
Purification and Characterization of the Protease from Staphylococcus xylosus A2 Isolated from Harbin Dry Sausages



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Microorganism and Protease Production
2.3. Protease Activity and Concentration Determination
2.4. Proteases Purification
2.4.1. Ammonium Sulfate Fractionation
2.4.2. Cation Exchange Chromatography
2.4.3. Gel-Filtration Chromatography
2.5. SDS-PAGE
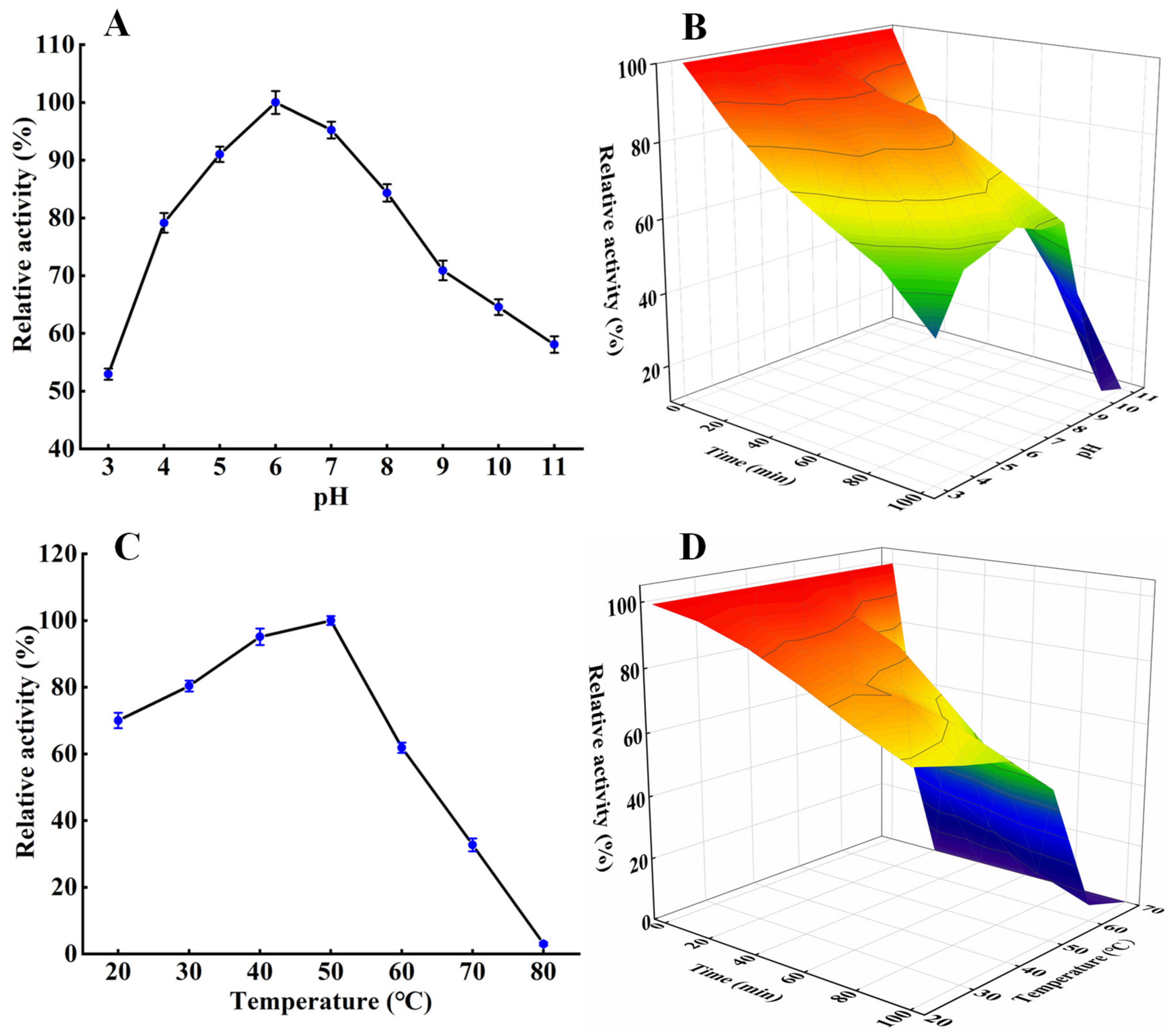
2.6. Effects of pH and Temperature on the Protease Activity and Stability
2.7. Effects of Metallic Ions and Inhibitors on Protease Activity
2.8. Determination of Activation Energy
2.9. Determination of Kinetic and Thermodynamics Parameters of the Protease
2.10. Thermal Inactivation
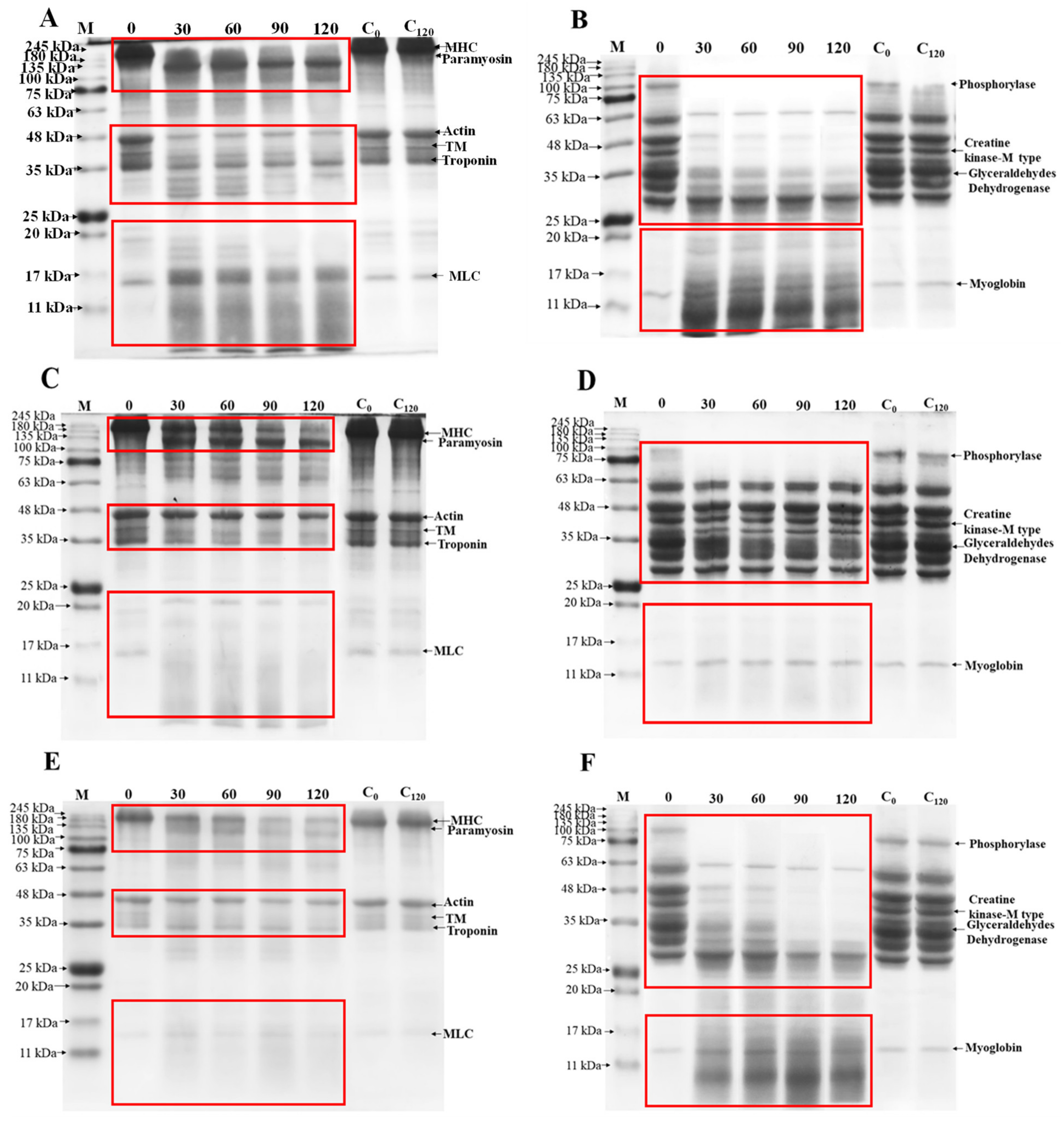
2.11. Extraction and Degradation of MP and SP
2.12. Determination of Particle Size
2.13. Confocal Laser Scanning Microscopy (CLSM)
2.14. Cell Toxicity Assay
2.15. Cell Morphology
2.16. The Peptide Concentration Measurements
2.17. Statistical Analysis
3. Results and Discussion
3.1. The Growth Curve of S. xylosus A2
3.2. Protease Purification
3.3. Effects of pH and Temperature on the Protease Activity and Stability
3.4. Influence of Metallic Ions and Inhibitors on Activity of the S. xylosus A2 Protease
3.5. Ea Analysis
3.6. Kinetic Constants and Thermal Stability of Casein Hydrolysis
3.7. Protein Profiles
3.8. Distribution and Particle Diameter
3.9. Cell Toxicity Assay
3.10. Morphological Features of Cell
3.11. Peptide Concentrations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, F.; Hu, Y.; Chen, Q.; Kong, B.; Liu, Q. Purification and biochemical characteristics of the extracellular protease from Pediococcus pentosaceus isolated from Harbin dry sausages. Meat Sci. 2019, 156, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, S.; Bkhairia, I.; Nasri, R.; Mechichi, T.; Nasri, M.; Kamoun, A.S. Evaluation of the biotechnological potential of a novel purified protease BS1 from Bacillus safensis S406 on the chitin extraction and detergent formulation. Int. J. Biol. Macromol. 2017, 104, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Greppi, A.; Ferrocino, I.; Storia, A.L.; Rantsiou, K.; Ercolini, D.; Cocolin, L. Monitoring of the microbiota of fermented sausages by culture independent rRNA-based approaches. Int. J. Food Microbiol. 2015, 212, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Mauriello, G.; Casaburi, A.; Blaiotta, G.; Villani, F. Isolation and technological properties of coagulase negative staphylococci from fermented sausages of Southern Italy. Meat Sci. 2004, 67, 149–158. [Google Scholar] [CrossRef]
- Sun, F.; Li, Q.; Liu, H.; Kong, B.; Chen, Q. Purification and biochemical characteristics of the protease from Lactobacillus brevis R4 isolated from Harbin dry sausages. LWT Food Sci. Technol. 2019, 113, 108287. [Google Scholar] [CrossRef]
- Cachaldora, A.; Fonseca, S.; Franco, I.; Carballo, J. Technological and safety characteristics of Staphylococcaceae isolated from Spanish traditional dry-cured sausages. Food Microbiol. 2013, 33, 61–68. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, Q.; Wen, R.; Wang, Y.; Qin, L.; Kong, B. Quality characteristics and flavor profile of Harbin dry sausages inoculated with lactic acid bacteria and Staphylococcus xylosus. LWT Food Sci. Technol. 2019, 114, 108392. [Google Scholar] [CrossRef]
- Cruxen, C.E.D.S.; Funck, G.D.; Dannenberg, G.D.S.; Haubert, L.; Marques, J.D.L.; Kroning, I.S.; Chaves, F.C.; Da Silva, W.P.; Fiorentini, A.M. Characterization of Staphylococcus xylosus LQ3 and its application in dried cured sausage. LWT Food Sci. Technol. 2017, 86, 538–543. [Google Scholar] [CrossRef]
- Hu, M.; Yu, J.; Yu, J.; Pan, Y.; Ou, Y. Isolation and screening of Staphylococcus xylosus P2 from Chinese bacon: A novel starter culture in fermented meat products. Int. J. Food Eng. 2018, 15, 20180021. [Google Scholar]
- Berardo, A.; Devreese, B.; Maere, H.D.; Stavropoulou, D.A.; Royen, G.V.; Leroy, F.; De Smet, S. Actin proteolysis during ripening of dry fermented sausages at different pH values. Food Chem. 2017, 221, 1322–1332. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowska, A.; Bajzert, J.; Babij, K.; Szołtysik, M.; Stefaniak, T.; Willak-Janc, E.; Chrzanowska, J. Reduced IgE and IgG antigenic response to milk proteins hydrolysates obtained with the use of non-commercial serine protease from Yarrowia lipolytica. Food Chem. 2020, 302, 125350. [Google Scholar] [CrossRef]
- Yu, P.; Huang, X.; Ren, Q.; Wang, X. Purification and characterization of a H2O2-tolerant alkaline protease from Bacillus sp. ZJ1502, a newly isolated strain from fermented bean curd. Food Chem. 2019, 274, 510–517. [Google Scholar] [CrossRef]
- Chen, Q.; Kong, B.; Han, Q.; Liu, Q.; Xu, L. The role of bacterial fermentation in the hydrolysis and oxidation of sarcoplasmic and myofibrillar proteins in Harbin dry sausages. Meat Sci. 2016, 121, 196–206. [Google Scholar] [CrossRef]
- Sun, F.; Sun, Q.; Zhang, H.; Kong, B.; Xia, X. Purification and biochemical characteristics of the microbial extracellular protease from Lactobacillus curvatus isolated from Harbin dry sausages. Int. J. Biol. Macromol. 2019, 133, 987–997. [Google Scholar] [CrossRef]
- Sinha, R.; Khare, S.K. Characterization of detergent compatible protease of a halophilic Bacillus sp. EMB9: Differential role of metallic ions in stability and activity. Bioresour. Technol. 2013, 145, 357–361. [Google Scholar] [CrossRef]
- Ravindran, B.; Wong, J.W.C.; Selvam, A.; Thirunavukarasu, K.; Sekaran, G. Microbial biodegradation of proteinaceous tannery solid waste and production of a novel value added product-metalloprotease. Bioresour. Technol. 2016, 217, 150–156. [Google Scholar] [CrossRef]
- Afsharnezhad, M.; Shahangian, S.S.; Sariri, R. A novel milk-clotting cysteine protease from Ficus johannis: Purification and characterization. Int. J. Biol. Macromol. 2019, 121, 173–182. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during assembly of head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bhunia, B.; Basak, B.; Mandal, T.; Bhattacharya, P.; Dey, A. Effect of pH and temperature on stability and kinetics of novel extracellular serine alkaline protease (70 kDa). Int. J. Biol. Macromol. 2013, 54, 1–8. [Google Scholar] [CrossRef]
- Mostafa, F.A.; Wahab, W.A.A.; Salah, H.A.; Nawwar, G.A.M.; Esawy, M.A. Kinetic and thermodynamic characteristic of Aspergillus awamori EM66 levansucrase. Int. J. Biol. Macromol. 2018, 119, 232–239. [Google Scholar] [CrossRef]
- Da Silva, O.S.; de Oliveira, R.L.; Silva, J.D.C.; Converti, A.; Porto, T.S. Thermodynamic investigation of an alkaline protease from Aspergillus tamarii URM4634: A comparative approach between crude extract and purified enzyme. Int. J. Biol. Macromol. 2018, 109, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, H.; Liu, Q.; Chen, Q.; Kong, B. Solubilization and stable dispersion of myofibrillar proteins in water through the destruction and inhibition of the assembly of filaments using high-intensity ultrasound. Ultrason. Sonochem. 2020, 67, 105160. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, J.; Zheng, B.; Guo, Z. Impact of combined ultrasound-microwave treatment on structural and functional properties of golden threadfin bream (Nemipterus virgatus) myofibrillar proteins and hydrolysates. Ultrason. Sonochem. 2020, 65, 105063. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Hu, Y.; Yin, X.; Kong, B.; Qin, L. Production, purification and biochemical characterization of the microbial protease produced by Lactobacillus fermentum R6 isolated from Harbin dry sausages. Process Biochem. 2020, 89, 37–45. [Google Scholar] [CrossRef]
- Hou, E.; Xia, T.; Zhang, Z.; Mao, X. Purification and characterization of an alkaline protease from Micrococcus sp. isolated from the South China Sea. J. Ocean Univ. China 2017, 16, 319–325. [Google Scholar] [CrossRef]
- Negi, S.; Banerjee, R. Characterization of amylase and protease produced by Aspergillus awamori in a single bioreactor. Food Res. Int. 2009, 42, 443–448. [Google Scholar] [CrossRef]
- Reddy, M.R.; Reddy, K.S.; Chouhan, Y.R.; Bee, H.; Reddy, G. Effective feather degradation and keratinase production by Bacillus pumilus GRK for its application as bio-detergent additive. Bioresour. Technol. 2017, 243, 254–263. [Google Scholar] [CrossRef]
- Benkiar, A.; Nadia, Z.J.; Badis, A.; Rebzani, F.; Soraya, B.T.; Rekik, H.; Naili, B.; Ferradji, F.Z.; Bejar, S.; Jaouadi, B. Biochemical and molecular characteriza tion of a thermo- and detergent-stable alkaline serine keratinolytic protease from Bacillus circulans strain DZ100 for detergent formulations and feather-biodegradation process. Int. Biodeterior. Biodegrad. 2013, 83, 129–138. [Google Scholar] [CrossRef]
- Thakrar, F.J.; Singh, S.P. Catalytic, thermodynamic and structural properties of an immobilized and highly thermostable alkaline protease from a haloalkaliphilic actinobacteria, Nocardiopsis alba TATA-5. Bioresour. Technol. 2019, 278, 150–158. [Google Scholar] [CrossRef]
- Hernández-Martínez, R.; Gutiérrez-Sánchez, G.; Bergmann, C.W.; Loera-Corral, O.; Rojo-Domínguez, A.; Huerta-Ochoa, S.; Regalado-Gonzalez, C.; Prado-Barragan, L.A. Purification and characterization of a thermodynamic stable serine protease from Aspergillus fumigatus. Process Biochem. 2011, 46, 2001–2006. [Google Scholar] [CrossRef]
- Abdel-Naby, M.A.; Ahmed, S.A.; Wehaidy, H.R.; El-Mahdy, S.A. Catalytic, kinetic and thermodynamic properties of stabilized Bacillus stearothermophilus alkaline protease. Int. J. Biol. Macromol. 2017, 96, 265–271. [Google Scholar] [CrossRef]
- De Borba, T.M.; Machado, T.B.; Brandelli, A.; Kalil, S.J. Thermal stability and catalytic properties of protease from Bacillus sp. P45 active in organic solvents and ionic liquid. Biotechnol. Progr. 2018, 34, 1102–1108. [Google Scholar] [CrossRef]
- Salehi, M.; Aghamaali, M.R.; Sajedi, R.H.; Asghari, S.M.; Jorjani, E. Purification and characterization of a milk-clotting aspartic protease from Withania coagulans fruit. Int. J. Biol. Macromol. 2017, 98, 847–854. [Google Scholar] [CrossRef]
- Sharma, A.K.; Kikani, B.A.; Singh, S.P. Biochemical, thermodynamic and structural characteristics of a biotechnologically compatible alkaline protease from a haloalkaliphilic, Nocardiopsis dassonvillei OK-18. Int. J. Biol. Macromol. 2020, 153, 680–696. [Google Scholar] [CrossRef]
- Wehaidy, H.R.; Abdel-Naby, M.A.; Shousha, W.G.; Elmallah, M.I.Y.; Shawky, M.M. Improving the catalytic, kinetic and thermodynamic properties of Bacillus subtilis KU710517 milk clotting enzyme via conjugation with polyethylene glycol. Int. J. Biol. Macromol. 2018, 111, 296–301. [Google Scholar] [CrossRef]
- Liu, R.; Liu, Q.; Xiong, S.; Fu, Y.; Chen, L. Effects of high intensity unltrasound on structural and physicochemical properties of myosin from Silver Carp. Ultrason. Sonochem. 2017, 37, 150–157. [Google Scholar] [CrossRef]
- Liu, J.; Chen, Z.; He, J.; Zhang, Y.; Zhang, T.; Jiang, Y. Anti-oxidative and anti-apoptosis effects of egg white peptide, Trp-Asn-Trp-Ala-Asp, against H2O2-induced oxidative stress in human embryonic kidney 293 cells. Food Funct. 2014, 5, 3179–3188. [Google Scholar] [CrossRef]
- El-Sayed, N.S.; Sajid, M.I.; Parang, K.; Tiwari, R.K. Synthesis, characterization, and cytotoxicity evaluation of dextran-myristoyl-ECGKRK peptide conjugate. Int. J. Biol. Macromol. 2021, 191, 1204–1211. [Google Scholar] [CrossRef]
- Lei, F.; Zhao, Q.; Sun-Waterhouse, D.; Zhao, M. Characterization of a salt-tolerant aminopeptidase from marine Bacillus licheniformis SWJS33 that improves hydrolysis and debittering efficiency for soy protein isolate. Food Chem. 2017, 214, 347–353. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Steps | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg Protein) | Purification Fold | Yield (%) |
|---|---|---|---|---|---|
| Culture broth | 68.23 ± 1.26 | 29.23 ± 1.10 | 2.34 ± 0.21 | 1 | 100 |
| Ammonium sulfate precipitation | 35.54 ± 1.06 | 13.24 ± 0.36 | 2.69 ± 0.30 | 1.15 | 51.4 |
| DEAE-Sepharose™ FF | 19.34 ± 1.11 | 1.25 ± 0.04 | 15.81 ± 0.56 | 6.75 | 27.9 |
| Sephadex-G75 chromatography | 10.36 ± 1.20 | 0.33 ± 0.02 | 33.31 ± 0.68 | 14.23 | 14.7 |
| Parameter | Data |
|---|---|
| Ea (kJ·mol−1) | 13.80 ± 0.81 |
| Vmax (mg·min−1) | 19.45 ± 0.69 |
| Km (mg·mL−1) | 2.94 ± 0.14 |
| kcat (s−1) | 3.24 ± 0.29 |
| ΔH* (kJ·mol−1) | 11.11 ± 0.45 |
| ΔG* (kJ·mol−1) | 45.16 ± 1.48 |
| ΔS* (J·mol−1K−1) | −105.42 ± 2.17 |
| Temperature | T (°K) | T1/2 (min) | Kd (min−1) | D (min) | ΔH*d (kJ·mol−1) | ΔG*d (kJ·mol−1) | ΔS*d (J·mol−1K−1) | E(a)d (kJ·mol−1) |
|---|---|---|---|---|---|---|---|---|
| 40 °C | 313 | 102.64 ± 2.88 | 0.0067 ± 0.0004 | 343.73 ± 25.63 | 25.92 ± 1.38 | 89.80 ± 4.11 | −204.09 ± 10.67 | - |
| 50 °C | 323 | 77.02 ± 1.71 | 0.009 ± 0.0003 | 255.89 ± 17.37 | 25.83 ± 2.61 | 91.97 ± 5.63 | −204.77 ± 8.83 | 28.52 ± 2.13 |
| 60 °C | 333 | 52.89 ± 1.95 | 0.013 ± 0.0072 | 177.15 ± 16.74 | 25.75 ± 1.93 | 93.88 ± 4.31 | −204.59 ± 6.57 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Xu, J.; Kong, B.; Liu, Q.; Xia, X.; Sun, F. Purification and Characterization of the Protease from Staphylococcus xylosus A2 Isolated from Harbin Dry Sausages. Foods 2022, 11, 1094. https://doi.org/10.3390/foods11081094
Wang H, Xu J, Kong B, Liu Q, Xia X, Sun F. Purification and Characterization of the Protease from Staphylococcus xylosus A2 Isolated from Harbin Dry Sausages. Foods. 2022; 11(8):1094. https://doi.org/10.3390/foods11081094
Chicago/Turabian StyleWang, Hui, Jianhang Xu, Baohua Kong, Qian Liu, Xiufang Xia, and Fangda Sun. 2022. "Purification and Characterization of the Protease from Staphylococcus xylosus A2 Isolated from Harbin Dry Sausages" Foods 11, no. 8: 1094. https://doi.org/10.3390/foods11081094