
Functional Food Based on Potato

Abstract
:1. Introduction
2. Potato Starch and Its Functional Properties
2.1. Structure of Potato Starch
2.2. Digestive Properties of Potato Starch
2.3. Potato Starch and Edible Films
3. Potato Protein as Functional Food Ingredients
3.1. Potato Starch and Edible Films
3.2. Amino Acid Composition of Potato Protein
3.3. Fractions of Potato Protein
3.4. Potential Applications of Potato Protein
4. Potato Phytochemicals and Nutritional Potential
4.1. Functional Phytochemicals in Potato
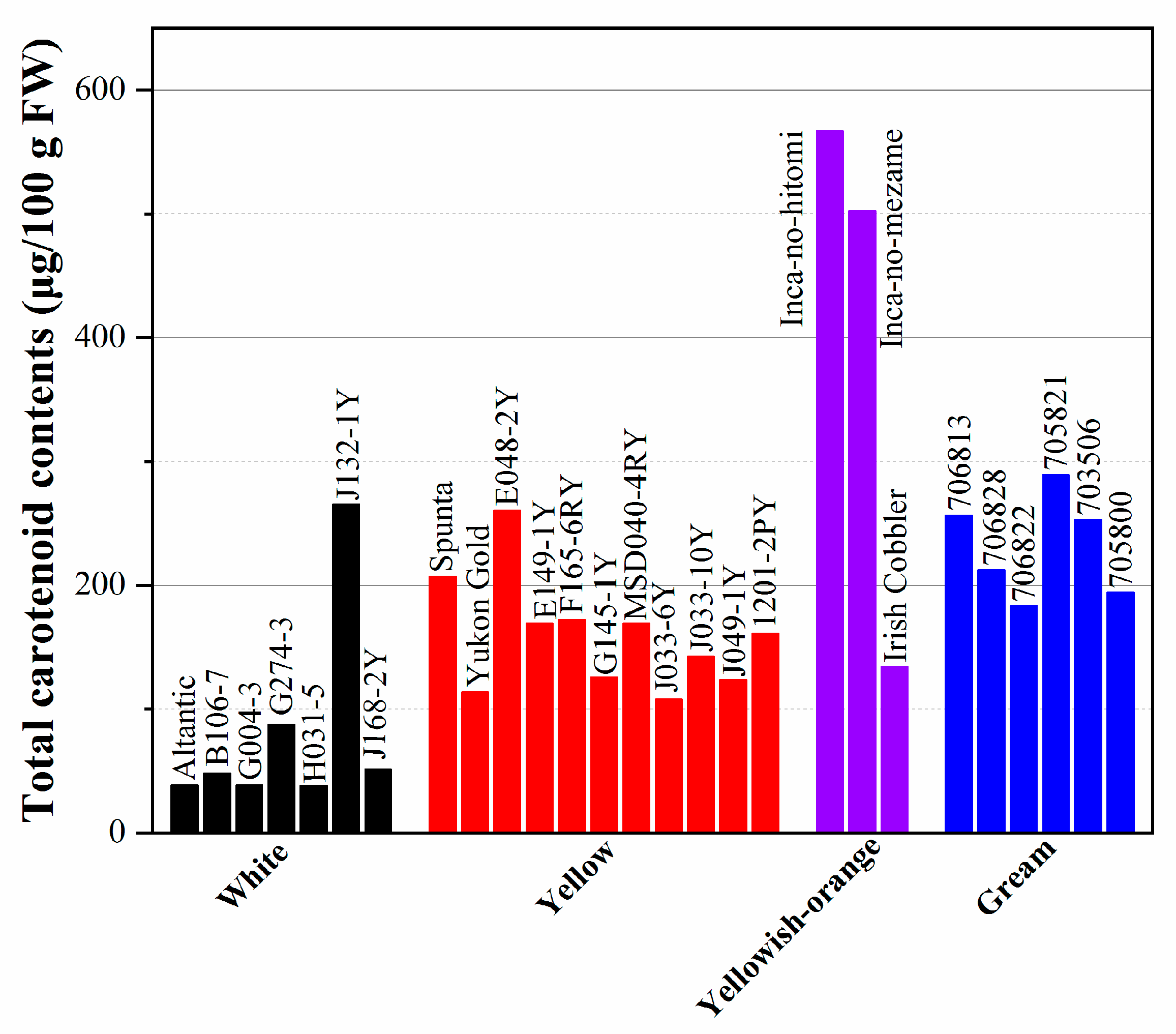
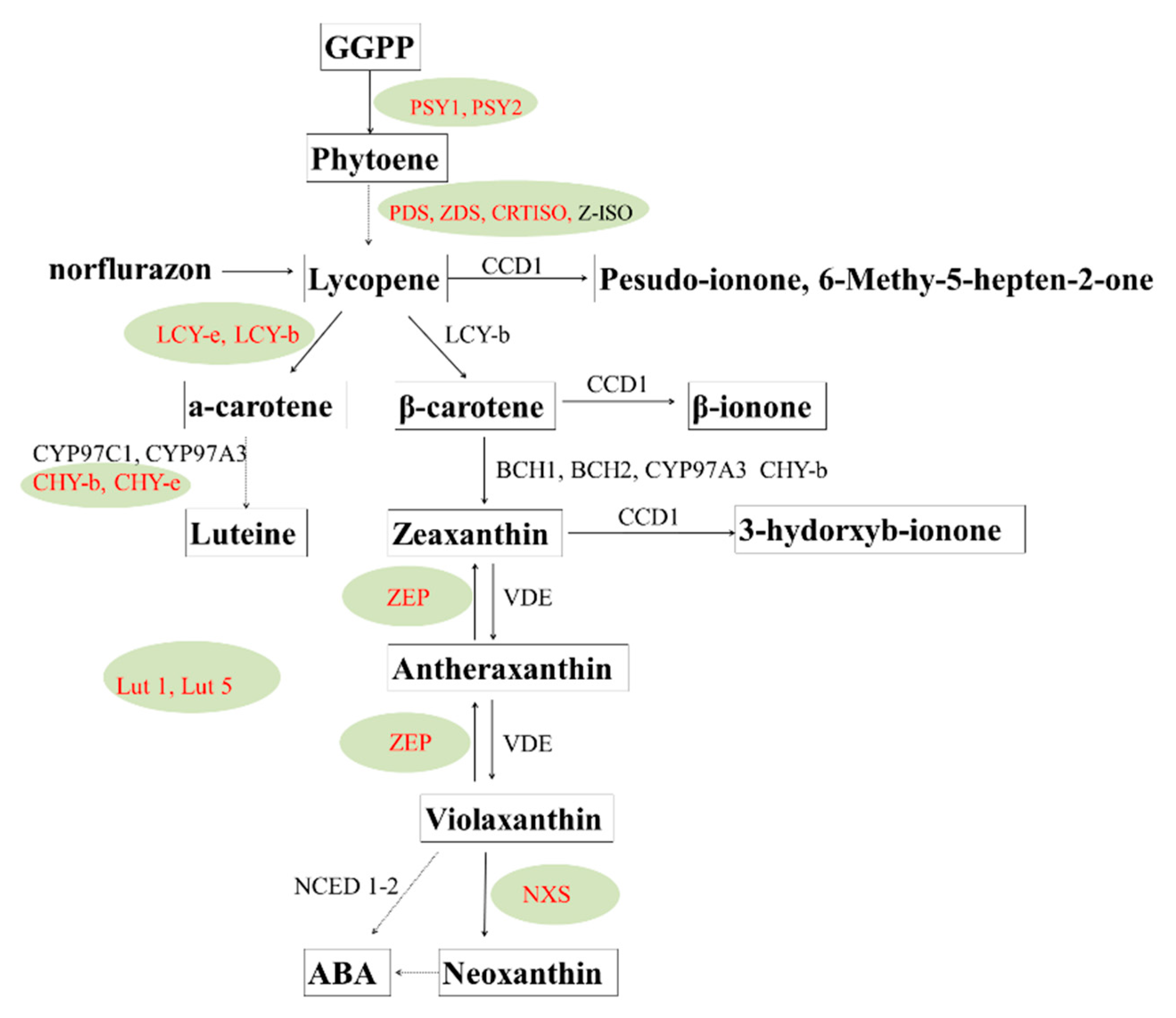
4.2. Carotenoids and Health Benefits
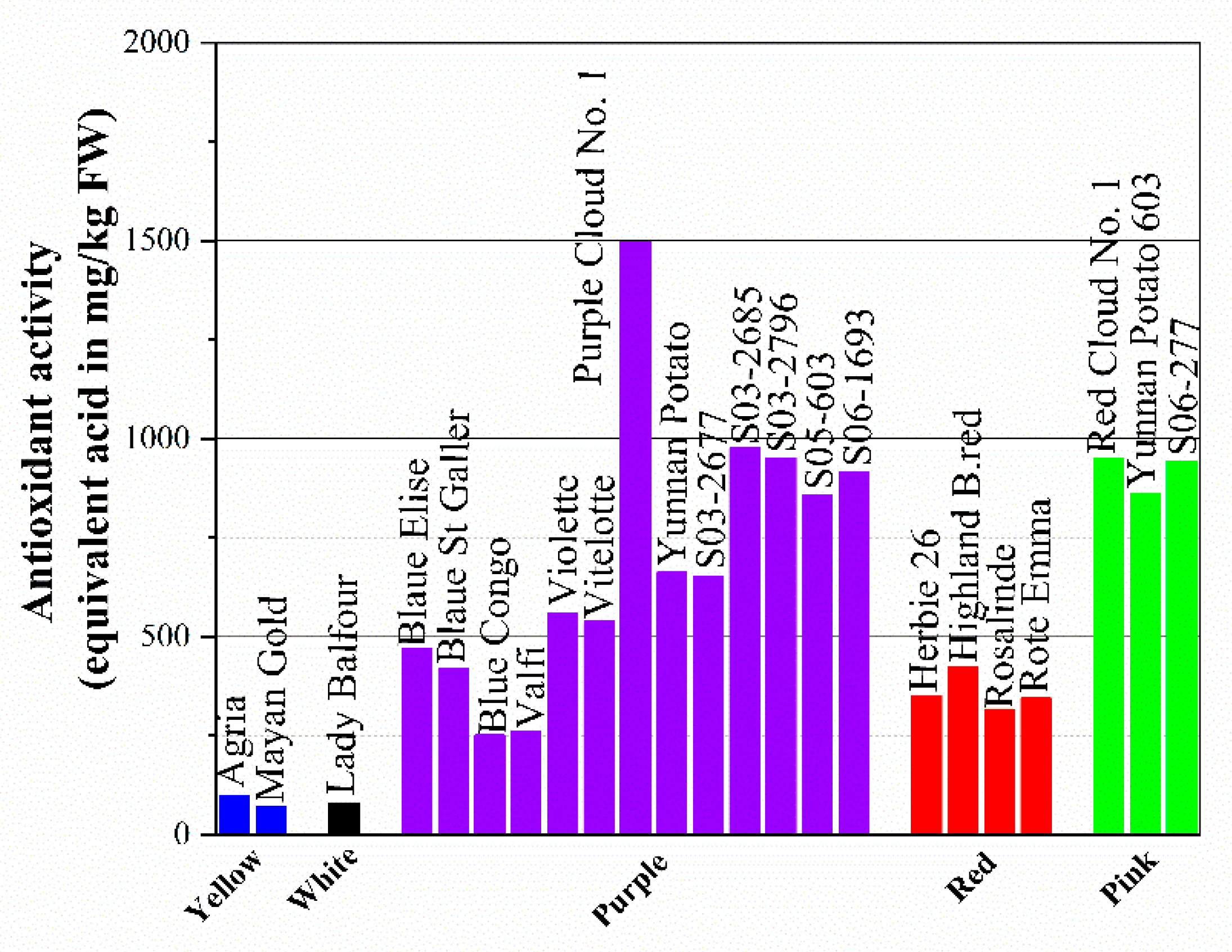
4.3. Phenolic Compounds and Antioxidant Activities
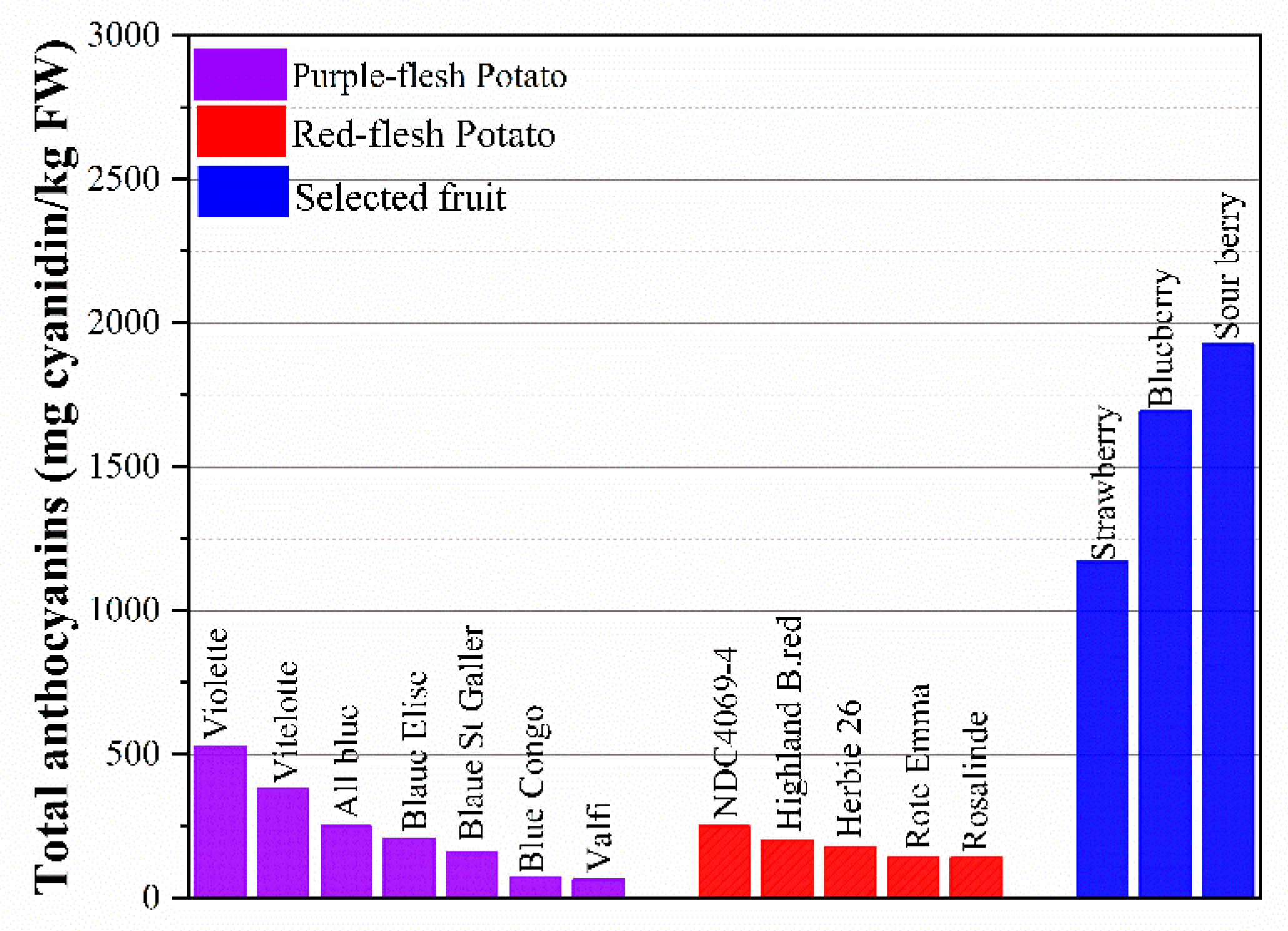
4.4. Flavonoids and Health Care Functions
4.5. Vitamins and Nutritional Potential
4.5.1. Vitamin C
4.5.2. Vitamin B9
4.5.3. Vitamin B6
5. Potato Minerals and Health Benefits
5.1. Potassium
5.2. Iron
5.3. Magnesium
5.4. Phosphorus
5.5. Acrylamide
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cesari, A.; Falcinelli, A.L.; Mendieta, J.R.; Pagano, M.R.; Mucci, N.; Daleo, G.R.; Guevara, M.G. Potato aspartic proteases (StAPs) exert cytotoxic activity on bovine and human spermatozoa. Fertil. Steril. 2007, 88, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Burgos, G.; Zum Felde, T.; Andre, C.; Kubow, S. The Potato and Its Contribution to the Human Diet and Health. Potato Crop Agric. Nutr. Soc. Contrib. Hum. 2020, 5, 73–74. [Google Scholar]
- Rommens, C.M.; Yan, H.; Swords, K.; Richael, C.; Ye, J. Low-acrylamide French fries and potato chips. Plant Biotechnol. J. 2008, 6, 843–853. [Google Scholar] [CrossRef]
- Kaguongo, W.; Lungaho, C.; Borus, D.; Kipkoech, D.; Ng’Ang’A, N. A Policymakers’ Guide to Crop Diversification: The Case of the Potato in Kenya; Food and Agriculture Organization of the United Nations Rome: Rome, Italy, 2013. [Google Scholar]
- Camire, M.E.; Kubow, S.; Donnelly, D.J. Potatoes and Human Health. Crit. Rev. Food Sci. Nutr. 2009, 49, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Visvanathan, R.; Jayathilake, C.; Jayawardana, B.; Liyanage, R. Health-beneficial properties of potato and compounds of interest. J. Sci. Food Agric. 2016, 96, 4850–4860. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, F.S.; Foster-Powell, K.; Brand-Miller, J.C. International Tables of Glycemic Index and Glycemic Load Values: 2008. Diabetes Care 2008, 31, 2281–2283. [Google Scholar] [CrossRef]
- Farhadnejad, H.; Teymoori, F.; Asghari, G.; Mirmiran, P.; Azizi, F. The Association of Potato Intake with Risk for Incident Type 2 Diabetes in Adults. Can. J. Diabetes 2018, 42, 613–618. [Google Scholar] [CrossRef]
- Khosravi-Boroujeni, H.; Mohammadifard, N.; Sarrafzadegan, N.; Sajjadi, F.; Maghroun, M.; Khosravi, A.; Alikhasi, H.; Rafieian-Kopaei, M.; Azadbakht, L. Potato consumption and cardiovascular disease risk factors among Iranian population. Int. J. Food Sci. Nutr. 2012, 63, 913–920. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Hao, T.; Rimm, E.B.; Willett, W.C.; Hu, F.B. Changes in Diet and Lifestyle and Long-Term Weight Gain in Women and Men. N. Engl. J. Med. 2011, 364, 2392–2404. [Google Scholar] [CrossRef]
- King, J.C.; Slavin, J.L. White Potatoes, Human Health, and Dietary Guidance. Adv. Nutr. 2013, 4, 393–401. [Google Scholar] [CrossRef]
- Huang, M.; Zhuang, P.; Jiao, J.; Wang, J.; Chen, X.; Zhang, Y. Potato consumption is prospectively associated with risk of hyper-tension: An 11.3-year longitudinal cohort study. Clin. Nutr. 2019, 38, 1936–1944. [Google Scholar] [CrossRef]
- Aburto, N.J.; Hanson, S.; Gutierrez, H.; Hooper, L.; Elliott, P.; Cappuccio, F.P. Effect of increased potassium intake on cardiovascular risk factors and disease: Systematic review and meta-analyses. BMJ 2013, 346, f1378. [Google Scholar] [CrossRef]
- van Niekerk, C.; Schönfeldt, H.; Hall, N.; Pretorius, B. The Role of Biodiversity in Food Security and Nutrition: A Potato Cultivar Case Study. Food Nutr. Sci. 2016, 07, 371–382. [Google Scholar] [CrossRef]
- Whistler, R.L.; Daniel, J.R. Chapter 5-Molecular Structure of Starch. In Strach: Chemistry and Technology; Elsevier: Amsterdam, The Netherlands, 1984; pp. 153–182. [Google Scholar]
- Osawa, M.; Inoue, N. Studies on the in vitro starch digestibility and the glycemic index in rice cultivars differing in amylose content. Jpn. J. Crop Sci. 2007, 76, 410–415. [Google Scholar] [CrossRef]
- Lin, A.H.-M. Structure and Digestion of Common Complementary Food Starches. J. Craniofacial Surg. 2018, 66, S35–S38. [Google Scholar] [CrossRef]
- Hall, A.; Versalovic, J. Microbial Metabolism in the Mammalian Gut: Molecular Mechanisms and Clinical Implications. J. Craniofacial Surg. 2018, 66, S72–S79. [Google Scholar] [CrossRef]
- Tan, X.; Li, X.; Chen, L.; Xie, F.; Li, L.; Huang, J. Effect of heat-moisture treatment on multi-scale structures and physicochemical properties of breadfruit starch. Carbohydr. Polym. 2017, 161, 286–294. [Google Scholar] [CrossRef]
- Lin, A.H.-M.; Lee, B.-H.; Nichols, B.L.; Quezada-Calvillo, R.; Rose, D.R.; Naim, H.Y.; Hamaker, B.R. Starch Source Influences Dietary Glucose Generation at the Mucosal α-Glucosidase Level. J. Biol. Chem. 2012, 287, 36917–36921. [Google Scholar] [CrossRef]
- Sawicka, B. Resistant starch in potato. Acta Sci. Pol. Technol. Aliment. 2018, 17, 153–169. [Google Scholar]
- Lockyer, S.; Nugent, A.P. Health effects of resistant starch. Nutr. Bull. 2017, 42, 10–41. [Google Scholar] [CrossRef]
- Brahma, B.; Sit, N. Physicochemical properties, and digestibility of heat moisture–treated potato starches for different treatment conditions. Potato Res. 2020, 63, 367–383. [Google Scholar] [CrossRef]
- Shortt, C.; Hasselwander, O.; Meynier, A.; Nauta, A.; Fernández, E.N.; Putz, P.; Rowland, I.; Swann, J.; Türk, J.; Vermeiren, J.; et al. Systematic review of the effects of the intestinal microbiota on selected nutrients and non-nutrients. Eur. J. Nutr. 2018, 57, 25–49. [Google Scholar] [CrossRef] [PubMed]
- Jane, J. Structural features of starch granules II. In Starch: Chemistry and Technology, 3rd ed.; BeMiller, J.N., Whistler, R.L., Eds.; Academic Press: New York, NY, USA, 2009; pp. 193–236. [Google Scholar]
- Lin, H.-M.; Chang, Y.-H.; Lin, J.-H.; Jane, J.-L.; Sheu, M.-J.; Lu, T.-J. Heterogeneity of lotus rhizome starch granules as revealed by α-amylase degradation. Carbohydr. Polym. 2006, 66, 528–536. [Google Scholar] [CrossRef]
- Zhang, G.; Hamaker, B.R. Slowly Digestible Starch: Concept, Mechanism, and Proposed Extended Glycemic Index. Crit. Rev. Food Sci. Nutr. 2009, 49, 852–867. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Sotomayor, M.; Quezada-Calvillo, R.; Avery, S.E.; Chacko, S.K.; Yan, L.-K.; Lin, A.H.-M.; Ao, Z.-H.; Hamaker, B.R.; Nichols, B.L. Maltase-Glucoamylase Modulates Gluconeogenesis and Sucrase-Isomaltase Dominates Starch Digestion Glucogenesis. J. Craniofacial Surg. 2013, 57, 704–712. [Google Scholar] [CrossRef]
- Lin, A.H.-M.; Lee, B.-H.; Chang, W.-J. Small intestine mucosal α-glucosidase: A missing feature of in vitro starch digestibility. Food Hydrocoll. 2016, 53, 163–171. [Google Scholar] [CrossRef]
- Butterworth, P.J.; Warren, F.J.; Grassby, T.; Patel, H.; Ellis, P.R. Analysis of starch amylolysis using plots for first-order kinetics. Carbohydr. Polym. 2012, 87, 2189–2197. [Google Scholar] [CrossRef]
- Amy, H.M.; Buford, L.; Nichols, L. The digestion of complementary feeding starches in the young child. Starch Strke 2017, 69, 170–178. [Google Scholar]
- Ben Jeddou, K.; Bouaziz, F.; Zouari-Ellouzi, S.; Chaari, F.; Ellouz-Chaabouni, S.; Ellouz-Ghorbel, R.; Nouri-Ellouz, O. Improvement of texture and sensory properties of cakes by addition of potato peel powder with high level of dietary fiber and protein. Food Chem. 2017, 217, 668–677. [Google Scholar] [CrossRef]
- Bajer, D.; Janczak, K.; Bajer, K. Novel Starch/Chitosan/Aloe Vera Composites as Promising Biopackaging Materials. J. Polym. Environ. 2020, 28, 1021–1039. [Google Scholar] [CrossRef]
- Zhang, R.; Cheng, M.; Wang, X.; Wang, J. Bioactive mesoporous nano-silica/potato starch films against molds commonly found in post-harvest white mushrooms. Food Hydrocoll. 2019, 95, 517–525. [Google Scholar] [CrossRef]
- Farajpour, R.; Djomeh, Z.E.; Moeini, S.; Tavakolipour, H.; Safayan, S. Structural and physico-mechanical properties of potato starch-olive oil edible films reinforced with zein nanoparticles. Int. J. Biol. Macromol. 2020, 149, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Liu, P.; Cui, B.; Kang, X.; Yu, B. Effects of different treatment methods on properties of potato starch-lauric acid complex and potato starch-based films. Int. J. Biol. Macromol. 2018, 124, 34–40. [Google Scholar] [CrossRef]
- Ayquipa-Cuellar, E.; Salcedo-Sucasaca, L.; Azamar-Barrios, J.A.; Chaquilla-Quilca, G. Assessment of Prickly Pear Peel Mucilage and Potato Husk Starch for Edible Films Production for Food Packaging Industries. Waste Biomass Valorization 2020, 12, 321–331. [Google Scholar] [CrossRef]
- Pęksa, A.; Miedzianka, J.; Nemś, A. Amino acid composition of flesh-coloured potatoes as affected by storage conditions. Food Chem. 2018, 266, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Pranaitiene, R.; Danilcenko, H.; Jariene, E.; Dabkevicius, Z. The effect of inhibitors on the changes of potato tuber quality during the storage period. J. Food Agric. Environ. 2008, 6, 231–235. [Google Scholar]
- Bartova, V.; Barta, J. Chemical composition and nutritional value of protein concentrates isolated from potato (Solanum tuberosum L.) fruit juice by precipitation with ethanol or ferric chloride. J. Agric. Food Chem. 2009, 57, 9028–9034. [Google Scholar] [CrossRef]
- AWaglay; Karboune, S. Chapter 4—Potato proteins: Functional food ingredients. In Advances in Potato Chemistry and Technology; Academic Press: San Diego, CA, USA, 2016; pp. 75–104. [Google Scholar]
- Fu, T.-J.; Abbott, U.R.; Hatzos, C. Digestibility of Food Allergens and Nonallergenic Proteins in Simulated Gastric Fluid and Simulated Intestinal FluidA Comparative Study. J. Agric. Food Chem. 2002, 50, 7154–7160. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Park, S.-C.; Kim, M.-H.; Lim, H.-T.; Park, Y.; Hahm, K.-S. Antimicrobial activity studies on a trypsin–chymotrypsin protease inhibitor obtained from potato. Biochem. Biophys. Res. Commun. 2005, 330, 921–927. [Google Scholar] [CrossRef]
- Pihlanto, A.; Akkanen, S.; Korhonen, H.J. ACE-inhibitory and antioxidant properties of potato (Solanum tuberosum). Food Chem. 2008, 109, 104–112. [Google Scholar] [CrossRef]
- Liyanage, R.; Han, K.-H.; Watanabe, S.; Shimada, K.-I.; Sekikawa, M.; Ohba, K.; Tokuji, Y.; Ohnishi, M.; Shibayama, S.; Nakamori, T.; et al. Potato and Soy Peptide Diets Modulate Lipid Metabolism in Rats. Biosci. Biotechnol. Biochem. 2008, 72, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Aparicio, C.; Molina, M.A.; Fernández-Salas, E.; Frazier, M.L.; Mas, J.M.; Querol, E.; Avilés, F.X.; de Llorens, R. Potato Carboxypeptidase Inhibitor, a T-knot Protein, Is an Epidermal Growth Factor Antagonist That Inhibits Tumor Cell Growth. J. Biol. Chem. 1998, 273, 12370–12377. [Google Scholar] [CrossRef] [PubMed]
- Mitrus, J.; Stankiewicz, C.; Steć, E.; Kamecki, M.; Starczewski, J. The influence of selected cultivation on the content of total protein and amino acids in the potato tubers. Plant Soil Environ. 2003, 49, 131–134. [Google Scholar] [CrossRef]
- Eppendorfer, W.H.; Eggum, B.O. Effects of sulphur, nitrogen, phosphorus, potassium, and water stress on dietary fibre fractions, starch, amino acids and on the biological value of potato protein. Plant Foods Hum. Nutr. 1994, 45, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, A.; Contreras, E.; Sousa, H.; Hernandez, V. Nutrient Composition and Toxic Factor Content of Four Wild Species of Mexican Potato. J. Agric. Food Chem. 1998, 46, 1355–1358. [Google Scholar] [CrossRef]
- Van Gelder, W.M.J.; Vonk, C.R. Amino acid composition of coagulable protein from tubers of 34 potato varieties and its relationship with protein content. Potato Res. 1980, 23, 427–434. [Google Scholar] [CrossRef]
- Kärenlampi, S.O.; White, P.J. Chapter 5—Potato proteins, lipids, and minerals. In Advances in Potato Chemistry and Technology; Academic Press: San Diego, CA, USA, 2009; pp. 99–125. [Google Scholar]
- Jin, C.-Y.; Zeng, F.-K.; Liu, G. Recovery of Protease Inhibitors from Potato Fruit Water by Expanded Bed Adsorption Chromatography in Pilot Scale. Am. J. Potato Res. 2017, 95, 1–8. [Google Scholar] [CrossRef]
- Lavoie, J.L.; Sigmund, C.D. Minireview: Overview of the Renin-Angiotensin System—An Endocrine and Paracrine System. Endocrinology 2003, 144, 2179–2183. [Google Scholar] [CrossRef]
- Miyamoto, H.; Ito, K.; Ito, K.; Wakabayashi, S.; Suzaka, H.; Matsuo, H.; Iga, T.; Sawada, Y. Comparative study of effects of angiotensin II receptor antagonist, KD3-671, and angiotensin converting enzyme inhibitor, enalaprilat, on cough reflex in guinea pig. Eur. J. Drug Metab. Pharmacokinet. 2001, 26, 47–52. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Bioactive Peptides. J. Aoac Int. 2010, 91, 914–931. [Google Scholar] [CrossRef]
- Guang, C.; Phillips, R.D.; Jiang, B.; Milani, F. Three key proteases angiotensin-I-converting enzyme (ACE), ACE2 and renin within and beyond the renin-angiotensin system. Arch. Cardiovasc. Dis. 2012, 105, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Gerocarni, B.; Laghi, L.; Borghi, C. Blood pressure lowering effect of lactotripeptides assumed as functional foods: A meta-analysis of current available clinical trials. J. Hum. Hypertens. 2010, 25, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, S.; Streng, T.; Larsen, L.B.; Laine, A.; Pihlanto, A. Angiotensin I-converting enzyme inhibitory and antihypertensive properties of potato and rapeseed protein-derived peptides. J. Funct. Foods 2016, 25, 160–173. [Google Scholar] [CrossRef]
- Hill, A.J.; Peikin, S.R.; Ryan, C.A.; Blundell, J.E. Oral administration of proteinase inhibitor II from potatoes reduces energy intake in man. Physiol. Behav. 1990, 48, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Edmondson, B.; Radosevich, J. Potato Proteinase Inhibitor II Exhibits Activity in Elevating Fasting Plasma Cholecystokinin Concentrations. U.S. Patent Application No. 20060204567, 14 September 2006. [Google Scholar]
- Ryan, C.A.; Hass, G.M.; Kuhn, R.W. Purification and Properties of a Carboxypeptidase Inhibitor from Potatoes. J. Biol. Chem. 1974, 249, 5495–5499. [Google Scholar] [CrossRef]
- Bryant, J.; Green, T.R.; Gurusaddaiah, T.; Ryan, C.A. Proteinase inhibitor II from potatoes: Isolation and characterization of its protomer components. Biochemistry 1976, 15, 3418–3424. [Google Scholar] [CrossRef]
- Barrette, I.H.; Ng, K.S.K.; Cherney, M.M.; Pearce, G.J. Chemistry unbound form of tomato inhibitor reveals interdomain flexibility, conformational variability in the reactive site loops. J. Biol. Chem. 2003, 278, 31391–31400. [Google Scholar] [CrossRef]
- AGambuti; Rinaldi, A.; Moio, L. Use of patatin, a protein extracted from potato, as alternative to animal proteins in fining of red wine. Eur. Food Res. Technol. 2012, 235, 753–765. [Google Scholar] [CrossRef]
- Nagashima, M.; Werner, M.; Wang, M.; Zhao, L.; Light, D.R.; Pagila, R.; Morser, J.; Verhallen, P. An Inhibitor of Activated Thrombin-Activatable Fibrinolysis Inhibitor Potentiates Tissue-Type Plasminogen Activator-Induced Thrombolysis in a Rabbit Jugular Vein Thrombolysis Model. Thromb. Res. 2000, 98, 333–342. [Google Scholar] [CrossRef]
- Schneider, M.; Nesheim, M. Reversible inhibitors of TAFIa can both promote and inhibit fibrinolysis. J. Thromb. Haemost. 2003, 1, 147–154. [Google Scholar] [CrossRef]
- John, B.W.; Bernadette, H.; Ian, J.; Peter, H.; Cornelis, K.; Laszlo, B. Stabilization versus inhibition of TAFIa by competitive inhibitors in vitro. J. Biol. Chem. 2003, 278, 8913–8921. [Google Scholar]
- Frenkel, K.; Chrzan, K.; Ryan, C.A.; Wiesner, R.; Troll, W. Chymotrypsin-specific protease inhibitors decrease H2O2 formation by activated human polymorphonuclear leukocytes. Carcinogenesis 1987, 8, 1207–1212. [Google Scholar] [CrossRef]
- Huang, C.; Ma, W.Y.; Ryan, C.A.; Dong, Z. Proteinase inhibitors I and II from potatoes specifically block UV-induced ac-tivator protein-1 activation through a pathway that is independent of extracellular signal-regulated kinases, c-Jun N-terminal kinases, and P38 kinase. Proc. Natl. Acad. Sci. USA 1997, 94, 11957–11962. [Google Scholar] [CrossRef]
- Park, Y.; Choi, B.H.; Kwak, J.-S.; Kang, C.-W.; Lim, H.-T.; Cheong, H.-S.; Hahm, K.-S. Kunitz-Type Serine Protease Inhibitor from Potato. J. Agric. Food Chem. 2005, 53, 6491–6496. [Google Scholar] [CrossRef]
- Tripathi, G.R.; Park, J.; Park, Y.; Hwang, I.; Cheong, H. Potide-G derived from potato (Solanum tuberosum L.) is active against potato virus infection. J. Agric. Food Chem. 2006, 54, 8437–8443. [Google Scholar] [CrossRef]
- Albishi, T.; John, J.A.; Al-Khalifa, A.S.; Shahidi, F. Antioxidant, anti-inflammatory and DNA scission inhibitory activities of phenolic compounds in selected onion and potato varieties. J. Funct. Foods 2013, 5, 930–939. [Google Scholar] [CrossRef]
- Müller, H. Determination of the carotenoid content in selected vegetables and fruit by HPLC and photodiode array detection. Z. Lebensm. Forsch. A 1997, 204, 88–94. [Google Scholar] [CrossRef]
- Nesterenko, S.; Sink, K.C. Carotenoid Profiles of Potato Breeding Lines and Selected Cultivars. Hortscience 2003, 38, 1173–1177. [Google Scholar] [CrossRef]
- Burgos, G.; Salas, E.; Amoros, W.; Auqui, M.; Muñoa, L.; Kimura, M.; Bonierbale, M. Total and individual carotenoid profiles in Solanum phureja of cultivated potatoes: I. Concentrations and relationships as determined by spectrophotometry and HPLC. J. Food Compos. Anal. 2009, 22, 503–508. [Google Scholar] [CrossRef]
- Kobayashi, A.; Ohara-Takada, A.; Tsuda, S.; Matsuura-Endo, C.; Takada, N.; Umemura, Y.; Nakao, T.; Yoshida, T.; Hayashi, K.; Mori, M. Breeding of potato variety “Inca-no-hitomi” with a very high carotenoid content. Breed. Sci. 2008, 58, 77–82. [Google Scholar] [CrossRef]
- Hossain, A.; Begum, P.; Zannat, M.S.; Rahman, M.H.; Ahsan, M.; Islam, S.N. Nutrient composition of strawberry genotypes cul-tivated in a horticulture farm. Food Chem. 2016, 199, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, K.; Rangasamy, R.; Krishnan, V. Total antioxidant activity and radical scavenging capacity of selected fruits and vegetables from south India. Int. Food Res. J. 2014, 21, 1003–1007. [Google Scholar]
- Chassy, A.W.; Bui, L.; Renaud, E.N.C.; Van Horn, M.; Mitchell, A.E. Three-year comparison of the content of antioxidant micro-constituents and several quality characteristics in organic and conventionally managed tomatoes and bell peppers. J. Agric. Food Chem. 2006, 54, 8244–8252. [Google Scholar] [CrossRef]
- Valcarcel, J.; Reilly, K.; Gaffney, M.; O’brien, N. Total Carotenoids and l-Ascorbic Acid Content in 60 Varieties of Potato (Solanum tuberosum L.) Grown in Ireland. Potato Res. 2014, 58, 29–41. [Google Scholar] [CrossRef]
- Fernandez-Orozco, R.; Gallardo-Guerrero, L.; Hornero-Mendez, D. Carotenoid profiling in tubers of different potato (Solarium sp) cultivars: Accumulation of carotenoids mediated by xanthophyll esterification. Food Chem. 2013, 141, 2864–2872. [Google Scholar] [CrossRef]
- Brown, C.R. Antioxidants in potato. Am. J. Potato Res. 2005, 82, 163–172. [Google Scholar] [CrossRef]
- Fraser, P.D.; Bramley, P.M. The biosynthesis and nutritional uses of carotenoids. Prog. Lipid Res. 2004, 43, 228–265. [Google Scholar] [CrossRef]
- Gammone, M.A.; Riccioni, G.; D’Orazio, N. Carotenoids: Potential allies of cardiovascular health. Food Nutr. Res. 2015, 59, 26762–26771. [Google Scholar]
- Abdel-Aal, E.-S.M.; Akhtar, H.; Zaheer, K.; Ali, R. Dietary Sources of Lutein and Zeaxanthin Carotenoids and Their Role in Eye Health. Nutrients 2013, 5, 1169–1185. [Google Scholar] [CrossRef]
- Chucair, A.J.; Rotstein, N.P.; SanGiovanni, J.P.; During, A.; Chew, E.Y.; Politi, L.E. Lutein and Zeaxanthin Protect Photoreceptors from Apoptosis Induced by Oxidative Stress: Relation with Docosahexaenoic Acid. Investig. Opthalmol. Vis. Sci. 2007, 48, 5168–5177. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.S.; Wang, J.J.; Flood, V.; Rochtchina, E.; Smith, W.; Mitchell, P. Dietary Antioxidants and the Long-term Incidence of Age-Related Macular Degeneration. Ophthalmology 2008, 115, 334–341. [Google Scholar] [CrossRef]
- Lu, S.; Li, L. Carotenoid metabolism: Biosynthesis, regulation, and beyond. J. Integr. Plant Biol. 2008, 50, 778–785. [Google Scholar] [CrossRef]
- Cazzonelli, C.; Pogson, B. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef]
- Shewmaker, C.K.; Sheehy, J.A.; Daley, M.; Colburn, S.; Ke, D.Y. Seed-specific overexpression of phytoene synthase: Increase in carotenoids and other metabolic effects. Plant J. 1999, 20, 401–412. [Google Scholar] [CrossRef]
- Fraser, P.D.; Romer, S.; Shipton, C.A.; Mills, P.B.; Kiano, J.W.; Misawa, N.; Drake, R.G.; Schuch, W.; Bramley, P.M. Evaluation of transgenic tomato plants expressing an additional phytoene synthase in a fruit-specific manner. Proc. Natl. Acad. Sci. USA 2001, 99, 1092–1097. [Google Scholar] [CrossRef]
- Ducreux, L.J.; Morris, W.; Hedley, L.; Shepherd, P.E. Metabolic engineering of high carotenoid potato tubers containing enhanced levels of b-carotene and lutein. J. Exp. Bot. 2005, 56, 81–89. [Google Scholar] [CrossRef]
- Kato, M.; Matsumoto, H.; Ikoma, Y.; Okuda, H.; Yano, M. The role of carotenoid cleavage dioxygenases in the regulation of carotenoid profiles during maturation in citrus fruit. J. Exp. Bot. 2006, 57, 2153–2164. [Google Scholar] [CrossRef]
- Campbell, R.; Ducreux, L.J.; Morris, W.L.; Morris, J.A.; Suttle, J.C.; Ramsay, G.; Bryan, G.J.; Hedley, P.E.; Taylor, M.A. The Metabolic and Developmental Roles of Carotenoid Cleavage Dioxygenase4 from Potato. Plant Physiol. 2010, 154, 656–664. [Google Scholar] [CrossRef]
- Haynes, K.G.; Clevidence, B.A.; Rao, D.; Vinyard, B.T.; White, J.M. Genotype environment interactions for potato tuber carotenoid content. J. Am. Soc. Hortic. Sci. Am. Soc. Hortic. Sci. 2010, 135, 250–258. [Google Scholar] [CrossRef]
- Brown, C.R.; Kim, T.S.; Ganga, Z.; Haynes, K.; De Jong, D.; Jahn, M.; Paran, I. Segregation of total carotenoid in high level potato germplasm and its relationship to beta-carotene hydroxylase polymorphism. Am. J. Potato Res. 2006, 83, 365–372. [Google Scholar] [CrossRef]
- Wolters, A.M.A.; Uitdewilligen, J.G.; Kloosterman, B.A.; Hutten, R.C.B.; Visser, R.G.F.; Eck, H.J.V. Identification of alleles of ca-rotenoid pathway genes important for zeaxanthin accumulation in potato tubers. Plant Mol. Biol. 2010, 73, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.; Pont, S.D.A.; Morris, J.A.; McKenzie, G.; Sharma, S.K.; Hedley, P.E.; Ramsay, G.; Bryan, G.J.; Taylor, M.A. Genome-wide QTL and bulked transcriptomic analysis reveals new candidate genes for the control of tuber carotenoid content in potato (Solanum tuberosum L.). Theor. Appl. Genet. 2014, 127, 1917–1933. [Google Scholar] [CrossRef] [PubMed]
- Sulli, M.; Mandolino, G.; Sturaro, M.; Onofri, C.; Diretto, G.; Parisi, B.; Giuliano, G. Molecular and biochemical characterization of a potato collection with contrasting tuber carotenoid content. PLoS ONE 2017, 12, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Diretto, G.; Al-Babili, S.; Tavazza, R.; Papacchioli, V.; Beyer, P.; Giuliano, G. Metabolic Engineering of Potato Carotenoid Content through Tuber-Specific Overexpression of a Bacterial Mini-Pathway. PLoS ONE 2007, 2, 350–361. [Google Scholar] [CrossRef]
- Chitchumroonchokchai, C.; Diretto, G.; Parisi, B.; Giuliano, G.; Failla, M.L. Potential of golden potatoes to improve vitamin A and vitamin E status in developing countries. PLoS ONE 2017, 12, 187–195. [Google Scholar] [CrossRef]
- Im, H.W.; Suh, B.-S.; Lee, S.-U.; Kozukue, N.; Ohnisi-Kameyama, M.; Levin, C.E.; Friedman, M. Analysis of Phenolic Compounds by High-Performance Liquid Chromatography and Liquid Chromatography/Mass Spectrometry in Potato Plant Flowers, Leaves, Stems, and Tubers and in Home-Processed Potatoes. J. Agric. Food Chem. 2008, 56, 3341–3349. [Google Scholar] [CrossRef]
- Bassoli, B.K.; Cassolla, P.; Borba-Murad, G.R.; Constantin, J.; Salgueiro-Pagadigorria, C.L.; Bazotte, R.B.; Silva, R.S.D.S.F.D.; de Souza, H.M. Chlorogenic acid reduces the plasma glucose peak in the oral glucose tolerance test: Effects on hepatic glucose release and glycaemia. Cell Biochem. Funct. 2007, 26, 320–328. [Google Scholar] [CrossRef]
- Niggeweg, R.; Michael, A.J.; Martin, C. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol. 2004, 22, 746–754. [Google Scholar] [CrossRef]
- Payyavula, R.S.; Shakya, R.; Sengoda, V.G.; Munyaneza, J.E.; Swamy, P.; Navarre, D.A. Synthesis and regulation of chlorogenic acid in potato: Rerouting phenylpropanoid flux in HQT-silenced lines. Plant Biotechnol. J. 2014, 13, 551–564. [Google Scholar] [CrossRef]
- Payyavula, R.S.; Singh, R.K.; Navarre, D.A. Transcription factors, sucrose, and sucrose metabolic genes interact to regulate potato phenylpropanoid metabolism. J. Exp. Bot. 2013, 64, 5115–5131. [Google Scholar] [CrossRef]
- Navarre, D.A.; Brown, C.R.; Sathuvalli, V.R. Potato Vitamins, Minerals and Phytonutrients from a Plant Biology Perspective. Am. J. Potato Res. 2019, 96, 111–126. [Google Scholar] [CrossRef]
- Andre, C.M.; Ghislain, M.; Bertin, P.; Oufir, M.; Herrera, M.D.R.; Hoffmann, L.; Hausman, J.-F.; Larondelle, A.Y.; Evers, D. Andean Potato Cultivars (Solanum tuberosum L.) as a Source of Antioxidant and Mineral Micronutrients. J. Agric. Food Chem. 2007, 55, 366–378. [Google Scholar] [CrossRef]
- Ayvaz, H.; Bozdogan, A.; Giusti, M.M.; Mortas, M.; Gomez, R.; Rodriguez-Saona, L.E. Improving the screening of potato breeding lines for specific nutritional traits using portable mid-infrared spectroscopy and multivariate analysis. Food Chem. 2016, 211, 374–382. [Google Scholar] [CrossRef]
- Valiñas, M.A.; Lanteri, M.L.; Have, A.T.; Andreu, A.B. Chlorogenic acid, anthocyanin and flavan-3-ol biosynthesis in flesh and skin of Andean potato tubers (Solanum tuberosum subsp. andigena). Food Chem. 2017, 229, 837–846. [Google Scholar] [CrossRef]
- Kong, A.H.; Carolina, F.; Susan, H.; Andrés, C.; Antonio, V.G.; Roberto, L.M. Antioxidant capacity and total phenolic compounds of twelve selected potato landrace clones grown in Southern Chile. Chil. J. Agric. Res. 2012, 72, 3–9. [Google Scholar]
- Tudela, J.A.; Cantos, E.; Espín, J.C.; Tomás-Barberán, F.A.; Gil, M.I. Chemistry, induction of antioxidant flavonol bio-synthesis in fresh-cut potatoes. Effect of Domestic Cooking. J. Agric. Food Chem. 2002, 50, 5925–5931. [Google Scholar] [CrossRef]
- Kawabata, K.; Mukai, R.; Ishisaka, A. Quercetin and related polyphenols: New insights and implications for their bioactivity and bioavailability. Food Funct. 2015, 6, 1399–1417. [Google Scholar] [CrossRef]
- Yıldırım, S.; Kadıoğlu, A.; Sağlam, A.; Yaşar, A.; Sellitepe, H.E. Fast determination of anthocyanins and free pelargonidin in fruits, fruit juices, and fruit wines by high-performance liquid chromatography using a core-shell column. J. Sep. Sci. 2016, 39, 3927–3935. [Google Scholar] [CrossRef]
- Wu, X.; Pittman, H.E.; Mckay, S.; Prior, R.L. Aglycones and Sugar Moieties Alter Anthocyanin Absorption and Metabolism after Berry Consumption in Weanling Pigs. J. Nutr. 2005, 135, 2417–2424. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Sun, Y.; Huang, Y.; He, J.; Zhu, Z. Transport of flavanolic monomers and procyanidin dimer A2 across human adeno-carcinoma stomach cells (MKN-28). J. Agric. Food Chem. 2019, 67, 3354–3362. [Google Scholar] [CrossRef] [PubMed]
- Jansen, G.; Flamme, W. Coloured potatoes (Solanum tuberosum L.)—Anthocyanin content and tuber quality. Genet. Resour. Crop Evol. 2006, 53, 1321–1331. [Google Scholar] [CrossRef]
- Jung, C.S.; Griffiths, H.M.; Jong, D.M.D.; Cheng, S.; Bodis, M.; Jong, W.S.D. The potato P locus codes for flavonoid 3′,5′-hydroxylase. Theor. Appl. Genet. 2005, 110, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.S.; Griffiths, H.M.; De Jong, D.M.; Cheng, S.; Bodis, M.; Kim, T.S.; De Jong, W.S. The potato developer (D) locus encodes an R2R3 MYB transcription factor that regulates expression of multiple anthocyanin structural genes in tuber skin. Theor. Appl. Genet. 2009, 120, 45–57. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, S.; De Jong, D.; Griffiths, H.; Halitschke, R.; De Jong, W. The potato R locus codes for dihydroflavonol 4-reductase. Theor. Appl. Genet. 2009, 119, 931–937. [Google Scholar] [CrossRef]
- Payyavula, R.S.; Navarre, D.A.; Kuhl, J.; Pantoja, A. Developmental Effects on Phenolic, Flavonol, Anthocyanin, and Carotenoid Metabolites and Gene Expression in Potatoes. J. Agric. Food Chem. 2013, 61, 7357–7365. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, B.; Liu, J.; Guo, D.; Hou, J.; Chen, S.; Song, B.; Xie, C. Analysis of structural genes and key transcription factors related to anthocyanin biosynthesis in potato tubers. Sci. Hortic. 2017, 225, 310–316. [Google Scholar] [CrossRef]
- Kyoungwon, C.; Kwang-Soo, C.; Hwang-Bae, S.; Jin, H.I.; Su-Young, H.; Hyerim, L.; Young-Mi, K.; Hee, N.M. Network analysis of the metabolome and transcriptome reveals novel regulation of potato pigmentation. J. Exp. Bot. 2016, 67, 1515–1533. [Google Scholar]
- Liu, Y.; Lin-Wang, K.; Deng, C.; Warran, B.; Wang, L.; Yu, B.; Yang, H.; Wang, J.; Espley, R.V.; Zhang, J.; et al. Comparative Transcriptome Analysis of White and Purple Potato to Identify Genes Involved in Anthocyanin Biosynthesis. PLoS ONE 2015, 10, e0129148. [Google Scholar] [CrossRef]
- Stushnoff, C.; Ducreux, L.J.M.; Hancock, R.D.; Hedley, P.E.; Holm, D.G.; McDougall, G.J.; McNicol, J.W.; Morris, J.; Morris, W.L.; Sungurtas, J.A.; et al. Flavonoid profiling and transcriptome analysis reveals new gene–metabolite correlations in tubers of Solanum tuberosum L. J. Exp. Bot. 2010, 61, 1225–1238. [Google Scholar] [CrossRef]
- Tierno, R.; Ignacio, R.D.G. Jose, Characterization of high anthocyanin-producing tetraploid potato cultivars selected for breeding using morphological traits and microsatellite markers. Plant Genet. Resour. 2015, 15, 147–156. [Google Scholar] [CrossRef]
- Charepalli, V.; Reddivari, L.; Radhakrishnan, S.; Vadde, R.; Agarwal, R.; Vanamala, J.K. Anthocyanin-containing purple-fleshed potatoes suppress colon tumorigenesis via elimination of colon cancer stem cells. J. Nutr. Biochem. 2015, 26, 1641–1649. [Google Scholar] [CrossRef]
- Ombra, M.N.; Fratianni, F.; Granese, T.; Cardinale, F.; Cozzolino, A.; Nazzaro, F. In vitro antioxidant, antimicrobial and an-ti-proliferative activities of purple potato extracts (Solanum tuberosum cv Vitelotte noire) following simulated gastro-intestinal digestion. Nat. Prod. Res. 2015, 29, 1087–1091. [Google Scholar] [CrossRef]
- Sido, A.; Radhakrishnan, S.; Kim, S.W.; Eriksson, E.; Shen, F.; Li, Q.; Bhat, V.; Reddivari, L.; Vanamala, J.K. A food-based approach that targets interleukin-6, a key regulator of chronic intestinal inflammation and colon carcinogenesis. J. Nutr. Biochem. 2017, 43, 11–17. [Google Scholar] [CrossRef]
- Reddivari, L.; Vanamala, J.; Chintharlapalli, S.; Safe, S.H.; Miller, J.C., Jr. Anthocyanin fraction from potato extracts is cytotoxic to prostate cancer cells through activation of caspa-se-dependent and caspase-independent pathways. Carcinogenesis 2007, 28, 2227–2235. [Google Scholar] [CrossRef]
- Zhao, X.; Sheng, F.; Zheng, J.; Liu, R. Composition and Stability of Anthocyanins from Purple Solanum tuberosum and Their Protective Influence on Cr(VI) Targeted to Bovine Serum Albumin. J. Agric. Food Chem. 2011, 59, 7902–7909. [Google Scholar] [CrossRef]
- Jiang, Z.; Chen, C.; Wang, J.; Xie, W.; Wang, M.; Li, X.; Zhang, X. Purple potato (Solanum tuberosum L.) anthocyanins attenuate alcohol-induced hepatic injury by enhancing antioxidant defense. J. Nat. Med. 2015, 70, 45–53. [Google Scholar] [CrossRef]
- Kaspar, K.L.; Park, J.S.; Brown, C.R.; Mathison, B.D.; Navarre, D.A.; Chew, B.P. Pigmented Potato Consumption Alters Oxidative Stress and Inflammatory Damage in Men. J. Nutr. 2011, 141, 108–111. [Google Scholar] [CrossRef]
- Vinson, J.A.; Demkosky, C.A.; A Navarre, D.; Smyda, M.A. High-Antioxidant Potatoes: Acute in Vivo Antioxidant Source and Hypotensive Agent in Humans after Supplementation to Hypertensive Subjects. J. Agric. Food Chem. 2012, 60, 6749–6754. [Google Scholar] [CrossRef]
- Ramdath, D.D.; Padhi, E.; Hawke, A.; Sivaramalingam, T.; Tsao, R. The glycemic index of pigmented potatoes is related to their polyphenol content. Food Funct. 2014, 5, 909–915. [Google Scholar] [CrossRef]
- Ayoub, H.M.; McDonald, M.R.; Sullivan, J.A.; Tsao, R.; Platt, M.; Simpson, J.; Meckling, K.A. The Effect of Anthocyanin-Rich Purple Vegetable Diets on Metabolic Syndrome in Obese Zucker Rats. J. Med. Food 2017, 20, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- Viola, R.; Vreugdenhil, D.; Davies, H.; Sommerville, L. Accumulation of L-ascorbic acid in tuberising stolon tips of potato (Solanum tuberosum L). J. Plant Physiol. 1998, 152, 58–63. [Google Scholar] [CrossRef]
- Tedone, L.; Hancock, R.D.; Alberino, S.; Haupt, S.; Viola, R. Long-distance transport of L-ascorbic acid in potato. BMC Plant Biol. 2004, 4, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Cabezas-Serrano, A.B.; Amodio, M.L.; Cornacchia, R.; Rinaldi, R.; Colelli, G. Suitability of five different potato cultivars (Solanum tuberosum L.) to be processed as fresh-cut products. Postharvest Biol. Technol. 2009, 53, 138–144. [Google Scholar] [CrossRef]
- Kaur, S.; Aggarwal, P. Evaluation of antioxidant phytochemicals in different genotypes of potato. Int. J. Eng. Res. Appl. 2014, 4, 167–172. [Google Scholar]
- Adelanwa, E.B.; Medugu, J.M. Variation in the nutrition composition of red and green cabbge (Brassica oleracea) with respect to age at harvest. Sci. Hortic. 2015, 7, 183–189. [Google Scholar]
- Bhandari, S.R.; Chae, Y.; Lee, J.G. Assessment of Phytochemicals, Quality Attributes, and Antioxidant Activities in Commercial Tomato Cultivars. Korean J. Hortic. Sci. 2016, 34, 677–691. [Google Scholar]
- Love, S.L.; Salaiz, T.; Shafii, B.; Price, W.J.; Mosley, A.R.; Thornton, R.E. Stability of Expression and Concentration of Ascorbic Acid in North American Potato Germplasm. Hortscience 2004, 39, 156–160. [Google Scholar] [CrossRef]
- Love, S.L.; Pavek, J.J. Positioning the Potato as a Primary Food Source of Vitamin C. Am. J. Potato Res. 2008, 85, 277–285. [Google Scholar] [CrossRef]
- Keijbets, M.J.H.; Ebbenhorst-Seller, G. Loss of vitamin C (L-ascorbic acid) during long-term cold storage of Dutch table potatoes. Potato Res. 1990, 33, 125–130. [Google Scholar] [CrossRef]
- Dale, M.F.B.; Griffiths, D.W.; Todd, D.T. Effects of genotype, environment, and postharvest storage on the total ascorbate content of potato (Solanum tuberosum) tubers. J. Agric. Food Chem. 2003, 51, 244–248. [Google Scholar] [CrossRef]
- Külen, O.; Stushnoff, C.; Holm, D.G. Effect of cold storage on total phenolics content, antioxidant activity and vitamin C level of selected potato clones. J. Sci. Food Agric. 2013, 93, 2437–2444. [Google Scholar] [CrossRef]
- Blauer, J.M.; Kumar, G.M.; Knowles, L.O.; Dhingra, A.; Knowles, N. Changes in ascorbate and associated gene expression during development and storage of potato tubers (Solanum tuberosum L.). Postharvest Biol. Technol. 2013, 78, 76–91. [Google Scholar] [CrossRef]
- Tosun, B.N.; Yücecan, S. Influence of commercial freezing and storage on vitamin C content of some vegetables. Int. J. Food Sci. Technol. 2007, 43, 316–321. [Google Scholar] [CrossRef]
- Licciardello, F.; Lombardo, S.; Rizzo, V.; Pitino, I.; Pandino, G.; Strano, M.G.; Muratore, G.; Restuccia, C.; Mauromicale, G. Integrated agronomical and technological approach for the quality maintenance of ready-to-fry potato sticks during refrigerated storage. Postharvest Biol. Technol. 2018, 136, 23–30. [Google Scholar] [CrossRef]
- Amado, I.R.; Franco, D.; Sanchez, M.; Zapata, C.; Vazquez, J.A. Optimisation of antioxidant extraction from Solarium tuberosum potato peel waste by surface response methodology. Food Chem. 2014, 165, 290–299. [Google Scholar] [CrossRef]
- López-Cobo, A.; Gómez-Caravaca, A.M.; Cerretani, L.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Distribution of phenolic compounds and other polar compounds in the tuber of Solanum tuberosum L. by HPLC-DAD-q-TOF and study of their an-tioxidant activity. J. Food Compos. Anal. 2014, 36, 1–11. [Google Scholar] [CrossRef]
- Nara, K.; Miyoshi, T.; Honma, T.; Koga, H. Antioxidative Activity of Bound-Form Phenolics in Potato Peel. Biosci. Biotechnol. Biochem. 2006, 70, 1489–1491. [Google Scholar] [CrossRef]
- Williams, D.J.; Edwards, D.; Hamernig, I.; Jian, L.; James, A.P.; Johnson, S.K.; Tapsell, L.C. Vegetables containing phytochemicals with potential anti-obesity properties: A review. Food Res. Int. 2013, 52, 323–333. [Google Scholar] [CrossRef]
- Navarre, D.A.; Payyavula, R.S.; Shakya, R.; Knowles, N.R.; Pillai, S.S. Changes in potato phenylpropanoid metabolism during tuber development. Plant Physiol. Biochem. 2013, 65, 89–101. [Google Scholar] [CrossRef]
- Tian, J.; Chen, J.; Lv, F.; Chen, S.; Chen, J.; Liu, D.; Ye, X. Domestic cooking methods affect the phytochemical composition and antioxidant activity of purple-fleshed potatoes. Food Chem. 2015, 197, 1264–1270. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, W.; Lu, Z.; Beta, T.; Hydamaka, A.W. Phenolic Content, Composition, Antioxidant Activity, and Their Changes during Domestic Cooking of Potatoes. J. Agric. Food Chem. 2009, 57, 10231–10238. [Google Scholar] [CrossRef] [PubMed]
- Blessington, T.; Nzaramba, M.N.; Scheuring, D.C.; Hale, A.L.; Reddivari, L.; Miller, J.C. Cooking Methods and Storage Treatments of Potato: Effects on Carotenoids, Antioxidant Activity, and Phenolics. Am. J. Potato Res. 2010, 87, 479–491. [Google Scholar] [CrossRef]
- Faller, A.; Fialho, E. The antioxidant capacity and polyphenol content of organic and conventional retail vegetables after domestic cooking. Food Res. Int. 2009, 42, 210–215. [Google Scholar] [CrossRef]
- Burgos, G.; Amoros, W.; Muñoa, L.; Sosa, P.; Cayhualla, E.; Sanchez, C.; Díaz, C.; Bonierbale, M. Total phenolic, total anthocyanin and phenolic acid concentrations and antioxidant activity of purple-fleshed potatoes as affected by boiling. J. Food Compos. Anal. 2013, 30, 6–12. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Tao, C.; Liu, M.; Pan, Y.; Lv, Z. Effect of temperature and pH on stability of anthocyanin obtained from blueberry. J. Food Meas. Charact. 2018, 12, 1744–1753. [Google Scholar] [CrossRef]
- Blancquaert, D.; Storozhenko, S.; Loizeau, K.; De Steur, H.; De Brouwer, V.; Viaene, J.; Ravanel, S.; Rébeillé, F.; Lambert, W.; Van Der Straeten, D. Folates and Folic Acid: From Fundamental Research Toward Sustainable Health. Crit. Rev. Plant Sci. 2010, 29, 14–35. [Google Scholar] [CrossRef]
- Alfthan, G.; Laurinen, M.S.; Valsta, L.M.; Pastinen, T.; Aro, A. Folate intake, plasma folate and homocysteine status in a random Finnish population. Eur. J. Clin. Nutr. 2003, 57, 81–88. [Google Scholar] [CrossRef]
- Hatzis, C.M.; Bertsias, G.K.; Linardakis, M.; Scott, J.M.; Kafatos, A.G. Dietary and other lifestyle correlates of serum folate con-centrations in a healthy adult population in Crete, Greece: A cross-sectional study. Nutr. J. 2006, 5, 105–113. [Google Scholar] [CrossRef]
- Goyer, A.; Navarre, D.A. Determination of Folate Concentrations in Diverse Potato Germplasm Using a Trienzyme Extraction and a Microbiological Assay. J. Agric. Food Chem. 2007, 55, 3523–3528. [Google Scholar] [CrossRef]
- Goyer, A.; Sweek, K. Genetic Diversity of Thiamin and Folate in Primitive Cultivated and Wild Potato (Solanum) Species. J. Agric. Food Chem. 2011, 59, 13072–13080. [Google Scholar] [CrossRef]
- Kuhn, D.N.; Chappell, J.; Boudet, A.; Hahlbrock, K. Induction of phenylalanine ammonia-lyase and 4-coumarate: CoA ligase mRNAs in cultured plant cells by UV light or fungal elicitor. Proc. Natl. Acad. Sci. USA 1984, 81, 1102–1106. [Google Scholar] [CrossRef]
- Mooney, S.; Chen, L.; Kühn, C.; Navarre, R.; Knowles, N.R.; Hellmann, H. Genotype-Specific Changes in Vitamin B6 Content and the PDX Family in Potato. BioMed Res. Int. 2013, 2013, 389723. [Google Scholar] [CrossRef]
- Holland, B.; Unwin, I.D.; Buss, D.H. Vegetables, herbs and spices. Nutr. Bull. 1991, 16, 386–414. [Google Scholar]
- Goyer, A.; Haynes, K.G. Vitamin B1 Content in Potato: Effect of Genotype, Tuber Enlargement, and Storage, and Estimation of Stability and Broad-Sense Heritability. Am. J. Potato Res. 2011, 88, 374–385. [Google Scholar] [CrossRef]
- Lisinska, G.; Leszczynski, W. Potato Science and Technology; Springer: Berlin/Heidelberg, Germany, 1989; ISBN 978-1-85166-307-1. [Google Scholar]
- Burrowes, J.D.; Ramer, N.J. Changes in the Potassium Content of Different Potato Varieties after Cooking. J. Ren. Nutr. 2008, 18, 249. [Google Scholar] [CrossRef]
- Sanchez-Castillo, C.P.; Dewey, P.J.; Aguirre, A.; Lara, J.J.; Vaca, R.; de la Barra, P.L.; Ortiz, M.; Escamilla, I.; James, W.T. The Mineral Content of Mexican Fruits and Vegetables. J. Food Compos. Anal. 1998, 11, 340–356. [Google Scholar] [CrossRef]
- Novy, C.R. Stability and broad-sense heritability of mineral content in potato: Potassium and phosphorus. Am. J. Potato Res. 2013, 90, 516–523. [Google Scholar]
- Subramanian, N.K.; White, P.J.; Broadley, M.R.; Ramsay, G. Variation in tuber mineral concentrations among accessions of so-lanum species held in the commonwealth potato collection. Genet. Resour. Crop Evol. 2017, 64, 1927–1935. [Google Scholar] [CrossRef]
- Sun, Y.; Byon, C.H.; Yang, Y.; Bradley, W.E.; Dell’Italia, L.J.; Sanders, P.W.; Agarwal, A.; Wu, H.; Chen, Y. Dietary potassium regulates vascular calcification and arterial stiffness. JCI Insight 2017, 2, e94920. [Google Scholar] [CrossRef]
- Hoy, M.K. Dietary fiber intake of the U.S. population, what we eat in America. FASEB J. 2014, 27, 2019–2025. [Google Scholar]
- Storey, M.L.; Anderson, A.P. Nutrient Intakes and Vegetable and White Potato Consumption by Children Aged 1 to 3 Years. Adv. Nutr. Int. Rev. J. 2016, 7, 241S–246S. [Google Scholar] [CrossRef] [PubMed]
- Lewu, M.N.; Adebola, P.O.; Afolayan, A.J. Comparative Assessment of the Nutritional Value of Commercially Available Cocoyam and Potato Tubers in South Africa. J. Food Qual. 2010, 33, 461–476. [Google Scholar] [CrossRef]
- Subramanian, N.K.; White, P.; Broadley, M.; Ramsay, G. The three-dimensional distribution of minerals in potato tubers. Ann. Bot. 2011, 107, 681–691. [Google Scholar] [CrossRef] [PubMed]
- Furrer, A.N.; Chegeni, M.; Ferruzzi, M.G. Impact of potato processing on nutrients, phytochemicals and human health. Crit. Rev. Food Sci. Nutr. 2016, 58, 168–173. [Google Scholar] [CrossRef]
- Wegener, C.B.; Jurgens, H.-U.; Jansen, G. Drought stress affects bioactive compounds in potatoes (Solanum tuberosum L.) relevant to non-communicable diseases. Funct. Foods Health Dis. 2017, 7, 17–35. [Google Scholar] [CrossRef]
- WHO. Micronutrient Deficiencies. 2017. Available online: https://www.who.int/nutrition/topics/vad/en/ (accessed on 21 April 2023).
- Brown, C.R.; Haynes, K.G.; Moore, M.; Pavek, M.J.; Hane, D.C.; Love, S.L.; Novy, R.G.; Miller, J.C. Stability and broad-sense her-itability of mineral content in potato: Iron. Am. J. Potato Res. 2010, 87, 390–396. [Google Scholar] [CrossRef]
- Paget, M.; Amoros, W.; Salas, E.; Eyzaguirre, R.; Alspach, P.; Apiolaza, L.; Noble, A.; Bonierbale, M. Genetic Evaluation of Micronutrient Traits in Diploid Potato from a Base Population of Andean Landrace Cultivars. Crop. Sci. 2014, 54, 1949–1959. [Google Scholar] [CrossRef]
- True, R.H.; Hogan, J.M.; Augustin, J.; Johnson, S.J.; Teitzel, C.; Toma, R.B.; Shaw, R.L. Mineral composition of freshly harvested potatoes. Am. J. Potato Res. 1978, 55, 511–519. [Google Scholar] [CrossRef]
- Scurrah, M.; Amoros, W.; Burgos, G.; Schafleitner, R.; Bonierbale, M. Back to the Future: Millennium Traits in Native Varieties. Acta Hortic. 2007, 745, 369–378. [Google Scholar] [CrossRef]
- Sales, C.H.; Pedrosa, L.D.F.C. Magnesium and diabetes mellitus: Their relation. Clin. Nutr. 2006, 25, 554–562. [Google Scholar] [CrossRef]
- Dibaba, D.T.; Xun, P.; He, K. Dietary magnesium intake is inversely associated with serum C-reactive protein levels: Meta-analysis and systematic review. Eur. J. Clin. Nutr. 2014, 68, 971. [Google Scholar] [CrossRef]
- Ricardo, C.M.; González, E.; Rodríguez, A.; Marrero, D. Carlos, Chemometric studies of chemical compounds in five cultivars of potatoes from Tenerife. J. Agric. Food Chem. 2002, 50, 2076–2082. [Google Scholar]
- Ramírez-Jiménez, A.; Guerra-Hernández, E.; García-Villanova, B. Evolution of non-enzymatic browning during storage of infant rice cereal. Food Chem. 2003, 83, 219–225. [Google Scholar] [CrossRef]
- Peña, C.; Restrepo-Sánchez, L.-P.; Kushalappa, A.; Rodríguez-Molano, L.-E.; Mosquera, T.; Narváez-Cuenca, C.-E. Nutritional contents of advanced breeding clones of Solanum tuberosum group Phureja. LWT 2015, 62, 76–82. [Google Scholar] [CrossRef]
- Tein, B.; Kauer, K.; Eremeev, V.; Luik, A.; Selge, A.; Loit, E. Farming systems affect potato (Solanum tuberosum L.) tuber and soil quality. Field Crop. Res. 2014, 156, 1–11. [Google Scholar] [CrossRef]
- Navarre, D.; Goyer, A.; Payyavula, R.; Hellmann, H.; Navarre, R.; Pavek, M.J. Nutritional Characteristics of Potatoes; CABI: Oxfordshire, UK, 2014; ISBN 9781780642802. [Google Scholar]
- Randhawa, K.S.; Sandhu, K.S.; Kaur, G.; Singh, D. Studies of the evaluation of different genotypes of potato (Solanum tuberosum L.) for yield and mineral contents. Plant Foods Hum. Nutr. 1984, 34, 239–242. [Google Scholar] [CrossRef]
- Tamasi, G.; Cambi, M.; Gaggelli, N.; Autino, A.; Cresti, M.; Cini, R. The content of selected minerals and vitamin C for potatoes (Solanum tuberosum L.) from the high Tiber Valley area, southeast Tuscany. J. Food Compos. Anal. 2015, 41, 157–164. [Google Scholar] [CrossRef]
- Potarzycki, J.; Grzebisz, W. Trends in phosphorus concentrations in potato organs during the growing season. J. Elem. 2019, 24, 1777–1783. [Google Scholar] [CrossRef]
- Pursglove, J.; Sanders, F. The growth and phosphorus economy of the early potato (Solanum tuberosum). Commun. Soil Sci. Plant Anal. 1981, 12, 1105–1121. [Google Scholar] [CrossRef]
- Mottram, D.S.; Wedzicha, B.L.; Dodson, A.T. Acrylamide is formed in the Maillard reaction. Nature 2002, 419, 448–449. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Luo, Y.; Sun, G.; Wang, P.; Hu, X.; Chen, F. Inhibition of acrylamide by glutathione in asparagine/glucose model systems and cookies. Food Chem. 2020, 329, 127171. [Google Scholar] [CrossRef] [PubMed]
- Zyzak, D.V.; Sanders, R.A.; Stojanovic, M.; Tallmadge, D.H.; Eberhart, B.L.; Ewald, D.K.; Gruber, D.C.; Morsch, T.R.; Strothers, M.A.; Rizzi, G.P.; et al. Acrylamide Formation Mechanism in Heated Foods. J. Agric. Food Chem. 2003, 51, 4782–4787. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.J.; Mottram, D.S.; Curtis, A. Acrylamide concentrations in potatocrisps in Europe from 2002 to 2011. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2013, 9, 30. [Google Scholar]
- Maan, A.A.; Anjum, M.A.; Khan, M.K.I.; Nazir, A.; Saeed, F.; Afzaal, M.; Aadil, R.M. Acrylamide Formation and Different Mitigation Strategies during Food Processing—A Review. Food Rev. Int. 2020, 38, 70–87. [Google Scholar] [CrossRef]
- Rifai, L.; Saleh, F.A. A Review on Acrylamide in Food: Occurrence, Toxicity, and Mitigation Strategies. Int. J. Toxicol. 2020, 39, 93–102. [Google Scholar] [CrossRef]
- Yagua, C.V.; Moreira, R.G. Physical and thermal properties of potato chips during vacuum frying. J. Food Eng. 2011, 104, 272–283. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Essential Amino Acids | Limiting Amino Acids | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Variety | EAAI | Thr | Met + Cys | Val | Ile | Leu | Phe + Tyr | Lys | ∑eaa | 1st | 2nd |
| Blaue Annelise | 79 | 71 | 63 | 93 | 92 | 63 | 118 | 65 | 81 | Met + Cys | Lys + Leu |
| Blue Congo | 86 | 76 | 59 | 114 | 102 | 63 | 134 | 79 | 91 | Met + Cys | Leu |
| Herbie 26 | 96 | 82 | 78 | 125 | 112 | 72 | 134 | 88 | 79 | Leu | Met + Cys |
| Rote Emma | 110 | 93 | 78 | 146 | 129 | 88 | 165 | 101 | 116 | Met + Cys | Leu |
| Vineta | 82 | 82 | 63 | 103 | 89 | 63 | 115 | 75 | 85 | Met + Cys | Lys + Leu |
| Fresco | 118 | 115 | 92 | 154 | 122 | 84 | 182 | 102 | 123 | Leu | Met + Cys |
| Protein standard (g/16 g N) | 3.4 | 2.5 | 3.5 | 2.8 | 6.6 | 6.3 | 5.8 | 30.9 | - | - | |
| Variety/Plant Cultivar | FCR | HPLC | ||
|---|---|---|---|---|
| g GAE/kg FW | g GAE/kg DW | mg/100 g FW | mg/100 g DW | |
| Cauliflower | 0.57–2.55 | 5.60–29.13 | 30–217 | |
| Cabbage | 1.70–2.53 | 1.90–26.29 | 0.50 | 125–387 |
| Broccoli | 0.96–3.76 | 1.10–1.14 | 1173 | |
| Spinach | 0.50–2.34 | 11.8 | ||
| Carrot (orange & black) | 0.16–10.29 | 1.10–1.90 | 0.30–39.76 | |
| Brinjal | 0.82–2.92 | |||
| Green bean | 0.78–4.58 | 17.10–66.3 | ||
| Mushroom | 0.09–1.80 | |||
| Tomato | 0.14–2.91 | 26.57 | ||
| Strawberry | 0.99–3.05 | |||
| Blueberry | 2.20–7.53 | |||
| Kiwifruit | 0.80–0.96 | |||
| Pea | 0.80–1.20 | |||
| Brussels sprouts | 147 | |||
| Italian kale | 1127 | |||
| Potato | 0.31–8.83 | 4.48–11.19 | 23.2–67.4 | 260–2852 |
| Macro-Elements | Micro-Elements | |||||
|---|---|---|---|---|---|---|
| K | Ca | Mg | P | Fe | Zn | |
| Recommended intake/day | 3.1 g | 800 mg | 280 mg | 12–18 mg | 7 mg | |
| Banana | 261–546 | 0.35~1.35 | 80.0~84.0 | / | 17.15~47.19 | 0.34 |
| Cauliflower | 389–705 | 6.33 | 5.00 | / | 0.29 | 0.12 |
| Broccoli | 405–727 | 8.87–11.40 | 5.9 | / | 0.26 | 0.87 |
| Spinach | 210–630 | 98.0–241.39 | 11.1–85.0 | / | 3.90–9.49 | 0.2–0.55 |
| Pepper | 159–244 | 8.8–13.9 | 11.2–18.8 | 17.6–37.6 | 288–975 | 270–775 |
| Cabbage | 165–340 | 70–152 | 12.0–42.0 | 32 | 1.4–1.7 | 0.3–0.5 |
| Carrot | 357 | 25 | 71.0 | / | 9.28 | 1.28 |
| Strawberry | 153–312 | 7.6–29 | 11.0–23.4 | 0.5–24 | 0.20–0.62 | 0.14–1.5 |
| Cow milk | 152 | 122 | 12.0 | 119 | 0.08 | 0.530 |
| Kiwifruit | 254–300 | 15.4–26 | 11.0–13.1 | 24–25 | 0.19–0.25 | 0.09–0.14 |
| Potato | 104–540 | 4.98–21.29 | 13.7–41.9 | 30–60 | 0.34–1.80 | 0.27–0.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Li, Y.; Kaur, L.; Singh, J.; Zeng, F. Functional Food Based on Potato. Foods 2023, 12, 2145. https://doi.org/10.3390/foods12112145
Xu J, Li Y, Kaur L, Singh J, Zeng F. Functional Food Based on Potato. Foods. 2023; 12(11):2145. https://doi.org/10.3390/foods12112145
Chicago/Turabian StyleXu, Jian, Yang Li, Lovedeep Kaur, Jaspreet Singh, and Fankui Zeng. 2023. "Functional Food Based on Potato" Foods 12, no. 11: 2145. https://doi.org/10.3390/foods12112145