
Foodborne Parasites and Their Complex Life Cycles Challenging Food Safety in Different Food Chains

{kind=link}
Abstract
:1. Introduction
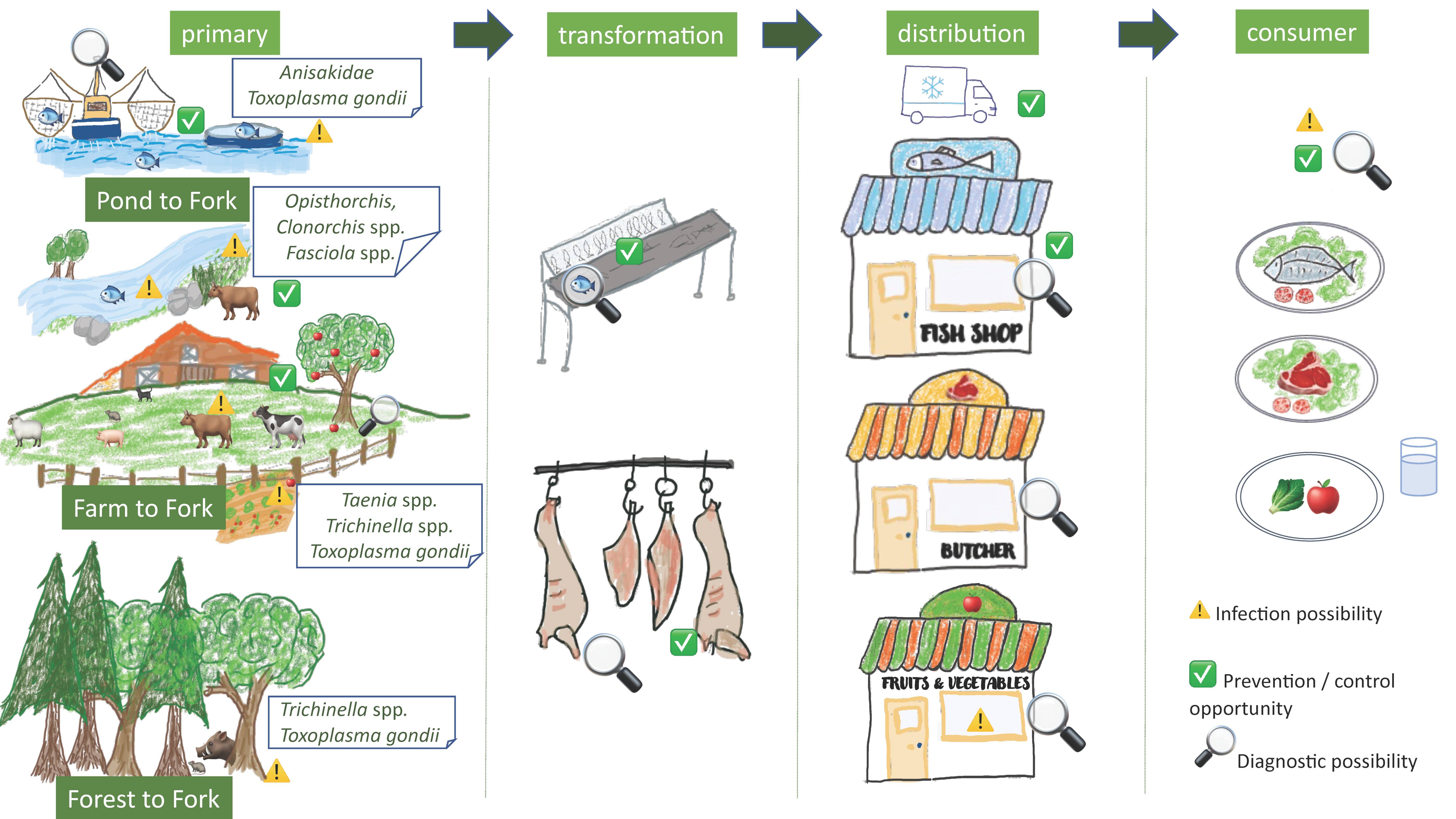
2. Farm to Fork and Forest to Fork
2.1. Taenia solium, Taenia saginata
2.1.1. Introduction
2.1.2. Localisation of the Infection Risks for the Consumer in the Food Chain
2.1.3. Diagnostic Options in the Food Chain
2.1.4. Prevention and Control Options in the Food Chain
2.2. Trichinella spp.
2.2.1. Introduction
2.2.2. Localisation of the Infection Risks for the Consumer in the Food Chain
2.2.3. Diagnostic Options in the Food chain
2.2.4. Prevention and Control Options in the Food Chain
2.3. Toxoplasma gondii
2.3.1. Introduction
2.3.2. Localisation of the Infection Risks for the Consumer in the Food Chain
2.3.3. Diagnostic Options in the Food Chain
2.3.4. Prevention and Control Options in the Food Chain
2.4. Fasciola spp.
2.4.1. Introduction
2.4.2. Localisation of the Infection Risks for the Consumer in the Food Chain
2.4.3. Diagnostic Options in the Food Chain
2.4.4. Prevention and Control Options in the Food Chain
3. Pond/Ocean/River to Fork: Fish-Borne Parasites
3.1. Fish Borne Trematodes: Clonorchis, Opisthorchis spp.
3.1.1. Introduction
3.1.2. Localisation of the Infection Risks for the Consumer in the Food Chain
3.1.3. Diagnostic Options in the Food Chain
3.1.4. Prevention and Control Options in the Food Chain
3.2. Anisakidae
3.2.1. Introduction
3.2.2. Localisation of the Infection Risks for the Consumer in the Food Chain
3.2.3. Diagnostic Options in the Food Chain
3.2.4. Prevention and Control Options in the Food Chain
4. Discussion and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- FAO/WHO (Food and Agriculture Organization of the United Nations/World Health Organization). Multicriteria-Based Ranking for Risk Management of Food-Borne Parasites; Microbiological Risk Assessment Series No. 23, Rome; World Health Organization: Geneva, Switzerland, 2014; p. 302. [Google Scholar]
- WHO. World Health Organisation, WHO Estimates of the Global Burden of Foodborne Diseases Foodborne Disease Burden Epidemiology Reference Group 2007–2015; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Gabriël, S.; Johansen, M.V.; Pozio, E.; Smit, G.S.; Devleesschauwer, B.; Allepuz, A.; Papadopoulos, E.; van der Giessen, J.; Dorny, P. Human migration and pig/pork import in the European Union: What are the implications for Taenia solium infections? Vet. Parasitol. 2015, 213, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Murrell, K.D.; Dorny, P.; Flisser, A.; Geerts, S.; Kyvsgaard, N.C.; McManus, D.; Nash, T.E.; Pawlowski, Z.S. WHO/FAO/OIE Guidelines for the Surveillance, Prevention & Control of Taeniosis/Cysticercosis; Murrell, K.D., Ed.; OIE: Paris, France, 2005. [Google Scholar]
- Jansen, F.; Dorny, P.; Berkvens, D.; Van Hul, A.; Van den Broeck, N.; Makay, C.; Praet, N.; Eichenberger, R.M.; Deplazes, P.; Gabriël, S. High prevalence of bovine cysticercosis found during evaluation of different post-mortem detection techniques in Belgian slaughterhouses. Vet. Parasitol. 2017, 244, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, C.; Mkupasi, E.M.; Ngowi, H.A.; Forkman, B.; Johansen, M.V. Severe seizures in pigs naturally infected with Taenia solium in Tanzania. Vet. Parasitol. 2016, 220, 67–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevisan, C.; Johansen, M.V.; Mkupasi, E.M.; Ngowi, H.A.; Forkman, B. Disease behaviours of sows naturally infected with Taenia solium in Tanzania. Vet. Parasitol. 2017, 235, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Jansen, F.; Dorny, P.; Trevisan, C.; Dermauw, V.; Laranjo-González, M.; Allepuz, A.; Dupuy, C.; Krit, M.; Gabriël, S.; Devleesschauwer, B. Economic impact of bovine cysticercosis and taeniosis caused by Taenia saginata in Belgium. Parasit. Vectors 2018, 11, 241. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, E.C.; Mwape, K.E.; Devleesschauwer, B.; Gabriël, S.; Chembensofu, M.; Mambwe, M.; Phiri, I.K.; Masuku, M.; Zulu, G.; Colston, A.; et al. Taenia solium from a community perspective: Preliminary costing data in the Katete and Sinda districts in Eastern Zambia. Vet. Parasitol. 2018, 251, 63–67. [Google Scholar] [CrossRef]
- Trevisan, C.; Devleesschauwer, B.; Schmidt, V.; Winkler, A.S.; Harrison, W.; Johansen, M.V. The societal cost of Taenia solium cysticercosis in Tanzania. Acta Trop. 2017, 165, 141–154. [Google Scholar] [CrossRef]
- Saelens, G.; Robertson, L.; Gabriël, S. Diagnostic tools for the detection of taeniid eggs in different environmental matrices: A systematic review. Food Waterborne Parasitol. 2022, 26, e00145. [Google Scholar] [CrossRef]
- Jansen, F.; Dorny, P.; Gabriël, S.; Dermauw, V.; Johansen, M.V.; Trevisan, C. The survival and dispersal of Taenia eggs in the environment: What are the implications for transmission? A systematic review. Parasit. Vectors 2021, 14, 88. [Google Scholar] [CrossRef]
- Bucur, I.; Gabriël, S.; Van Damme, I.; Dorny, P.; Vang Johansen, M. Survival of Taenia saginata eggs under different environmental conditions. Vet. Parasitol. 2019, 266, 88–95. [Google Scholar] [CrossRef]
- Gabriël, S.; Mwape, K.E.; Phiri, I.K.; Devleesschauwer, B.; Dorny, P. Taenia solium control in Zambia: The potholed road to success. Parasite Epidemiol. Control 2018, 4, e00082. [Google Scholar] [CrossRef]
- Thys, S.; Mwape, K.E.; Lefèvre, P.; Dorny, P.; Phiri, A.M.; Marcotty, T.; Phiri, I.K.; Gabriël, S. Why pigs are free-roaming: Communities’ perceptions, knowledge and practices regarding pig management and taeniosis/cysticercosis in a Taenia solium endemic rural area in Eastern Zambia. Vet. Parasitol. 2016, 225, 33–42. [Google Scholar] [CrossRef]
- Dorny, P.; Phiri, I.K.; Vercruysse, J.; Gabriel, S.; Willingham, A.L., 3rd; Brandt, J.; Victor, B.; Speybroeck, N.; Berkvens, D. A Bayesian approach for estimating values for prevalence and diagnostic test characteristics of porcine cysticercosis. Int. J. Parasitol. 2004, 34, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Chembensofu, M.; Mwape, K.E.; Van Damme, I.; Hobbs, E.; Phiri, I.K.; Masuku, M.; Zulu, G.; Colston, A.; Willingham, A.L.; Devleesschauwer, B.; et al. Re-visiting the detection of porcine cysticercosis based on full carcass dissections of naturally Taenia solium infected pigs. Parasit. Vectors 2017, 10, 572. [Google Scholar] [CrossRef] [Green Version]
- Jansen, F.; Dorny, P.; Berkvens, D.; Gabriël, S. Bovine cysticercosis and taeniosis: The effect of an alternative post-mortem detection method on prevalence and economic impact. Prev. Vet. Med. 2018, 161, 1–8. [Google Scholar] [CrossRef]
- Waema, M.W.; Misinzo, G.; Kagira, J.M.; Agola, E.L.; Ngowi, H.A. DNA-Detection Based Diagnostics for Taenia solium Cysticercosis in Porcine. J. Parasitol. Res. 2020, 2020, 5706981. [Google Scholar] [CrossRef]
- EU. Commission implementing regulation (EU) 2019/627 of 15 March 2019 laying down uniform practical arrangements for the performance of official controls on products of animal origin intended for human consumption in accordance with Regulation (EU) 2017/625 of the European Parliament and of the Council and amending Commission Regulation (EC) No 2074/2005 as regards official controls. Off. J. Eur. Union 2019, L 131, 72. [Google Scholar]
- Alban, L.; Häsler, B.; van Schaik, G.; Ruegg, S. Risk-based surveillance for meat-borne parasites. Exp. Parasitol. 2020, 208, 107808. [Google Scholar] [CrossRef]
- Lightowlers, M.W. Vaccines for prevention of cysticercosis. Acta Trop. 2003, 87, 129–135. [Google Scholar] [CrossRef]
- Lightowlers, M.W. Eradication of Taenia solium cysticercosis: A role for vaccination of pigs. Int. J. Parasitol. 2010, 40, 1183–1192. [Google Scholar] [CrossRef]
- Gabriël, S.; Mwape, K.E.; Hobbs, E.C.; Devleesschauwer, B.; Van Damme, I.; Zulu, G.; Mwelwa, C.; Mubanga, C.; Masuku, M.; Mambwe, M.; et al. Evidence for potential elimination of active Taenia solium transmission in Africa? N. Engl. J. Med. 2020, 383, 396–397. [Google Scholar] [CrossRef] [PubMed]
- De Coster, T.; Van Damme, I.; Baauw, J.; Gabriël, S. Recent advancements in the control of Taenia solium: A systematic review. Food Waterborne Parasitol. 2018, 13, e00030. [Google Scholar] [CrossRef] [PubMed]
- Pozio, E. How globalization and climate change could affect foodborne parasites. Exp. Parasitol. 2020, 208, 107807. [Google Scholar] [CrossRef] [PubMed]
- Murrell, K.D.; Pozio, E. Worldwide Occurrence and Impact of Human Trichinellosis, 1986–2009. Emerg. Infect. Dis. 2011, 17, 2194–2202. [Google Scholar] [CrossRef] [PubMed]
- Pozio, E. World distribution of Trichinella spp. infections in animals and humans. Vet. Parasitol. 2007, 149, 3–21. [Google Scholar] [CrossRef]
- Pozio, E.; Hoberg, E.; La Rosa, G.; Zarlenga, D.S. Molecular taxonomy, phylogeny and biogeography of nematodes belonging to the Trichinella genus. Infect Genet. Evol. 2009, 9, 606–616. [Google Scholar] [CrossRef]
- Crisóstomo-Jorquera, V.; Landaeta-Aqueveque, C. The genus Trichinella and its presence in wildlife worldwide: A review. Transbound. Emerg. Dis. 2022, 69, e1269–e1279. [Google Scholar] [CrossRef]
- Boireau, P.; Vallée, I.; Roman, T.; Perret, C.; Mingyuan, L.; Gamble, H.R.; Gajadhar, A. Trichinella in horses: A low frequency infection with high human risk. Vet. Parasitol. 2000, 93, 309–320. [Google Scholar] [CrossRef]
- Gottstein, B.; Pozio, E.; Nöckler, K. Epidemiology, diagnosis, treatment, and control of trichinellosis. Clin. Microbiol. Rev. 2009, 22, 127–145. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Boireau, P. Trichinellosis in China: Epidemiology and control. Trends Parasitol. 2002, 18, 553–556. [Google Scholar] [CrossRef]
- Noeckler, K.; Pozio, E.; van der Giessen, J.; Hill, D.E.; Gamble, H.R. International Commission on Trichinellosis: Recommendations on post-harvest control of Trichinella in food animals. Food Waterborne Parasitol. 2019, 21, e00041. [Google Scholar] [CrossRef]
- Gamble, H.R. Trichinella spp. control in modern pork production systems. Food Waterborne Parasitol. 2022, 28, e00172. [Google Scholar] [CrossRef]
- Bruschi, F.; Gómez-Morales, M.A.; Hill, D.E. International Commission on Trichinellosis: Recommendations on the use of serological tests for the detection of Trichinella infection in animals and humans. Food Waterborne Parasitol. 2019, 5, e00032. [Google Scholar] [CrossRef]
- Barlow, A.; Roy, K.; Hawkins, K.; Ankarah, A.A.; Rosenthal, B. A review of testing and assurance methods for Trichinella surveillance programs. Food Waterborne Parasitol 2021, 9, e00129. [Google Scholar] [CrossRef]
- Li, X.; Liu, W.; Wang, J.; Zou, D.; Wang, X.; Yang, Z.; Yin, Z.; Cui, Q.; Shang, W.; Li, H.; et al. Rapid detection of Trichinella spiralis larvae in muscles by loop-mediated isothermal amplification. Int. J. Parasitol 2012, 42, 1119–1126. [Google Scholar] [CrossRef]
- Van Knapen, F. Control of trichinellosis by inspection and farm management practices. Vet. Parasitol. 2000, 93, 385–392. [Google Scholar] [CrossRef]
- Kapel, C.M. Changes in the EU legislation on Trichinella inspection—New challenges in the epidemiology. Vet. Parasitol. 2005, 132, 189–194. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Trichinellosis. In ECDC. Annual Epidemiological Report for 2019; ECDC: Stockholm, Sweden, 2021; Available online: https://www.ecdc.europa.eu/en/publications-data/trichinellosis-annual-epidemiological-report-2019 (accessed on 23 November 2022).
- Dupouy-Camet, J.; Murrell, K.D. FAO/WHO/OIE Guidelines for the Surveillance, Management. Prevention and Control of Trichinellosis; OIE: Paris, France, 2007; pp. 101–110. [Google Scholar]
- Burke, R.; Masuoka, P.; Murrell, K.D. Swine Trichinella infection and geographic information system tools. Emerg. Infect Dis. 2008, 14, 1109–1111. [Google Scholar] [CrossRef]
- Tang, B.; Li, J.; Li, T.; Xie, Y.; Guan, W.; Zhao, Y.; Yang, S.; Liu, M.; Xu, D. Vaccines as a Strategy to Control Trichinellosis. Front. Microbiol. 2022, 23, 857786. [Google Scholar] [CrossRef]
- Tenter, A.M.; Heckeroth, A.; Weiss, L. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef] [Green Version]
- Torgerson, P.R.; Mastroiacovo, P. The global burden of congenital toxoplasmosis: A systematic review. Bull. WHO 2013, 91, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Toxoplasmosis of Animals and Humans; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Cook, A.J.; Gilbert, R.E.; Buffolano, W.; Zufferey, J.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of toxoplasma infection in pregnant women: European multi- centre case-control study. European Research Network on Congenital Toxoplasmosis. British Med. J. 2000, 321, 142–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenter, A.M. Toxoplasma gondii in animals used for human consumption. Mem. Inst. Oswaldo Cruz. 2009, 104, 364–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, D.; Coss, C.; Dubey, J.P.; Wroblewski, K.; Sautter, M.; Hosten, T.; Munoz-Zanzi, C.; Mui, E.; Withers, S.; Boyer, K.; et al. Identification of a sporozoite-specific antigen from Toxoplasma gondii. J. Parasitol. 2011, 97, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Kijlstra, A.; Meerburg, B.G.; Bos, A.P. Food safety in free-range and organic livestock systems: Risk management and responsibility. J. Food Prot. 2009, 72, 2629–2637. [Google Scholar] [CrossRef]
- Verhelst, D.; De Craeye, S.; Vanrobaeys, M.; Czaplicki, G.; Dorny, P.; Cox, E. Seroprevalence of Toxoplasma gondii in domestic sheep in Belgium. Vet. Parasitol. 2014, 205, 57–61. [Google Scholar] [CrossRef]
- Dubey, J.P.; Jones, J.L. Toxoplasma gondii infection in humans and animals in the United States. Int. J. Parasitol. 2008, 38, 1257–1278. [Google Scholar] [CrossRef]
- Bacci, C.; Vismarra, A.; Mangia, C.; Bonardi, S.; Bruini, I.; Genchi, M.; Kramer, L.; Brindani, F. Detection of Toxoplasma gondii in free-range, organic pigs in Italy using serological and molecular methods. Int. J. Food Microbiol. 2015, 202, 54–56. [Google Scholar] [CrossRef]
- De Berardinis, A.; Paludi, D.; Pennisi, L.; Vergara, A. Toxoplasma gondii, a foodborne pathogen in the swine production chain from a European perspective. Foodborne Pathogens Dis. 2017, 14, 637–648. [Google Scholar] [CrossRef]
- Kijlstra, A.; Eissen, O.A.; Cornelissen, J.; Munniksma, K.; Eijck, I.; Kortbeek, T. Toxoplasma gondii infection in animal-friendly pig production systems. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3165–3169. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.; Aubert, D.; Escotte-Binet, S.; Durand, B.; Robert, C.; Geers, R.; Alliot, A.; Belbis, G.; Villena, I.; Blaga, R. Anatomical distribution of Toxoplasma gondii in naturally and experimentally infected lambs. Parasite 2022, 29, 3. [Google Scholar] [CrossRef]
- Shapiro, K.; Kim, M.; Rajal, V.B.; Arrowood, M.J.; Packham, A.; Aguilar, B.; Wuertz, S. Simultaneous detection of four protozoan parasites on leafy greens using a novel multiplex PCR assay. Food Microbiol. 2019, 84, 103252. [Google Scholar] [CrossRef]
- Dumètre, A.; Dardé, M.L. How to detect Toxoplasma gondii oocysts in environmental samples? FEMS Microbiol. Rev. 2003, 27, 651–661. [Google Scholar] [CrossRef] [Green Version]
- Mirza Alizadeh, A.; Jazaeri, S.; Shemshadi, B.; Hashempour-Baltork, F.; Sarlak, Z.; Pilevar, Z.; Hosseini, H.A. Review on inactivation methods of Toxoplasma gondii in foods. Pathog. Glob. Health 2018, 112, 306–319. [Google Scholar] [CrossRef]
- Shapiro, K.; Vanwormer, E.; Aguilar, B.; Conrad, P.A. Surveillance for Toxoplasma gondii in California mussels (Mytilus californianus) reveals transmission of atypical genotypes from land to sea. Environ. Microbiol. 2015, 17, 4177–4188. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Collins, M.V.; Mitchell, S.M.; Wetch, C.N.; Rosypal, A.C.; Flick, G.J.; Zajac, A.M.; Lindquist, A.; Dubey, J.P. Survival of Toxoplasma gondii oocysts in eastern oysters (Crassostrea virginica). J. Parasitol. 2004, 90, 1054–1057. [Google Scholar] [CrossRef]
- Hohweyer, J.; Dumètre, A.; Aubert, D.; Azas, N.; Villena, I. Tools and methods for detecting and characterizing Giardia, Cryptosporidium, and Toxoplasma parasites in marine mollusks. J. Food Prot. 2013, 76, 1649–1657. [Google Scholar] [CrossRef]
- Gisbert Algaba, I.; Geerts, M.; Jennes, M.; Coucke, W.; Opsteegh, M.; Cox, E.; Dorny, P.; Dierick, K.; De Craeye, S. A more sensitive, efficient and ISO 17025 validated Magnetic Capture real time PCR method for the detection of archetypal Toxoplasma gondii strains in meat. Int. J. Parasitol. 2017, 47, 875–884. [Google Scholar] [CrossRef]
- Opsteegh, M.; Dam-Deisz, C.; de Boer, P.; De Craeye, S.; Faré, A.; Hengeveld, P.; Luiten, R.; Schares, G.; van Solt-Smits, C.; Verhaegen, B.; et al. Methods to assess the effect of meat processing on viability of Toxoplasma gondii: Towards replacement of mouse bioassay by in vitro testing. Int. J. Parasitol. 2020, 50, 357–369. [Google Scholar] [CrossRef]
- Opsteegh, M.; Maas, M.; Schares, G.; van der Giessen, J. Relationship between seroprevalence in the main livestock species and presence of Toxoplasma gondii in meat (GP/EFSA/BIOHAZ/2013/01) An extensive literature review, Final report. EFSA Support Publ. 2016, 13, 1–294. [Google Scholar] [CrossRef] [Green Version]
- Zdolec, N.; Kiš, M. Meat safety from farm to slaughter—Risk-based control of Yersinia enterocolitica and Toxoplasma gondii. Processes 2021, 9, 815. [Google Scholar] [CrossRef]
- Wehbe, K.; Pencole, L.; Lhuaire, M.; Sibiude, J.; Mandelbrot, L.; Villena, I.; Picone, O. Hygiene measures as primary prevention of toxoplasmosis during pregnancy: A systematic review. J. Gynecol. Obstet. Hum. Reprod. 2022, 51, 102300. [Google Scholar] [CrossRef] [PubMed]
- EU. Commission Regulation No. 219/2014 Commission Regulation (EU) No 219/2014 of 7 March 2014 amending Annex I to Regulation (EC) No 854/2004 of the European Parliament and of the Council as regards the specific requirements for post-mortem inspection of domestic swine. Off. J. Eur. Union 2014, L 69, 99–100. [Google Scholar]
- EFSA (European Food Safety Agency). Scientific Opinion on the public health hazard to be covered by inspection of meat (swine). EFSA J. 2011, 9, 2351. [Google Scholar] [CrossRef] [Green Version]
- Rani, S.; Pradhan, A.K. Evaluating uncertainty and variability associated with Toxoplasma gondii survival during cooking and low temperature storage of fresh cut meats. Int. J. Food Microbiol. 2021, 341, 109031. [Google Scholar] [CrossRef]
- Buxton, D.; Thomson, K.; Maley, S.; Wright, S.; Bos, H.J. Vaccination of sheep with a live incomplete strain (S48) of Toxoplasma gondii and their immunity to challenge when pregnant. Vet. Rec. 1991, 129, 89–93. [Google Scholar] [CrossRef]
- Yuan, Z.G.; Zhang, X.X.; Lin, R.Q.; Petersen, E.; He, S.; Yu, M.; He, X.H.; Zhou, D.H.; He, Y.; Li, H.X.; et al. Protective effect against toxoplasmosis in mice induced by DNA immunization with gene encoding Toxoplasma gondii ROP18. Vaccine 2011, 29, 6614–6619. [Google Scholar] [CrossRef]
- Bonačić Marinović, A.A.; Opsteegh, M.; Deng, H.; Suijkerbuijk, A.W.M.; van Gils, P.F.; van der Giessen, J. Prospects of toxoplasmosis control by cat vaccination. Epidemics 2020, 30, 100380. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Esteban, J.; Bargues, M. Epidemiology of human fascioliasis: A review and proposed new classification. Bull. WHO 1999, 77, 340–346. [Google Scholar]
- Chen, M.G.; Mott, K.E. Progress in assessment of morbidity due to Fasciola hepatica infection: A review of recent literature. Trop Dis. Bull 1990, 87, 1–38. [Google Scholar]
- WHO. Working to Overcome the Global Impact of Neglected Tropical Diseases. First WHO Report on Neglected Tropical Diseases; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar] [CrossRef]
- Fürst, T.; Keiser, J.; Utzinger, J. Global burden of human food-borne trematodiasis: A systematic review and meta-analysis. Lancet Inf. Dis. 2012, 12, 210–221. [Google Scholar] [CrossRef]
- Havelaar, A.; Kirk, M.; Torgerson, P.; Gibb, H.; Hald, T.; Lake, R.; Praet, N.; Bellinger, D.; de Silva, N.; Gargouri, N.; et al. Disease Burden Epidemiology Reference Group. World Health Organization global estimates and regional comparisons of the burden of foodborne disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, D. Homing in on helminths. Am. J. Trop. 1992, 46, 626–634. [Google Scholar] [CrossRef]
- Rim, H.; Madsen, H.; Cross, J.H.; Rim, H.; Farag, H.F.; Sornmani, S. Food-borne trematodes: Ignored or emerging? Food-borne Ignored or Emerging? Parasitol. Today 1994, 10, 207–209. [Google Scholar] [CrossRef]
- Keiser, J.; Utzinger, J. Emerging foodborne trematodiasis. Emerg. Inf. Dis. 2005, 11, 1507–1514. [Google Scholar] [CrossRef]
- WHO. Control of Foodborne Trematode infections: Report of a WHO Study Group; 1995 WHO Technical Report Series 894; WHO: Geneva, Switzerland, 1995. [Google Scholar]
- Dermauw, V.; Muchai, J.; Kappany, Y.; Al Fajardo, A.L.; Dorny, P. Human fascioliasis in Africa: A systematic review. PLoS ONE 2021, 16, e0261166. [Google Scholar] [CrossRef] [PubMed]
- Dorny, P.; Praet, N.; Deckers, N.; Gabriel, S. Emerging food-borne parasites. Vet. Parasitol. 2009, 163, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Mas-Coma, S.; Valero, M.; Bargues, M. Chapter 2—Fasciola, lymnaeids and human fascioliasis, with a global overview on disease transmission, epidemiology, evolutionary genetics, molecular epidemiology and control. In Advances in Parasitology, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar] [CrossRef]
- Fox, N.J.; White, P.C.L.; McClean, C.J.; Marion, G.; Evans, A.; Hutchings, M.R. Predicting impacts of climate change on fasciola hepatica risk. PLoS ONE 2011, 6, 19–21. [Google Scholar] [CrossRef] [PubMed]
- Mas-Coma, S.; Valero, M.A.; Bargues, M.D. Climate change effects on trematodiases, with emphasis on zoonotic fascioliasis and schistosomiasis. Vet. Parasitol. 2009, 163, 264–280. [Google Scholar] [CrossRef]
- Mas-Coma, S. Human fascioliasis emergence risks in developed countries: From individual patients and small epidemics to climate and global change impacts. Enferm. Infec. y Microbiol. Clinic. 2020, 38, 253–256. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Bargues, M.D.; Valero, M.A. Human fascioliasis infection sources, their diversity, incidence factors, analytical methods and prevention measures. Parasitology 2018, 145, 1665–1699. [Google Scholar] [CrossRef] [Green Version]
- Keiser, J.; Utzinger, J. Food-borne trematodiases. Clin. Microbiol. Rev. 2009, 22, 466–483. [Google Scholar] [CrossRef]
- Mas-Coma, S. Human Fascoliasis: Epidemiological patterns in human endemic areas of South America, Africa and Asia. Southeast Asian J. Trop. Med. Public Health 2004, 35, 1–11. [Google Scholar]
- Taghipour, A.; Zaki, L.; Rostami, A.; Foroutan, M.; Ghaffarifar, F.; Fathi, A.; Abdoli, A. Highlights of human ectopic fascioliasis: A systematic review. Inf. Dis. 2019, 51, 785–792. [Google Scholar] [CrossRef]
- Mas-Coma, S.; Agramunt, V.; Valero, M. Chapter 2. Neurological and Ocular Fascioliasis in Humans. In Advances in Parasitology; Academic Press: Cambridge, MA, USA, 2014; Volume 84. [Google Scholar] [CrossRef]
- González-Miguel, J.; Valero, M.A.; Reguera-Gomez, M.; Mas-Bargues, C.; Bargues, M.D.; Simón, F.; Mas-Coma, S. Numerous Fasciola plasminogen-binding proteins may underlie blood-brain barrier leakage and explain neurological disorder complexity and heterogeneity in the acute and chronic phases of human fascioliasis. Parasitol 2019, 146, 284–298. [Google Scholar] [CrossRef] [Green Version]
- Mailles, A.; Capek, A.; Ajana, F.; Schepens, C.; Ilef, D.; Vaillant, V. Commercial watercress as an emerging source of fascioliasis in Northern France in 2002: Results from an outbreak investigation. Epidemiol. Inf. 2006, 134, 942–945. [Google Scholar] [CrossRef]
- Micic, D.; Oto, A.; Charlton, M.R.; Benoit, J.-L.; Siegler, M. Hiding in the Water. N. Engl. J. Med. 2020, 382, 1844–1849. [Google Scholar] [CrossRef]
- Sah, R.; Khadka, S.; Khadka, M.; Gurubacharya, D.; Sherchand, J.B.; Parajuli, K.; Shah, N.P.; Kattel, H.P.; Pokharel, B.M.; Rijal, B. Human fascioliasis by Fasciola hepatica: The first case report in Nepal. BMC Res. Notes 2017, 10, 10–13. [Google Scholar] [CrossRef] [Green Version]
- Goral, V.; Senturk, S.; Mete, O.; Cicek, M.; Ebik, B.; Kaya, B. A Case of Biliary Fascioliasis by Fasciola gigantica in Turkey. Korean J. Parasitol. 2011, 49, 65–68. [Google Scholar] [CrossRef]
- Nino Incani, R.; Vieira, J.; Pacheco, M.; Planchart, S.; Amarista, M.; Lazdins, J. Human infection by Fasciola hepatica in Venezuela: Report of a geriatric case. Investig. Clin. 2003, 44, 255–260. [Google Scholar]
- Ashrafi, K.; Valero, M.A.; Massoud, J.; Sobhani, A.; Solaymani-Mohammadi, S.; Conde, P.; Khoubbane, M.; Bargues, M.D.; Mas-Coma, S. Plant-borne human contamination by fascioliasis. Am. J. Trop. Med. Hyg. 2006, 75, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milas, S.; Rossi, C.; Philippart, I.; Dorny, P.; Bottieau, E. Autochthonous human Fascioliasis, Belgium. Em. Infect. Dis. 2020, 26, 155–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Ulloa, C.; Rivera-jacinto, M.; del Valle-Mendoza, J.; Cerna, C.; Hoban, C.; Ortiz, P. Risk factors for human fascioliasis in schoolchildren in Baños del Inca. Trans. Royal Soc. Trop. Med. Hyg. 2018, 112, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Park, S.; Hong, S.; Shin, H.; Jung, B.K.; Kim, M.J. Green vegetable juice as a potential source of human fascioliasis in Korea. One Health 2022, 15, 100441. [Google Scholar] [CrossRef] [PubMed]
- Curtale, F.; Mas-Coma, S.; Hassanein, Y.A.W.; Barduagni, P.; Pezzotti, P.; Savioli, L. Clinical signs and household characteristics associated with human fascioliasis among rural population in Egypt: A case-control study. Parassitologia 2003, 45, 5–11. [Google Scholar]
- Esteban, J.G.; Gonzalez, C.; Curtale, F.; Muñoz-Antoli, C.; Valero, M.; Bargues, M.; El Sayed, M.; El Wakeel, A.; Abdel-Wahab, Y.; Montresor, A.; et al. Hyperendemic fascioliasis associated with schistosomiasis in villages in the Nile Delta of Egypt. Am J Trop. Med. Hyg. 2003, 69, 429–437. [Google Scholar] [CrossRef]
- Mas-Coma, S. Human fascioliasis. In Waterborne Zoonoses: Identification, Causes and Control; IWA Publish: London, UK, 2004. [Google Scholar]
- Taira, N.; Yoshifuji, H.; Boray, J.C. Zoonotic potential of infection with Fasciola spp. by consumption of freshly prepared raw liver containing immature flukes. Int. J. Parasitol. 1997, 27, 775–779. [Google Scholar] [CrossRef]
- Dreyfuss, G.; Vignoles, P.; Rondelaud, D. Fasciola hepatica: Epidemiological surveillance of natural watercress beds in central France. Parasitol. Res. 2005, 95, 278–282. [Google Scholar] [CrossRef]
- Gaasenbeek, C.P.H.; Over, H.J.; Noorman, N.; Leeuw, W.A.; An, W.L. An epidemiological study of Fasciola hepatica in the Netherlands. Vet. Q. 2011, 2176, 140–144. [Google Scholar] [CrossRef]
- Ueno, H. Metacercaria detecting buoy (MDB) method for estimating contamination of rice fields with Fasciola metacercariae. Jarq 1976, 10, 149–152. [Google Scholar]
- El-Sayad, M.; Allam, A.; Osman, M. Prevention of human fascioliasis: A study on the role of acids detergents and potassium permenganate in clearing salads from metacercariae. J. Egyptical. Soc. Parasitol. 1997, 27, 163–169. [Google Scholar]
- Hassan, A.; Shoukary, N.; El-Motayam, M.; Morsy, A. Efficacy of five chemicals on Fasciola gigantica encysted metacercariae infectivity. J. Egyptical. Soc. Parasitol. 2008, 38, 919–928. [Google Scholar]
- Fennell, J. Potential for Watercress Production in AUSTRALIA. Rural Industries Research and Development Corporation, Australian Government, Report; RIRDC Publication No. 06/105, Project No. DAS-49A (Issue 06); RIRDC Publication: Canberra, Australia.
- King, S.; Scholz, T. Trematodes of the family Opisthorchiidae: A minireview. Korean J. Parasitol. 2001, 39, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Bürli, C.; Harbrecht, H.; Odermatt, P.; Sayasone, S.; Chitnis, N. Mathematical analysis of the transmission dynamics of the liver fluke, Opisthorchis viverrini. J. Theoretical. Biol. 2018, 439, 181–194. [Google Scholar] [CrossRef] [Green Version]
- Sripa, B.; Kaewkes, S.; Sithithaworn, P.; Mairiang, E.; Laha, T.; Smout, M.; Pairojkul, C.; Bhudhisawasdi, V.; Tesana, S.; Thinkamrop, B.; et al. Liver fluke induces cholangiocarcinoma. PLoS Med. 2007, 4, 1148–1155. [Google Scholar] [CrossRef]
- Sithithaworn, P.; Andrews, R.H.; Van De, N.; Wongsaroj, T.; Sinuon, M.; Odermatt, P.; Nawa, Y.; Liang, S.; Brindley, P.J.; Sripa, B. The current status of opisthorchiasis and clonorchiasis in the Mekong Basin. Parasitol. Int. 2012, 61, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Kaewkes, S. Taxonomy and biology of liver flukes. Acta Trop. 2003, 88, 177–186. [Google Scholar] [CrossRef]
- Na, B.K.; Pak, J.H.; Hong, S.J. Clonorchis sinensis and clonorchiasis. Acta Trop. 2020, 203, 105309. [Google Scholar] [CrossRef]
- Sripa, B.; Tangkawattana, S.; Brindley, P.J. Update on Pathogenesis of Opisthorchiasis and Cholangiocarcinoma. In Advances in Parasitology; Academic Press: Cambridge, MA, USA, 2018; Volume 12. [Google Scholar] [CrossRef]
- Woradet, S.; Promthet, S.; Songserm, N.; Parkin, D.M. Factors affecting survival time of cholangiocarcinoma patients: A prospective study in Northeast Thailand. Asian Pacific. J. Cancer Prev. 2013, 14, 1623–1627. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, M.J.; Pakharukova, M.Y.; Laha, T.; Sripa, B.; Maksimova, G.A.; Rinaldi, G.; Brindley, P.J.; Mordvinov, V.A.; Amaro, T.; Santos, L.L.; et al. Infection with Opisthorchis felineus induces intraepithelial neoplasia of the biliary tract in a rodent model. Carcinogenesis 2017, 38, 929–937. [Google Scholar] [CrossRef] [Green Version]
- Pakharukova, M.Y.; Mordvinov, V.A. The liver fluke Opisthorchis felineus: Biology, epidemiology and carcinogenic potential. Trans. Royal Soc. Trop. Med. Hyg. 2015, 110, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Accelerating Work to Overcome the Global Impact of Neglected Tropical Diseases—A Roadmap for Implementation; WHO: Geneva, Switzerland, 2012. [Google Scholar]
- Chai, J.; Jung, B.K. General overview of the current status of human foodborne trematodiasis. Parasitol 2022, 149, 1262–1285. [Google Scholar] [CrossRef] [PubMed]
- Murrell, K.; Pozio, E. The liver flukes: Clonorchis sinensis, Opisthorchis spp., and Metorchis spp. In Global Water Pathogen Project. Part Three. Specific Excreted Pathogens: Environmental and Epidemiology Aspects; Michigan State University: East Lansing, MI, USA, 2017; pp. 3–17. [Google Scholar]
- Rim, H. The current pathobiology and chemotherapy of clonorchiasis. Korean J. Parasitol. 1986, 24, 1–141. [Google Scholar] [CrossRef] [PubMed]
- Manivong, K.; Komalamisra, C.; Waikagul, J.; Radomyos, P. Opisthorchis viverrini Metacercariae in Cyprinoid fish from three rivers in Khammouane Province, Lao PDR. J. Trop. Med. Parasitol. 2009, 32, 23–29. [Google Scholar] [CrossRef]
- Araki, H.; Ong, K.I.C.; Lorphachan, L.; Soundala, P.; Iwagami, M.; Shibanuma, A.; Hongvanthong, B.; Brey, P.T.; Kano, S.; Jimba, M. Mothers’ Opisthorchis viverrini infection status and raw fish dish consumption in Lao People’s Democratic Republic: Determinants of child infection status. Trop. Med. Health 2018, 46, 1–7. [Google Scholar] [CrossRef]
- Nguyen, T.T.B.; Dermauw, V.; Dahma, H.; Bui, D.T.; Le, T.T.H.; Phi, N.T.T.; Lempereur, L.; Losson, B.; Vandenberg, O.; Do, D.T.; et al. Prevalence and risk factors associated with Clonorchis sinensis infections in rural communities in northern Vietnam. PLoS Negl. Trop. Dis. 2020, 14, e0008483. [Google Scholar] [CrossRef]
- Zhang, Y.; Gong, Q.L.; Lv, Q.B.; Qiu, Y.Y.; Wang, Y.C.; Qiu, H.Y.; Guo, X.R.; Gao, J.F.; Chang, Q.C.; Wang, C.R. Prevalence of Clonorchis sinensis infection in fish in South-East Asia: A systematic review and meta-analysis. J. Fish Dis. 2020, 43, 1409–1418. [Google Scholar] [CrossRef]
- Sun, J.; Xin, H.; Jiang, Z.; Qian, M.; Duan, K.; Li, W.; Huang, S.; Gan, X.; Yang, Y.; Li, Z. High endemicity of Clonorchis sinensis infection in Binyang County, Southern China. PLoS Negl. Trop. Dis. 2020, 14, e0008540. [Google Scholar] [CrossRef]
- Le, T.H.; Van De, N.; Blair, D.; Sithithaworn, P.; McManus, D.P. Clonorchis sinensis and Opisthorchis viverrini: Development of a mitochondrial-based multiplex PCR for their identification and discrimination. Exp. Parasitol. 2006, 112, 109–114. [Google Scholar] [CrossRef]
- Cai, X.Q.; Xu, M.; Wang, W.H.; Qiu, D.Y.; Liu, G.X.; Lin, A.; Tang, J.D.; Zhang, R.L.; Zhu, X.Q. Sensitive and rapid detection of Clonorchis sinensis infection in fish by loop-mediated isothermal amplification (LAMP). Parasitol. Res. 2010, 106, 1379–1383. [Google Scholar] [CrossRef]
- Lee, S.E.; Shin, H.E.; Lee, M.R.; Kim, Y.H.; Cho, S.H.; Ju, J.W. Risk factors of clonorchis sinensis human infections in endemic areas, Haman-gun, Republic of Korea: A case-control study. Korean J. Parasitol. 2020, 58, 647–652. [Google Scholar] [CrossRef]
- Qian, M.B.; Li, H.M.; Jiang, Z.H.; Yang, Y.C.; Lu, M.F.; Wei, K.; Wei, S.L.; Chen, Y.; Zhou, C.H.; Chen, Y.D.; et al. Severe hepatobiliary morbidity is associated with clonorchis sinensis infection: The evidence from a cross-sectional community study. PLoS Negl. Trop. Dis. 1 2021, 5, e0009116. [Google Scholar] [CrossRef]
- Steele, J.A.; Richter, C.H.; Echaubard, P.; Saenna, P.; Stout, V.; Sithithaworn, P.; Wilcox, B.A. Thinking beyond Opisthorchis viverrini for risk of cholangiocarcinoma in the lower Mekong region: A systematic review and meta-analysis. Inf. Dis. Poverty 2018, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Qian, M.B.; Utzinger, J.; Keiser, J.; Zhou, X.N. Clonorchiasis. Lancet 2016, 387, 800–810. [Google Scholar] [CrossRef]
- Wang, X.; Chen, W.; Tian, Y.; Mao, Q.; Lv, X.; Shang, M.; Li, X.; Yu, X.; Huang, Y. Surface display of Clonorchis sinensis enolase on Bacillus subtilis spores potentializes an oral vaccine candidate. Vaccine 2014, 32, 1338–1345. [Google Scholar] [CrossRef]
- Huyen, T.; Thi, D.B.; Trung, D.D.; Phi, N.; Thuy, T. Knowledge, attitude and practices related to small liver fluke infection in rural communities of Ha Trung District, Thanh Hoa Province, Vietnam. J. Kasetsart Vet. 2021, 31, 15–32. [Google Scholar]
- Sripa, B.; Tangkawattana, S.; Laha, T.; Kaewkes, S.; Mallory, F.F.; Smith, J.F.; Wilcox, B.A. Toward integrated opistorchiasis control in Northeast Thailand: The Lawa Project. Acta Trop. 2015, 141, 361–367. [Google Scholar] [CrossRef]
- Sripa, B.; Tangkawattana, S.; Sangnikul, T. The Lawa model: A sustainable, integrated opisthorchiasis control program using the EcoHealth approach in the Lawa Lake region of Thailand. Parasitol. Int. 2017, 66, 346–354. [Google Scholar] [CrossRef]
- Aibinu, I.E.; Smooker, P.M.; Lopata, A.L. Anisakis Nematodes in Fish and Shellfish- from infection to allergies. Int. J. Parasitol. Parasites Wildl. 2019, 6, 384–393. [Google Scholar] [CrossRef]
- Baird, F.J.; Gasser, R.B.; Jabbar, A.; Lopata, A.L. Foodborne anisakiasis and allergy. Mol. Cell Probes. 2014, 28, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Mercken, E.; Van Damme, I.; Vangeenberghe, S.; Serradell, A.; De Sterck, T.; Lumain, J.P.L.; Gabriël, S. Ascaridoids in commercial fish: Occurrence, intensity and localization in whole fish and fillets destined for the Belgian market. Int. J. Food Microbiol. 2020, 16, 108657. [Google Scholar] [CrossRef] [PubMed]
- Adroher-Auroux, F.J.; Benítez-Rodríguez, R. Anisakiasis and Anisakis: An underdiagnosed emerging disease and its main etiological agents. Res. Vet. Sci. 2020, 132, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, P.; Acerra, V.; Bellisario, B.; Sbaraglia, G.L.; Cheleschi, R.; Nascetti, G.; Mattiucci, S. Larval migration of the zoonotic parasite Anisakis pegreffii (Nematoda: Anisakidae) in European anchovy, Engraulis encrasicolus: Implications to seafood safety. Food Control 2016, 519, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Bucci, C.; Gallotta, S.; Morra, I.; Fortunato, A.; Ciacci, C.; Iovino, P. Anisakis, just think about it in an emergency! Int J Infect Dis 2013, 17, e1071–e1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieuwenhuizen, N.E. Anisakis-immunology of a foodborne parasitosis. Parasite Immunol. 2016, 38, 548–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzucco, W.; Raia, D.D.; Marotta, C.; Costa, A.; Ferrantelli, V.; Vitale, F.; Casuccio, A. Anisakis sensitization in different population groups and public health impact: A systematic review. PLoS ONE 2018, 20, e0203671. [Google Scholar] [CrossRef] [PubMed]
- Rahmati, A.R.; Kiani, B.; Afshari, A.; Moghaddas, E.; Williams, M.; Shamsi, S. World-wide prevalence of Anisakis larvae in fish and its relationship to human allergic anisakiasis: A systematic review. Parasitol. Res. 2020, 119, 3585–3594. [Google Scholar] [CrossRef]
- Añíbarro, B.; Seoane, F.J.; Múgica, M.V. Involvement of hidden allergens in food allergic reactions. J. Investig. Allergol. Clin. Immunol. 2007, 17, 168–172. [Google Scholar]
- Armentia, A.; Martin-Gil, F.J.; Pascual, C.; Martín-Esteban, M.; Callejo, A.; Martínez, C. Anisakis simplex allergy after eating chicken meat. J. Investig. Allergol. Clin. Immunol. 2006, 16, 258–263. [Google Scholar]
- Fæste, C.; Levsen, A.; Lin, A.H.; Larsen, N.; Plassen, C.; Moen, A.; Van Do, T.; Egaas, E. Fish feed as source of potentially allergenic peptides from the fish parasite Anisakis simplex (s.l.). Anim. Feed Sci. Technol. 2015, 202, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Polimeno, L.; Lisanti, M.T.; Rossini, M.; Giacovazzo, E.; Polimeno, L.; Debellis, L.; Ballini, A.; Topi, S.; Santacroce, L. Anisakis Allergy: Is aquacultured fish a safe and alternative food to wild-capture fisheries for Anisakis simplex-sensitized patients? Biology 2021, 10, 106. [Google Scholar] [CrossRef]
- Saelens, G.; Planckaert, S.; Devreese, B.; Gabriël, S. Transmissibility of anisakid allergenic peptides from animal feed to chicken meat: Proof of concept. J. Food Comp. Anal. 2023, 115, 104939. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.E.; Lopata, A.L. Anisakis-A food-borne parasite that triggers allergic host defences. Int. J. Parasitol. 2013, 43, 1047–1057. [Google Scholar] [CrossRef]
- Ivanović, J.; Baltić, M.Ž.; Bošković, M.; Kilibarda, N.; Dokmanović, M.; Marković, R.; Janjić, J.; Baltić, B. Anisakis allergy in human. Trends Food Sci. Technol. 2017, 59, 25–29. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.E.; Lopata, A. Allergic reactions to Anisakis found in fish. Curr. Allergy Asthma Rep. 2014, 14, 455. [Google Scholar] [CrossRef]
- Levsen, A.; Lunestad, B.T.; Berland, B. Low detection dfficiency of candling as a commonly recommended inspection method for nematode larvae in the flesh of Pelagic fish. J. Food Protect 2005, 68, 828–832. [Google Scholar] [CrossRef]
- Karl, H.; Leinemann, M. A fast and quantitative detection method for nematodes in fish fillets and fishery products. Arch. Lebensmittelhyg. 1993, 44, 124–125. [Google Scholar]
- Petrie, A.; Wootten, R.; Bruno, D.; MacKenzie, K.; Bron, J. A Survey of Anisakis and Pseudoterranova in Scottish Fisheries and the Efficacy of Current Detection Methods FSAS Project S14008; Food Standards Scotland: Aberdeen, UK, 2007. [Google Scholar]
- Mercken, E.; Van Damme, I.; Šoba Šparl, B.; Vangeenberghe, S.; Serradell, A.; De Sterck, T. Sensitivity of candling as routine method for the detection and recovery of ascaridoids in commercial fish fillets. Sci. Rep. 2022, 12, 1358. [Google Scholar] [CrossRef]
- Pippy, J. Use of ultraviolet light to find parasitic nematodes in situ. J. Fish Res. Board. Can. 1970, 27, 963–965. [Google Scholar] [CrossRef]
- Llarena-Reino, M.; Piñeiro, C.; Antonio, J.; Outeriño, L.; Vello, C.; González, Á.F.; Pascual, S. Optimization of the pepsin digestion method for anisakids inspection in the fishing industry. Vet. Parasitol. 2013, 191, 276–283. [Google Scholar] [CrossRef]
- Kochanowski, M.; Różycki, M.; Dąbrowska, J.; Karamon, J.; Sroka, J.; Antolak, E.; Bełcik, A.; Cencek, T. Development and application of novel chemiluminescence immunoassays for highly sensitive detection of Anisakis simplex proteins in thermally processed seafood. Pathogens 2020, 23, 777. [Google Scholar] [CrossRef] [PubMed]
- Werner, M.T.; Fæste, C.K.; Levsen, A.; Egaas, E. A quantitative sandwich ELISA for the detection of Anisakis simplex protein in seafood. Eur. Food Res. Technol. 2011, 232, 967–973. [Google Scholar] [CrossRef] [Green Version]
- Fæste, C.K.; Moen, A.; Schniedewind, B.; Haug Anonsen, J.; Klawitter, J.; Christians, U. Development of liquid chromatography-tandem mass spectrometry methods for the quantitation of Anisakis simplex proteins in fish. J. Chromatogr. A 2016, 5, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Mahillo, A.I.; González-Muñoz, M.; de las Heras, C.; Tejada, M.; Moneo, I. Quantification of Anisakis simplex allergens in fresh, long-term frozen, and cooked fish muscle. Foodborne Pathog. Dis. 2010, 7, 967–973. [Google Scholar] [CrossRef] [Green Version]
- McClelland, G. Larval anisakine nematodes in various fish species from Sable Island Bank and vicinity. Can. Bull Fish Aquat. Sci. 1990, 222, 83–118. [Google Scholar]
- EU. Commission regulation no 1276/2011 of 8 December 2011 amending annex III to regulation (EC) no 853/2004 of the European Parliament and of the council as regards the treatment to kill viable parasites in fishery products for human consumption. Off. J. Eur. Union 2011, L 327, 39–41. [Google Scholar]
- FDA. Parasites. In Fish and Fishery Products Hazards and Controls Guidance, 4th ed.; Food and Drug Administration, U.S. Department of Health and Human Services: Silver Spring, MD, USA, 2019; pp. 91–98. [Google Scholar]
- Chen, H.Y.; Cheng, Y.S.; Grabner, D.S.; Chang, S.H.; Shih, H.H. Effect of different temperatures on the expression of the newly characterized heat shock protein 90 (Hsp90) in L3 of Anisakis spp. isolated from Scomber australasicus. Vet. Parasitol. 2014, 205, 540–550. [Google Scholar] [CrossRef]
- Donadeu, M.; Fahrion, A.S.; Olliaro, P.L.; Abela-Ridder, B. Target product profiles for the diagnosis of Taenia solium taeniasis, neurocysticercosis and porcine cysticercosis. PLoS Negl. Trop. Dis. 2017, 11, e0005875. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues da Costa, M.; Pessoa, J.; Meemken, D.; Nesbakken, T. A systematic review on the effectiveness of pre-harvest meat safety interventions in pig herds to control Salmonella and other foodborne pathogens. Microorganisms 2021, 9, 1825. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gabriël, S.; Dorny, P.; Saelens, G.; Dermauw, V. Foodborne Parasites and Their Complex Life Cycles Challenging Food Safety in Different Food Chains. Foods 2023, 12, 142. https://doi.org/10.3390/foods12010142
Gabriël S, Dorny P, Saelens G, Dermauw V. Foodborne Parasites and Their Complex Life Cycles Challenging Food Safety in Different Food Chains. Foods. 2023; 12(1):142. https://doi.org/10.3390/foods12010142
Chicago/Turabian StyleGabriël, Sarah, Pierre Dorny, Ganna Saelens, and Veronique Dermauw. 2023. "Foodborne Parasites and Their Complex Life Cycles Challenging Food Safety in Different Food Chains" Foods 12, no. 1: 142. https://doi.org/10.3390/foods12010142