
Application of Two-Dimensional Fluorescence Spectroscopy for the On-Line Monitoring of Teff-Based Substrate Fermentation Inoculated with Certain Probiotic Bacteria

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Starter Culture Preparation and Storage
2.3. Off-Line Measurement of Microbial Viability
2.4. Fermentation Process Conditions
2.5. Off-Line Measurement of Glucose and Lactic Acid
2.6. On-Line Measurement Using 2D-Fluorescence Spectroscopy
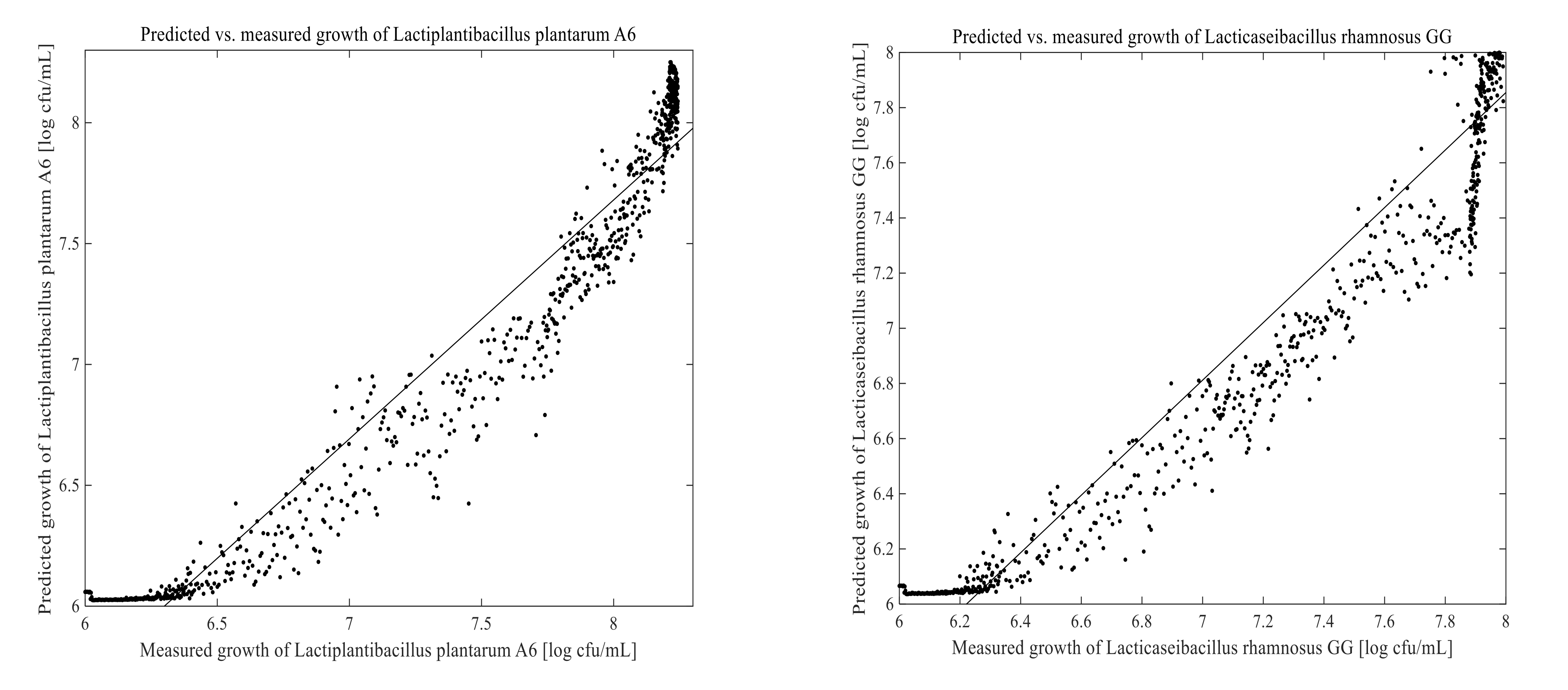
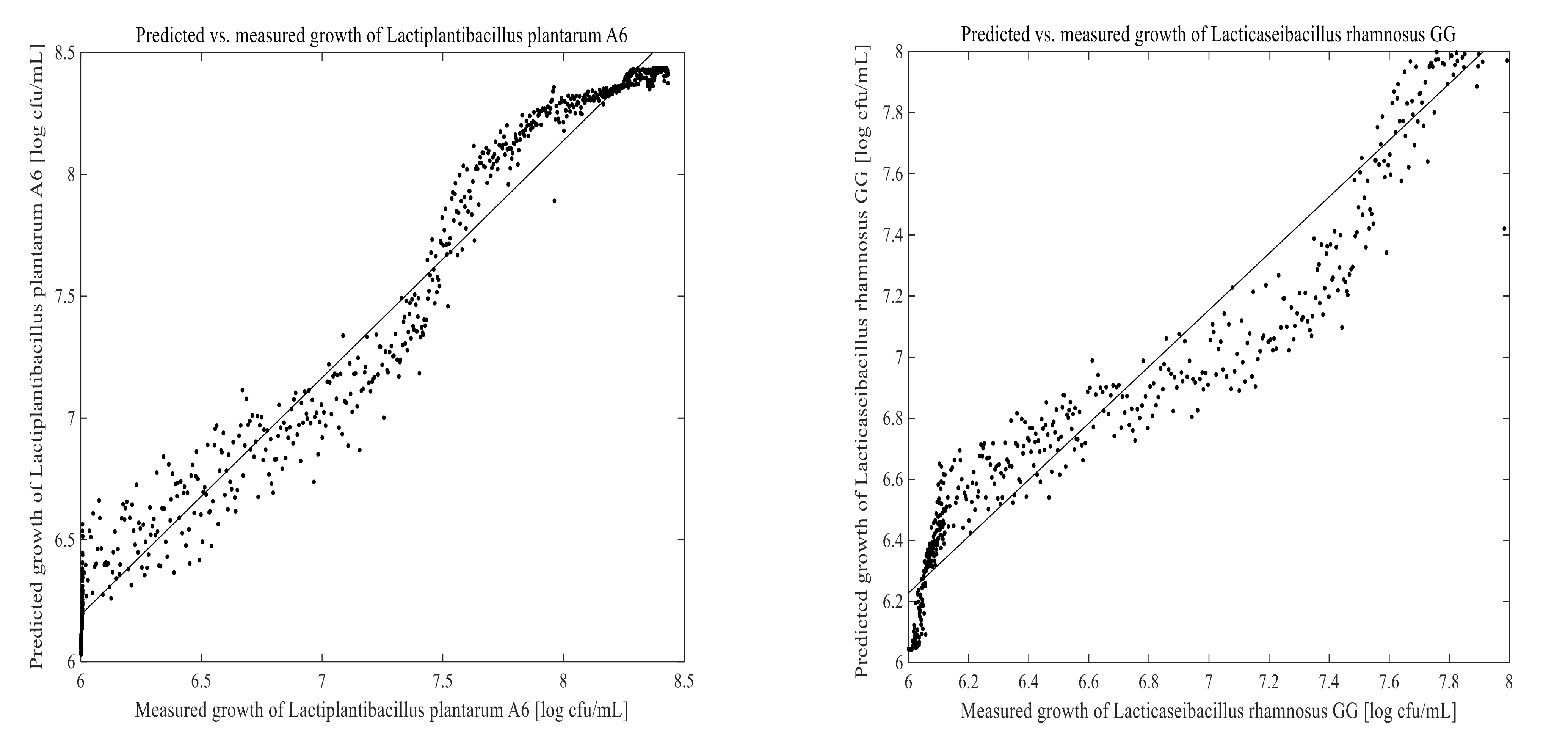
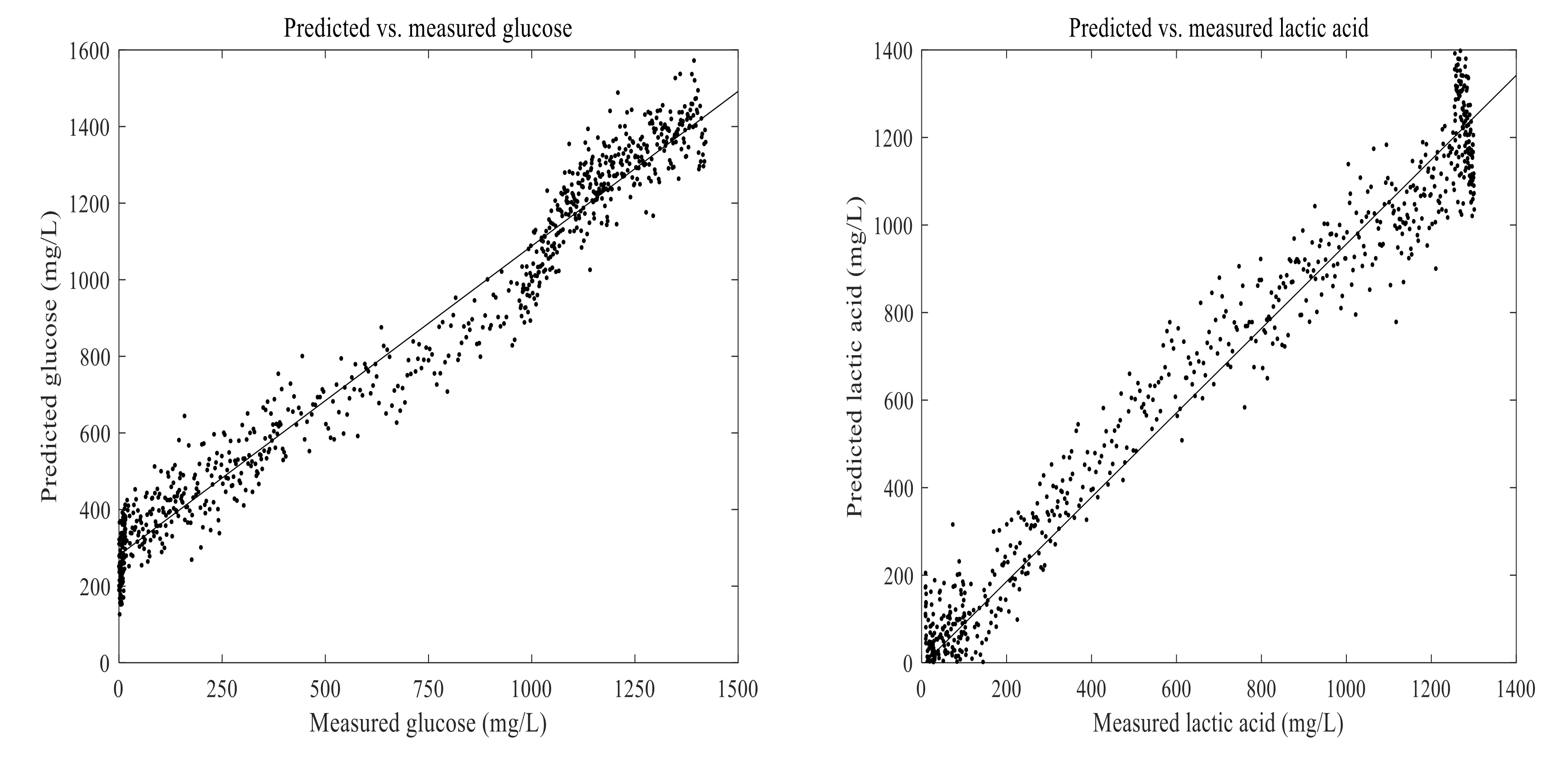
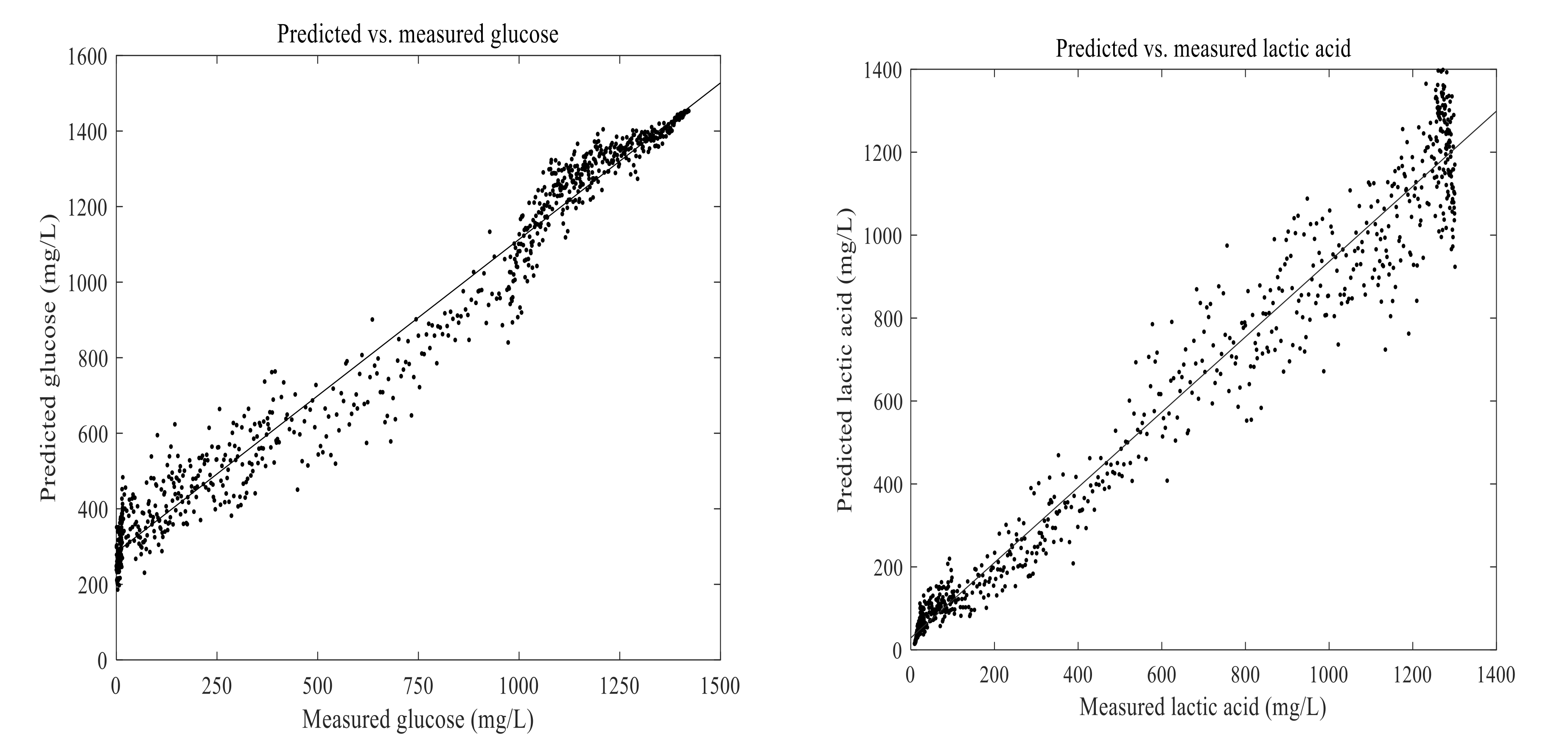
2.7. Examination of the Model Performance
2.8. Statistical Analysis
3. Results and Discussion
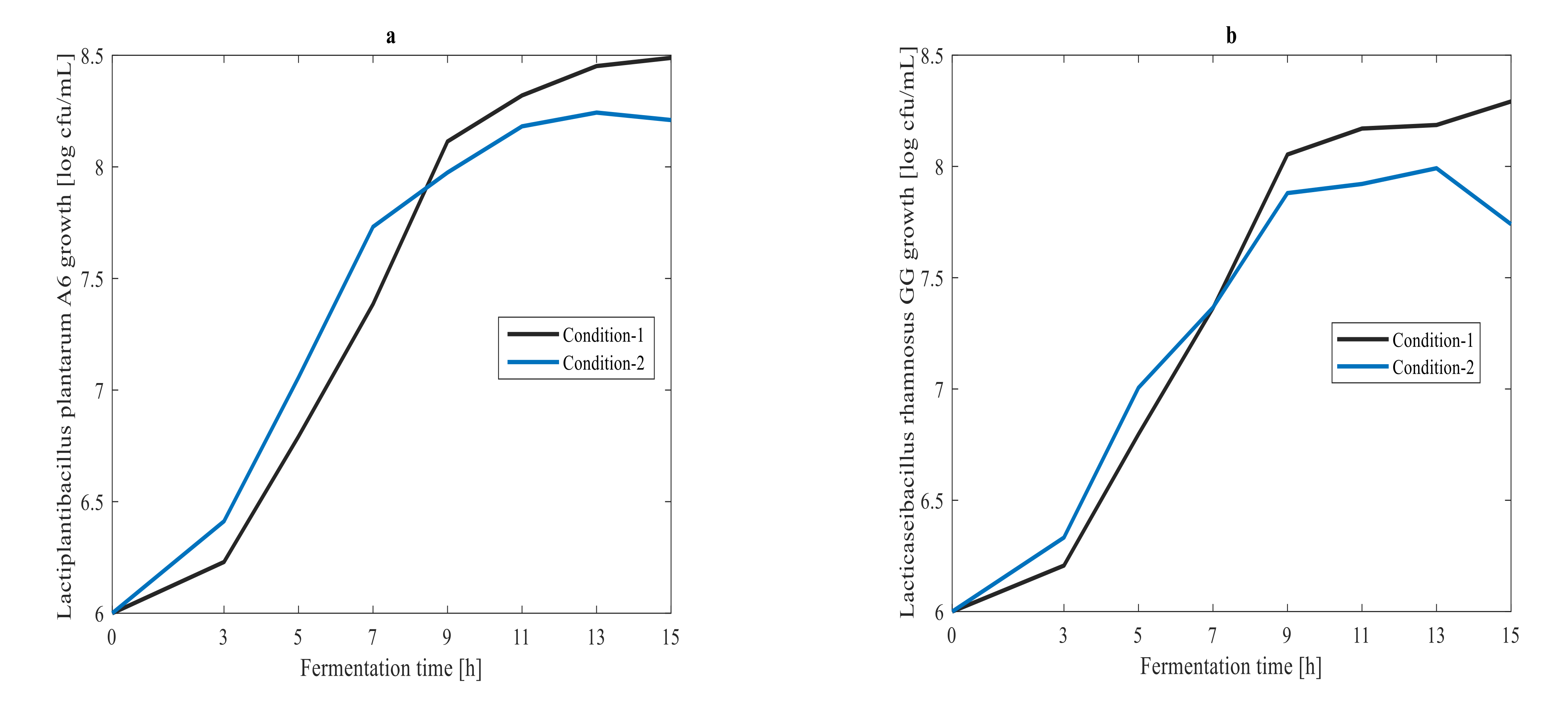
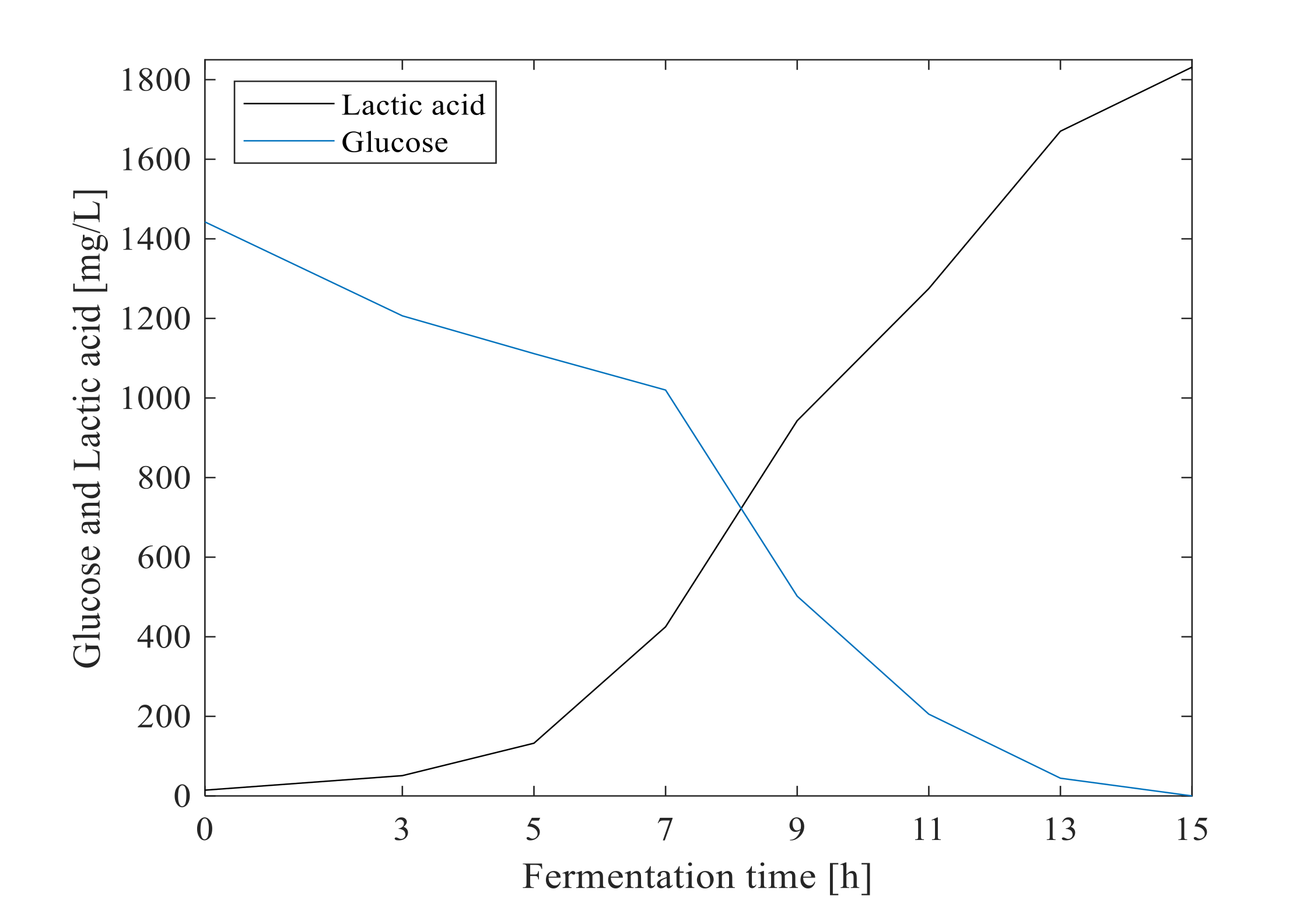
3.1. Off-Line Measurement of Cell Growth, Glucose and Lactic Acid
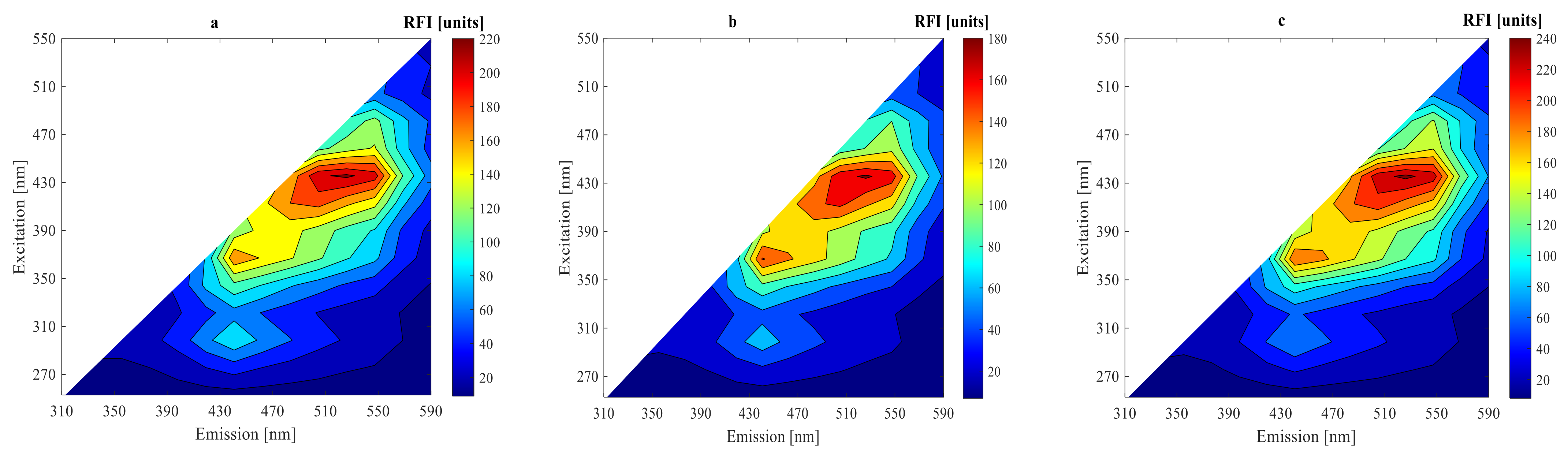
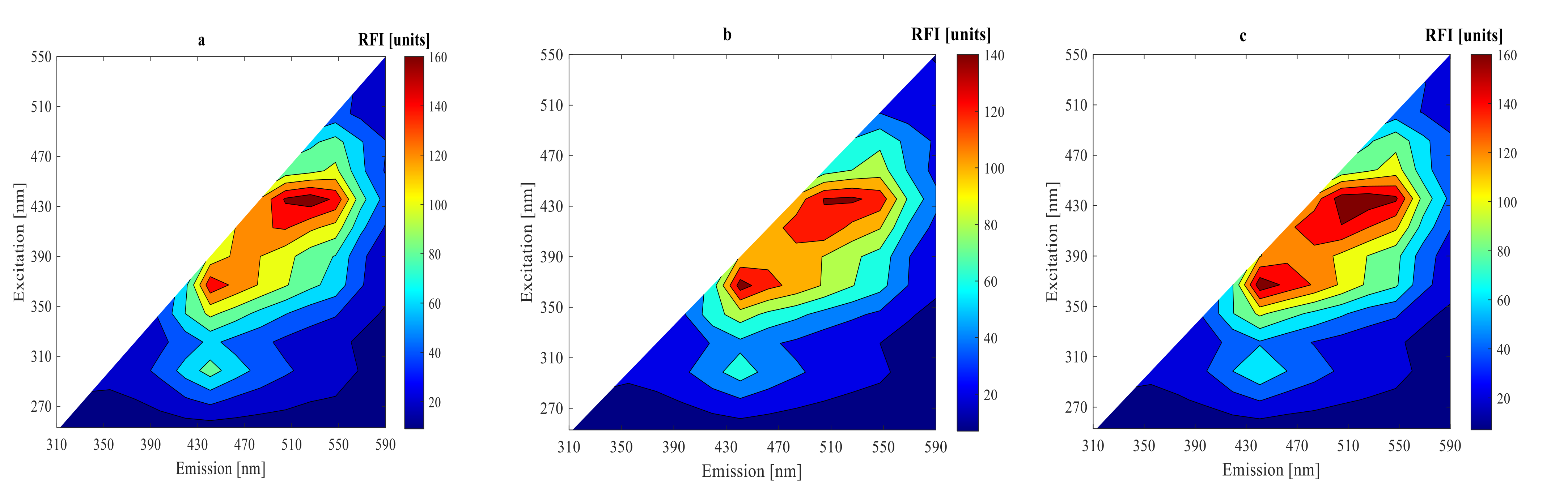
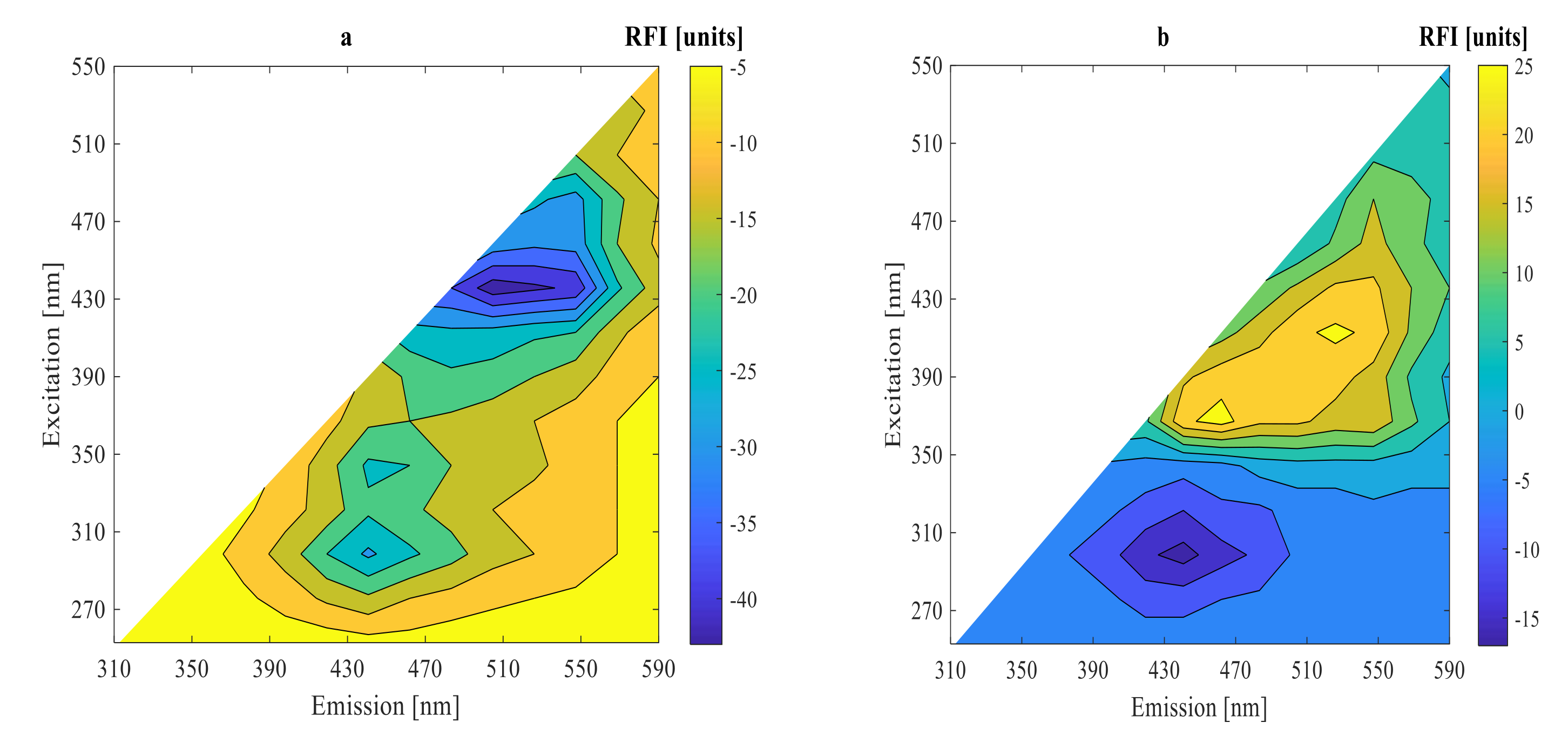
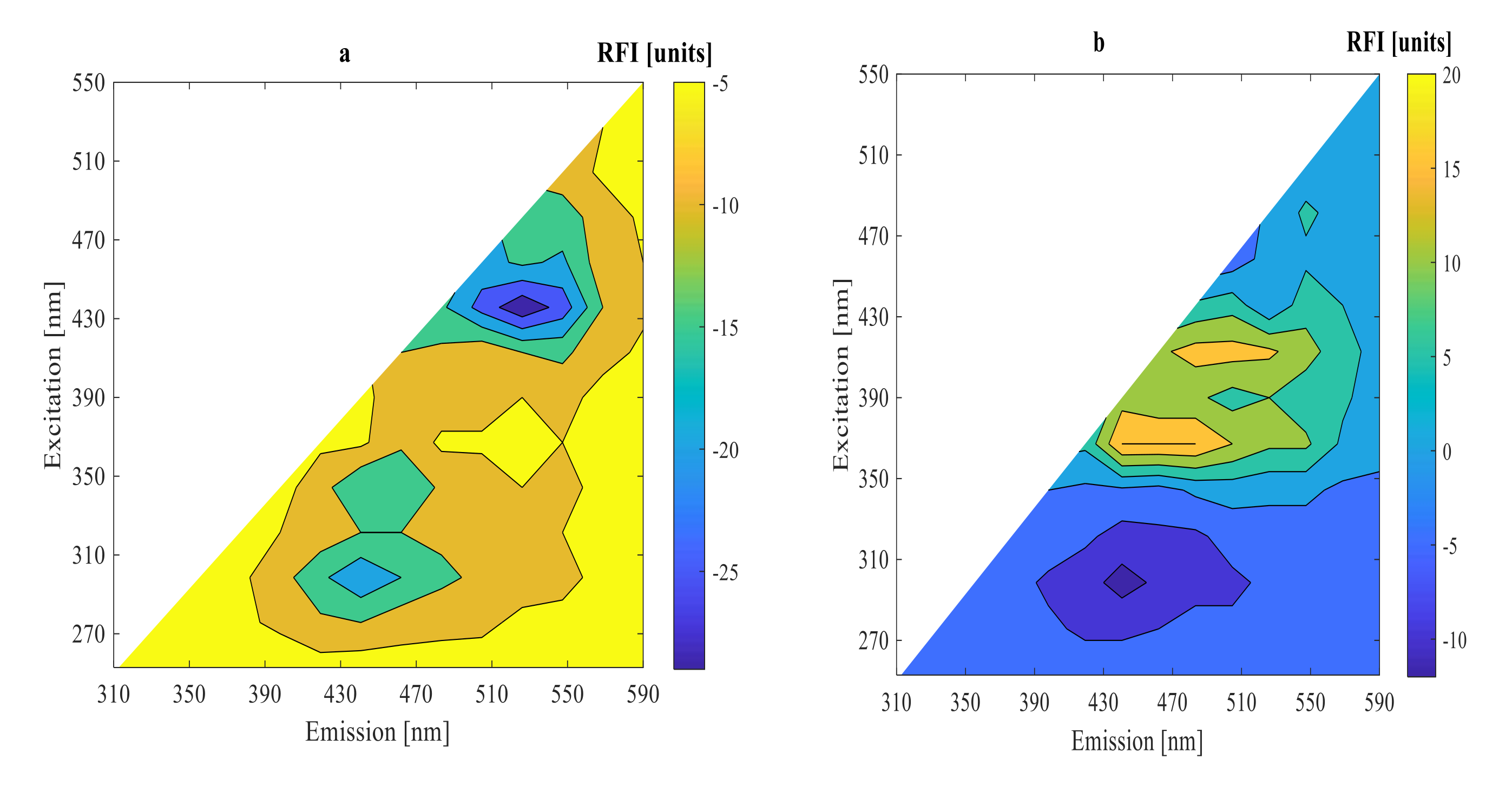
3.2. On-Line Measurement Using 2D-Fluorescence Spectroscopy
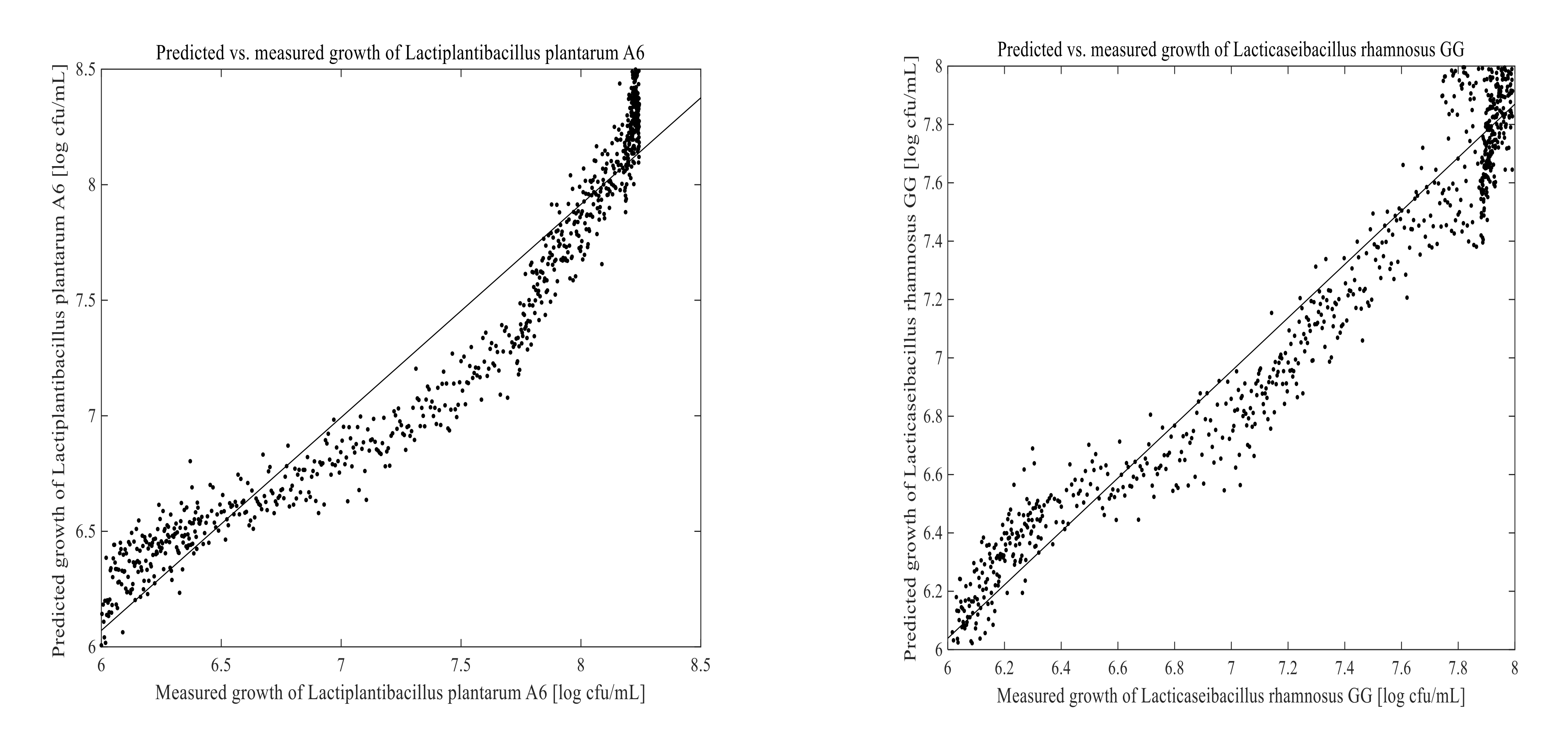
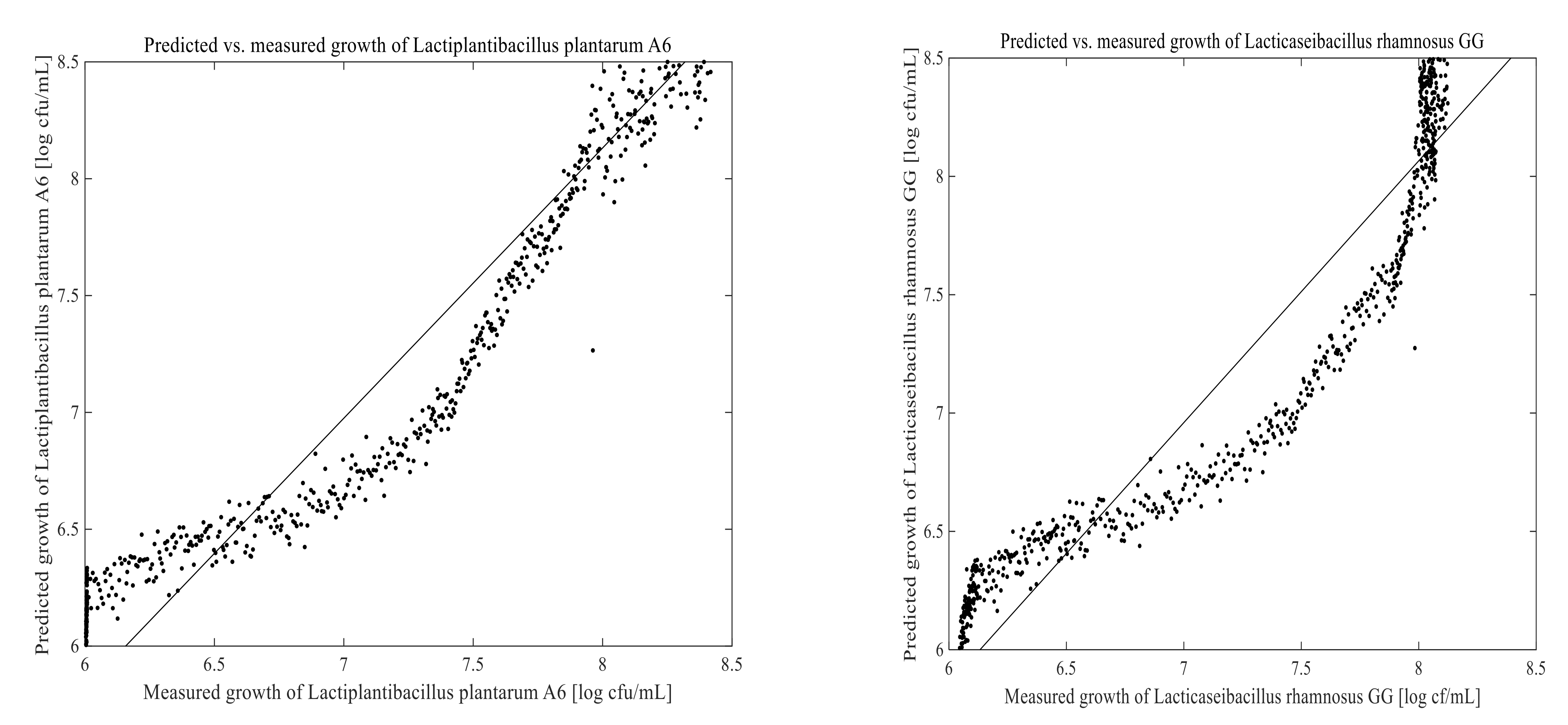
3.3. Prediction of Cell Growth in the Fermentation Process
3.4. Prediction of Lactic Acid and Glucose in the Fermentation Process
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kokwar, M.A.; Arya, S.S.; Bhat, M.S. A cereal-based nondairy probiotic functional beverage: An insight into the improvement in quality characteristics, sensory profile, and shelf-life. J. Food Processing Preserv. 2022, 46, e16147. [Google Scholar] [CrossRef]
- Li, B.; Shanahan, M.; Calvet, A.; Leister, K.J.; Ryder, A.G. Comprehensive, quantitative bioprocess productivity monitoring using fluorescence EEM spectroscopy and chemometrics. Analyst 2014, 139, 1661–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindemann, C.; Marose, S.; Nielsen, H.; Scheper, T. 2-Dimensional fluorescence spectroscopy for on-line bioprocess monitoring. Sens. Actuators Chem. 1998, 51, 273–277. [Google Scholar] [CrossRef]
- Masiero, S.S.; Trierweiler, J.O.; Farenzena, M.; Escobar, M.; Trierweiler, L.F.; Ranzan, C. Evaluation of wavelength selection methods for 2D fluorescence spectra applied to bioprocesses characterization. Braz. J. Chem. Eng. 2013, 30, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Grassi, S.; Alamprese, C.; Bono, V.; Picozzi, C.; Foschino, R.; Casiraghi, E. Monitoring of lactic acid fermentation process using Fourier transform near infrared spectroscopy. J. Near Infrared Spectrosc. 2013, 21, 417–425. [Google Scholar] [CrossRef]
- Alemneh, S.T.; Emire, S.A.; Hitzmann, B. Teff-Based Probiotic Functional Beverage Fermented with Lactobacillus rhamnosus and Lactobacillus plantarum. Foods 2021, 10, 2333. [Google Scholar] [CrossRef] [PubMed]
- Alemneh, S.T.; Emire, S.A.; Hitzmann, B.; Zettel, V. Comparative Study of Chemical Composition, Pasting, Thermal and Functional properties of Teff (Eragrostis tef) Flours Grown in Ethiopia and South Africa. Int. J. Food Prop. 2022, 25, 144–158. [Google Scholar] [CrossRef]
- Svendsen, C.; Skov, T.; Van den Berg, F.W. Monitoring fermentation processes using in-process measurements of different orders. J. Chem. Technol. Biotechnol. 2015, 90, 244–254. [Google Scholar] [CrossRef]
- Assawajaruwan, S.; Reinalter, J.; Hitzmann, B. Comparison of methods for wavelength combination selection from multi-wavelength fluorescence spectra for on-line monitoring of yeast cultivations. Anal. Bioanal. Chem. 2017, 409, 707–717. [Google Scholar] [CrossRef]
- Rhee, J.I.; Kang, T.-H. On-line process monitoring and chemometric modeling with 2D fluorescence spectra obtained in recombinant E. coli fermentations. Process Biochem. 2007, 42, 1124–1134. [Google Scholar] [CrossRef]
- Khajehsharifi, H.; Pourbasheer, E.; Tavallali, H.; Sarvi, S.; Sadeghi, M. The comparison of partial least squares and principal component regression in simultaneous spectrophotometric determination of ascorbic acid, dopamine and uric acid in real samples. Arab. J. Chem. 2017, 10, S3451–S3458. [Google Scholar] [CrossRef] [Green Version]
- Wentzell, P.D.; Montoto, L.V. Comparison of principal components regression and partial least squares regression through generic simulations of complex mixtures. Chemom. Intell. Lab. Syst. 2003, 65, 257–279. [Google Scholar] [CrossRef]
- Li, F.; Gu, Z.; Ge, L.; Sun, D.; Deng, X.; Wang, S.; Hu, B.; Xu, J. Application of artificial neural networks to X-ray fluorescence spectrum analysis. X-ray Spectrom. 2019, 48, 138–150. [Google Scholar] [CrossRef]
- Yang, J.; Du, L.; Cheng, Y.; Shi, S.; Xiang, C.; Sun, J.; Chen, B. Assessing different regression algorithms for paddy rice leaf nitrogen concentration estimations from the first-derivative fluorescence spectrum. Opt. Express 2020, 28, 18728–18741. [Google Scholar] [CrossRef]
- Bationo, F.; Songré-Ouattara, L.T.; Hemery, Y.M.; Hama-Ba, F.; Parkouda, C.; Chapron, M.; Le Merrer, M.; Leconte, N.; Sawadogo-Lingani, H.; Diawara, B. Improved processing for the production of cereal-based fermented porridge enriched in folate using selected lactic acid bacteria and a back slopping process. LWT 2019, 106, 172–178. [Google Scholar] [CrossRef]
- Matejčeková, Z.; Liptáková, D.; Valík, Ľ. Functional probiotic products based on fermented buckwheat with Lactobacillus rhamnosus. LWT-Food Sci. Technol. 2017, 81, 35–41. [Google Scholar] [CrossRef]
- Yan, Y.; Zhang, F.; Chai, Z.; Liu, M.; Battino, M.; Meng, X. Mixed fermentation of blueberry pomace with L. rhamnosus GG and L. plantarum-1: Enhance the active ingredient, antioxidant activity and health-promoting benefits. Food Chem. Toxicol. 2019, 131, 110541. [Google Scholar] [CrossRef]
- Terpou, A.; Papadaki, A.; Lappa, I.K.; Kachrimanidou, V.; Bosnea, L.A.; Kopsahelis, N. Probiotics in food systems: Significance and emerging strategies towards improved viability and delivery of enhanced beneficial value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef] [Green Version]
- Johansson, L.; Lidén, G. A study of long-term effects on plasmid-containing Escherichia coli in carbon-limited chemostat using 2D-fluorescence spectrofluorimetry. Biotechnol. Prog. 2006, 22, 1132–1139. [Google Scholar] [CrossRef]
- Yang, H.; Xiao, X.; Zhao, X.S.; Hu, L.; Xue, X.F.; Ye, J.S. Study on Fluorescence Spectra of Thiamine and Riboflavin. In Proceedings of the MATEC Web of Conferences: 2016; EDP Sciences: Les Ulis, France, 2016; p. 03013. [Google Scholar]
- Díez-Pascual, A.M.; García-García, D.; San Andrés, M.P.; Vera, S. Determination of riboflavin based on fluorescence quenching by graphene dispersions in polyethylene glycol. RSC Adv. 2016, 6, 19686–19699. [Google Scholar] [CrossRef]
- Bottiroli, G.; Croce, A.C.; Locatelli, D.; Marchesini, R.; Pignoli, E.; Tomatis, S.; Cuzzoni, C.; Di Palma, S.; Dalfante, M.; Spinelli, P. Natural fluorescence of normal and neoplastic human colon: A comprehensive “ex vivo” study. Lasers Surg. Med. 1995, 16, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Grote, B.; Zense, T.; Hitzmann, B. 2D-fluorescence and multivariate data analysis for monitoring of sourdough fermentation process. Food Control 2014, 38, 8–18. [Google Scholar] [CrossRef]
- Faassen, S.M.; Hitzmann, B. Fluorescence spectroscopy and chemometric modeling for bioprocess monitoring. Sensors 2015, 15, 10271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graf, A.; Claßen, J.; Solle, D.; Hitzmann, B.; Rebner, K.; Hoehse, M. A novel LED-based 2D-fluorescence spectroscopy system for in-line monitoring of Chinese hamster ovary cell cultivations–Part, I. Eng. Life Sci. 2019, 19, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Thakur, K.; Tomar, S.K.; De, S. Lactic acid bacteria as a cell factory for riboflavin production. Microb. Biotechnol. 2016, 9, 441–451. [Google Scholar] [CrossRef]
- Thakur, K.; Tomar, S.K. Exploring indigenous Lactobacillus species from diverse niches for riboflavin production. J. Young Pharm. 2015, 7, 126. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-Y.; Choi, E.-J.; Lee, J.-H.; Yoo, M.-S.; Heo, K.; Shim, J.-J.; Lee, J.-L. Probiotic Potential of a Novel Vitamin B2-Overproducing Lactobacillus plantarum Strain, HY7715, Isolated from Kimchi. Appl. Sci. 2021, 11, 5765. [Google Scholar] [CrossRef]
- Jain, G.; Jayaraman, G.; Kökpinar, Ö.; Rinas, U.; Hitzmann, B. On-line monitoring of recombinant bacterial cultures using multi-wavelength fluorescence spectroscopy. Biochem. Eng. J. 2011, 58, 133–139. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predicted with PLSR | Condition 1 | Condition 2 | ||||
|---|---|---|---|---|---|---|
| RMSEP (log cfu/mL) | pRMSEP (%) | R2 | RMSEP (log cfu/mL) | pRMSEP (%) | R2 | |
| LPA6 | 0.31 | 3.7 | 0.88 | 0.22 | 2.7 | 0.92 |
| LCGG | 0.32 | 3.9 | 0.85 | 0.20 | 2.4 | 0.92 |
| Predicted with ANN | RMSEP (log cfu/mL) | pRMSEP (%) | R2 | RMSEP (log cfu/mL) | pRMSEP (%) | R2 |
| LPA6 | 0.21 | 2.5 | 0.95 | 0.37 | 4.5 | 0.78 |
| LCGG | 0.20 | 2.5 | 0.94 | 0.29 | 3.6 | 0.83 |
| Analyte | PLSR | ANN | ||||
|---|---|---|---|---|---|---|
| RMSEP (log cfu/mL) | pRMSEP (%) | R2 | RMSEP (log cfu/mL) | pRMSEP (%) | R2 | |
| Glucose | 191.72 | 13.5 | 0.86 | 199.92 | 14.1 | 0.85 |
| Lactic acid | 98.41 | 7.6 | 0.96 | 100.09 | 7.7 | 0.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alemneh, S.T.; Emire, S.A.; Jekle, M.; Paquet-Durand, O.; von Wrochem, A.; Hitzmann, B. Application of Two-Dimensional Fluorescence Spectroscopy for the On-Line Monitoring of Teff-Based Substrate Fermentation Inoculated with Certain Probiotic Bacteria. Foods 2022, 11, 1171. https://doi.org/10.3390/foods11081171
Alemneh ST, Emire SA, Jekle M, Paquet-Durand O, von Wrochem A, Hitzmann B. Application of Two-Dimensional Fluorescence Spectroscopy for the On-Line Monitoring of Teff-Based Substrate Fermentation Inoculated with Certain Probiotic Bacteria. Foods. 2022; 11(8):1171. https://doi.org/10.3390/foods11081171
Chicago/Turabian StyleAlemneh, Sendeku Takele, Shimelis Admassu Emire, Mario Jekle, Olivier Paquet-Durand, Almut von Wrochem, and Bernd Hitzmann. 2022. "Application of Two-Dimensional Fluorescence Spectroscopy for the On-Line Monitoring of Teff-Based Substrate Fermentation Inoculated with Certain Probiotic Bacteria" Foods 11, no. 8: 1171. https://doi.org/10.3390/foods11081171