
Application of iTRAQ Technology to Identify Differentially Expressed Proteins of Sauce Lamb Tripe with Different Secondary Pasteurization Treatments

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Samples
2.3. Determination of Texture Profile Analysis (TPA)
2.4. Protein Extraction
2.5. Protein Digestion and Labeling of Peptides
2.6. Pre-Separation and Mass Spectrum Identification
2.7. Western Blot
2.8. Bioinformatics and Statistical Analysis
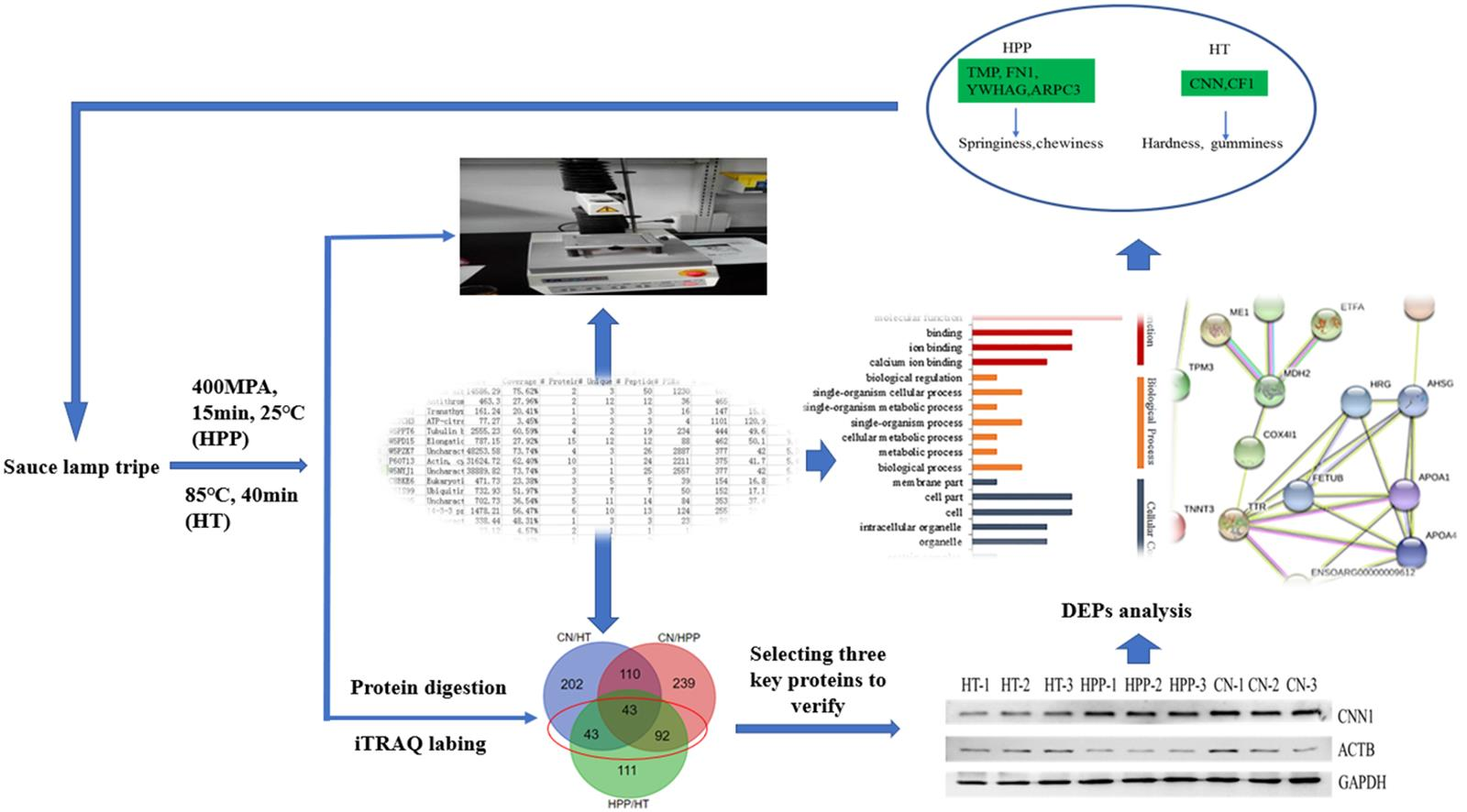
3. Results
3.1. TPA Analysis
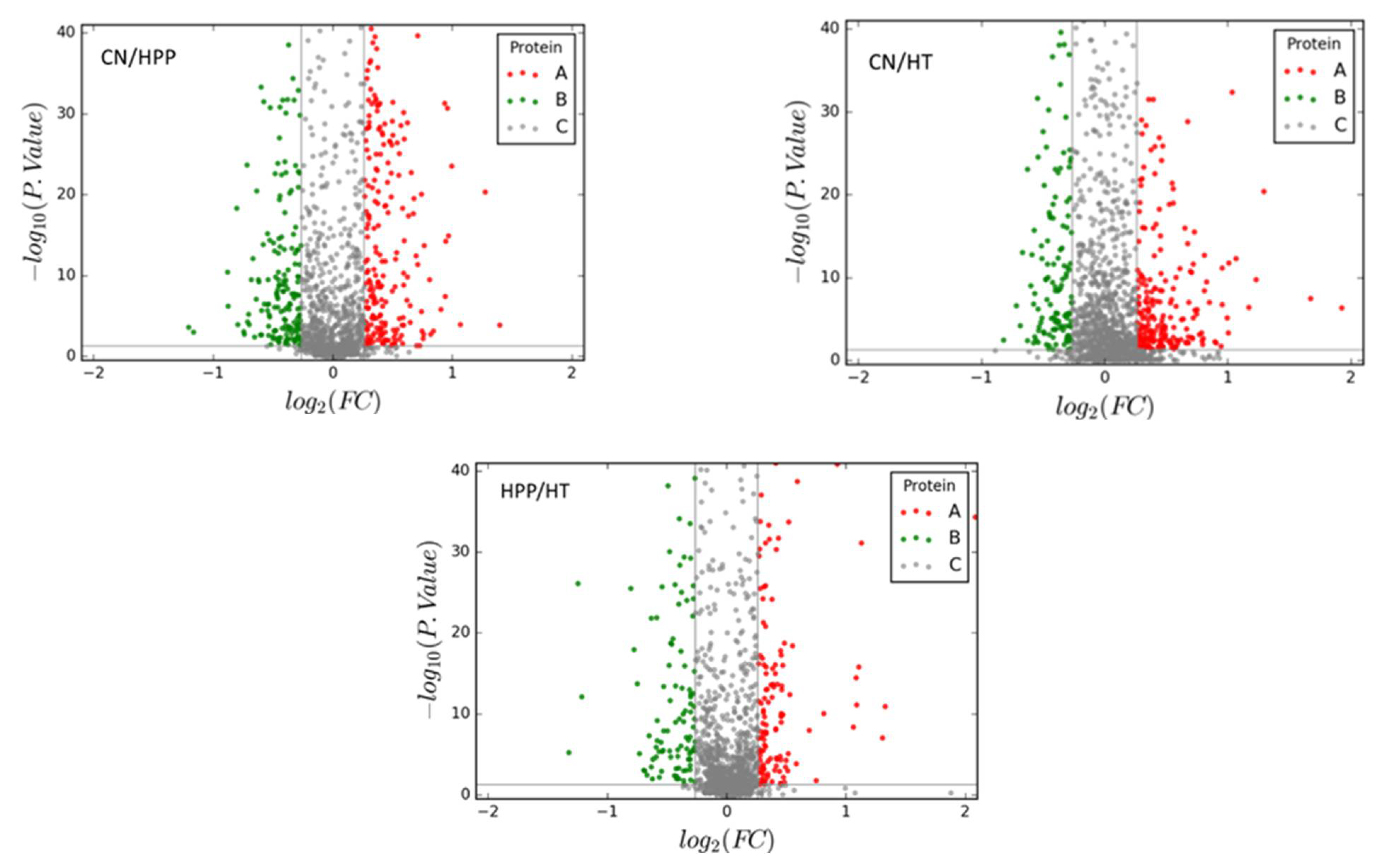
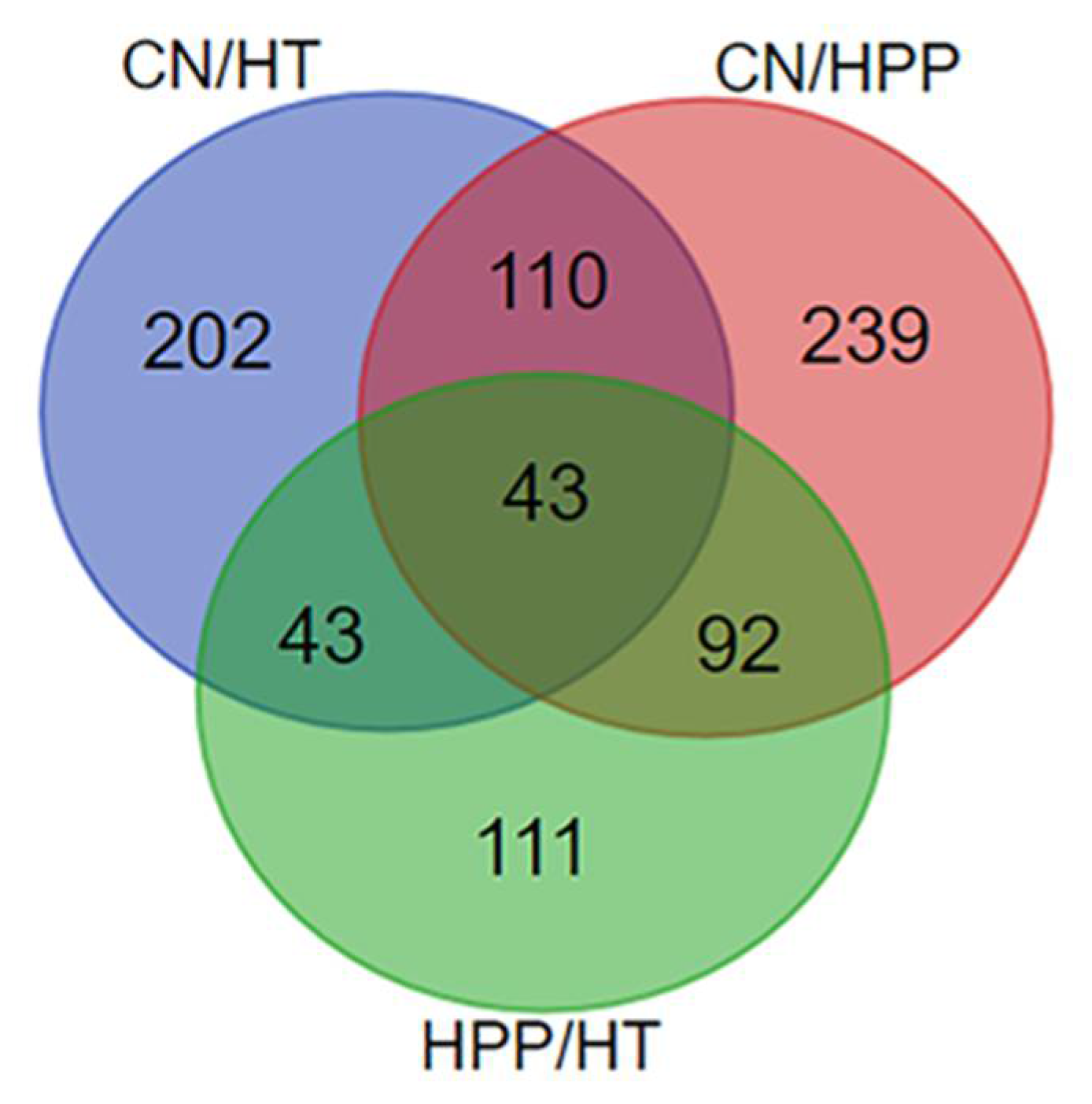
3.2. Proteomic Analysis
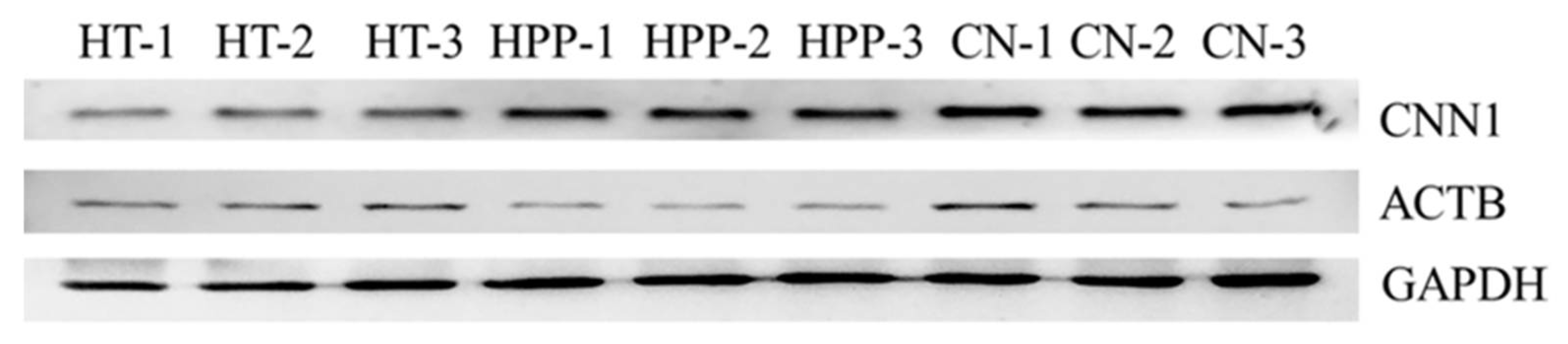
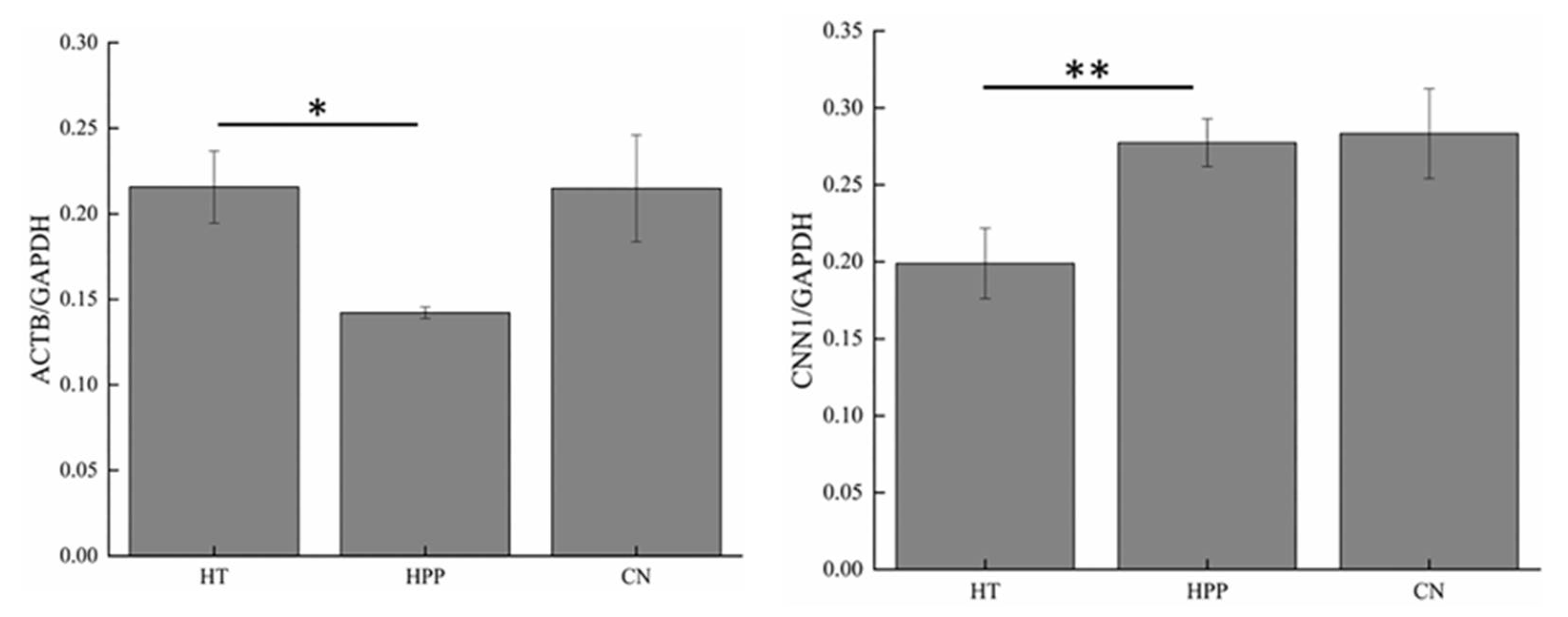
3.3. Western Blot Analysis
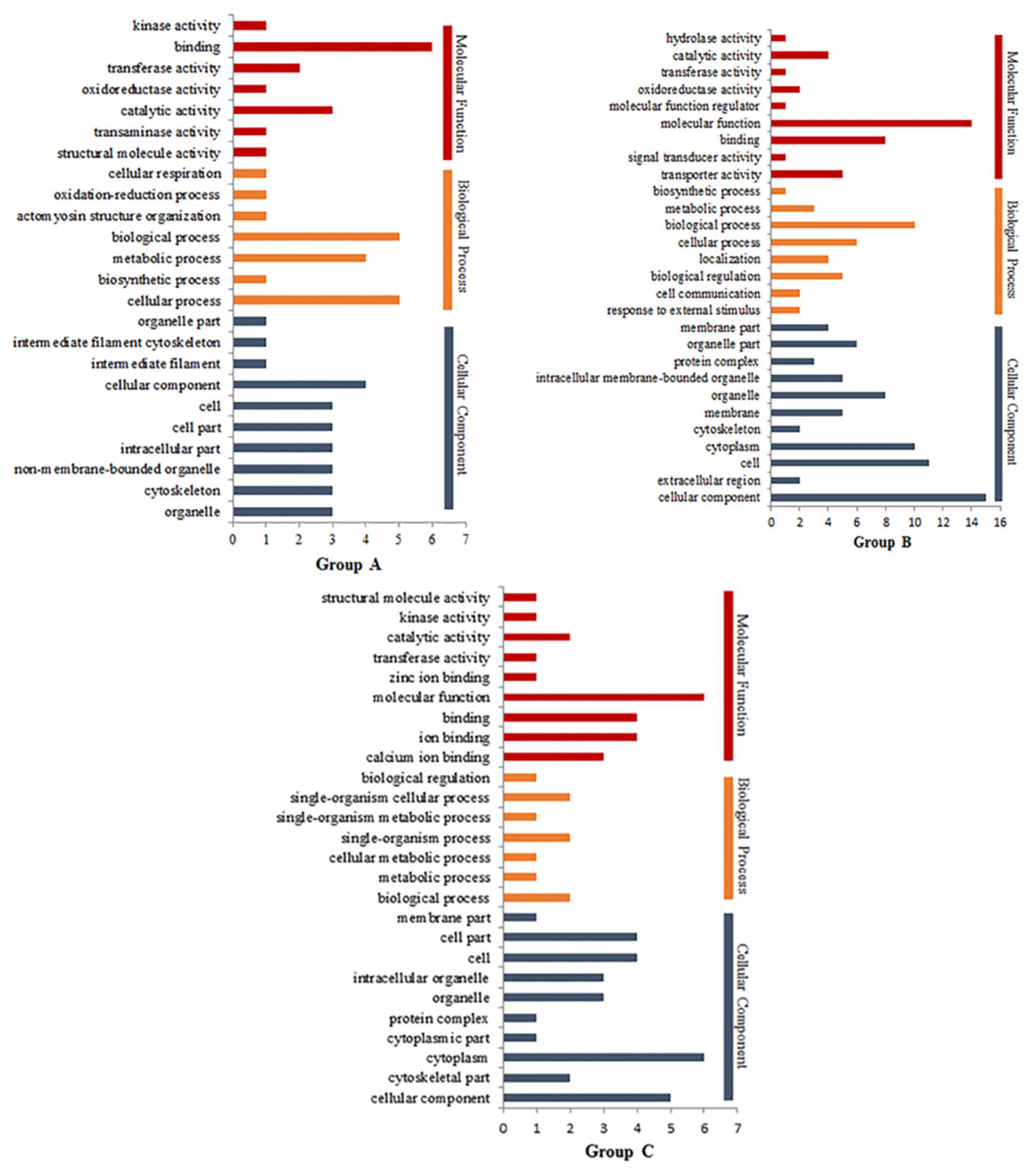
3.4. Gene Ontology Nalysis of DEPs
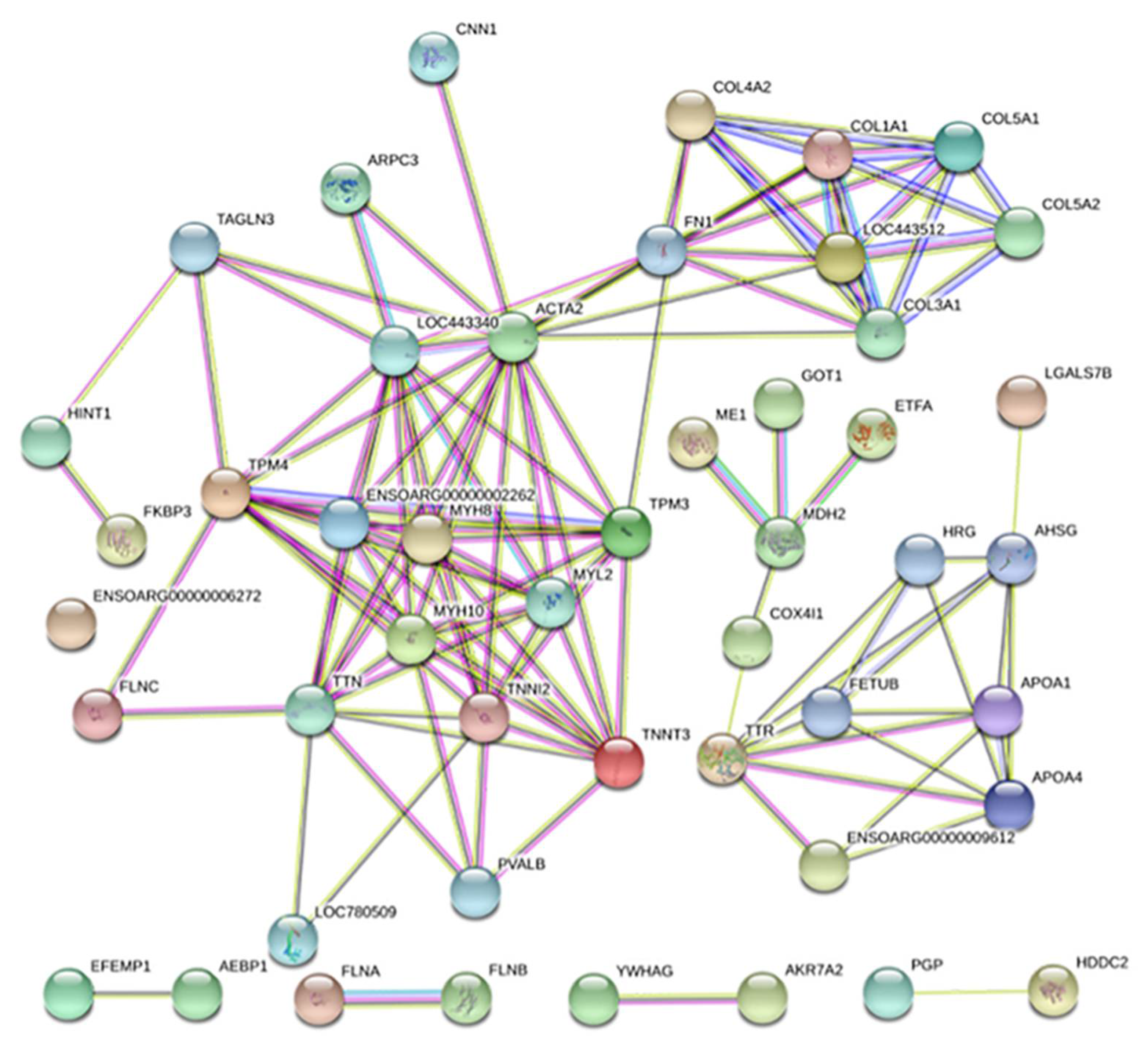
3.5. Protein–Protein Interaction Networks of DEPs
3.6. Potential Key Proteins Associated with Texture
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Number | Protein Name | Gene | Score | Difference Multiple | ||
|---|---|---|---|---|---|---|
| CN/HT | CN/HPP | HPP/HT | ||||
| Group A | ||||||
| W5PZK7 | Actin alpha 2 | ACTA2 | 48253.58 | 0.719385185 | —— | 1.231099206 |
| Q7YRL2 | Calponin-1 | CNN1 | 37163.94 | 1.637111111 | —— | 1.306886777 |
| Q9MZW1 | Beta-actin | —— | 19378.69 | 1.488666667 | —— | 1.406111111 |
| Q6J275 | Galectin | —— | 5735.86 | 0.465888889 | —— | 1.271913651 |
| W5PW05 | Malate dehydrogenase 2 | MDH2 | 4477.01 | 0.758929276 | —— | 1.367714286 |
| A0A0U1Z4T4 | Myosin light chain 3 | MYL3 | 1463.82 | 1.47012037 | —— | 0.690777778 |
| B7TJ13 | Phosphoglycerate kinase | PGK1 | 1439.86 | 0.764185185 | —— | 1.278442153 |
| W5QDF3 | Myosin heavy chain 7 | MYH7 | 983.21 | 0.757269841 | —— | 0.602759259 |
| W5QDG7 | Fibronectin 1 | FN1 | 630.48 | 0.725157088 | —— | 1.268772487 |
| W5Q611 | Keratin 1 | KRT1 | 454.6 | 1.46988889 | —— | 0.759463 |
| W5Q5N9 | Keratin 8 | KRT8 | 430.17 | 0.758 | —— | 0.77182963 |
| W5Q6G0 | IF rod domain-containing protein | LOC101111440 | 303.71 | 1.24622222 | —— | 0.594079365 |
| W5PKE2 | ETF domain-containing protein (Fragment) | —— | 280.19 | 1.33751852 | —— | 1.274928571 |
| W5Q6N7 | Thioredoxin domain containing 5 | TXNDC5 | 280.05 | 0.711518519 | —— | 1.256707071 |
| W5NRF4 | Amine oxidase | LOC101113086 | 190.23 | 1.593555556 | —— | 3.346490741 |
| W5PS88 | Aspartate aminotransferase | GOT1 | 144.92 | 1.257992467 | —— | 1.240838384 |
| W5P031 | Haloacid dehalogenase like hydrolase domain containing 2 | HDHD2 | 141.96 | 0.646388889 | —— | 1.331708333 |
| W5Q633 | Biliverdin reductase A | BLVRA | 75.7 | 0.779333333 | —— | 1.239148148 |
| W5PIJ2 | Adenosylhomocysteinase | —— | 70.68 | 1.241181818 | —— | 1.255685185 |
| W5PPE8 | Cytochrome c oxidase subunit 4I1 | COX4I1 | 67.18 | 1.306912698 | —— | 1.218055556 |
| W5PC82 | Malic enzyme | ME1 | 47.43 | 0.687777778 | —— | 1.236022222 |
| W5NU56 | SLC25A12 | SLC25A12 | 39.77 | 1.361592593 | —— | 1.2035 |
| Group B | ||||||
| W5P5A0 | Uncharacterized protein | FLNA | 31757.43 | —— | 0.77936248 | 1.482596475 |
| P60713 | Actin, cytoplasmic 1 | ACTB | 31624.72 | —— | 1.566222222 | 1.223288889 |
| B2LU28 | TPM1 | TPM1 | 18167.25 | —— | 1.263462963 | 0.778351852 |
| W5NRW4 | Tropomyosin 4 | TPM4 | 15549.46 | —— | 1.636562092 | 0.73072549 |
| W5P481 | Collagen type I alpha 1 chain | COL1A1 | 14420.3 | —— | 1.495122306 | 0.665721805 |
| W5NZK9 | Filamin C | FLNC | 13158.81 | —— | 0.811444758 | 1.343351199 |
| W5NTT7 | Collagen type I alpha 2 chain | COL1A2 | 6289.12 | —— | 1.619763957 | 0.744984553 |
| W5NU63 | Myosin heavy chain 10 | MYH10 | 5804.57 | —— | 0.734888889 | 1.292046296 |
| W5NX51 | Apolipoprotein A1 | APOA1 | 2679.74 | —— | 1.543137778 | 1.346196296 |
| P68057 | Hemoglobin subunit beta-C | —— | 2398.75 | —— | 1.270794872 | 1.371555556 |
| W5PT68 | Filamin B | FLNB | 2086.94 | —— | 0.771263889 | 1.341930556 |
| C6ZP47 | I alpha globin | —— | 1919.18 | —— | 1.308829736 | 1.42840048 |
| W5NW47 | Myosin tail 1 domain-containing protein | LOC101106313 | 1650.75 | —— | 1.223851852 | 0.82562963 |
| W5PWD6 | 14-3-3 protein gamma | YWHAG | 1485.72 | —— | 1.324644444 | 0.731088889 |
| W5PBN5 | Myosin heavy chain 8 | MYH8 | 1107.28 | —— | 0.616444444 | 0.430333333 |
| W5Q4S0 | Collagen type III alpha 1 chain | COL3A1 | 995.06 | —— | 1.459057613 | 0.705459534 |
| A8WEG2 | Troponin C | —— | 861.18 | —— | 1.506585185 | 0.604841975 |
| W5QAA9 | Aconitate hydratase | ACO2 | 733.61 | —— | 0.815577061 | 1.485605735 |
| W5P983 | Collagen type IV alpha 2 chain | COL4A2 | 685.25 | —— | 0.798526455 | 1.398399471 |
| P29701 | Alpha-2-HS-glycoprotein | AHSG | 599.54 | —— | 1.420521368 | 1.243254274 |
| P02190 | Myoglobin | MB | 579.37 | —— | 1.402347826 | 0.628536232 |
| Q9N114 | Cytochrome oxidase subunit Va | —— | 556.35 | —— | 1.405451537 | 1.233250591 |
| W5PPG3 | Aldedh domain-containing protein | ALDH9A1 | 546.07 | —— | 0.785277778 | 0.774794118 |
| W5NRC7 | Troponin T3 | TNNT3 | 486.89 | —— | 1.234900463 | 0.830506944 |
| W5P748 | Galectin | LOC101112249 | 479.39 | —— | 3.122481481 | 2.185203704 |
| W5NWM2 | Apolipoprotein A4 | APOA4 | 318.78 | —— | 1.662979328 | 1.208015504 |
| W5P5C5 | Glycogenin 1 | GYG1 | 263.65 | —— | 0.576125 | 0.809409722 |
| W5PGC5 | Aldose 1-epimerase | GALM | 241.97 | —— | 1.401569444 | 1.897486111 |
| A0A075B6D9 | Caveolin | —— | 227.88 | —— | 0.763111111 | 0.620555556 |
| W5QCI5 | Parvalbumin | PVALB | 206.89 | —— | 1.406911111 | 0.566125926 |
| W5QCI3 | NADH-cytochrome b5 reductase | CYB5R3 | 206.13 | —— | 0.818513889 | 1.251319444 |
| W5PJJ7 | Aldo_ket_red domain-containing protein | LOC101109421 | 187.86 | —— | 1.563177778 | 1.366955556 |
| W5PMM4 | Aldo_ket_red domain-containing protein | LOC106990122 | 177.11 | —— | 2.092444444 | 2.513333333 |
| P12303 | Transthyretin | TTR | 161.24 | —— | 1.343354167 | 1.234902778 |
| W5Q754 | Titin | TTN | 150.01 | —— | 1.299432099 | 0.628021605 |
| W5PIN8 | Clusterin | —— | 143.04 | 4.406469136 | 4.23745679 | |
| B2MVW8 | SLC25A4 | SLC25A4 | 136.7 | —— | 0.573244444 | 1.2426 |
| W5P8F9 | BPI1 domain-containing protein | —— | 126.58 | —— | 1.431587302 | 1.385888889 |
| W5QH50 | Histidine rich glycoprotein | HRG | 124.12 | —— | 1.227418803 | 1.232094017 |
| B0LXN9 | Glutathione S-transferase | —— | 121.87 | —— | 2.417412698 | 2.150555556 |
| W5Q5K9 | Actin-related protein 2/3 complex subunit 3 | ARPC3 | 117.72 | —— | 0.785027778 | 0.783152778 |
| B0LRN3 | Histone H3 | —— | 101.87 | —— | 1.275039216 | 0.815529412 |
| G3FIP6 | Glutathione peroxidase-1 | —— | 86.49 | —— | 1.389916667 | 1.378027778 |
| W5Q8X9 | Phosphoglycolate phosphatase | PGP | 79.37 | —— | 0.824740741 | 1.208703704 |
| B2MVW1 | Tetraspanin | CD81 | 73.64 | —— | 0.542555556 | 0.743333333 |
| W5Q7Q6 | Epoxide hydrolase | EPHX1 | 69.85 | —— | 0.688037037 | 1.240259259 |
| W5QD62 | Transgelin | TAGLN3 | 66.76 | —— | 0.771888889 | 0.738555556 |
| W5NVY3 | Clathrin light chain | CLTB | 53.41 | —— | 1.243481481 | 0.827277778 |
| W5PHQ8 | Rhodanese domain-containing protein | TSTD1 | 51.71 | —— | 0.776 | 0.649666667 |
| W5NSG2 | Transmembrane protein 43 | TMEM43 | 44.24 | —— | 0.724555556 | 1.21562963 |
| C8BKE1 | Signal transducer and activator of transcription | —— | 40.49 | —— | 0.697333333 | 1.242222222 |
| W5PXA7 | Collagen type V alpha 3 chain | COL5A3 | 40.13 | —— | 1.435722222 | 0.692888889 |
| W5P013 | Syntrophin beta 2 | SNTB2 | 38.85 | —— | 0.688666667 | 0.779222222 |
| B7U168 | SLC25A3 | —— | 36 | —— | 0.544277778 | 1.399611111 |
| M4WDN5 | Thymosin beta 4 | ThymB4X | 35.27 | —— | 1.509222222 | 0.674484848 |
| W5Q227 | Doublecortin domain containing 1 | DCDC1 | 32.86 | —— | 1.201888889 | 1.495444444 |
| W5NPT4 | Sulfotransferase | SULT1C4 | 25.77 | —— | 1.447148148 | 1.36862963 |
| W5PDL8 | HD domain containing 2 | HDDC2 | 21.82 | —— | 1.251555556 | 1.328111111 |
| Group C | ||||||
| W5NUU3 | Tropomyosin 3 | TPM3 | 12393.68 | 0.829888889 | 1.333767574 | 0.619770975 |
| B6E3I6 | Creatine kinase B | —— | 4518.13 | 0.686777778 | 1.290932099 | 1.465969136 |
| Q9MZW2 | Type VI collagen alpha 3 chain | COL6A1 | 2804.42 | 0.766555556 | 0.804592593 | 0.811851852 |
| A0A0H3V7A0 | Myosin light chain 1 transcript variant 2 | MYL1b | 1426.27 | 0.72 | 1.249492063 | 0.621801587 |
| W5P5I7 | Calmodulin | CALM2 | 873.05 | 0.72337037 | 1.462655123 | 0.712036075 |
| W5Q5Z3 | Keratin 77 | KRT77 | 600.69 | 1.523703704 | 0.797222222 | 0.666833333 |
| W5PUV4 | Carbonic anhydrase 1 | CA1 | 451.62 | 0.819305556 | 2.631666667 | 2.471962963 |
| W5P742 | Galectin | LOC101111992 | 295.19 | 1.582037037 | 1.672111111 | 1.233611111 |
| W5NQ46 | Fibrinogen beta chain | FGB | 293.82 | 0.757711111 | 1.317646091 | 1.236897119 |
| O02762 | Apolipoprotein A1 | —— | 290.43 | 1.428259259 | 1.465598291 | 1.201623932 |
| Q6B3Y2 | Caveolin-1 | CAV1 | 265.49 | 1.402 | 0.71 | 1.332555556 |
| W5Q4M3 | Collagen type V alpha 2 chain | COL5A2 | 264.15 | 1.483449612 | 1.488806584 | 0.819353909 |
| W5Q6F0 | Histone H2A | H2AFX | 251.36 | 1.324444444 | 1.496111111 | 0.745111111 |
| W5Q8N4 | Myosin light chain 2 | MYL2 | 238.73 | 0.811814815 | 0.627362963 | 0.364422222 |
| W5NVR8 | Collagen type V alpha 1 chain | COL5A1 | 140.88 | 1.972472813 | 1.582311111 | 0.706472222 |
| W5PJ97 | Apolipoprotein A2 | APOA2 | 119.62 | 1.333074074 | 1.451166667 | 1.300888889 |
| W5PHT7 | Peptidylprolyl isomerase | FKBP3 | 102.29 | 1.201654971 | 1.292506173 | 1.215654321 |
| W5PJU2 | Aldo_ket_red domain-containing protein | AKR7A2 | 90.79 | 1.297293651 | 1.508711111 | 1.305244444 |
| W5PPE0 | Aldo_ket_red domain-containing protein | LOC101109633 | 90.59 | 1.211777778 | 1.508666667 | 1.240888889 |
| W5P118 | Troponin I2 | TNNI2 | 75.62 | 1.367791328 | 1.207730159 | 0.810222222 |
| W5P6M2 | Proteasome 26S subunit | PSMD5 | 72.87 | 0.806111111 | 0.693194444 | 0.805472222 |
| W5Q701 | HIT domain-containing protein | LOC101112491 | 64.26 | 0.731018519 | 1.338018519 | 1.390018519 |
| W5P642 | Cytochrome b-c1 complex subunit 7 | UQCRB | 58.27 | 0.700046296 | 1.394111111 | 1.298777778 |
| W5Q7R8 | Junction plakoglobin | JUP | 54.68 | 1.537444444 | 0.69 | 0.629055556 |
| A0A0R5Z2R2 | Cysteine and glycine-rich protein 2 | CSRP2 | 37.12 | 1.506833333 | 1.656666667 | 0.675222222 |
| W5P1T1 | Anoctamin | —— | 36.26 | 1.363444444 | 0.762277778 | 1.277777778 |
| Q6Q298 | Beta actin | —— | 2527.29 | 2.446611111 | 0.815611111 | 1.377444444 |
References
- Zeng, W.; Wen, W.; Yue, D.; Tian, Y.; Sun, Q. Chinese ethnic meat products: Continuity and development. Meat Sci. 2016, 120, 37–46. [Google Scholar] [CrossRef]
- Grossi, A.; Bolumar, T.; Soltoft-Jensen, J.; Orlien, V. High pressure treatment of brine enhanced pork semitendinosus: Effect on microbial stability, drip loss, lipid and protein oxidation, and sensory properties. Innov. Food Sci. Emerg. Technol. 2014, 22, 11–21. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, Y.; Wang, Y.; Sun, Y.; Pan, D.; Cao, J. Effect of high pressure treatment on metabolite profile of marinated meat in soy sauce. Food Chem. 2018, 240, 662–669. [Google Scholar] [CrossRef]
- Yang, Y.; Sun, Y.; Pan, D.; Wang, Y.; Cao, J. Effects of high pressure treatment on lipolysis-oxidation and volatiles of marinated pork meat in soy sauce. Meat Sci. 2018, 145, 186–194. [Google Scholar] [CrossRef]
- Wang, Z.; He, F.; Rao, W.; Ni, N.; Shen, Q.; Zhang, D. Proteomic Analysis of Goat Longissimus dorsi Muscles with Different Drip Loss Values Related to Meat Quality Traits. Food Sci. Biotechnol. 2016, 25, 425–431. [Google Scholar] [CrossRef]
- Li, J.; Ding, X.; Han, S.; He, T.; Zhang, H.; Yang, L.; Yang, S.; Gai, J. Differential proteomics analysis to identify proteins and pathways associated with male sterility of soybean using iTRAQ-based strategy. J. Proteom. 2016, 138, 72–82. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, L.; Lei, Y.; Shen, H.; Yu, X.; Luo, Y. Differential proteomic analysis to identify proteins associated with quality traits of frozen mud shrimp (Solenocera melantho) using an iTRAQ-based strategy. Food Chem. 2018, 251, 25–32. [Google Scholar] [CrossRef]
- Wei, Y.; Li, X.; Zhang, D.; Liu, Y. Comparison of protein differences between high- and low-quality goat and bovine parts based on iTRAQ technology. Food Chem. 2019, 289, 240–249. [Google Scholar] [CrossRef]
- Sun, X.B.; Huang, J.C.; Li, T.T.; Ang, Y.; Xu, X.L.; Huang, M. Effects of preslaughter shackling on postmortem glycolysis, meat quality, changes of water distribution, and protein structures of broiler breast meat. Poult. Sci. 2019, 98, 4212–4220. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Dominguez-Hernandez, E.; Salaseviciene, A.; Ertbjerg, P. Low-temperature long-time cooking of meat: Eating quality and underlying mechanisms. Meat Sci. 2018, 143, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Cepero-Betancourt, Y.; Opazo-Navarrete, M.; Janssen, A.E.M.; Tabilo-Munizagab, G.; Perez-Won, M. Effects of high hydrostatic pressure (HHP) on protein structure and digestibility of red abalone (Haliotis rufescens) muscle. Innov. Food Sci. Emerg. Technol. 2020, 60, 102282. [Google Scholar] [CrossRef]
- Picard, B.; Gagaoua, M. Meta-proteomics for the discovery of protein biomarkers of beef tenderness: An overview of integrated studies. Food Res. Int. 2020, 127, 108739. [Google Scholar] [CrossRef]
- Liu, R.; Jin, J.P. Calponin isoforms CNN1, CNN2 and CNN3: Regulators for actin cytoskeleton functions in smooth muscle and non-muscle cells. Gene 2016, 585, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Polati, R.; Menini, M.; Robotti, E.; Millioni, R.; Marengo, E.; Novelli, E.; Balzan, S.; Cecconi, D.J.F.C. Proteomic changes involved in tenderization of bovine Longissimus dorsi muscle during prolonged ageing. Food Chem. 2012, 135, 2052–2069. [Google Scholar] [CrossRef]
- D’Alessandro, A.; Marrocco, C.; Zolla, V.; D’Andrea, M.; Zolla, L. Meat quality of the longissimus lumborum muscle of Casertana and Large White pigs: Metabolomics and proteomics intertwined. J. Proteom. 2012, 75, 610–627. [Google Scholar] [CrossRef]
- Jeong, J.Y.; Ibrahim, M.; Kim, M.J.; So, K.; Jeong, Y.D.; Park, S.; Kim, M.; Lee, H.-J. Comparisons of extracellular matrix-related gene expression levels in different adipose tissues from Korean cattle. Livest. Sci. 2017, 198, 138–146. [Google Scholar] [CrossRef]
- van der Flier, A.; Kuikman, I.; Kramer, D.; Geerts, D.; Kreft, M.; Takafuta, T.; Shapiro, S.S.; Sonnenberg, A. Different splice variants of filamin-B affect myogenesis, subcellular distribution, and determine binding to integrin beta subunits. J. Cell Biol. 2002, 156, 361–376. [Google Scholar] [CrossRef]
- Kim, H.; McCulloch, C.A. Filamin A mediates interactions between cytoskeletal proteins that control cell adhesion. FEBS Lett. 2011, 585, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Lana, A.; Zolla, L. Proteolysis in meat tenderization from the point of view of each single protein: A proteomic perspective. J. Proteom. 2016, 147, 85–97. [Google Scholar] [CrossRef]
- Fidler, A.L.; Boudko, S.P.; Rokas, A.; Hudson, B.G. The triple helix of collagens—An ancient protein structure that enabled animal multicellularity and tissue evolution. J. Cell Sci. 2018, 131, jcs203950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bella, J. Collagen structure: New tricks from a very old dog. Biochem. J. 2016, 473, 1001–1025. [Google Scholar] [CrossRef] [PubMed]
- Chaosap, C.; Sitthigripong, R.; Sivapirunthep, P.; Pungsuk, A.; Adeyemi, K.D.; Sazili, A.Q. Myosin heavy chain isoforms expression, calpain system and quality characteristics of different muscles in goats. Food Chem. 2020, 321, 126677. [Google Scholar] [CrossRef] [PubMed]
- Granzier, H.L.; Labeit, S. Titin and its associated proteins: The third myofilament system of the sarcomere. In Fibrous Proteins: Muscle and Molecular Motors; Gulf Professional Publishing: Houston, TX, USA, 2005; Volume 71, p. 89. [Google Scholar]
- Keller, T.C.S.; Eilertsen, K.; Higginbotham, M.; Kazmierski, S.; Kim, K.T.; Velichkova, M. Role of titin in nonmuscle and smooth muscle cells. In Elastic Filaments of the Cell; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2000; Volume 481, pp. 265–281. [Google Scholar]
- Rubio, M.P.; Geraghty, K.M.; Wong, B.H.C.; Wood, N.T.; Campbell, D.G.; Morrice, N.; Mackintosh, C. 14-3-3-affinity purification of over 200 human phosphoproteins reveals new links to regulation of cellular metabolism, proliferation and trafficking. Biochem. J. 2004, 379, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Obsil, T.; Obsilova, V. Structural basis of 14-3-3 protein functions. Semin. Cell Dev. Biol. 2011, 22, 663–672. [Google Scholar] [CrossRef]
- D’Alessandro, A.; Rinalducci, S.; Marrocco, C.; Zolla, V.; Napolitano, F.; Zolla, L. Love me tender: An Omics window on the bovine meat tenderness network. J. Proteom. 2012, 75, 4360–4380. [Google Scholar] [CrossRef]
- Higgs, H.N.; Pollard, T.D. Regulation of actin filament network formation through Arp2/3 complex: Activation by a diverse array of proteins. Annu. Rev. Biochem. 2001, 70, 649–676. [Google Scholar] [CrossRef]
- Poleti, M.D.; Regitano, L.C.A.; Souza, G.H.M.F.; Cesar, A.S.M.; Simas, R.C.; Silva-Vignato, B.; Oliveira, G.B.; Andrade, S.C.S.; Cameron, L.C.; Coutinho, L.L. Data from proteomic analysis of bovine Longissimus dorsi muscle associated with intramuscular fat content. Data Brief 2018, 19, 1314–1317. [Google Scholar] [CrossRef]





 from curated databases;
from curated databases;  experimentally determined), predicted interactions (
experimentally determined), predicted interactions (  gene neighborhood;
gene neighborhood;  gene fusions;
gene fusions;  gene co-occurrence) and others (
gene co-occurrence) and others (  textmining;
textmining;  co-expression;
co-expression;  protein homology)). Some proteins without protein identifiers do not appear in the figure.
from curated databases; experimentally determined), predicted interactions ( gene neighborhood; gene fusions; gene co-occurrence) and others ( textmining; co-expression; protein homology)). Some proteins without protein identifiers do not appear in the figure.
protein homology)). Some proteins without protein identifiers do not appear in the figure.
from curated databases; experimentally determined), predicted interactions ( gene neighborhood; gene fusions; gene co-occurrence) and others ( textmining; co-expression; protein homology)). Some proteins without protein identifiers do not appear in the figure.
| CN | HT | HPP | |
|---|---|---|---|
| Hardness | 247.63 ± 50.366 a | 435.314 ± 87.039 b | 255.633 ± 64.275 a |
| Springiness | 0.889 ± 0.054 a | 0.912 ± 0.125 a | 2.401 ± 0.2 b |
| Gumminess | 206.986 ± 35.032 a | 369.368 ± 51.983 b | 240.797 ± 58.856 a |
| Chewiness | 185.185 ± 40.766 a | 332.062 ± 4.675 a | 584.52 ± 180.439 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, N.; Hou, R.; Liu, Y.; Han, P.; Zhao, W.; Wu, W.; Lu, S.; Ji, H.; Dong, J. Application of iTRAQ Technology to Identify Differentially Expressed Proteins of Sauce Lamb Tripe with Different Secondary Pasteurization Treatments. Foods 2022, 11, 1166. https://doi.org/10.3390/foods11081166
An N, Hou R, Liu Y, Han P, Zhao W, Wu W, Lu S, Ji H, Dong J. Application of iTRAQ Technology to Identify Differentially Expressed Proteins of Sauce Lamb Tripe with Different Secondary Pasteurization Treatments. Foods. 2022; 11(8):1166. https://doi.org/10.3390/foods11081166
Chicago/Turabian StyleAn, Ning, Ran Hou, Yangming Liu, Ping Han, Wei Zhao, Wenxia Wu, Shiling Lu, Hua Ji, and Juan Dong. 2022. "Application of iTRAQ Technology to Identify Differentially Expressed Proteins of Sauce Lamb Tripe with Different Secondary Pasteurization Treatments" Foods 11, no. 8: 1166. https://doi.org/10.3390/foods11081166