [Google Scholar] [CrossRef] [PubMed]
- Valenta, R.; Duchene, M.; Pettenburger, K.; Sillaber, C.; Valent, P.; Bettelheim, P.; Breitenbach, M.; Rumpold, H.; Kraft, D.; Scheiner, O. Identification of profilin as a novel pollen allergen; IgE autoreactivity in sensitized individuals. Science 1991, 253, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Hirschwehr, R.; Valenta, R.; Ebner, C.; Ferreira, F.; Sperr, W.R.; Valent, P.; Rohac, M.; Rumpold, H.; Scheiner, O.; Kraft, D. Identification of common allergenic structures in hazel pollen and hazelnuts: A possible explanation for sensitivity to hazelnuts in patients allergic to tree pollen. J. Allergy Clin. Immunol. 1992, 90, 927–936. [Google Scholar] [CrossRef]
- Rihs, H.P.; Chen, Z.; Rueff, F.; Petersen, A.; Rozynek, P.; Heimann, H.; Baur, X. IgE binding of the recombinant allergen soybean profilin (rGly m 3) is mediated by conformational epitopes. J. Allergy Clin. Immunol. 1999, 104, 1293–1301. [Google Scholar] [CrossRef]
- Reindl, J.; Rihs, H.P.; Scheurer, S.; Wangorsch, A.; Haustein, D.; Vieths, S. IgE reactivity to profilin in pollen-sensitized subjects with adverse reactions to banana and pineapple. Int. Arch. Allergy Immunol. 2002, 128, 105–114. [Google Scholar] [CrossRef]
- Lopez-Torrejon, G.; Crespo, J.F.; Sanchez-Monge, R.; Sanchez-Jimenez, M.; Alvarez, J.; Rodriguez, J.; Salcedo, G. Allergenic reactivity of the melon profilin Cuc m 2 and its identification as major allergen. Clin. Exp. Allergy 2005, 35, 1065–1072. [Google Scholar] [CrossRef]
- Lopez-Torrejon, G.; Ibanez, M.D.; Ahrazem, O.; Sanchez-Monge, R.; Sastre, J.; Lombardero, M.; Barber, D.; Salcedo, G. Isolation, cloning and allergenic reactivity of natural profilin Cit s 2, a major orange allergen. Allergy 2005, 60, 1424–1429. [Google Scholar] [CrossRef]
- Ballmer-Weber, B.K.; Wuthrich, B.; Wangorsch, A.; Fotisch, K.; Altmann, F.; Vieths, S. Carrot allergy: Double-blinded, placebo-controlled food challenge and identification of allergens. J. Allergy Clin. Immunol. 2001, 108, 301–307. [Google Scholar] [CrossRef]
- Luttkopf, D.; Ballmer-Weber, B.K.; Wuthrich, B.; Vieths, S. Celery allergens in patients with positive double-blind placebo-controlled food challenge. J. Allergy Clin. Immunol. 2000, 106, 390–399. [Google Scholar] [CrossRef]
- Santos, A.; Van Ree, R. Profilins: Mimickers of allergy or relevant allergens? Int. Arch. Allergy Immunol. 2011, 155, 191–204. [Google Scholar] [CrossRef]
- Scala, E.; Alessandri, C.; Palazzo, P.; Pomponi, D.; Liso, M.; Bernardi, M.L.; Ferrara, R.; Zennaro, D.; Santoro, M.; Rasi, C.; et al. IgE recognition patterns of profilin, PR-10, and tropomyosin panallergens tested in 3,113 allergic patients by allergen microarray-based technology. PLoS ONE 2011, 6, e24912. [Google Scholar] [CrossRef] [PubMed]
- Scheurer, S.; Wangorsch, A.; Nerkamp, J.; Skov, P.S.; Ballmer-Weber, B.; Wuthrich, B.; Haustein, D.; Vieths, S. Cross-reactivity within the profilin panallergen family investigated by comparison of recombinant profilins from pear (Pyr c 4), cherry (Pru av 4) and celery (Api g 4) with birch pollen profilin Bet v 2. J. Chromatogr. B Biomed. Sci. Appl. 2001, 756, 315–325. [Google Scholar] [CrossRef]
- Bauer, L.; Ebner, C.; Hirschwehr, R.; Wuthrich, B.; Pichler, C.; Fritsch, R.; Scheiner, O.; Kraft, D. IgE cross-reactivity between birch pollen, mugwort pollen and celery is due to at least three distinct cross-reacting allergens: Immunoblot investigation of the birch-mugwort-celery syndrome. Clin. Exp. Allergy 1996, 26, 1161–1170. [Google Scholar] [CrossRef] [PubMed]
- Tawde, P.; Venkatesh, Y.P.; Wang, F.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Cloning and characterization of profilin (Pru du 4), a cross-reactive almond (Prunus dulcis) allergen. J. Allergy Clin. Immunol. 2006, 118, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.S.; Ballmer-Weber, B.K.; Luttkopf, D.; Skov, P.S.; Wuthrich, B.; Bindslev-Jensen, C.; Vieths, S.; Poulsen, L.K. Roasted hazelnuts—Allergenic activity evaluated by double-blind, placebo-controlled food challenge. Allergy 2003, 58, 132–138. [Google Scholar] [CrossRef]
- Wang, Y.; Fu, T.J.; Howard, A.; Kothary, M.H.; McHugh, T.H.; Zhang, Y. Crystal structure of peanut (Arachis hypogaea) allergen Ara h 5. J. Agric. Food Chem. 2013, 61, 1573–1578. [Google Scholar] [CrossRef]
- Kapingidza, A.B.; Pye, S.E.; Hyduke, N.; Dolamore, C.; Pote, S.; Schlachter, C.R.; Commins, S.P.; Kowal, K.; Chruszcz, M. Comparative structural and thermal stability studies of Cuc m 2.0101, Art v 4.0101 and other allergenic profilins. Mol. Immunol. 2019, 114, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, A.A.; Ball, T.; Mahoney, N.M.; Valenta, R.; Almo, S.C. The molecular basis for allergen cross-reactivity: Crystal structure and IgE-epitope mapping of birch pollen profilin. Structure 1997, 5, 33–45. [Google Scholar] [CrossRef]
- Mares-Mejia, I.; Martinez-Caballero, S.; Garay-Canales, C.; Cano-Sanchez, P.; Torres-Larios, A.; Lara-Gonzalez, S.; Ortega, E.; Rodriguez-Romero, A. Structural insights into the IgE mediated responses induced by the allergens Hev b 8 and Zea m 12 in their dimeric forms. Sci. Rep. 2016, 6, 32552. [Google Scholar] [CrossRef]
- Breiteneder, H.; Ebner, C. Molecular and biochemical classification of plant-derived food allergens. J. Allergy Clin. Immunol. 2000, 106, 27–36. [Google Scholar] [CrossRef]
- Hoffmann-Sommergruber, K. Pathogenesis-related (PR)-proteins identified as allergens. Biochem. Soc. Trans. 2002, 30, 930–935. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212–213, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, H.; Michalska, K.; Sikorski, M.; Jaskolski, M. Structural and functional aspects of PR-10 proteins. FEBS J. 2013, 280, 1169–1199. [Google Scholar] [CrossRef] [PubMed]
- Koistinen, K.M.; Soininen, P.; Venalainen, T.A.; Hayrinen, J.; Laatikainen, R.; Perakyla, M.; Tervahauta, A.I.; Karenlampi, S.O. Birch PR-10c interacts with several biologically important ligands. Phytochemistry 2005, 66, 2524–2533. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, J.E.; Wimmer, R.; Larsen, J.N.; Spangfort, M.D.; Otzen, D.E. The major birch allergen, Bet v 1, shows affinity for a broad spectrum of physiological ligands. J. Biol. Chem. 2002, 277, 23684–23692. [Google Scholar] [CrossRef]
- Bantignies, B.; Seguin, J.; Muzac, I.; Dedaldechamp, F.; Gulick, P.; Ibrahim, R. Direct evidence for ribonucleolytic activity of a PR-10-like protein from white lupin roots. Plant Mol. Biol. 2000, 42, 871–881. [Google Scholar] [CrossRef]
- Koistinen, K.M.; Kokko, H.I.; Hassinen, V.H.; Tervahauta, A.I.; Auriola, S.; Karenlampi, S.O. Stress-related RNase PR-10c is post-translationally modified by glutathione in birch. Plant Cell Environ. 2002, 25, 707–715. [Google Scholar] [CrossRef]
- Mittag, D.; Akkerdaas, J.; Ballmer-Weber, B.K.; Vogel, L.; Wensing, M.; Becker, W.M.; Koppelman, S.J.; Knulst, A.C.; Helbling, A.; Hefle, S.L.; et al. Ara h 8, a Bet v 1-homologous allergen from peanut, is a major allergen in patients with combined birch pollen and peanut allergy. J. Allergy Clin. Immunol. 2004, 114, 1410–1417. [Google Scholar] [CrossRef]
- Kos, T.; Hoffmann-Sommergruber, K.; Ferreira, F.; Hirschwehr, R.; Ahorn, H.; Horak, F.; Jager, S.; Sperr, W.; Kraft, D.; Scheiner, O. Purification, characterization and N-terminal amino acid sequence of a new major allergen from European chestnut pollen-Cas s 1. Biochem. Biophys. Res. Commun. 1993, 196, 1086–1092. [Google Scholar] [CrossRef]
- Florvaag, E.; Holen, E.; Vik, H.; Elsayed, S. Comparative studies on tree pollen allergens. XIV. Characterization of the birch (Betula verrucosa) and hazel (Corylus avellana) pollen extracts by horizontal 2-D SDS-PAGE combined with electrophoretic transfer and IgE immunoautoradiography. Ann. Allergy 1988, 61, 392–400. [Google Scholar]
- Hauser, M.; Roulias, A.; Ferreira, F.; Egger, M. Panallergens and their impact on the allergic patient. Allergy Asthma Clin. Immunol. 2010, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Jacob, T.; von Loetzen, C.S.; Reuter, A.; Lacher, U.; Schiller, D.; Schobert, R.; Mahler, V.; Vieths, S.; Rosch, P.; Schweimer, K.; et al. Identification of a natural ligand of the hazel allergen Cor a 1. Sci. Rep. 2019, 9, 8714. [Google Scholar] [CrossRef] [PubMed]
- Hurlburt, B.K.; Offermann, L.R.; McBride, J.K.; Majorek, K.A.; Maleki, S.J.; Chruszcz, M. Structure and function of the peanut panallergen Ara h 8. J. Biol. Chem. 2013, 288, 36890–36901. [Google Scholar] [CrossRef] [PubMed]
- Berkner, H.; Neudecker, P.; Mittag, D.; Ballmer-Weber, B.K.; Schweimer, K.; Vieths, S.; Rosch, P. Cross-reactivity of pollen and food allergens: Soybean Gly m 4 is a member of the Bet v 1 superfamily and closely resembles yellow lupine proteins. Biosci. Rep. 2009, 29, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Li, H.; Fan, X.; Li, Y.; Zhang, C.; Zhu, L.; Hu, J.; Kombe Kombe, A.J.; Xie, J.; Yin, D.; et al. Identification of a novel major allergen in buckwheat seeds: Fag t 6. J. Agric. Food Chem. 2021, 69, 13315–13322. [Google Scholar] [CrossRef]
- Shao, Q.; Liu, X.; Su, T.; Ma, C.; Wang, P. New insights into the role of seed oil body proteins in metabolism and plant development. Front. Plant Sci. 2019, 10, 1568. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.H. Oleosins and oil bodies in seeds and other organs. Plant Physiol. 1996, 110, 1055–1061. [Google Scholar] [CrossRef]
- Nikiforidis, C.V. Structure and functions of oleosomes (oil bodies). Adv. Colloid Interface Sci. 2019, 274, 102039. [Google Scholar] [CrossRef]
- Jolivet, P.; Acevedo, F.; Boulard, C.; d’Andrea, S.; Faure, J.D.; Kohli, A.; Nesi, N.; Valot, B.; Chardot, T. Crop seed oil bodies: From challenges in protein identification to an emerging picture of the oil body proteome. Proteomics 2013, 13, 1836–1849. [Google Scholar] [CrossRef]
- Shimada, T.L.; Hayashi, M.; Hara-Nishimura, I. Membrane dynamics and multiple functions of oil bodies in seeds and leaves. Plant Physiol. 2018, 176, 199–207. [Google Scholar] [CrossRef]
- Parthibane, V.; Rajakumari, S.; Venkateshwari, V.; Iyappan, R.; Rajasekharan, R. Oleosin is bifunctional enzyme that has both monoacylglycerol acyltransferase and phospholipase activities. J. Biol. Chem. 2012, 287, 1946–1954. [Google Scholar] [CrossRef]
- Tzen, J.T.C. Integral Proteins in Plant Oil Bodies. ISRN Bot. 2012, 2012, 173954. [Google Scholar] [CrossRef]
- Geoghegan, I.; Steinberg, G.; Gurr, S. The role of the fungal cell wall in the infection of plants. Trends Microbiol. 2017, 25, 957–967. [Google Scholar] [CrossRef]
- Durkin, C.A.; Mock, T.; Armbrust, E.V. Chitin in diatoms and its association with the cell wall. Eukaryot. Cell 2009, 8, 1038–1050. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Peng, D. Nematode chitin and application. Adv. Exp. Med. Biol. 2019, 1142, 209–219. [Google Scholar]
- Merzendorfer, H.; Zimoch, L. Chitin metabolism in insects: Structure, function and regulation of chitin synthases and chitinases. J. Exp. Biol. 2003, 206, 4393–4412. [Google Scholar] [CrossRef]
- Guan, Y.; Chye, M.L. A Brassica juncea chitinase with two-chitin binding domains show anti-microbial properties against phytopathogens and Gram-negative bacteria. Plant Signal. Behav. 2008, 3, 1103–1105. [Google Scholar] [CrossRef]
- Fujimura, M.; Minami, Y.; Watanabe, K.; Tadera, K. Purification, characterization, and sequencing of a novel type of antimicrobial peptides, Fa-AMP1 and Fa-AMP2, from seeds of buckwheat (Fagopyrum esculentum Moench.). Biosci. Biotechnol. Biochem. 2003, 67, 1636–1642. [Google Scholar] [CrossRef]
- van der Wel, H.; Loeve, K. Isolation and characterization of thaumatin I and II, the sweet-tasting proteins from Thaumatococcus daniellii Benth. Eur. J. Biochem. 1972, 31, 221–225. [Google Scholar] [CrossRef]
- Masuda, T.; Okubo, K.; Murata, K.; Mikami, B.; Sugahara, M.; Suzuki, M.; Temussi, P.A.; Tani, F. Subatomic structure of hyper-sweet thaumatin D21N mutant reveals the importance of flexible conformations for enhanced sweetness. Biochimie 2019, 157, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Leone, P.; Menu-Bouaouiche, L.; Peumans, W.J.; Payan, F.; Barre, A.; Roussel, A.; Van Damme, E.J.; Rouge, P. Resolution of the structure of the allergenic and antifungal banana fruit thaumatin-like protein at 1.7-A. Biochimie 2006, 88, 45–52. [Google Scholar] [CrossRef]
- Muhammad, I.; Li, W.Q.; Jing, X.Q.; Zhou, M.R.; Shalmani, A.; Ali, M.; Wei, X.Y.; Sharif, R.; Liu, W.T.; Chen, K.M. A systematic in silico prediction of gibberellic acid stimulated GASA family members: A novel small peptide contributes to floral architecture and transcriptomic changes induced by external stimuli in rice. J. Plant Physiol. 2019, 234–235, 117–132. [Google Scholar] [CrossRef]
- Fan, S.; Zhang, D.; Zhang, L.; Gao, C.; Xin, M.; Tahir, M.M.; Li, Y.; Ma, J.; Han, M. Comprehensive analysis of GASA family members in the Malus domestica genome: Identification, characterization, and their expressions in response to apple flower induction. BMC Genom. 2017, 18, 827. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, S.; Xu, D.; Liu, X.; Li, X.; Xiao, W.; Cao, J.; Jiang, H.; Min, X.; Wang, J.; et al. Identification and analysis of the GASR Gene family in common wheat (Triticum aestivum L.) and characterization of TaGASR34, a gene associated with seed dormancy and germination. Front. Genet. 2019, 10, 980. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, X. One new kind of phytohormonal signaling integrator: Up-and-coming GASA family genes. Plant Signal. Behav. 2017, 12, e1226453. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, K.A.; Moskal, W.A., Jr.; Wu, H.C.; Underwood, B.A.; Graham, M.A.; Town, C.D.; VandenBosch, K.A. Small cysteine-rich peptides resembling antimicrobial peptides have been under-predicted in plants. Plant J. 2007, 51, 262–280. [Google Scholar] [CrossRef] [PubMed]
- Aubert, D.; Chevillard, M.; Dorne, A.M.; Arlaud, G.; Herzog, M. Expression patterns of GASA genes in Arabidopsis thaliana: The GASA4 gene is up-regulated by gibberellins in meristematic regions. Plant Mol. Biol. 1998, 36, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. The Clustal Omega multiple alignment package. Methods Mol. Biol. 2021, 2231, 3–16. [Google Scholar] [CrossRef]
- Rodriguez-Decuadro, S.; Barraco-Vega, M.; Dans, P.D.; Pandolfi, V.; Benko-Iseppon, A.M.; Cecchetto, G. Antimicrobial and structural insights of a new snakin-like peptide isolated from Peltophorum dubium (Fabaceae). Amino Acids 2018, 50, 1245–1259. [Google Scholar] [CrossRef]
- Almasia, N.I.; Nahirnak, V.; Hopp, H.E.; Vazquez-Rovere, C. Potato Snakin-1: An antimicrobial player of the trade-off between host defense and development. Plant Cell Rep. 2020, 39, 839–849. [Google Scholar] [CrossRef]
- Yeung, H.; Squire, C.J.; Yosaatmadja, Y.; Panjikar, S.; Lopez, G.; Molina, A.; Baker, E.N.; Harris, P.W.; Brimble, M.A. Radiation damage and racemic protein crystallography reveal the unique structure of the GASA/Snakin protein superfamily. Angew. Chem. Int. Ed. Engl. 2016, 55, 7930–7933. [Google Scholar] [CrossRef] [PubMed]
- Pomes, A.; Chruszcz, M.; Gustchina, A.; Minor, W.; Mueller, G.A.; Pedersen, L.C.; Wlodawer, A.; Chapman, M.D. 100 Years later: Celebrating the contributions of x-ray crystallography to allergy and clinical immunology. J. Allergy Clin. Immunol. 2015, 136, 29–37.e10. [Google Scholar] [CrossRef]
- Barlow, D.J.; Edwards, M.S.; Thornton, J.M. Continuous and discontinuous protein antigenic determinants. Nature 1986, 322, 747–748. [Google Scholar] [CrossRef]
- Dall’antonia, F.; Pavkov-Keller, T.; Zangger, K.; Keller, W. Structure of allergens and structure based epitope predictions. Methods 2014, 66, 3–21. [Google Scholar] [CrossRef]
- Burks, A.W.; Shin, D.; Cockrell, G.; Stanley, J.S.; Helm, R.M.; Bannon, G.A. Mapping and mutational analysis of the IgE-binding epitopes on Ara h 1, a legume vicilin protein and a major allergen in peanut hypersensitivity. Eur. J. Biochem. 1997, 245, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Shreffler, W.G.; Lencer, D.A.; Bardina, L.; Sampson, H.A. IgE and IgG4 epitope mapping by microarray immunoassay reveals the diversity of immune response to the peanut allergen, Ara h 2. J. Allergy Clin. Immunol. 2005, 116, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Shreffler, W.G.; Beyer, K.; Chu, T.H.; Burks, A.W.; Sampson, H.A. Microarray immunoassay: Association of clinical history, in vitro IgE function, and heterogeneity of allergenic peanut epitopes. J. Allergy Clin. Immunol. 2004, 113, 776–782. [Google Scholar] [CrossRef]
- Matsumoto, N.; Okochi, M.; Matsushima, M.; Ogawa, A.; Takase, T.; Yoshida, Y.; Kawase, M.; Isobe, K.; Kawabe, T.; Honda, H. Development of peptide arrays for detection of IgE-binding epitopes in cow’s milk allergens. J. Biosci. Bioeng. 2009, 107, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Jarvinen, K.M.; Chatchatee, P.; Bardina, L.; Beyer, K.; Sampson, H.A. IgE and IgG binding epitopes on alpha-lactalbumin and beta-lactoglobulin in cow’s milk allergy. Int. Arch. Allergy Immunol. 2001, 126, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Bardina, L.; Shreffler, W.G.; Andreae, D.A.; Ge, Y.; Wang, J.; Bruni, F.M.; Fu, Z.; Han, Y.; Sampson, H.A. Development of a novel peptide microarray for large-scale epitope mapping of food allergens. J. Allergy Clin. Immunol. 2009, 124, 315–322.e3. [Google Scholar] [CrossRef]
- Shin, D.S.; Compadre, C.M.; Maleki, S.J.; Kopper, R.A.; Sampson, H.; Huang, S.K.; Burks, A.W.; Bannon, G.A. Biochemical and structural analysis of the IgE binding sites on ara h1, an abundant and highly allergenic peanut protein. J. Biol. Chem. 1998, 273, 13753–13759. [Google Scholar] [CrossRef]
- Stanley, J.S.; King, N.; Burks, A.W.; Huang, S.K.; Sampson, H.; Cockrell, G.; Helm, R.M.; West, C.M.; Bannon, G.A. Identification and mutational analysis of the immunodominant IgE binding epitopes of the major peanut allergen Ara h 2. Arch. Biochem. Biophys. 1997, 342, 244–253. [Google Scholar] [CrossRef]
- Rouge, P.; Culerrier, R.; Sabatier, V.; Granier, C.; Rance, F.; Barre, A. Mapping and conformational analysis of IgE-binding epitopic regions on the molecular surface of the major Ara h 3 legumin allergen of peanut (Arachis hypogaea). Mol. Immunol. 2009, 46, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Flinterman, A.E.; Knol, E.F.; Lencer, D.A.; Bardina, L.; den Hartog Jager, C.F.; Lin, J.; Pasmans, S.G.; Bruijnzeel-Koomen, C.A.; Sampson, H.A.; van Hoffen, E.; et al. Peanut epitopes for IgE and IgG4 in peanut-sensitized children in relation to severity of peanut allergy. J. Allergy Clin. Immunol. 2008, 121, 737–743.e710. [Google Scholar] [CrossRef] [PubMed]
- Reese, G.; Ayuso, R.; Leong-Kee, S.M.; Plante, M.J.; Lehrer, S.B. Characterization and identification of allergen epitopes: Recombinant peptide libraries and synthetic, overlapping peptides. J. Chromatogr. B Biomed. Sci. Appl. 2001, 756, 157–163. [Google Scholar] [CrossRef]
- Robotham, J.M.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Linear IgE epitope mapping of the English walnut (Juglans regia) major food allergen, Jug r 1. J. Allergy Clin. Immunol. 2002, 109, 143–149. [Google Scholar] [CrossRef]
- Fang, L.; Li, G.; Zhang, J.; Gu, R.; Cai, M.; Lu, J. Identification and mutational analysis of continuous, immunodominant epitopes of the major oyster allergen Crag 1. Clin. Immunol. 2019, 201, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bhardwaj, S.R.; Vilches, A.; Breksa, A.; Lyu, S.C.; Chinthrajah, S.; Nadeau, K.C.; Jin, T. IgE binding epitope mapping with TL1A tagged peptides. Mol. Immunol. 2022, 153, 194–199. [Google Scholar] [CrossRef]
- Yang, L.; Kulis, M. Hypoallergenic proteins for the treatment of food allergy. Curr. Allergy Asthma Rep. 2019, 19, 15. [Google Scholar] [CrossRef]
- Niemi, M.; Jylha, S.; Laukkanen, M.L.; Soderlund, H.; Makinen-Kiljunen, S.; Kallio, J.M.; Hakulinen, N.; Haahtela, T.; Takkinen, K.; Rouvinen, J. Molecular interactions between a recombinant IgE antibody and the beta-lactoglobulin allergen. Structure 2007, 15, 1413–1421. [Google Scholar] [CrossRef]
- Padavattan, S.; Flicker, S.; Schirmer, T.; Madritsch, C.; Randow, S.; Reese, G.; Vieths, S.; Lupinek, C.; Ebner, C.; Valenta, R.; et al. High-affinity IgE recognition of a conformational epitope of the major respiratory allergen Phl p 2 as revealed by X-ray crystallography. J. Immunol. 2009, 182, 2141–2151. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ramirez, B.; Mares-Mejia, I.; Rodriguez-Hernandez, A.; Cano-Sanchez, P.; Torres-Larios, A.; Ortega, E.; Rodriguez-Romero, A. A native IgE in complex with profilin provides insights into allergen recognition and cross-reactivity. Commun. Biol. 2022, 5, 748. [Google Scholar] [CrossRef] [PubMed]
- Mitropoulou, A.N.; Bowen, H.; Dodev, T.S.; Davies, A.M.; Bax, H.J.; Beavil, R.L.; Beavil, A.J.; Gould, H.J.; James, L.K.; Sutton, B.J. Structure of a patient-derived antibody in complex with allergen reveals simultaneous conventional and superantigen-like recognition. Proc. Natl. Acad. Sci. USA 2018, 115, E8707–E8716. [Google Scholar] [CrossRef]
- James, L.C.; Roversi, P.; Tawfik, D.S. Antibody multispecificity mediated by conformational diversity. Science 2003, 299, 1362–1367. [Google Scholar] [CrossRef] [PubMed]
- NIH_Expert_Panel. Food Allergey: Report of the NIH Expert Panel on Food Allergy Research; NIH: Bethesda, MD, USA, 2006.
- Wildner, S.; Griessner, I.; Stemeseder, T.; Regl, C.; Soh, W.T.; Stock, L.G.; Volker, T.; Alessandri, C.; Mari, A.; Huber, C.G.; et al. Boiling down the cysteine-stabilized LTP fold—Loss of structural and immunological integrity of allergenic Art v 3 and Pru p 3 as a consequence of irreversible lanthionine formation. Mol. Immunol. 2019, 116, 140–150. [Google Scholar] [CrossRef]
- Koppelman, S.J.; Nieuwenhuizen, W.F.; Gaspari, M.; Knippels, L.M.; Penninks, A.H.; Knol, E.F.; Hefle, S.L.; de Jongh, H.H. Reversible denaturation of Brazil nut 2S albumin (Ber e1) and implication of structural destabilization on digestion by pepsin. J. Agric. Food Chem. 2005, 53, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Van Boxtel, E.L.; van den Broek, L.A.; Koppelman, S.J.; Gruppen, H. Legumin allergens from peanuts and soybeans: Effects of denaturation and aggregation on allergenicity. Mol. Nutr. Food Res. 2008, 52, 674–682. [Google Scholar] [CrossRef]
- Tuppo, L.; Spadaccini, R.; Alessandri, C.; Wienk, H.; Boelens, R.; Giangrieco, I.; Tamburrini, M.; Mari, A.; Picone, D.; Ciardiello, M.A. Structure, stability, and IgE binding of the peach allergen Peamaclein (Pru p 7). Biopolymers 2014, 102, 416–425. [Google Scholar] [CrossRef]
- Rib-Schmidt, C.; Riedl, P.; Meisinger, V.; Schwaben, L.; Schulenborg, T.; Reuter, A.; Schiller, D.; Seutter von Loetzen, C.; Rosch, P. pH and heat resistance of the major celery allergen Api g 1. Mol. Nutr. Food Res. 2018, 62, e1700886. [Google Scholar] [CrossRef]
- Cabanos, C.; Katayama, H.; Tanaka, A.; Utsumi, S.; Maruyama, N. Expression and purification of peanut oleosins in insect cells. Protein J. 2011, 30, 457–463. [Google Scholar] [CrossRef]
- Smole, U.; Bublin, M.; Radauer, C.; Ebner, C.; Breiteneder, H. Mal d 2, the thaumatin-like allergen from apple, is highly resistant to gastrointestinal digestion and thermal processing. Int. Arch. Allergy Immunol. 2008, 147, 289–298. [Google Scholar] [CrossRef]
- Nesbit, J.B.; Hurlburt, B.K.; Schein, C.H.; Cheng, H.; Wei, H.; Maleki, S.J. Ara h 1 structure is retained after roasting and is important for enhanced binding to IgE. Mol. Nutr. Food Res. 2012, 56, 1739–1747. [Google Scholar] [CrossRef]
- Mattison, C.P.; Bren-Mattison, Y.; Vant-Hull, B.; Vargas, A.M.; Wasserman, R.L.; Grimm, C.C. Heat-induced alterations in cashew allergen solubility and IgE binding. Toxicol. Rep. 2016, 3, 244–251. [Google Scholar] [CrossRef]
- Mattison, C.P.; Grimm, C.C.; Li, Y.; Chial, H.J.; McCaslin, D.R.; Chung, S.Y.; Bren-Mattison, Y.; Wasserman, R.L. Identification and characterization of Ana o 3 modifications on arginine-111 residue in heated cashew nuts. J. Agric. Food Chem. 2017, 65, 411–420. [Google Scholar] [CrossRef]
- Verhoeckx, K.; Bogh, K.L.; Dupont, D.; Egger, L.; Gadermaier, G.; Larre, C.; Mackie, A.; Menard, O.; Adel-Patient, K.; Picariello, G.; et al. The relevance of a digestibility evaluation in the allergenicity risk assessment of novel proteins. Opinion of a joint initiative of COST action ImpARAS and COST action INFOGEST. Food Chem. Toxicol. 2019, 129, 405–423. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.J.; Abbott, U.R.; Hatzos, C. Digestibility of food allergens and nonallergenic proteins in simulated gastric fluid and simulated intestinal fluid-a comparative study. J. Agric. Food Chem. 2002, 50, 7154–7160. [Google Scholar] [CrossRef] [PubMed]
- Bogh, K.L.; Barkholt, V.; Rigby, N.M.; Mills, E.N.; Madsen, C.B. Digested Ara h 1 loses sensitizing capacity when separated into fractions. J. Agric. Food Chem. 2012, 60, 2934–2942. [Google Scholar] [CrossRef] [PubMed]
- Scheurer, S.; Lauer, I.; Foetisch, K.; San Miguel Moncin, M.; Retzek, M.; Hartz, C.; Enrique, E.; Lidholm, J.; Cistero-Bahima, A.; Vieths, S. Strong allergenicity of Pru av 3, the lipid transfer protein from cherry, is related to high stability against thermal processing and digestion. J. Allergy Clin. Immunol. 2004, 114, 900–907. [Google Scholar] [CrossRef] [PubMed]
- Kenna, J.G.; Evens, R.M. Digestibility of proteins in simulated gastric fluid [Abstract]. Toxicologist 2000, 54, 141. [Google Scholar]
- Smith, F.; Pan, X.Y.; Bellido, V.; Toole, G.A.; Gates, F.K.; Wickham, M.S.J.; Shewry, P.R.; Bakalis, S.; Padfield, P.; Mills, E.N.C. Digestibility of gluten proteins is reduced by baking and enhanced by starch digestion. Mol. Nutr. Food Res. 2015, 59, 2034–2043. [Google Scholar] [CrossRef] [PubMed]
- Mayer, L. Mucosal immunity. Pediatrics 2003, 111, 1595–1600. [Google Scholar] [CrossRef] [PubMed]
- Husby, S.; Foged, N.; Host, A.; Svehag, S.E. Passage of dietary antigens into the blood of children with coeliac disease. Quantification and size distribution of absorbed antigens. Gut 1987, 28, 1062–1072. [Google Scholar] [CrossRef]
- Martinez San Ireneo, M.; Ibanez Sandin, M.D.; Fernandez-Caldas, E. Hypersensitivity to members of the botanical order Fabales (legumes). J. Investig. Allergol. Clin. Immunol. 2000, 10, 187–199. [Google Scholar] [PubMed]
- de Leon, M.P.; Drew, A.C.; Glaspole, I.N.; Suphioglu, C.; O’Hehir, R.E.; Rolland, J.M. IgE cross-reactivity between the major peanut allergen Ara h 2 and tree nut allergens. Mol. Immunol. 2007, 44, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Yanagida, N.; Takei, M.; Saito, A.; Sato, S.; Ebisawa, M. Clinical cross-reactivity of wheat and barley in children with wheat allergy. Pediatr. Allergy Immunol. 2022, 33, e13878. [Google Scholar] [CrossRef]
- Smits, M.; Verhoeckx, K.; Knulst, A.; Welsing, P.; de Jong, A.; Gaspari, M.; Ehlers, A.; Verhoeff, P.; Houben, G.; Le, T.M. Co-sensitization between legumes is frequently seen, but variable and not always clinically relevant. Front. Allergy 2023, 4, 1115022. [Google Scholar] [CrossRef]
- Bublin, M.; Kostadinova, M.; Radauer, C.; Hafner, C.; Szepfalusi, Z.; Varga, E.M.; Maleki, S.J.; Hoffmann-Sommergruber, K.; Breiteneder, H. IgE cross-reactivity between the major peanut allergen Ara h 2 and the nonhomologous allergens Ara h 1 and Ara h 3. J. Allergy Clin. Immunol. 2013, 132, 118–124. [Google Scholar] [CrossRef]
- Warmenhoven, H.J.M.; Hulsbos, L.; Dreskin, S.C.; Akkerdaas, J.H.; Versteeg, S.A.; van Ree, R. IgE cross-inhibition between Ara h 1 and Ara h 2 is explained by complex formation of both major peanut allergens. J. Allergy Clin. Immunol. 2023. [Google Scholar] [CrossRef]
- EFSA Panel on Genetically Modified Organisms (GMO); Mullins, E.; Bresson, J.L.; Dalmay, T.; Dewhurst, I.C.; Epstein, M.M.; George Firbank, L.; Guerche, P.; Hejatko, J.; Naegeli, H.; et al. Scientific opinion on development needs for the allergenicity and protein safety assessment of food and feed products derived from biotechnology. EFSA J. 2022, 20, e07044. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Family a | No. of Allergens | Proteins | No. of Allergens |
|---|---|---|---|
| PF00234: Tryp_alpha_amyl | 74 | Nsltp | 42 |
| 2S albumin | 26 | ||
| Others | 6 | ||
| PF00190: Cupin_1 | 39 | 11S | 19 |
| 7S | 20 | ||
| PF00235: Profilin | 27 | Profilin | |
| PF00407: Bet_v_1 | 19 | PR-10 | |
| PF01277: Oleosin | 10 | Oleosin | |
| PF00187: Chitin_bind_1 | 7 | Chitinases | |
| (PF00182: Glyco_hydro_19) | |||
| PF00314: Thaumatin | 6 | Thaumatin-like | |
| PF02704: GASA | 6 | Gibberellin regulated protein | |
| PF04702: Vicilin_N | 6 | N-terminal of vicilin cC3C antimicrobial protein | |
| PF00112: Peptidase_C1 | |||
| PF08246: Inhibitor_I29 | 3 | Cysteine protease | |
| PF00197: Kunitz_legume | 3 | Protease inhibitors | |
| PF00304: Gamma-thionin | 3 | Defensin | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Che, H.; Li, C.; Jin, T. Food Allergens of Plant Origin. Foods 2023, 12, 2232. https://doi.org/10.3390/foods12112232
Zhang Y, Che H, Li C, Jin T. Food Allergens of Plant Origin. Foods. 2023; 12(11):2232. https://doi.org/10.3390/foods12112232
Chicago/Turabian StyleZhang, Yuzhu, Huilian Che, Caiming Li, and Tengchuan Jin. 2023. "Food Allergens of Plant Origin" Foods 12, no. 11: 2232. https://doi.org/10.3390/foods12112232