
Chemical Composition and Biological Activity of Commelina erecta: An Edible Wild Plant Consumed in Brazil

 ,
,  ,
,  ,
,  , , , , and
, , , , and
Abstract
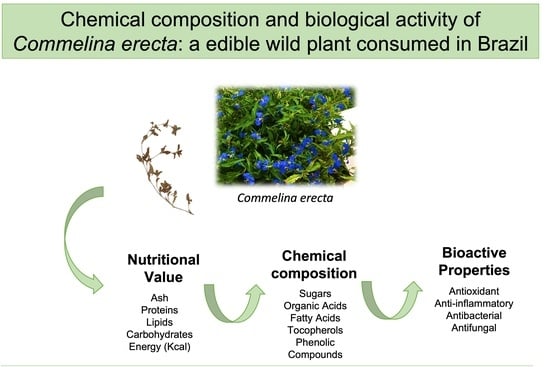
:
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Preparation of Extracts
2.3. Chemical Parameters
2.3.1. Nutritional Value
2.3.2. Free Sugars
2.3.3. Organic Acids
2.3.4. Fatty Acids
2.3.5. Tocopherols
2.3.6. Phenolic Compounds
2.4. Bioactive Properties
2.4.1. Evaluation of In Vitro Antioxidant Properties
2.4.2. Anti-Inflammatory Activity
2.4.3. Antimicrobial Activity
2.5. Statistical Analysis
3. Results and Discussion
3.1. Chemical and Nutritional Characterization of C. erecta Aerial Parts
3.2. Bioactive Properties of C. erecta Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Alam, M.K.; Rana, Z.H.; Islam, S.N.; Akhtaruzzaman, M. Comparative assessment of nutritional composition, polyphenol profile, antidiabetic and antioxidative properties of selected edible wild plant species of Bangladesh. Food Chem. 2020, 320, 126646. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.M.; Shah, S.Z.; Khan, M.S.; Akhtar, N. Evaluation of indigenous knowledge of medicinal plants from Tall Dardyal Hills, Khyber Pakhtunkhwa, Pakistan. J. Herb. Med. 2020, 20, 100314. [Google Scholar] [CrossRef]
- Shabab, S.; Gholamnezhad, Z.; Mahmoudabady, M. Protective effects of medicinal plant against diabetes induced cardiac disorder: A review. J. Ethnopharmacol. 2021, 265, 113328. [Google Scholar] [CrossRef] [PubMed]
- Iyda, J.H.; Fernandes, Â.; Calhelha, R.C.; Alves, M.J.; Ferreira, F.D.; Barros, L.; Amaral, J.S.; Ferreira, I.C.F.R. Nutritional composition and bioactivity of Umbilicus rupestris (Salisb.) Dandy: An underexploited edible wild plant. Food Chem. 2019, 295, 341–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geraci, A.; Amato, F.; Di Noto, G.; Bazan, G.; Schicchi, R. The wild taxa utilized as vegetables in Sicily (Italy): A traditional component of the Mediterranean diet. J. Ethnobiol. Ethnomed. 2018, 14, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bredariol, L.R. Levantamento e caracterização das Plantas Alimentícias Não Convencionais (PANC’S) Espontâneas Presentes em um Sistema Agroflorestal No Município de Rio Claro-SP, Trabalho de Conclusão de curso (Ecologia). Universidade Estadual Paulista “Julio de Mesquita Filho”—Instituto de Biociências, Rio Claro, Brazil. 2015. Available online: https://repositorio.unesp.br/bitstream/handle/11449/139038/000865068.pdf?sequence=1&isAllowed=y (accessed on 1 November 2022).
- Huergo, E.M.; Galeano, Y.P.G.; Lima, L.C.P. Plantas Alimentícias Não Convencionais (PANC) do município de Foz do Iguaçu, Paraná, Brasil. Heringeriana 2020, 14, 107–132. [Google Scholar] [CrossRef]
- Misra, A.; Srivastava, A.; Srivastava, S.; Rawat, A.K.S. Simultaneous reverse-phase HPLC determination of major antioxidant phenolics in Commelina benghalensis L. tubers. Acta Chromatogr. 2016, 28, 541–554. [Google Scholar] [CrossRef] [Green Version]
- Seixas, S.; Mangueira, S.; Gonçalves, C.M.; Pereira, D.P.; Mendes, J.E. Eixo temático: Saúde e Agroecologia. In Proceedings of the Anais do XI Congresso Brasileiro de Agroecologia, São Cristóvão, Brazil, 2020; Non-Conventional Food Plants with Food Potential in the CENTEC Abaitará Campus. p. 6. [Google Scholar]
- Sfoglia, N.; Biondo, E.; Zanetti, C.; Cherobini, L.; Kolchinski, E.M.; Sant’Anna, V. Caracterização da agrobiodiversidade no Vale do Taquari, RS: Levantamento florístico, consumo e agroindustrialização de hortaliças não convencionais. Cad. Ciência Tecnol. 2019, 36, 26489. [Google Scholar] [CrossRef]
- De Albuquerque, U.P.; de Medeiros, P.M.; de Almeida, A.L.S.; Monteiro, J.M.; de Freitas Lins Neto, E.M.; de Melo, J.G.; dos Santos, J.P. Medicinal plants of the caatinga (semi-arid) vegetation of NE Brazil: A quantitative approach. J. Ethnopharmacol. 2007, 114, 325–354. [Google Scholar] [CrossRef]
- Brandão, M.G.L.; Pignal, M.; Romaniuc, S.; Grael, C.F.F.; Fagg, C.W. Useful Brazilian plants listed in the field books of the French naturalist Auguste de Saint-Hilaire (1779–1853). J. Ethnopharmacol. 2012, 143, 488–500. [Google Scholar] [CrossRef]
- Malami, I.; Jagaba, N.M.; Abubakar, I.B.; Muhammad, A.; Alhassan, A.M.; Waziri, P.M.; Yahaya, I.Z.Y.; Mshelia, H.E.; Mathias, S.N. Integration of medicinal plants into the traditional system of medicine for the treatment of cancer in Sokoto State, Nigeria. Heliyon 2020, 6, e04830. [Google Scholar] [CrossRef]
- Ricardo, L.M.; De Paula-Souza, J.; Andrade, A.; Brandão, M.G.L. Plants from the Brazilian traditional medicine: Species from the books of the polish physician piotr czerniewicz (Pedro Luiz Napoleão Chernoviz, 1812–1881). Rev. Bras. Farmacogn. 2017, 27, 388–400. [Google Scholar] [CrossRef]
- Siew, Y.Y.; Yew, H.C.; Neo, S.Y.; Seow, S.V.; Lew, S.M.; Lim, S.W.; Lim, C.S.E.S.; Ng, Y.C.; Seetoh, W.G.; Ali, A.; et al. Evaluation of anti-proliferative activity of medicinal plants used in Asian Traditional Medicine to treat cancer. J. Ethnopharmacol. 2019, 235, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Batool, R.; Aziz, E.; Mahmood, T.; Tan, B.; Chow, V. Inhibitory activities of extracts of Rumex dentatus, Commelina benghalensis, Ajuga bracteosa, Ziziphus mauritiana as well as their compounds of gallic acid and emodin against dengue virus. Asian Pac. J. Trop. Med. 2018, 11, 265–271. [Google Scholar]
- Lebogo, K.W.; Mokgotho, M.P.; Bagla, V.P.; Matsebatlela, T.M.; Mbazima, V.; Shai, L.J.; Mampuru, L. Semi-purified extracts of Commelina benghalensis (Commelinaceae) induce apoptosis and cell cycle arrest in Jurkat-T cells. BMC Complement. Altern. Med. 2014, 14, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokilavani, P.; Suriyakalaa, U.; Elumalai, P.; Abirami, B.; Ramachandran, R.; Sankarganesh, A.; Achiraman, S. Antioxidant mediated ameliorative steroidogenesis by Commelina benghalensis L. and Cissus quadrangularis L. against quinalphos induced male reproductive toxicity. Pestic. Biochem. Physiol. 2014, 109, 18–33. [Google Scholar] [CrossRef] [PubMed]
- Mbazima, V.G.; Mokgotho, M.P.; February, F.; Rees, D.J.G.; Mampuru, L.J. Alteration of Bax-to-Bcl-2 ratio modulates the anticancer activity of methanolic extract of Commelina benghalensis (Commelinaceae) in Jurkat T cells. African J. Biotechnol. 2008, 7, 3569–3576. [Google Scholar]
- Frankova, A.; Vistejnova, L.; Merinas-Amo, T.; Leheckova, Z.; Doskocil, I.; Soon, J.W.; Kudera, T.; Laupua, F.; Alonso-Moraga, A.; Kokoska, L. In vitro antibacterial activity of extracts from Samoan medicinal plants and their effect on proliferation and migration of human fibroblasts. J. Ethnopharmacol. 2021, 264, 113220. [Google Scholar] [CrossRef]
- Houghton, P.J.; Hylands, P.J.; Mensah, A.Y.; Hensel, A.; Deters, A.M. In vitro tests and ethnopharmacological investigations: Wound healing as an example. J. Ethnopharmacol. 2005, 100, 100–107. [Google Scholar] [CrossRef]
- Kuppusamy, P.; Ilavenil, S.; Srigopalram, S.; Kim, D.H.; Govindan, N.; Maniam, G.P.; Yusoff, M.M.; Choi, K.C. Synthesis of Bimetallic Nanoparticles (Au–Ag Alloy) Using Commelina nudiflora L. Plant Extract and Study its on Oral Pathogenic Bacteria. J. Inorg. Organomet. Polym. Mater. 2017, 27, 562–568. [Google Scholar] [CrossRef]
- Shah, M.D.; D’Souza, U.J.A.; Iqbal, M. The potential protective effect of Commelina nudiflora L. against carbon tetrachloride (CCl4)-induced hepatotoxicity in rats, mediated by suppression of oxidative stress and inflammation. Environ. Health Prev. Med. 2017, 22, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Review, P.A.; Stress, O.; Alagar, B.; Maran, V.; Iqbal, M.; Gangadaran, P.; Ahn, B. Hepatoprotective Potential of Malaysian Medicinal. Molecules 2022, 27, 1533. [Google Scholar]
- Otsuka, F.A.M.; Santos, R.B.; Chaves, L.F.; Santos, R.S.; Chaves Filho, A.B.; Miyamoto, S.; Matos, H.R. Identification of caffeic acid and rutin by UHPLC MS/MS and antioxidant activity of Commelina erecta lineu. In cell culture. An. Acad. Bras. Cienc. 2020, 92, e20190491. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, J.A.; Corrêa, R.F.; Salomé, K.; Santos, A.D.C.; Lima, E.S.; Campos, F.R.; Barison, A.; Pinheiro, M.L.B. Phytochemical Study and Antioxidant Evaluation of Commelina erecta (Commelinaceae) Stems. Rev. Virtual Quim. 2019, 11, 255–263. [Google Scholar] [CrossRef]
- Olano, I.; Paz, E.A.; Cerdeíras, M.P.; Fernández, J.; Ferreira, F.; Moyna, P.; Soubes, M.; Vázquez, A.; Vero, S.; Bassagoda, M.J. Screening of Uruguayan medicinal plants for antimicrobial activity. Part II. J. Ethnopharmacol. 1996, 53, 111–115. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of Association of Official Analytical Chemists International, 21st ed.; Horwitz, W., Latimer, G., Eds.; AOAC International: Gaithersburg, MD, USA, 2016; ISBN 0935584773. [Google Scholar]
- Spréa, R.M.; Fernandes, Â.; Calhelha, R.C.; Pereira, C.; Pires, T.C.S.P.; Alves, M.J.; Canan, C.; Barros, L.; Amaral, J.S.; Ferreira, I.C.F.R. Chemical and bioactive characterization of the aromatic plant: Levisticum officinale W.D.J. Koch: A comprehensive study. Food Funct. 2020, 11, 1292–1303. [Google Scholar] [CrossRef]
- Bessada, S.M.F.; Barreira, J.C.M.; Barros, L.; Ferreira, I.C.F.R.; Oliveira, M.B.P.P. Phenolic profile and antioxidant activity of Coleostephus myconis (L.) Rchb.f.: An underexploited and highly disseminated species. Ind. Crops Prod. 2016, 89, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Silva, I.; Sá, D.; Paula, A.; Vitória, F.; Nicola, G.; Nunes, B.; Fachinetto, R.; Pinela, J.; Costa, R.; Filomena, M.; et al. In vitro and in vivo evaluation of enzymatic and antioxidant activity, cytotoxicity and genotoxicity of curcumin-loaded solid dispersions. Food Chem. Toxicol. 2019, 125, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Essoh, A.P.; Liberal, Â.; Fernandes, Â.; Dias, M.I.; Pereira, C.; Mandim, F.; Moldão-Martins, M.; Cravo, P.; Duarte, M.P.; Moura, M.; et al. Evaluation of the Polyphenolic Composition and Bioactivities of Three Native Cabo Verde Medicinal Plants. Pharmaceuticals 2022, 15, 1162. [Google Scholar] [CrossRef]
- Soković, M.; Glamočlija, J.; Marin, P.D.; Brkić, D.; Griensven, L.J.L.D. van Antibacterial Effects of the Essential Oils of Commonly Consumed Medicinal Herbs Using an in vitro Model. Molecules 2010, 15, 7532–7546. [Google Scholar] [CrossRef] [Green Version]
- Fotakis, C.; Tsigrimani, D.; Tsiaka, T.; Lantzouraki, D.Z.; Strati, I.F.; Makris, C.; Tagkouli, D.; Proestos, C.; Sinanoglou, V.J.; Zoumpoulakis, P. Metabolic and antioxidant profiles of herbal infusions and decoctions. Food Chem. 2016, 211, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, J.P.L.; Yuyama, L.K.O.; Souza, F. das C. do A.; Pessoa, A. Biodisponibilidade do ferro do jambu (Spilanthes oleracea L.): Estudo em murinos. Rev. Pan-Amazônica Saúde 2014, 5, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Duarte, J.M.; Areco, K.C.N. Tabela de Composição Química Dos Alimentos. Available online: http://tabnut.dis.epm.br/Credito (accessed on 14 November 2022).
- Queiroz, C.R.A.d.A.; Melo, C.M.T.; Andrade, R.R.; Pavani, L.C.; Morais, S.A.L. 34a Reunião Anual da Sociedade Brasileira de Química; Instituto Federal do Triângulo Mineiro (IFTM) Campus Uberlândia: Uberlandia, Brazil, 2009. [Google Scholar]
- Monerri, C.; Fortunato-Almeida, A.; Molina, R.V.; Nebauer, S.G.; García-Luis, A.; Guardiola, J.L. Relation of carbohydrate reserves with the forthcoming crop, flower formation and photosynthetic rate, in the alternate bearing “Salustiana” sweet orange (Citrus sinensis L.). Sci. Hortic. 2011, 129, 71–78. [Google Scholar] [CrossRef]
- Priecina, L.; Karklina, D. Composition of Major Organic Acids in Vegetables and Spices. CBU Int. Conf. Proc. 2015, 3, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Abdelhamid, A.S.; Brown, T.J.; Brainard, J.S.; Biswas, P.; Thorpe, G.C.; Moore, H.J.; Deane, K.H.O.; Alabdulghafoor, F.K.; Summerbell, C.D.; Worthington, H.V.; et al. Omega-3 fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2018, 2018, 1–30. [Google Scholar]
- Ferreres, F.; Gomes, N.G.M.; Valentão, P.; Pereira, D.M.; Gil-Izquierdo, A.; Araújo, L.; Silva, T.C.; Andrade, P.B. Leaves and stem bark from Allophylus africanus P. Beauv.: An approach to anti-inflammatory properties and characterization of their flavonoid profile. Food Chem. Toxicol. 2018, 118, 430–438. [Google Scholar] [CrossRef]
- Cao, J.; Yin, C.; Qin, Y.; Cheng, Z.; Chen, D. Approach to the study of flavone di-C-glycosides by high performance liquid chromatography-tandem ion trap mass spectrometry and its application to characterization of flavonoid composition in Viola yedoensis. J. Mass Spectrom. 2014, 49, 1010–1024. [Google Scholar] [CrossRef]
- Figueirinha, A.; Paranhos, A.; Pérez-Alonso, J.J.; Santos-Buelga, C.; Batista, M.T. Cymbopogon citratus leaves: Characterization of flavonoids by HPLC-PDA-ESI/MS/MS and an approach to their potential as a source of bioactive polyphenols. Food Chem. 2008, 110, 718–728. [Google Scholar] [CrossRef]
- Niu, Y.; Li, H.; Dong, J.; Wang, H.; Hashi, Y.; Chen, S. Identification of isoflavonoids in Radix Puerariae for quality control using on-line high performance liquid chromatography-diode array detector-electrospray ionization-mass spectrometry coupled with post-column derivatization. Food Res. Int. 2012, 48, 528–537. [Google Scholar] [CrossRef]
- Prasain, J.K.; Reppert, A.; Jones, K.; Moore, D.R.; Barnes, S.; Lila, M.A. Identification of isoflavone glycosides in pueraria lobata cultures by tandem mass spectrometry. Phytochem. Anal. 2007, 18, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.A.-D.P.; Swain, T. Flavonoids and Chemotaxonomy of the Commelinaceae. Biochem. Syst. Ecol. 1985, 13, 391–402. [Google Scholar] [CrossRef]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The therapeutic potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurcheshen, M.; Seehuus, M.; Pigeon, W. Updates on Nutraceutical Sleep Therapeutics and Investigational Research. Evid.-Based Complement. Altern. Med. 2015, 2015, 9. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Flowering Stem | Stem after Flowering | Flowering Leaf | Leaf after Flowering | |
|---|---|---|---|---|
| Nutritional value (g/100 g fw 1) | ||||
| Moisture | 94 ± 6 a | 90.7 ± 9 b | 94 ± 6 a | 89 ± 11 c |
| Lipids | 0.08 ± 0.01 d | 1.86 ± 0.06 b | 0.14 ± 0.01 c | 2.03 ± 0.04 a |
| Protein | 0.53 ± 0.03 d | 0.97 ± 0.01 c | 1.64 ± 0.07 b | 1.99 ± 0.05 a |
| Ash | 0.73 ± 0.01 d | 1.02 ± 0.02 c | 0.92 ± 0.02 b | 1.39 ± 0.02 a |
| Carbohydrates | 4.55 ± 0.01 b | 5.5 ± 0.3 a | 3.50 ± 0.07 c | 5.9 ± 0.9 a |
| Energy (kcal/100 g fw) | 21.09 ± 0.03 c | 42.6 ± 0.7 b | 21.82 ± 0.04 d | 50 ± 3 a |
| Free sugars (g/100 g fw) | ||||
| Fructose | 0.079 ± 0.001 d | 0.540 ± 0.002 a | 0.212 ± 0.003 c | 0.311 ± 0.004 b |
| Sucrose | tr 2 | tr | 0.014 ± 0.002 | tr |
| Total free sugars | 0.079 ± 0.001 c | 0.540 ± 0.002 a | 0.230 ± 0.004 d | 0.311 ± 0.004 b |
| Organic acids (mg/100 g fw) | ||||
| Oxalic | 256.8 ± 0.1 c | 507 ± 1 a | 208 ± 1 d | 383 ± 1 b |
| Malic | 160 ± 2 a | 84 ± 1 b | 68 ± 1 c | 39 ± 1 d |
| Shikimic | 41.4 ± 0.1 c | 98.01 ± 0.01 b | 9.6 ± 0.1 d | 98.1 ± 0.2 a |
| Ascorbic | 2.18 ± 0.09 c | 9.5 ± 0.8 a | nd | 7.1 ± 0.1 b |
| Total organic acids | 460 ± 2 c | 699 ± 1 a | 285 ± 2 d | 527 ± 2 b |
| Flowering Stem | Stem after Flowering | Flowering Leaf | Leaf after Flowering | |
|---|---|---|---|---|
| Fatty acids (relative %) | ||||
| C6:0 | 0.36 ± 0.01 | nd | nd | nd |
| C8:0 | 0.11 ± 0.01 | nd | nd | nd |
| C11:0 | 0.12 ± 0.01 | nd | nd | nd |
| C12:0 | nd | nd | nd | nd |
| C13:0 | 0.48 ± 0.01 | nd | 0.76± 0.04 | 1.08 ± 0.01 |
| C14:0 | 0.45 ± 0.01 | nd | 0.33 ± 0.01 | nd |
| C15:0 | 0.31 ± 0.02 | nd | nd | nd |
| C16:0 | 23.31 ± 0.01 | 30.51 ± 0.05 | 21.9 ± 0.2 | 16.60 ± 0.07 |
| C16:1 | 2.08 ± 0.04 | 0.94 ± 0.01 | 2.8 ± 0.1 | nd |
| C17:0 | 0.99 ± 0.01 | 0.65 ± 0.01 | nd | nd |
| C18:0 | 6.41 ± 0.08 | 4.60 ± 0.05 | 4.28 ± 0.02 | 4.91 ± 0.05 |
| C18:1n9c | 15.1 ± 0.1 | 8.11 ± 0.07 | 4.22 ± 0.04 | 10.9 ± 0.1 |
| C18:2n6c | 26.95 ± 0.04 | 28.5 ± 0.1 | 19.24 ± 0.02 | 20.03 ± 0.02 |
| C18:3n3 | 17.9 ± 0.2 | 18.17 ± 0.05 | 43.6 ± 0.3 | 42.74 ± 0.08 |
| C20:0 | 0.95 ± 0.01 | nd | 1.72 ± 0.08 | 1.57 ± 0.08 |
| C22:0 | 1.68 ± 0.04 | 2.70 ± 0.09 | nd | nd |
| C23:0 | 1.11 ± 0.03 | 2.48 ± 0.01 | nd | nd |
| C24:0 | 1.82 ± 0.02 | 3.50 ± 0.06 | 1.25 ± 0.01 | 2.27 ± 0.01 |
| SFA (%) | 38.11 ± 0.01 b | 44.4 ± 0.2 a | 30.2 ± 0.2 c | 26.37 ± 0.08 d |
| MUFA (%) | 17.1 ± 0.2 a | 9.06 ± 0.06 c | 7.0 ± 0.2 d | 10.9 ± 0.1 b |
| PUFA (%) | 44.8 ± 0.1 c | 46.5 ± 0.2 b | 62.8 ± 0.3 a | 62.77 ± 0.06 a |
| Tocopherols (mg/100 g fw) | ||||
| α-Tocopherol | 2.14 ± 0.05 d | 6.3 ± 0.1 c | 17.0 ± 0.2 a | 15.9 ± 0.4 b |
| β-Tocopherol | 6.8 ± 0.3 c | 5.8 ± 0.3 d | 13.8 ± 0.4 a | 12.7 ± 0.9 b |
| γ-Tocopherol | 21.2 ± 0.1 a | 9.5 ± 0.4 c | 12.4 ± 0.4 b | 8.4 ± 0.4 d |
| δ-Tocopherol | nd | nd | 10.2 ± 0.4 | nd |
| Total tocopherols | 30.1 ± 0.5 c | 21.7 ± 0.8 d | 53.4 ± 0.1 a | 37.0 ± 0.1 b |
| Peak | Rt (min) | λmax (nm) | [M-H]− (m/z) | MS2 (m/z) | Tentative Identification |
|---|---|---|---|---|---|
| 1 | 7.19 | 324 | 593 | 503 (31), 473 (10), 431 (100), 311 (14) | Apigenin-7-O-hexosyl-8-C-hexoside |
| 2 | 8.74 | 272/327 | 593 | 533 (10), 503 (61), 473 (100), 413 (11), 383 (25), 353 (32) | Apigenin-6,8-di-C-hexoside |
| 3 | 9.24 | 271/345 | 579 | 561 (11), 519 (19), 489 (23), 459 (100), 399 (31), 369 (15) | Luteolin-6-C-hexosyl-8-C-pentoside derivative I |
| 4 | 10.19 | 270/343 | 579 | 561 (12), 519 (13), 489 (45), 459 (100), 399 (39), 369 (11) | Luteolin-6-C-hexosyl-8-C-pentoside derivative II |
| 5 | 12.15 | 272/331 | 563 | 503 (12), 443 (100), 383 (23), 353 (32) | Apigenin 6-C-hexosyl-8-C-pentoside |
| 6 | 13.19 | 272/327 | 563 | 503 (71), 443 (100), 383 (34), 353 (63) | Apigenin 6-C-pentosyl-8-C-hexoside |
| 7 | 14.18 | 271/343 | 549 | 531 (18), 489 (32), 459 (100), 441 (23), 429 (13), 399 (23), 369 (22) | Luteolin-6-C-pentosyl-8-C-pentoside derivative I |
| 8 | 15.89 | 271/342 | 549 | 531 (23), 489 (34), 459 (100), 441 (18), 429 (5), 399 (35), 369 (36) | Luteolin-6-C-pentosyl-8-C-pentoside derivative II |
| 9 | 16.16 | 272/328 | 533 | 515 (52), 473 (71), 443 (100), 383 (28), 353 (25) | Apigenin-6,8-di-C-pentoside |
| 10 | 17.09 | 335 | 431 | 311 (30), 283 (100) | Hydroxy-Puerarin |
| 11 | 17.96 | 327 | 463 | 301 (100) | Quercetin-3-O-hexoside |
| 12 | 19.21 | 320 | 549 | 505 (10), 463 (32), 301 (100) | Quercetin-O-malonyl-hexoside |
| 13 | 21.19 | 325 | 447 | 301 (100) | Quercetin-O-deoxyhexoside |
| Peak | Flowering Stem | Stem after Flowering | Flowering Leaf | Leaf after Flowering | ||||
|---|---|---|---|---|---|---|---|---|
| Hydroethanolic | Aqueous | Hydroethanolic | Aqueous | Hydroethanolic | Aqueous | Hydroethanolic | Aqueous | |
| 1 | nd | nd | nd | nd | nd | 0.334 ± 0.07 | nd | nd |
| 2 | 0.396 ± 0.002 a | nd | 0.09 ± 0.002 f | 0.22 ± 0.004 d | 0.284 ± 0.003 c | 0.33 ± 0.03 b | 0.32 ± 0.03 b | 0.171 ± 0.004 e |
| 3 | tr | nd | tr | nd | tr | nd | nd | nd |
| 4 | 0.384 ± 0.009 | tr | nd | tr | tr | nd | nd | tr |
| 5 | 2.96 ± 0.06 a | 0.39 ± 0.02 b | nd | 0.235 ± 0.004 d | 0.258 ± 0.001 c | 0.39 ± 0.02 b | 0.184 ± 0.004 e | 0.088 ± 0.001 f |
| 6 | 1.485 ± 0.005 a | 0.124 ± 0.005 f | nd | 0.258 ± 0.01 c | 0.21 ± 0.003 d | 0.29 ± 0.03 b | nd | nd |
| 7 | 3.27 ± 0.01 | tr | nd | tr | nd | tr | nd | tr |
| 8 | 0.47 ± 0.03 | tr | nd | tr | nd | nd | tr | tr |
| 9 | 0.891 ± 0.009 a | 0.17 ± 0.003 c | nd | nd | 0.349 ± 0.001 b | nd | nd | nd |
| 10 | nd | nd | nd | nd | nd | 0.77 ± 0.04 | nd | nd |
| 11 | 0.237 ± 0.015 | nd | nd | nd | nd | nd | nd | nd |
| 12 | 0.064 ± 0.001 | nd | nd | nd | nd | nd | nd | nd |
| 13 | 0.104 ± 0.009 | nd | nd | tr | tr | nd | tr | tr |
| TFC | 10.26 ± 0.01 a | 0.679 ± 0.02 e | 0.09 ± 0.002 h | 0.71 ± 0.02 d | 1.103 ± 0.003 c | 2.1 ± 0.1 b | 0.51 ± 0.02 f | 0.259 ± 0.005 g |
| TCGF | 9.849 ± 0.005 a | 0.679 ± 0.02 e | 0.09 ± 0.002 h | 0.71 ± 0.02 d | 1.102 ± 0.003 c | 2.1 ± 0.1 b | 0.51 ± 0.02 f | 0.259 ± 0.005 g |
| TOGF | 0.406 ± 0.005 | nd | nd | tr | tr | nd | tr | tr |
| Flowering Stem | Stem after Flowering | Flowering Leaf | Leaf after Flowering | |||
|---|---|---|---|---|---|---|
| TBARS inhibition (EC50; µg/mL) a | Aqueous | 74.8 ± 0.7 a | 18.4 ± 0.2 c | 63.4 ± 0.9 b | 63 ± 1 b | |
| Hydroethanolic | 75.1 ± 0.7 c | 638 ± 8 b | 1060 ± 78 a | 63 ± 2 d | ||
| OxHLIA (EC50; µg/mL) | ∆t = 60 min | Aqueous | 97 ± 6 b | 115 ± 7 a | 53 ± 3 d | 56 ± 3 c |
| Hydroethanolic | 80 ± 3 c | 103 ± 2 a | 89 ± 4 b | 58 ± 3 d | ||
| ∆t = 120 min | Aqueous | 392 ± 11 a | 353 ± 6 b | 211 ± 9 c | 139 ± 6 d | |
| Hydroethanolic | 170 ± 9 c | 189 ± 2 b | 248 ± 5 a | 113 ± 5 d | ||
| Anti-inflammatory activity (EC50 μg/mL) | Aqueous | >400 | >400 | >400 | >400 | |
| Hydroethanolic | 157 ± 7 | >400 | >400 | >400 | ||
| Antibacterial Activity (mg/mL) | |||||||
|---|---|---|---|---|---|---|---|
| Gram-Positive Bacteria | Gram-Negative Bacteria | ||||||
| S. aureus | B. cereus | L. monocytogenes | E. coli | S. Typhimurium | E. cloacae | ||
| MIC/MBC | MIC/MBC | MIC/MBC | MIC/MBC | MIC/MBC | MIC/MBC | ||
| Flowering stem | Aqueous | 0.5/1 | 1/2 | 1/2 | 1/2 | 2/4 | 2/4 |
| Hydroethanolic | 1/2 | 0.5/1 | 1/2 | 0.5/1 | 1/2 | 1/2 | |
| Stem after flowering | Aqueous | 1/2 | 1/2 | 1/2 | 1/2 | 1/2 | 1/2 |
| Hydroethanolic | 1/2 | 0.5/1 | 1/2 | 0.5/1 | 1/2 | 1/2 | |
| Flowering leaf | Aqueous | 1/2 | 1/2 | 1/2 | 1/2 | 1/2 | 1/2 |
| Hydroethanolic | 0.5/1 | 0.25/0.5 | 0.5/1 | 0.5/1 | 1/2 | 0.5/1 | |
| Leaf after flowering | Aqueous | 1/2 | 0.5/1 | 1/2 | 1/2 | 1/2 | 1/2 |
| Hydroethanolic | 1/2 | 0.5/1 | 1/2 | 1/2 | 1/2 | 1/2 | |
| E211 | 4/4 | 0.5/0.5 | 1/2 | 1/2 | 1/2 | 2/4 | |
| E224 | 1/1 | 2/4 | 0.5/1 | 0.5/1 | 1/1 | 0.5/0.5 | |
| Antifungal activity (mg/mL) | |||||||
| A. fumigatus | A. niger | A. ochraceus | P. funiculosum | P. verrucosum var. cyclopium | T. viride | ||
| MIC/MFC | MIC/MFC | MIC/MFC | MIC/MFC | MIC/MFC | MIC/MFC | ||
| Flowering stem | Aqueous | 0.5/1 | 0.5/1 | 0.5/1 | 0.5/1 | 1/2 | 1/2 |
| Hydroethanolic | 0.5/1 | 1/2 | 0.5/1 | 0.5/1 | 0.5/1 | 1/2 | |
| Stem after flowering | Aqueous | 0.25/0.5 | 0.5/1 | 0.5/1 | 0.5/1 | 1/2 | 1/2 |
| Hydroethanolic | 0.5/1 | 1/2 | 0.5/1 | 0.5/1 | 0.5/1 | 0.5/1 | |
| Flowering leaf | Aqueous | 0.5/1 | 0.5/1 | 0.5/1 | 0.5/1 | 0.5/1 | 0.5/1 |
| Hydroethanolic | 0.5/1 | 0.5/1 | 0.5/1 | 0.5/1 | 0.5/1 | 0.5/1 | |
| Leaf after flowering | Aqueous | 0.5/1 | 0.5/1 | 0.5/1 | 0.5/1 | 1/2 | 1/2 |
| Hydroethanolic | 1/1 | 1/2 | 0.5/1 | 0.5/1 | 0.5/1 | 0.5/1 | |
| E211 | 1/2 | 1/2 | 1/2 | 1/2 | 2/4 | 1.0/2.0 | |
| E224 | 1/1 | 1/1 | 1/1 | 0.5/0.5 | 1/1 | 0.5/0.5 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavichi, L.V.; Liberal, Â.; Dias, M.I.; Mandim, F.; Pinela, J.; Kostić, M.; Soković, M.; Kalschne, D.L.; Fernandes, Â.; Canan, C.; et al. Chemical Composition and Biological Activity of Commelina erecta: An Edible Wild Plant Consumed in Brazil. Foods 2023, 12, 192. https://doi.org/10.3390/foods12010192
Cavichi LV, Liberal Â, Dias MI, Mandim F, Pinela J, Kostić M, Soković M, Kalschne DL, Fernandes Â, Canan C, et al. Chemical Composition and Biological Activity of Commelina erecta: An Edible Wild Plant Consumed in Brazil. Foods. 2023; 12(1):192. https://doi.org/10.3390/foods12010192
Chicago/Turabian StyleCavichi, Lucas Vinicius, Ângela Liberal, Maria Inês Dias, Filipa Mandim, José Pinela, Marina Kostić, Marina Soković, Daneysa Lahis Kalschne, Ângela Fernandes, Cristiane Canan, and et al. 2023. "Chemical Composition and Biological Activity of Commelina erecta: An Edible Wild Plant Consumed in Brazil" Foods 12, no. 1: 192. https://doi.org/10.3390/foods12010192