
Composition of Phenolic Compounds in South African Schinus molle L. Berries



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Materials
2.3. Sample Methanolic Extraction
2.4. UPLC-QTOF-MS Profiling and Quantification
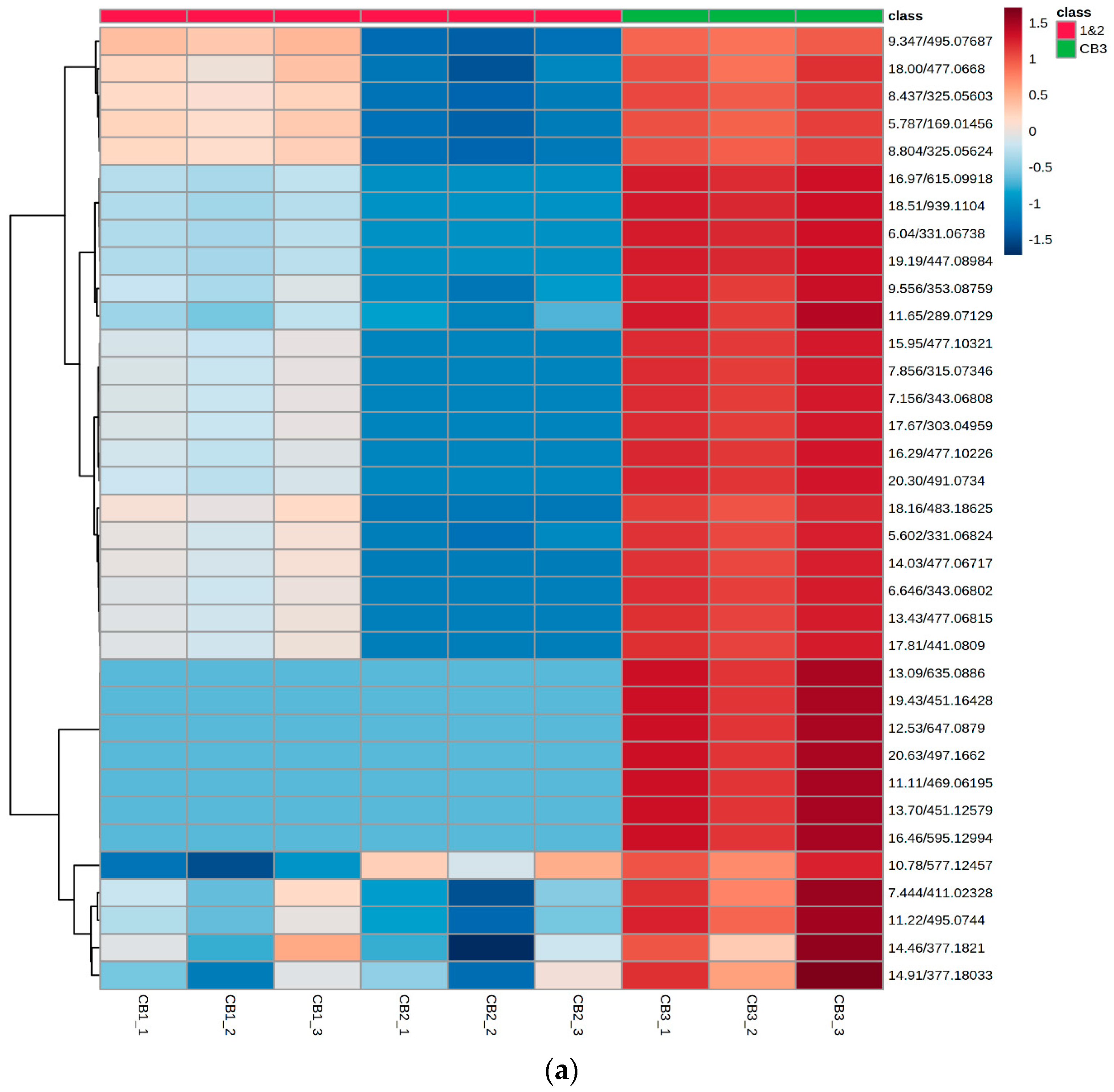
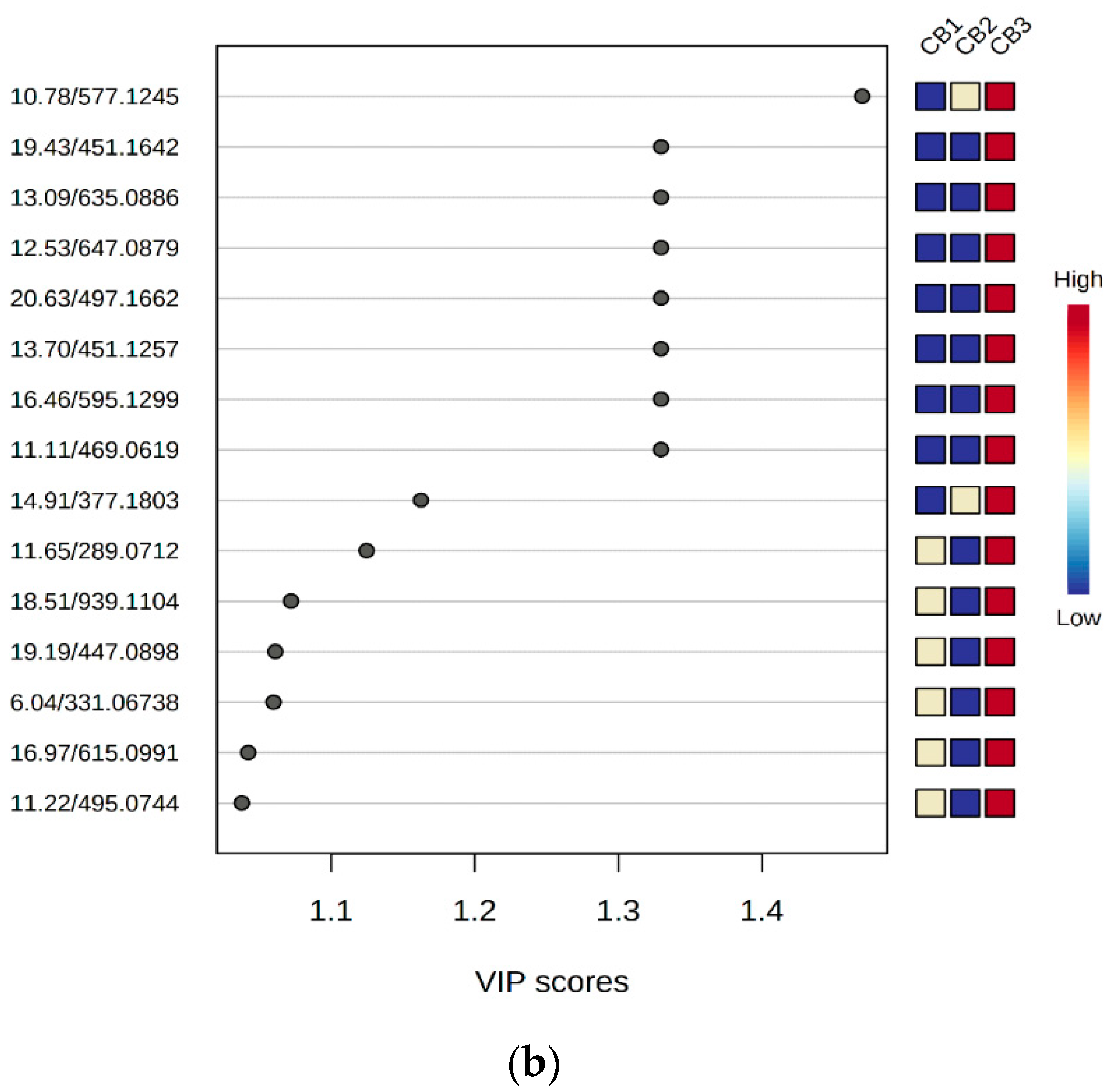
2.5. Chemometric Data Analysis
3. Results
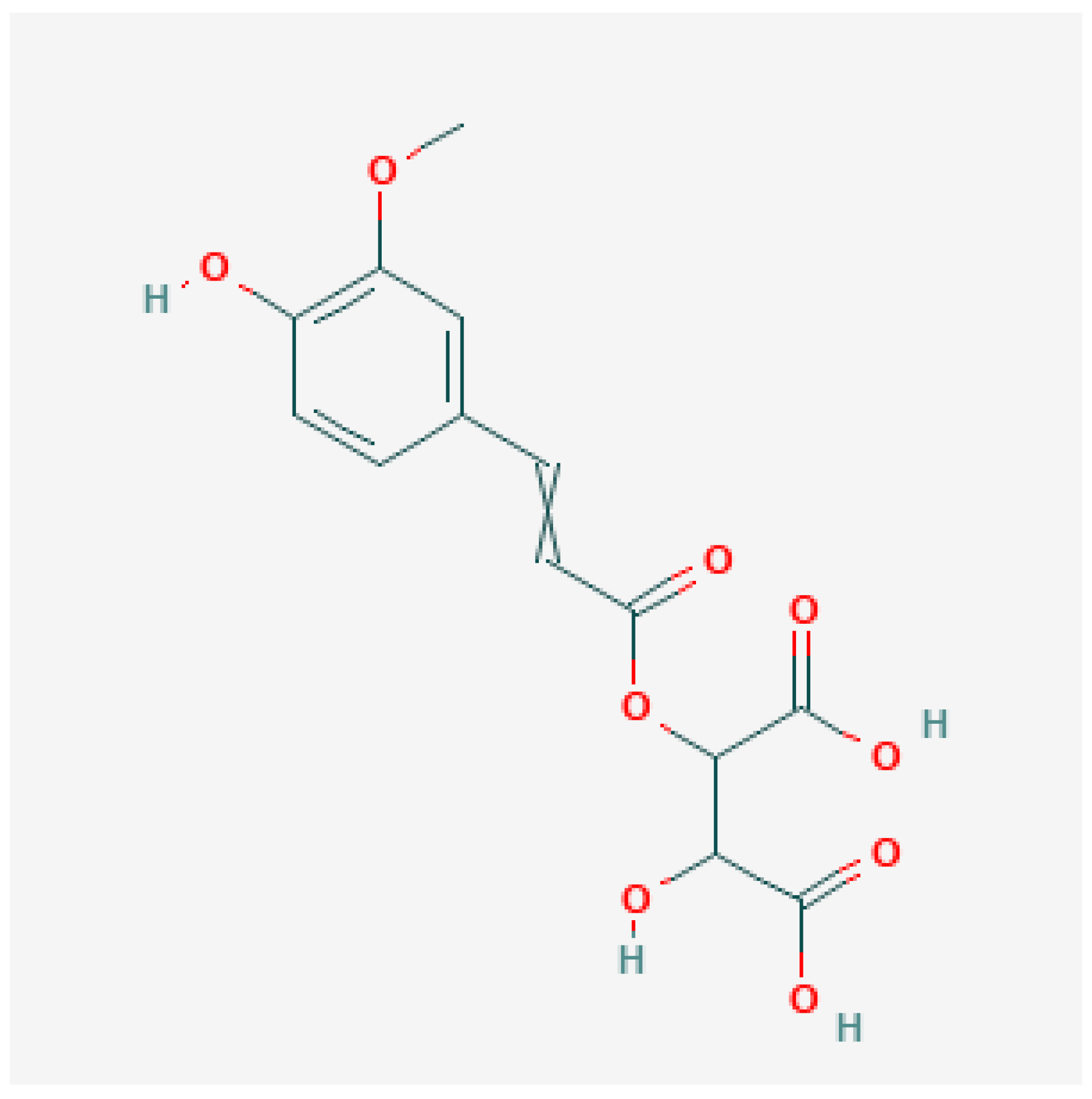
3.1. Phenolic Acids and/or Derivatives
3.2. Flavonoids
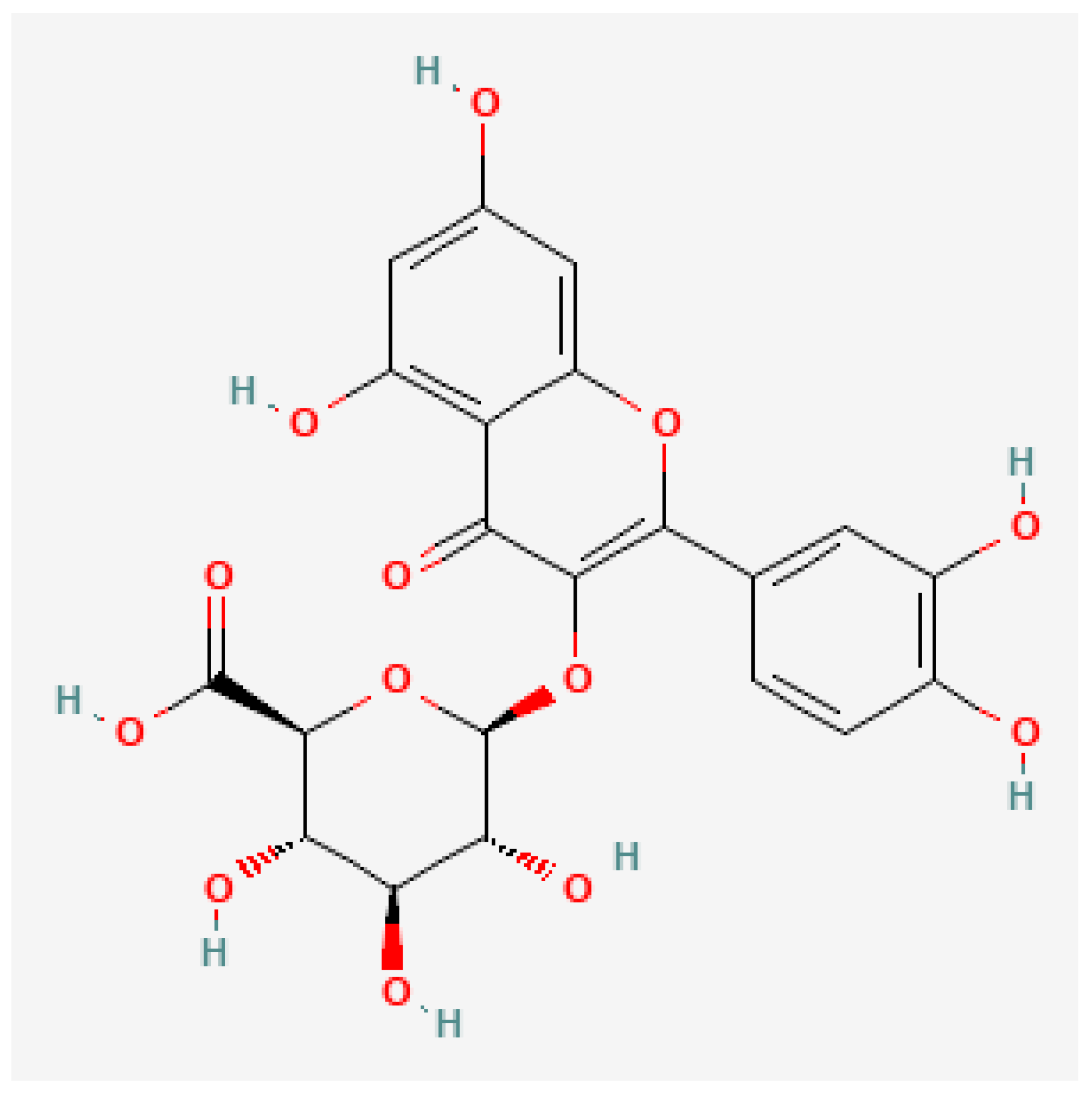
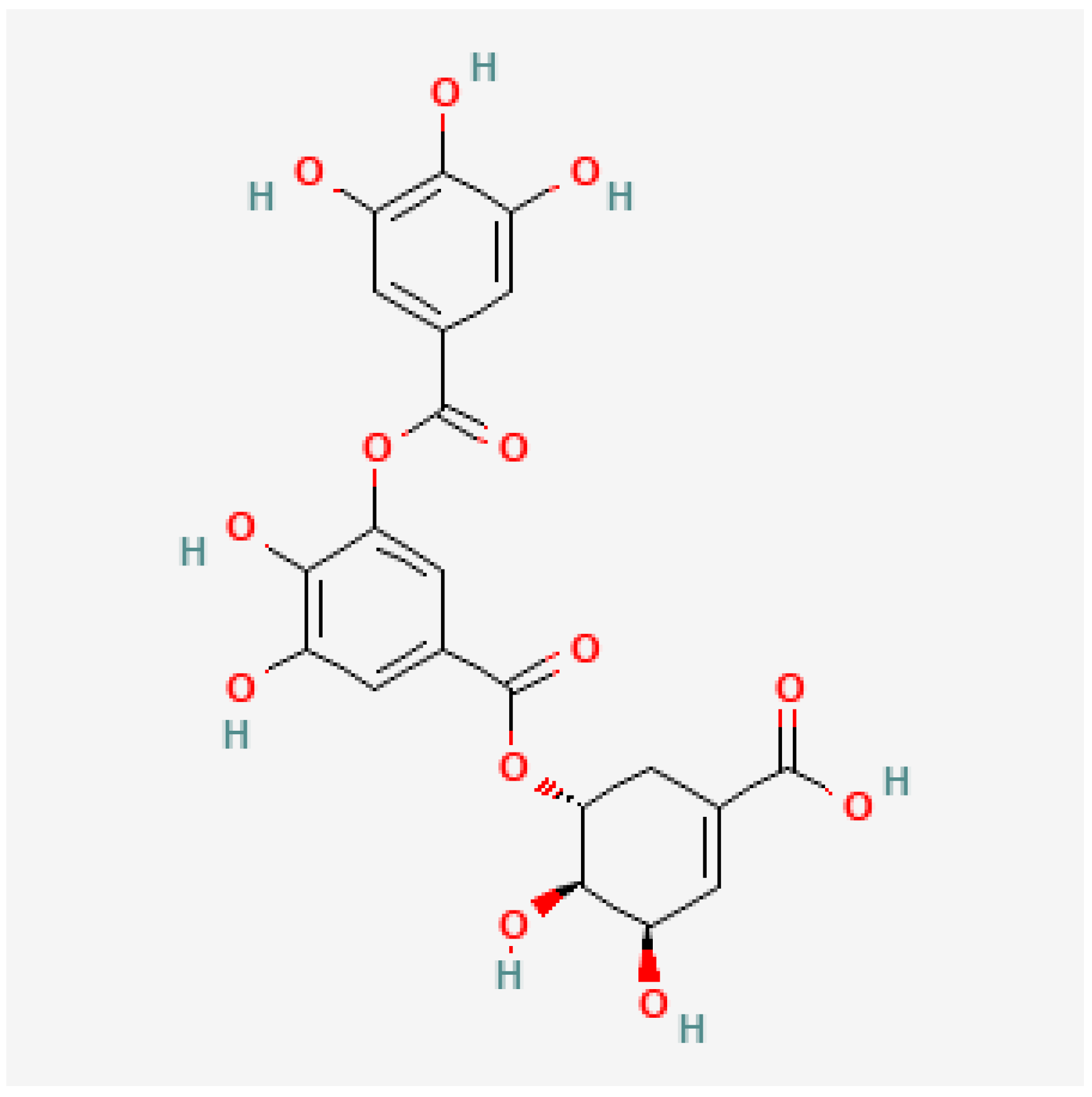
3.3. Tannins
4. Discussion
4.1. Phenolic Acids and/or Derivatives
4.2. Flavonoids
4.3. Tannins
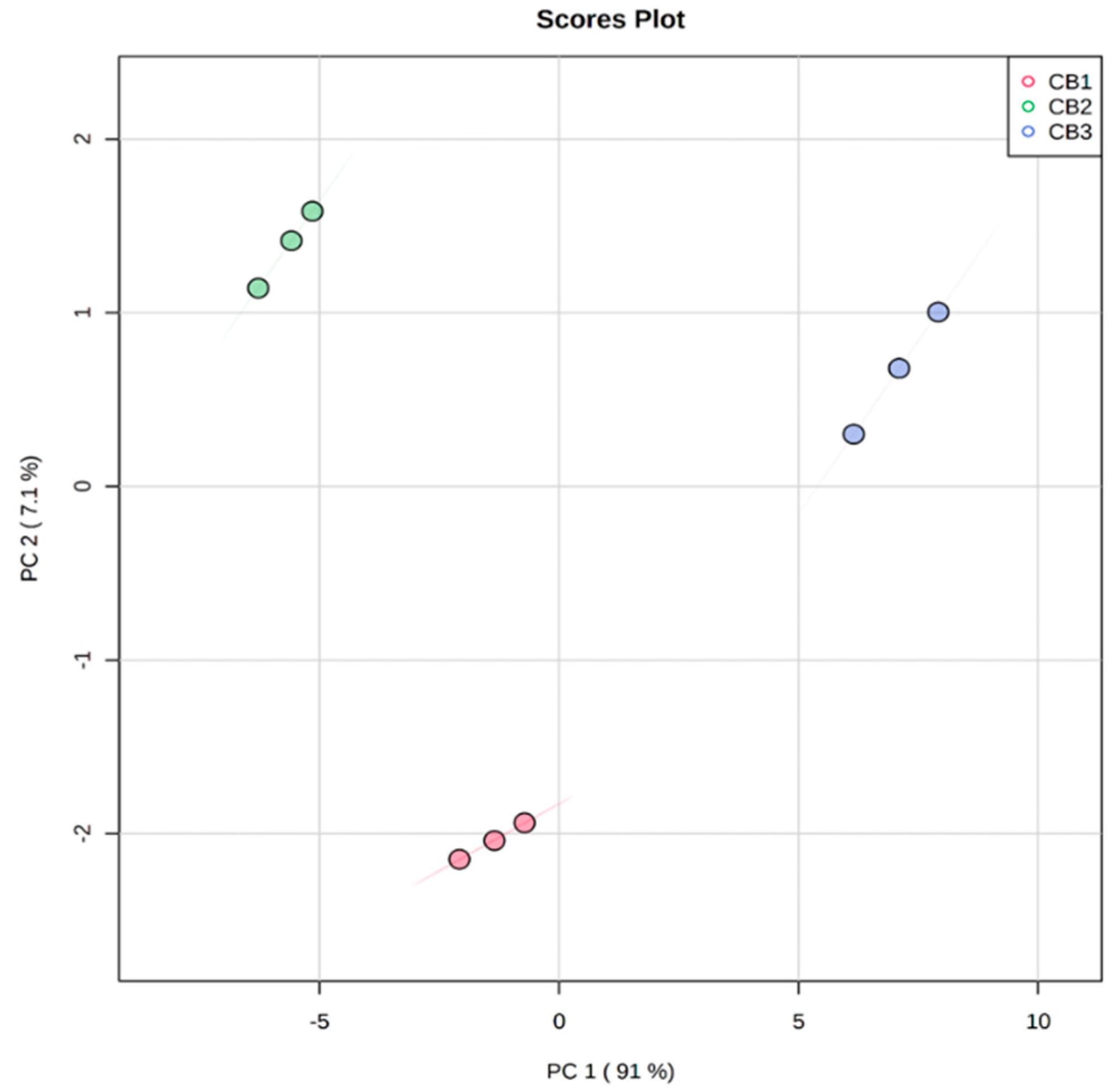
4.4. Multivariate Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Wang, J. Modelling and prediction of global non-communicable diseases. BMC Public Health 2020, 20, 822. [Google Scholar] [CrossRef] [PubMed]
- Bvenura, C.; Sivakumar, D. The role of wild fruits and vegetables in delivering a healthy and balanced diet. Food Res. Int. 2017, 99, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.I.; Wagner, R.G. Weighing up the Costs of Treating ‘Lifestyle’ Diseases in South Africa. 2019. Available online: https//www.wits.ac.za/news/latest-news/opinion/2019/2019-02/weighing-up-the-costs-of-treating-lifestyle-diseases-in-south africa.html#,~,text=Although%20HIV%20still%20causes%20the,are%20poor%2C%20black%20and%20male (accessed on 18 March 2021).
- WHO. Global Action Plan for the Prevention and Control of Noncommunicable Diseases, 2013–2020; WHO Press: Geneva, Switzerland, 2013. [Google Scholar]
- Asowata-Ayodele, A.M.; Afolayan, A.J.; Otunola, G.A. Ethnobotanical survey of culinary herbs and spices used in the traditional medicinal system of Nkonkobe Municipality, Eastern Cape, South Africa. South Afr. J. Bot. 2016, 104, 69–75. [Google Scholar] [CrossRef]
- UNSCN. A World Free from Hunger and All Forms of Malnutrition Is Attainable in This Generation. 2021. Available online: https//www.unscn.org/en/topics/ncds (accessed on 18 March 2021).
- Abegunde, D.O.; Mathers, C.D.; Adam, T.; Ortegon, M.; Strong, K. The burden and costs of chronic diseases in low-income and middle-income countries. Lance 2008, 370, 1929–1938. [Google Scholar] [CrossRef]
- WHO/NCD. Country Profiles, South Africa. 2018. Available online: https//www.who.int/nmh/countries/zaf_en.pdf (accessed on 18 March 2021).
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Murray, A.P.; Murray, M.G. Phytochemistry, traditional uses and bioactivity of the medicinal plant Schinus areira L. (Anacardiaceae), A review. Nat. Prod. J. 2017, 7, 97. [Google Scholar] [CrossRef]
- García, N.; Ormazabal, C. Árboles Nativos de Chile; Enersis, S.A.: Santiago, Chile, 2008; p. 196. [Google Scholar]
- Dellacassa, E. Normalización de Productos Naturales Obtenidos de Especies de la Flora Aromática Latinoamericana, Proyecto CYTED IV.20; Edipucrs: Porto Alegre, Brasil, 2010. [Google Scholar]
- Rebolledo, V.; Otero, M.C.; Delgado, J.M.; Torres, F.; Herrera, M.; Ríos, M.; Cabañas, M.; Martinez, J.L.; Rodríguez-Díaz, M. Phytochemical profile and antioxidant activity of extracts of the peruvian peppertree Schinus areira L. from Chile. Saudi J. Biol. Sci. 2021, 28, 1052–1062. [Google Scholar] [CrossRef]
- Guala, M.S.; Lapissonde, M.O.; Elder, H.V.; van Baren, C.M.; Bandoni, A.L.; Dellacassa, E. Chapter 78—Rose Pepper (Schinus molle L.) Oils. In Essential Oils in Food Preservation, Flavour and Safety; Academic Press: Cambridge, MA, USA, 2016; pp. 689–695. [Google Scholar]
- Gundidza, M. Antimicrobial activity of essential oil from Schinus molle Linn. Cent. Afr. J. Med. 1993, 39, 231–234. [Google Scholar]
- Bendaoud, H.; Romdhane, M.; Souchard, J.P.; Cazaux, S.; Bouajila, J. Chemical composition and anticancer and antioxidant activities of Schinus molle L. and Schinus terebinthifolius Raddi berries essential oils. J. Food Sci. 2010, 75, C466–C472. [Google Scholar] [CrossRef]
- Martins Mdo, R.; Arantes, S.; Candeias, F.; Tinoco, M.T.; Cruz-Morais, J. Antioxidant, antimicrobial and toxicological properties of Schinus molle L. essential oils. J. Ethnopharmacol. 2014, 151, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Guala, M.; Elder, H.; Perez, G.; Chiesa, A. Evaluation of the antioxidant power of fractions of Schinus molle L. essential oil obtained by vacuum distillation. Inf. Technol. 2009, 20, 83–88. [Google Scholar]
- Silva-Junior, E.F.; Aquino, P.G.V.; Santos-Junior, P.F.S.; Nascimento, I.J.S.; Gomes, E.A.; Silva, A.L.L.; Verissimo, R.C.S.S.; Aquino, T.M.; Araújo-Junior, J.X. Phytochemical compounds and pharmacological properties from Schinus molle Linnaeus and Schinus terebinthifolius Raddi (Anacardiaceae). J. Chem. Pharm. Res. 2015, 7, 389–393. [Google Scholar]
- Belhamel, K.; Abderrahim, A.; Ludwig, R. Chemical composition and antibacterial activity of the essential oil of Schinus molle L. grown in Algeria. Int. J. Essent. Oil Ther. 2008, 2, 175–177. [Google Scholar]
- Doleski, M.P.S.; Ferreira, C.C.H.; Calil, B.J.; Palermo, M.M. Chemical composition of the Schinus molle L. essential oil and their biological activities. Rev. Cuba. Farm. 2015, 49, 132–143. [Google Scholar]
- El Hayouni, A.; Chraief, I.; Abedrabba, M.; Bouix, M.; Leveau, J.Y.; Mohammed, H.; Hamdi, M. Tunisian Salvia officinalis L. and Schinus mole L. essential oils, Their chemical compositions and their preservative effects against Salmonella inoculated in minced beef meat. Int. J. Food Microbiol. 2008, 125, 242–251. [Google Scholar] [CrossRef]
- Marzouk, M.S.; Moharram, F.A.; Haggag, E.G.; Ibrahim, M.T.; Badary, O.S. Antioxidant flavonol glycosides from Schinus molle. Phytother. Res. 2006, 20, 200–205. [Google Scholar] [CrossRef]
- Prado, A.C.; Garces, H.G.; Bagagli, E. Schinus molle essential oil as a potential source of bioactive compounds, Antifungal and antibacterial properties. J. Appl. Microbiol. 2018, 126, 516–522. [Google Scholar] [CrossRef]
- Turchetti, G.; Garzoli, S.; Laghezza Masci, V.; Sabia, C.; Iseppi, R.; Giacomello, P.; Tiezzi, A.; Ovidi, E. Antimicrobial Testing of Schinus molle (L.) Leaf Extracts and Fractions Followed by GC-MS Investigation of Biological Active Fractions. Molecules 2020, 25, 1977. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, Z.M.S.; Mohamed, Z.Z.; Hayssam, M.A.; Mamoun, S.M.A. Chemical composition, antioxidant and antibacterial activities of extracts from Schinus molle wood branch growing in Egypt. J. Wood Sci. 2016, 62, 548–561. [Google Scholar]
- Benzi, B.; Stefanazzi, N.; Ferrero, A. Biological activity of essential oils from leaves and fruits of pepper tree (Schinus molle L.) to control rice weevil (Sitophilus oryzae L.). Chil. J. Agric. Res. 2009, 69, 154–159. [Google Scholar] [CrossRef] [Green Version]
- Deveci, O.; Sukan, A.; Tuzun, N.; Kocabas, E.E.H. Chemical composition, repellent and antimicrobial activity of Schinus molle L. J. Med. Plants Res. 2010, 4, 2211–2216. [Google Scholar]
- Rocha, P.M.D.M.; Rodilla, J.M.; Díez, D.; Elder, H. Synergistic antibacterial activity of the essential oil of Aguaribay (Schinus mole L.). Molecules 2012, 17, 12023–12036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eryigit, T.; Yildirim, B.; Ekici, K.; Çirka, M. Chemical composition, antimicrobial and antioxidant properties of Schinus molle L. Essential oil from Turkey. J. Essent. Oil-Bear. Plants 2017, 20, 570–577. [Google Scholar] [CrossRef]
- Feriani, A.; Tir, M.; Mufti, A.; Caravaca, A.; Contreras, M.; Taamalli, A.; Carretero, A.S.; Aldawood, N.; Nahdi, S.; Alwasel, S.; et al. HPLC-ESI-QTOF-MS/MS profiling and therapeutic effects of Schinus terebinthifolius and Schinus molle fruits, investigation of their antioxidant, antidiabetic, anti-inflammatory and antinociceptive properties. Inflammopharmacology 2021, 29, 467–481. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, D.W.; Kim, J.G.; Shin, Y.; Jung, S.K.; Kim, Y.J. Analysis of the Chemical, Antioxidant, and Anti-Inflammatory Properties of Pink Pepper (Schinus molle L.). Antioxidants 2021, 10, 1062. [Google Scholar] [CrossRef]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL, data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI). PubChem Compound Summary for CID 69501207, Feruloyltartaric acid. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Feruloyltartaric-acid (accessed on 20 March 2022).
- National Center for Biotechnology Information (NCBI). PubChem Compound Summary for CID 5274585, Quercetin 3-O-Glucuronide. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/quercetin-3-O-glucuronide (accessed on 20 March 2022).
- National Center for Biotechnology Information (NCBI). PubChem Compound Summary for CID 475266, 3-O-Digalloylshikimic acid. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/3-O-Digalloylshikimic-acid (accessed on 20 March 2022).
- Al Zahrani, N.A.; El-Shishtawy, R.M.; Asiri, A.M. Recent developments of gallic acid derivatives and their hybrids in medicinal chemistry, A review. Eur. J. Med. Chem. 2020, 204, 112609. [Google Scholar] [CrossRef]
- Pawlowska, A.; De Leo, M.; Braca, A. Phenolics of Arbutus unedo L. (Ericaceae) Fruits, Identification of Anthocyanins and Gallic Acid Derivatives. J. Agric. Food Chem. 2006, 54, 10234–10238. [Google Scholar] [CrossRef]
- Inoue, M.; Suzuki, R.; Sakaguchi, N.; Li, Z.; Takeda, T.; Ogihara, Y.; Jiang, B.Y.; Chen, Y. Selective induction of cell death in cancer cells by gallic acid. Biol. Pharm. Bull. 1995, 18, 1526–1530. [Google Scholar] [CrossRef]
- Locatelli, C.; Filippin-Monteiro, F.B.; Creczynski-Pasa, T.B. Alkyl esters of gallic acid as anticancer agents, a review. Eur. J. Med. Chem. 2013, 60, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Swamy, S.K.P.; Govindaswamy, V. Therapeutical properties of ferulic acid and bioavailability enhancement through feruloyl esterase. J. Funct. Foods 2015, 17, 657–666. [Google Scholar] [CrossRef]
- Tlili, N.; Yahia, Y.; Feriani, A.; Labidi, A.; Ghazouani, L.; Nasri, N.; Saadaoui, E.; Khaldi, A. Schinus terebinthifolius vs Schinus molle: A comparative study of the effect of species and location on the phytochemical content of fruits. Ind. Crops Prod. 2018, 122, 559–565. [Google Scholar] [CrossRef]
- Andrae-Marobela, K.; Ghislain, F.W.; Okatch, H.; Majinda, R. Polyphenols, A diverse class of multi-target anti-HIV-1 agents. Curr. Drug Metab. 2013, 7, 392–413. [Google Scholar] [CrossRef] [PubMed]
- Bae, E.A.; Han, M.J.; Lee, M.; Kim, D.H. In vitro inhibitory effect of aome flavonoids on rotavirus infectivity. Biol. Pharm. Bull. 2000, 23, 1122–1124. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.R.; Li, A.X.; Deng, L.B. Progress in the research of inhibitory activities and structure-activity relationship of flavonoids on neuraminidases. Chin. J. New Drugs 2012, 21, 2272–2278. [Google Scholar]
- Morikawa, T.; Ninomiya, K.; Miyake, S.; Miki, Y.; Okamoto, M.; Yoshikawa, M.; Muraoka, O. Flavonol glycosides with lipid accu mulation inhibitory activity and simultaneous quantitative analysis of 15 polyphenols and caffeine in the flower buds of Camellia sinensis from different regions by LCMS. Food Chem. 2013, 140, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J. Dietary flavonoid aglycones and their glycosides, which show better biological significance? Crit. Rev. Food Sci. Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef]
- Bvenura, C.; Witbooi, H.; Kambizi, L. Pigmented Potatoes, A Potential Panacea for Food and Nutrition Security and Health. Foods 2022, 11, 175. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition, antioxidant potential and health benefits of citrus peel. Food Res. Int. 2020, 132, 109114. [Google Scholar] [CrossRef]
- Fan, F.Y.; Sang, L.X.; Jiang, M. Catechins and Their Therapeutic Benefits to Inflammatory Bowel Disease. Molecules 2017, 22, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izawa, K.; Amino, Y.; Kohmura, M.; Ueda, Y.; Kuroda, M. 4.16—Human–Environment Interactions—Taste. Compr. Nat. Prod. II Chem. Biol. 2010, 4, 631–671. [Google Scholar]
- Trugo, L.C.; von Baer, D.; Lupin, E. Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Academic Press: Cambridge, MA, USA, 2003; pp. 3623–3629. [Google Scholar]
- Cheng, H.Y.; Lin, C.C.; Lin, T.C. Antiherpes simplex virus type 2 activity of casuarinin from the bark of Terminalia arjuna Linn. Antivir. Res. 2002, 55, 447–455. [Google Scholar] [CrossRef]
- Quideau, S.; Varadinova, T.; Karagiozova, D.; Jourdes, M.; Pardon, P.; Baudry, C.; Genova, P.; Diakov, T.; Petrova, R. Main structural and stereochemical aspects of the antiherpetic activity of nonahydroxyterphenoyl-containing C-glycosidic ellagitannins. Chem. Biodivers. 2004, 1, 247–258. [Google Scholar] [CrossRef]
- Ashok, P.K.; Upadhyaya, K. Tannins are Astringent. J. Pharmacogn. Phytochem. 2012, 1, 45–50. [Google Scholar]
- Milledge, J.J.; Nielsen, B.V.; Harvey, P.J. The inhibition of anaerobic digestion by model phenolic compounds representative of those from Sargassum muticum. J. Appl. Phycol. 2019, 31, 779–786. [Google Scholar] [CrossRef]
- Nonaka, G.; Nishioka, I.; Nishizawa, M.; Yamagishi, T.; Kashiwada, Y.; Dutschman, G.E.; Bodner, A.J.; Kilkuskie, R.E.; Cheng, Y.C.; Lee, K.H. Anti-AIDS agents, 2, Inhibitory effects of tannins on HIV reverse transcriptase and HIV replication in H9 lymphocyte cells. J. Nat. Prod. 1990, 53, 587–595. [Google Scholar] [CrossRef]
- Wang, Z.; Guillen Quispe, Y.N.; Hwang, S.H.; Zuo, G.; Lim, S.S. Pistafolin B is the major aldose reductase inhibitor of the pods of tara [Caesalpinia spinose (Molina) Kuntze]. Ind. Crops Prod. 2018, 122, 709–715. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, H.; Hou, A.; Zhao, Q.; Wei, T.; Xin, W. Antioxidant properties of two gallotannins isolated from the leaves of Pistacia weinmannifolia. Biochim. Biophys. Acta 2005, 1725, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Sun, H.; Zhao, X.; Hou, J.; Hou, A.; Zhao, Q.; Xin, W. Scavenging of reactive oxygen species and prevention of oxidative neuronal cell damage by a novel gallotannin, pistafolia A. Life Sci. 2002, 70, 1889–1899. [Google Scholar] [CrossRef]
- Hasegawa, T.; Takano, F.; Takata, T.; Niiyama, M.; Ohta, T. Bioactive monoterpene glycosides conjugated with gallic acid from the leaves of Eucalyptus globulus. Phytochemistry 2008, 69, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Koreishi, M.; Tokuda, H.; Yoshida, H.; Nishino, T. Cypellocarpin A–C, phenolic glycosides esterified with oleuropeic acid, from Eucalyptus cypellocarpa. J. Nat. Prod. 2000, 63, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Dhondge, S.S.; Shende, P.H.; Paliwal, L.J.; Deshmukh, D.W. Volumetric and acoustic study of aqueous binary mixtures of quinine hydrochloride, guanidine hydrochloride and quinic acid at different temperatures. J. Chem. Thermodyn. 2015, 81, 34–43. [Google Scholar] [CrossRef]
- Pero, R.W.; Lund, H.; Leanderson, T. Antioxidant metabolism induced by quinic acid increased urinary excretion of tryptophan and nicotinamide. Phytother. Res. 2009, 23, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.A.; Park, D.W.; Kwon, J.E.; Song, H.S.; Park, B.; Jeon, H.; Sohn, E.H.; Koo, H.J.; Kang, S.C. Quinic acid inhibits vascular inflammation in TNF-α-stimulated vascular smooth muscle cells. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 96, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.F.; Li, W.X.; Zhang, F.; Li, Y.H.; Cao, Y.J.; Zhao, Y.; Li, X.L.; Ma, Z.J. Discrimination of Radix Polygoni Multiflori from different geographical areas by UPLC-QTOF/MS combined with chemometrics. Chin. Med. 2017, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Farag, M.A.; Gad, H.A.; Heiss, A.G.; Wessjohann, L.A. Metabolomics driven analysis of six Nigella species seeds via UPLC-qTOF-MS and GC-MS coupled to chemometrics. Food Chem. 2014, 151, 333–342. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT | UV Max | m/z [M-H]− | Tentative Identification | Formula | Class | SH (mg/kg *) | DH (mg/kg *) | H (mg/kg *) |
|---|---|---|---|---|---|---|---|---|
| 5.602 | 229.6 | 331.06824 | β-Glucogallin A | C13H16O10 | Gallotannin | 76.0 | 14.8 | 443.6 |
| 5.787 | 271.6 | 169.01456 | Gallic acid | C7H6O5 | Phenolic acid | 62.7 | 4.9 | 252.3 |
| 6.04 | 273.6 | 331.06738 | β-Glucogallin | C13H16O10 | Gallotannin | 2.1 | nd | 147.8 |
| 6.646 | 268.6 | 343.06802 | Galloyl quinic acid A | C14H16O10 | Gallotannin | 30.8 | nd | 265.8 |
| 7.156 | 280.6 | 343.06808 | Galloyl quinic acid B | C14H16O10 | Gallotannin | 23.9 | nd | 251.9 |
| 7.444 | 255.6 | 411.02328 | Gallic acid derivative | C9H16O18 | Phenolic acid | 99.0 | 77.0 | 166.4 |
| 7.856 | 231.6 | 315.07346 | Gentisic acid 5-O-glucoside | C13H16O9 | Phenolic acid glucoside | 12.2 | nd | 130.3 |
| 8.437 | 261.6 | 325.05603 | Feruloyl tartaric A | C14H14O9 | Phenolic acid ester | 295.7 | 20.2 | 1613.1 |
| 8.804 | 267.6 | 325.05624 | Feruloyl tartaric B | C14H14O9 | Phenolic acid ester | 157.0 | 7.2 | 908.8 |
| 9.347 | 272.6 | 495.07687 | Digalloyl quinic acid | C35H28O22 | Gallotannin | 146.6 | 2.0 | 561.6 |
| 9.556 | 229.6 | 353.08759 | 3-caffeoylquinic acid | C16H18O9 | Phenolic acid ester | 95.1 | 36.4 | 543.9 |
| 10.781 | 274.6 | 577.12457 | Quinic acid derivative | C26H26O15 | organic acid | 9.0 | 20.5 | 31.0 |
| 11.11 | 275.6 | 469.06195 | Gallic acid derivative | C19H18O14 | Phenolic acid | nd | nd | 89.7 |
| 11.227 | 272.6 | 495.0744 | Digalloyl quinic acid | C35H28O22 | Gallotannin | 28.5 | 21.8 | 60.8 |
| 12.537 | 237.6 | 647.0879 | Pistafolin | C28H24O18 | Gallotannin | nd | nd | 147.9 |
| 13.094 | 237.6 | 635.0886 | Trigalloyl glucose | C27H24O18 | Gallotannin | nd | nd | 48.1 |
| 13.432 | 235.6 | 477.06815 | Digalloylshikimic acid A | C21H18O13 | Gallotannin | 65.8 | nd | 524.6 |
| 13.705 | 235.6 | 451.12579 | Galloylsalidroside | C21H24O11 | Gallotannin | nd | nd | 88.1 |
| 14.034 | 274.6 | 477.06717 | Digalloylshikimic acid B | C21H18O13 | Gallotannin | 102.4 | nd | 664.8 |
| 14.464 | 235.6 | 377.1821 | Unidentified | C17H30O9 | 121.4 | 103.0 | 155.4 | |
| 14.919 | 235.6 | 377.18033 | Unidentified | C17H30O9 | 233.3 | 240.7 | 378.0 | |
| 18.168 | 237.6 | 483.18625 | Gallic acid glycoside | C23H32O11 | Gallotannin | 108.1 | nd | 469.1 |
| 18.513 | 237.6 | 939.1104 | Pentagalloyl-β-D-glucose | C41H32O26 | Gallotannin | 1.1 | nd | 96.3 |
| 20.639 | 276.6 | 497.1662 | Eucaglobulin | C23H30O12 | Gallotannins derivative | nd | nd | 174.5 |
| 15.952 | 235.6 | 477.10321 | Isorhamnetin galactoside | C22H22O12 | Flavonol | 38.4 | nd | 430.1 |
| 16.299 | 235.6 | 477.10226 | Isorhamnetin glucoside | C22H22O12 | Flavonol | 27.4 | nd | 377.3 |
| 16.461 | 235.6 | 595.12994 | Quercetin 3-lathyroside | C26H28O16 | Flavonol | nd | nd | 124.0 |
| 16.978 | 237.6 | 615.09918 | Quercetin 3-(2-galloylglucoside) | C28H24O16 | Flavonol | 4.2 | nd | 224.1 |
| 17.67 | 235.6 | 303.04959 | Pentahydroxyflavanone | C15H12O7 | Flavonol | 16.5 | nd | 172.6 |
| 18.004 | 254.6 | 477.0668 | Quercetin 3-O-glucuronide | C21H18O13 | Flavonol | 267.5 | 73.4 | 564.3 |
| 19.198 | 237.6 | 447.08984 | Quercitrin | C21H20O11 | Flavonol | 1.8 | nd | 126.3 |
| 17.812 | 234.6 | 441.0809 | Catechin 3-O-gallate | C22H18O10 | Flavanol | 41.7 | nd | 326.8 |
| 11.651 | 229.6 | 289.07129 | Catechin | C15H14O6 | Flavanol | 122.0 | 80.4 | 551.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bvenura, C.; Kambizi, L. Composition of Phenolic Compounds in South African Schinus molle L. Berries. Foods 2022, 11, 1376. https://doi.org/10.3390/foods11101376
Bvenura C, Kambizi L. Composition of Phenolic Compounds in South African Schinus molle L. Berries. Foods. 2022; 11(10):1376. https://doi.org/10.3390/foods11101376
Chicago/Turabian StyleBvenura, Callistus, and Learnmore Kambizi. 2022. "Composition of Phenolic Compounds in South African Schinus molle L. Berries" Foods 11, no. 10: 1376. https://doi.org/10.3390/foods11101376