
Recent Advances in Microbial Synthesis of Poly-γ-Glutamic Acid: A Review

 ,
,
Abstract
:1. Introduction
2. The Functional Properties of γ-PGA
3. Microbial Sources of γ-PGA
4. Synthetic Pathway of γ-PGA
4.1. The Racemization Pathway of γ-PGA
4.2. The Polymerization and Regulation Pathway of γ-PGA
4.3. The Catabolism Pathway of γ-PGA
5. Improvement of Microbial γ-PGA Synthesis
5.1. Genetic Manipulation
5.2. Culture Medium Optimization
6. Purification of γ-PGA
7. Application of γ-PGA
7.1. Food Industry
7.2. Agricultural Field
7.3. Bio-Medical Field
7.4. Other Fields
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shih, I.L.; Van, Y.T. The production of poly-(gamma-glutamic acid) from microorganisms and its various applications. Bioresour. Technol. 2001, 79, 207–225. [Google Scholar] [CrossRef]
- Ashiuchi, M.; Misono, H. Biochemistry and molecular genetics of poly-γ-glutamate synthesis. Appl. Microbiol. Biotechnol. 2002, 59, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Sirisansaneeyakul, S.; Cao, M.F.; Kongklom, N.; Chuensangjun, C.; Shi, Z.P.; Chisti, Y. Microbial production of poly-γ-glutamic acid. World J. Microbiol. Biotechnol. 2017, 33, 173. [Google Scholar] [CrossRef]
- Richard, A.; Margaritis, A. Poly (glutamic acid) for biomedical applications. Crit. Rev. Biotechnol. 2001, 21, 219–232. [Google Scholar] [CrossRef]
- Sung, M.H.; Park, C.; Kim, C.J.; Poo, H.; Soda, K.; Ashiuchi, M. Natural and edible biopolymer poly-γ-glutamic acid: Synthesis, production, and applications. Chem. Rec. 2005, 5, 352–366. [Google Scholar] [CrossRef] [PubMed]
- Akagi, T.; Matsusaki, M.; Akashi, M. Pharmaceutical and Medical Applications of Poly-Gamma-Glutamic Acid; Microbiology Monographs, (Volume: Amino-Acid Homopolymers Occurring in Nature: 119–153); Hamano, Y., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 15. [Google Scholar] [CrossRef]
- Ashiuchi, M. Occurrence and Biosynthetic Mechanism of Poly-Gamma-Glutamic Acid; Microbiology Monographs, (Volume: Amino-Acid Homopolymers Occurring in Nature: 77–93); Hamano, Y., Ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar] [CrossRef]
- Ashiuchi, M. Biochemical engineering of PGA. Microb. Biotechnol. 2013, 6, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, I.; Singhal, R. Poly (glutamic acid)—An emerging biopolymer of commercial interest. Bioresour. Technol. 2011, 102, 5551–5561. [Google Scholar] [CrossRef]
- Buescher, J.M.; Margaritis, A. Microbial biosynthesis of polyglutamic acid biopolymer and applications in the biopharmaceutical, biomedical and food industries. Crit. Rev. Biotechnol. 2007, 27, 1–19. [Google Scholar] [CrossRef]
- Luo, Z.; Guo, Y.; Liu, J.; Qiu, H.; Zhao, M.; Zou, W.; Li, S. Microbial synthesis of poly-γ-glutamic acid: Current progress, challenges, and future perspectives. Biotechnol. Biofuels 2016, 9, 134. [Google Scholar] [CrossRef] [Green Version]
- Ogunleye, A.; Bhat, A.; Irorere, V.U.; Hill, D.; Williams, C.; Radecka, I. Poly-γ-glutamic acid: Production, properties and applications. Microbiology 2014, 161, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wei, X.; Chen, S. Production and Application of Poly-γ-Glutamic Acid. Biotechnology of Microbial Enzymes, (Chapter 2: Production. Isolation and Purification of Industrial Products). 2017. Available online: https://www.sciencedirect.com/book/9780128037256/biotechnology-of-microbial-enzymes#book-description (accessed on 1 February 2022).
- Candela, T.; Moya, M.; Haustant, M.; Fouet, A. Fusobacterium nucleatum, the first gram-negative bacterium demonstrated to produce polyglutamate. Can. J. Microbiol. 2009, 55, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Hezayen, F.F.; Rehm, B.H.; Tindall, B.J.; Steinbuchel, A. Transfer of Natrialba asiatica B1T to Natrialba taiwanensis sp. nov. and description of Natrialba aegyptiaca sp. nov., a novel extremely halophilic, aerobic, non-pigmented member of the Archaea from Egypt that produces extracellular poly (glutamic acid). Int. J. Syst. Evol. Microbiol. 2001, 51, 1133–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamang, J.P.; Shin, D.H.; Jung, S.J.; Chae, S.W. Functional properties of microorganisms in fermented foods. Front. Microbiol. 2016, 7, 578. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Zhu, Y.; Sha, Y.; Lei, P.; Xu, H. Development of a robust bacillus amyloliquefaciens cell factory for efficient poly (γ-glutamic acid) production from jerusalem artichoke. ACS Sustain. Chem. Eng. 2020, 8, 9763–9774. [Google Scholar] [CrossRef]
- Bajaj, I.B.; Singhal, R.S. Enhanced Production of Poly (γ-glutamic acid) from Bacillus licheniformis NCIM 2324 by Using Metabolic Precursors. Appl. Biochem. Biotechnol. 2009, 159, 133–141. [Google Scholar] [CrossRef] [PubMed]
- CaO, M.; Feng, J.; Sirisansaneeyakul, S.; Song, C.; Chisti, Y. Genetic and metabolic engineering for microbial production of poly-γ-glutamic acid. Biotechnol. Adv. 2018, 36, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Hong, X.; Lin, X.; Ouyang, P. Biosynthesis of poly (γ-glutamic acid) in Bacillus subtilis nx-2: Regulation of stereochemical composition of poly (γ-glutamic acid). Process Biochem. 2006, 41, 1650–1655. [Google Scholar] [CrossRef]
- Ashiuchi, M.; Shimanouchi, K.; Nakamura, H.; Kamei, T.; Soda, K.; Park, C.; Sung, M.H.; Misono, H. Enzymatic synthesis of high-molecular-mass poly-γ-glutamate and regulation of its stereochemistry. Appl. Environ. Microbiol. 2004, 70, 4249–4255. [Google Scholar] [CrossRef] [Green Version]
- Ho, G.H.; Ho, T.I.; Hsieh, K.H.; Su, Y.C.; Lin, P.Y.; Yang, J.; Yang, K.H.; Yang, S.H. γ-polyglutamic acid produced by Bacillus subtilis (natto): Structural characteristics, chemical properties and biological functionalities. J. Chin. Chem. Soc. 2006, 53, 1363–1384. [Google Scholar] [CrossRef]
- Bhat, A.R.; Irorere, V.U.; Bartlett, T.; Hill, D.; Kedia, G.; Morris, M.R.; Charalampopoulos, D.; Radecka, I. Bacillus subtilis natto: A non-toxic source of poly-γ-glutamic acid that could be used as a cryoprotectant for probiotic bacteria. AMB Express 2013, 3, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Birrer, G.A.; Cromwick, A.M.; Gross, R.A. γ-poly (glutamic acid) formation by Bacillus licheniformis 9945a: Physiological and biochemical studies. Int. J. Biol. Macromol. 1994, 16, 265–275. [Google Scholar] [CrossRef]
- Chunhachart, O.; Itoh, T.; Sukchotiratana, M.; Tanimoto, H.; Tahara, Y. Characterization of γ-glutamyl hydrolase produced by Bacillus sp. isolated from thai thua-nao. Biosci. Biotechnol. Biochem. 2006, 70, 2779–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.; Choi, J.C.; Choi, Y.H.; Nakamura, H.; Shimanouchi, K.; Horiuchi, T.; Misono, H.; Sewaki, T.; Soda, K.; Ashiuchi, M.; et al. Synthesis of super-high-molecular-weight poly-γ-glutamic acid by Bacillus subtilis subsp. chungkookjang. J. Mol. Catal. B Enzym. 2005, 35, 128–133. [Google Scholar] [CrossRef]
- Peng, Y.; Bo, J.; Tao, Z.; Mu, W.; Ming, M.; Hua, Y. High-level production of poly (γ-glutamic acid) by a newly isolated glutamate-independent strain, Bacillus methylotrophicus. Process Biochem. 2015, 50, 329–335. [Google Scholar] [CrossRef]
- Yin, A.; Jia, Y.; Qiu, T.; Gao, M.; Cheng, S.; Wang, X.; Sun, Y. Poly-γ-glutamic acid improves the drought resistance of maize seedlings by adjusting the soil moisture and microbial community structure. Appl. Soil Ecol. 2018, 129, 128–135. [Google Scholar] [CrossRef]
- Jang, W.J.; Choi, S.Y.; Lee, J.M.; Lee, G.H.; Hasan, M.T.; Kong, I.S. Viability of Lactobacillus plantarum encapsulated with poly-γ-glutamic acid produced by Bacillus sp. SJ-10 during freeze-drying and in an in vitro gastrointestinal model. LWT Food Sci. Technol. 2019, 112, 108222. [Google Scholar] [CrossRef]
- Lee, C.Y.; Kuo, M.I. Effect of γ-polyglutamate on the rheological properties and microstructure of tofu. Food Hydrocoll. 2011, 25, 1034–1040. [Google Scholar] [CrossRef]
- Xie, X.; Wu, X.; Shen, Y.; Song, M.; Xu, C.; Zhang, B.; Aziz, U.; Xu, X. Effect of poly-γ-glutamic acid on hydration and structure of wheat gluten. J. Food Sci. 2020, 85, 3214–3219. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, C.; Ding, J.; Li, M.; Chen, X.; Tang, Z.; Zhuang, X.; Chen, X. A comparative study of linear, Y-shaped and linear-dendritic methoxy poly (ethylene glycol)-block-polyamidoamine-block-poly (γ-glutamic acid) block copolymers for doxorubicin delivery in vitro and in vivo. Acta Biomater. 2016, 40, 243–253. [Google Scholar] [CrossRef]
- Liang, H.F.; Chen, C.T.; Chen, S.C.; Kulkarni, A.R.; Chiu, Y.L.; Chen, M.C.; Sung, H.W. Paclitaxel-loaded poly (gamma glutamic acid)-poly(lactide) nanoparticles as a targeted drug delivery system for the treatment of liver cancer. Biomaterials 2006, 27, 2051–2059. [Google Scholar] [CrossRef]
- Manocha, B.; Margaritis, A. Production and characterization of gamma-polyglutamic acid nanoparticles for controlled anticancer drug release. Crit. Rev. Biotechnol. 2008, 28, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Manocha, B.; Margaritis, A. Controlled Release of Doxorubicin from Doxorubicin/gamma-Polyglutamic Acid Ionic Complex. J. Nanomater. 2010, 2010, 780171. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Song, W.; Tang, Z.; Lv, S.; Lin, L.; Sun, H.; Li, Q.; Yang, Y.; Hong, H.; Chen, X. Nanoscaled Poly (γ-glutamic acid)/Doxorubicin-Amphiphile Complex as pH-responsive Drug Delivery System for Effective Treatment of Nonsmall Cell Lung Cancer. ACS Appl. Mater. Interfaces 2013, 5, 1781–1792. [Google Scholar] [CrossRef]
- Zhang, L.; Chang, J.; Zhao, Y.; Xu, H.; Wang, T.; Li, Q.; Xing, L.; Huang, J.; Wang, Y.; Liang, Q. Fabrication of a triptolide-loaded and poly-γ-glutamic acid-based amphiphilic nanoparticle for the treatment of rheumatoid arthritis. Int. J. Nanomed. 2018, 13, 2051–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.H.; Dong, C.D.; Chen, C.W.; Shea, Y.T.; Kao, C.M. Using poly-glutamic acid as soil-washing agent to remediate heavy metal-contaminated soils. Environ. Sci. Pollut. Res. 2018, 25, 5231–5242. [Google Scholar] [CrossRef]
- Mitsunaga, H.; Meissner, L.; Bu¨chs, J.; Fukusaki, E. Branched chain amino acids maintain the molecular weight of poly (γ-glutamic acid) of Bacillus licheniformis ATCC 9945 during the fermentation. J. Biosci. Bioeng. 2016, 122, 400–405. [Google Scholar] [CrossRef]
- Lee, J.M.; Jang, W.J.; Park, S.H.; Kong, I.S. Antioxidant and gastrointestinal cytoprotective effect of edible polypeptide poly-γ-glutamic acid. Int. J. Biol. Macromol. 2020, 153, 616–624. [Google Scholar] [CrossRef]
- Ben-Zur, N.; Goldman, D.M. γ-Poly glutamic acid: A novel peptide for skin care. Cosmet Toilet. 2007, 122, 64–72. [Google Scholar]
- Nair, P.; Navale, G.R.; Dharne, M.S. Poly-gamma-glutamic acid biopolymer: A sleeping giant with diverse applications and unique opportunities for commercialization. Biomass Conv. Bioref. 2021, 1–19. [Google Scholar] [CrossRef]
- Cheng, C.; Asada, Y.; Aida, T. Production of γ-polyglutamic acid by Bacillus subtilis A35 under denitrifying conditions. Agric. Biol. Chem. 1989, 53, 2369–2375. [Google Scholar] [CrossRef]
- Kubota, H.; Matsunobu, T.; Uotani, K.; Takebe, H.; Satoh, A.; Tanaka, T.; Taniguchi, M. Production of poly (γ-glutamic acid) by Bacillus subtilis F-2-01. Biosci. Biotech. Biochem. 1993, 57, 1212–1213. [Google Scholar] [CrossRef] [PubMed]
- Ashiuchi, M.; Kamei, T.; Baek, D.H.; Shin, S.Y.; Sung, M.H.; Soda, K.; Yagi, T.; Mispno, H. Isolation of Bacillus subtilis (chungkookjang), a poly-γ-glutamate producer with high genetic competence. Appl. Microbiol. Biotechnol. 2001, 57, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Ji, Z.; Chen, S. Isolation of halotolerant Bacillus licheniformis WX-02 and regulatory effects of sodium chloride on yield and molecular sizes of poly-γ-glutamic acid. Appl. Microbiol. Biotechnol. 2010, 160, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, J.; Zhu, X.; Cai, J.; Zhang, A.; Hong, Y.; Huang, J.; Huang, L.; Xu, Z. High-level exogenous glutamic acid independent production of poly-(γ-glutamic acid) with organic acid addition in a new isolated Bacillus subtilis C10. Bioresour. Technol. 2012, 116, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Chen, G.; Wang, Q.; Zheng, S.; Shu, L.; Liang, Z. Metabolic studies of temperature control strategy on poly (gamma-glutamic acid) production in a thermophilic strain Bacillus subtilis GXA-28. Bioresour. Technol. 2014, 155, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Yoshihito, I.; Takeshi, T.; Tetsuo, O.; Ito, Y.; Tanaka, T.; Ohmachi, T.; Asada, Y. Glutamic acid independent production of poly (γ-glutamic acid) by Bacillus subtilis tam-4. Biosci. Biotechnol. Biochem. 1996, 66, 1239–1242. [Google Scholar] [CrossRef] [Green Version]
- Shih, I.L.; Wu, P.J.; Shieh, C.J. Microbial production of a poly (gamma-glutamic acid) derivative by Bacillus subtilis. Process. Biochem. 2005, 40, 2827–2832. [Google Scholar] [CrossRef]
- Cao, M.; Geng, W.; Liu, L.; Song, C.; Xie, H.; Guo, W.; Jin, Y.; Wang, S. Glutamic acid independent production of poly-γ-glutamic acid by Bacillus amyloliquefaciens LL3 and cloning of pgsBCA genes. Bioresour. Technol. 2011, 102, 4251–4257. [Google Scholar] [CrossRef]
- Cachat, E.; Barker, M.; Read, T.D.; Priest, F.G. A Bacillus thuringiensis strain producing a polyglutamate capsule resembling that of Bacillus anthracis. FEMS Microbiol. Lett. 2008, 285, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, P.M.; Datta, S.; Tiwari, D.P.; Raval, R.; Dubey, A.K. Selection of an effective indicator for rapid detection of microorganisms producing γ-polyglutamic acid and its biosynthesis under submerged fermentation conditions using Bacillus methylotrophicus. Appl. Biochem. Biotechnol. 2018, 185, 270–288. [Google Scholar] [CrossRef]
- Ashiuchi, M.; Shimanouchi, K.; Horiuchi, T.; Kame, T.; Misono, H. Genetically engineered poly-gamma-glutamate producer from Bacillus subtilis ISW1214. Biosci. Biotechnol. Biochem. 2006, 70, 1794–1797. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Geng, W.; Zhang, W.; Sun, J.; Wang, S.; Feng, J.; Zheng, P.; Jiang, A.; Song, C. Engineering of recombinant Escherichia coli cells co-expressing poly-γ-glutamic acid (γ-PGA) synthetase and glutamate racemase for differential yielding of γ-PGA. Microb. Biotechnol. 2013, 6, 675–684. [Google Scholar] [CrossRef]
- Sha, Y.; Sun, T.; Qiu, Y.; Zhu, Y.; Zhan, Y.; Zhang, Y.; Xu, Z.; Li, S.; Feng, X.; Xu, H. Investigation of Glutamate Dependence Mechanism for Poly-γ-glutamic Acid Production in Bacillus subtilis on the Basis of Transcriptome Analysis. J. Agric. Food Chem. 2019, 67, 6263–6274. [Google Scholar] [CrossRef] [PubMed]
- Ashiuchi, M.; Soda, K.; Misono, H. Characterization of yrpC gene product of Bacillus subtilis IFO 3336 as glutamate racemase isozyme. Biosci. Biotechnol. Biochem. 1999, 63, 792–798. [Google Scholar] [CrossRef]
- Bormann, E.; Eikmanns, B.; Sahm, H. Molecular analysis of the Corynebacterium glutamicum gdh gene encoding glutamate dehydrogenase. Mol. Microbiol. 1992, 6, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Urushibata, Y.; Tokuyama, S.; Tahara, Y. Difference in transcription levels of cap genes for y-polyglutamic acid production between Bacillus subtilis IFO 16449 and Marburg 168. J. Biosci. Bioeng. 2002, 93, 252–254. [Google Scholar] [CrossRef]
- Candela, T.; Fouet, A. Poly-gamma-glutamate in bacteria. Mol. Microbiol. 2006, 60, 1091–1098. [Google Scholar] [CrossRef]
- Yamashiro, D.; Yoshioka, M.; Ashiuchi, M. Bacillus subtilis pgsE (formerly ywtC) stimulates poly-γ-glutamate production in the presence of Zinc. Biotechnol. Bioeng. 2011, 108, 226–230. [Google Scholar] [CrossRef]
- Tran, L.S.; Nagai, T.; Itoh, Y. Divergent structure of the ComQXPA quorumsensing components: Molecular basis of strain-specific communication mechanism in Bacillus subtilis. Mol. Microbiol. 2000, 37, 1159–1171. [Google Scholar] [CrossRef]
- Do, T.H.; Suzuki, Y.; Abe, N.; Kaneko, J.; Itoh, Y.; Kimura, K. Mutations suppressing the loss of DegQ function in Bacillus subtilis (natto) poly-γ-glutamate synthesis. Appl. Environ. Microbiol. 2011, 77, 8249–8258. [Google Scholar] [CrossRef] [Green Version]
- Osera, C.; Amati, G.; Calvio, C.; Galizzi, A. SwrAA activates poly-gamma-glutamate synthesis in addition to swarming in Bacillus subtilis. Microbiology 2009, 155, 2282–2287. [Google Scholar] [CrossRef] [Green Version]
- Ohsawa, T.; Tsukahara, K.; Ogura, M. Bacillus subtilis response regulator DegU is a direct activator of pgsB transcription involved in gamma-poly-glutamic acid synthesis. Biosci. Biotechnol. Biochem. 2009, 73, 2096–2102. [Google Scholar] [CrossRef] [PubMed]
- Stanley, N.R.; Lazazzera, B.A. Defining the genetic differences between wild and domestic strains of Bacillus subtilis that affect poly-γ-DL-glutamic acid production and biofilm formation. Mol. Microbiol. 2005, 57, 1143–1158. [Google Scholar] [CrossRef] [PubMed]
- Ashiuchi, M.; Kamei, T.; Misono, H. Poly-γ-glutamate synthetase of Bacillus subtilis. J. Mol. Catal. B Enzym. 2003, 23, 101–106. [Google Scholar] [CrossRef]
- Feng, J.; Gao, W.; Gu, Y.; Zhang, W.; Cao, M.; Song, C.; Zhang, P.; Sun, M.; Yang, C.; Wang, S. Functions of poly-gamma-glutamic acid (γ-PGA)degradation genesin γ-PGA synthesis and cell morphology maintenance. Appl. Microbiol. Biotechnol. 2014, 98, 6397–6407. [Google Scholar] [CrossRef]
- Mitsui, N.; Murasawa, H.; Sekiguchi, J. Disruption of the cell wall lytic enzyme Cwlo affects the amount and molecular size of poly-γ-glutamic acid produced by Bacillus subtilis (natto). J. Gen. Appl. Microbiol. 2011, 57, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Haque, S.; Niwas, R.; Srivastava, A.; Pasupuleti, M.; Tripathi, C.K.M. Strategies for fermentation medium optimization: An in-depth review. Front. Microbiol. 2017, 7, 2087. [Google Scholar] [CrossRef]
- Gao, W.; Zhang, Z.; Feng, J.; Dang, Y.; Quan, Y.; Gu, Y.; Wang, S.; Song, C. Effects of MreB paralogs on poly-γ-glutamic acid synthesis and cell morphology in Bacillus amyloliquefaciens. FEMS Microbiol. Lett. 2016, 363, fnw187. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Du, Y.; Xu, G.; Zhang, H.; Zhu, F.; Huang, L.; Xu, Z. High yield and cost-effective production of poly (gammaglutamic acid) with Bacillus subtilis. Eng. Life Sci. 2011, 11, 291–297. [Google Scholar] [CrossRef]
- Xu, Z.; Feng, X.; Zhang, D.; Tang, B.; Lei, P.; Liang, J.; Xu, H. Enhanced poly (γ-glutamic acid) fermentation by Bacillus subtilis NX-2 immobilized in an aerobic plant fibrous-bed bioreactor. Bioresour. Technol. 2014, 155, 8–14. [Google Scholar] [CrossRef]
- Zhang, D.; Feng, X.; Zhou, Z.; Zhang, Y.; Xu, H. Economical production of poly (γ-glutamic acid) using untreated cane molasses and monosodium glutamate waste liquor by Bacillus subtilis NX-2. Bioresour. Technol. 2012, 114, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Ju, W.T.; Song, Y.S.; Jung, W.J.; Park, R.D. Enhanced production of poly-γ glutamic acid by a newly-isolated Bacillus subtilis. Biotechnol. Lett. 2014, 36, 2319–2324. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Cai, J.; Zheng, Q.; Zhu, X.; Cen, P.; Xu, Z. A novel approach for poly-gamma-glutamic acid production using xylose and corncob fibres hydrolysate in Bacillus subtillis. HB-1. J. Chem. Technol. Biotechnol. 2014, 89, 616–622. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, Y.; Wei, X.; Hu, Z.; Zhu, F.; Xu, L.; Luo, M. Production of ultra-high molecular weight poly-γ-glutamic acid with Bacillus licheniformis P-104 and characterization of its flocculation properties. Appl. Biochem. Biotechnol. 2013, 170, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Scoffone, V.; Dondi, D.; Biino, G.; Borghese, G.; Pasini, D.; Galizzi, A.; Calvio, C. Knockout of pgds and ggt genes improves y-PGA yield in B. subtilis. Biotechnol. Bioeng. 2013, 110, 2006–2012. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Qi, G.; Ji, Z.; Zhang, S.; Liu, J.; Ma, X.; Chen, S. Expression of glr gene encoding glutamate racemase in Bacillus licheniformis WX-02 and its regulatory effects on synthesis of poly-gamma-glutamic acid. Biotechnol. Lett. 2011, 33, 1837–1840. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Fu, J.; Wei, X.; Ji, Z.; Ma, X.; Qi, G.; Chen, S. Enhanced expression of pgdS gene for high production of poly-γ-glutamic acid with lower molecular weight in Bacillus licheniformis WX-02. J. Chem. Technol. Biotechnol. 2014, 89, 1825–1832. [Google Scholar] [CrossRef]
- Zhan, Y.; Sheng, B.; Wang, H.; Shi, J.; Cai, D.; Yi, L.; Yang, S.; Wen, Z.; Ma, X.; Chen, S. Rewiring glycerol metabolism for enhanced production of poly-γ-glutamic acid in Bacillus licheniformis. Biotechnol. Biofuels 2018, 11, 306. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Gu, Y.; Quan, Y.; Cao, M.; Gao, W.; Zhang, W.; Wang, S.; Yang, C.; Song, C. Improved poly-γ-glutamic acid production in Bacillus amyloliquefaciens bymodular pathway engineering. Metab. Eng. 2015, 32, 106–115. [Google Scholar] [CrossRef]
- Zhang, W.; He, Y.; Gao, W.; Feng, J.; Cao, M.; Yang, C.; Song, C.; Wang, S. Deletion of genes involved in glutamate metabolism to improve poly-gamma-glutamic acid production in Bacillus amyloliquefaciens LL3. J. Ind. Microbiol. Biotechnol. 2015, 42, 297–305. [Google Scholar] [CrossRef]
- Gao, W.; He, Y.; Zhang, F.; Zhao, F.; Huang, C.; Zhang, Y.; Zhao, Q.; Wang, S.; Yang, C. Metabolic engineering of Bacillus amyloliquefaciens LL3 for enhanced poly-γ-glutamic acid synthesis. Microb. Biotechnol. 2019, 12, 932–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, C.M.; Wang, J.P.; Lo, S.C.; Chan, W.C.; Lin, M.L.Y. Chromosomal integration of a synthetic expression control sequence achieves poly-y-glutamate production in a Bacillus subtilis strain. Biotechnol. Prog. 2010, 26, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Tran, L.S.; Uchida, L.; Itoh, Y. Characterization of Bacillus subtilis γ-glutamyltransferase and its involvement in the degradation of capsule poly-γ-glutamate. Microbiology 2004, 150, 4115–4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Zhang, T.; Mu, W.; Miao, M.; Jiang, B. Intracellular synthesis of glutamic acid in Bacillus methylotrophicus SK19.001, a glutamate-independent poly (γ-glutamic acid)-producing strain. J. Sci. Food Agric. 2016, 96, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Kang, J.W.; Kang, A.; Lee, T.S. Redirecting metabolic flux via combinatorial multiplex crispri-mediated repression for isopentenol production in Escherichia coli. ACS Synth Biol. 2019, 8, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Liu, F.; Zhang, W.; Quan, Y.; Dang, Y.; Feng, J.; Gu, Y.; Wang, S.; Song, C.J.; Yang, C. Mutations in genes encoding antibiotic substances increase the synthesis of poly-γ-glutamic acid in Bacillus amyloliquefaciens LI.3. Microbiologyopen 2017, 6, e00398. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; He, P.; Wan, N.; Cai, D.; Hu, S.; Chen, Y.; Li, S. 13C-Metabolic flux analysis reveals the metabolic flux redistribution for enhanced production of poly-γ-glutamic acid in dlt over-expressed Bacillus Licheniformis. Front. Microbiol. 2018, 10, 105. [Google Scholar] [CrossRef]
- Jiang, H.; Shang, L.; Yoon, S.H.; Lee, S.Y.; Yu, Z. Optimal production of poly-gamma-glutamic acid by metabolically engineered Escherichia coli. Biotechnol. Lett. 2006, 28, 1241–1246. [Google Scholar] [CrossRef]
- Yao, W.; Meng, G.; Zhang, W.; Chen, X.; Yin, R. China National Intellectual Property Administration. Vol CN103146630 (A), 2013. Available online: https://english.cnipa.gov.cn/col/col1584/index.html (accessed on 24 January 2022).
- Cao, M.; Song, C.; Jin, Y.; Liu, L.; Liu, J.; Xie, H.; Guo, W.; Wang, S. Synthesis of poly (γ-glutamic acid) and heterologous expression of pgsBCA genes. J. Mol. Catal. B Enzym. 2010, 67, 111–116. [Google Scholar] [CrossRef]
- Lin, B.; Li, Z.; Zhang, H.; Wu, J.; Luo, M. Cloning and expression of the γ-polyglutamic acid synthetase gene pgsBCA in Bacillus subtilis WB600. BioMed Res. Int. 2016, 2016, 3073949. [Google Scholar] [CrossRef] [Green Version]
- Moraes, L.P.; Alegre, R.M.; Brito, P.N. Optimisation of poly (γ-glutamic acid) production by Bacillus velezensis NRRL B-23189 in liquid fermentation with molasses as the carbon source without addition of glutamic acid. Int. Rev. Chem. Eng. 2012, 4, 618–623. [Google Scholar]
- Tork, S.E.; Aly, M.M.; Alakilli, S.Y.; Al-Seeni, M.N. Purification and characterization of gamma poly glutamic acid from newly Bacillus licheniformis NRC20. Int. J. Biol. Macromol. 2015, 74, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Hwang, J.S.; Kim, D.H.; Lee, S.; Joe, M.H. A newly isolated bacillus siamensis sb1001 for mass production of poly-γ-glutamic acid. Process. Biochem. 2019, 92, 164–173. [Google Scholar] [CrossRef]
- Kongklom, N.; Luo, H.; Shi, Z.; Pechyen, C.; Chisti, Y.; Sirisansaneeyakul, S. Production of poly-gamma-glutamic acid by glutamic acid-independent Bacillus licheniformis TISTR 1010 using different feeding strategies. Biochem. Eng. J. 2015, 100, 67–75. [Google Scholar] [CrossRef]
- Da Silva, S.B.; Cantarelli, V.V.; Ayub, M.A.Z. Production and optimization of poly-γ-glutamic acid by Bacillus subtilis BL53 isolated from the Amazonian environment. Bioprocess. Biosyst. Eng. 2014, 37, 469–479. [Google Scholar] [CrossRef]
- Lim, S.M.; Kim, J.I.; Shim, J.Y.; Imm, B.Y.; Sung, M.H.; Imm, J.Y. Effect of poly-γ-glutamic acids (PGA) on oil uptake and sensory quality in doughnuts. Food Sci. Biotechnol. 2012, 21, 247–252. [Google Scholar] [CrossRef]
- Du, G.; Yang, G.; Qu, Y.; Chen, J.; Lun, S. Effects of glycerol on the production of poly (γ-glutamic acid) by Bacillus licheniformis. Process. Biochem. 2005, 40, 2143–2147. [Google Scholar] [CrossRef]
- Shih, I.L.; Van, Y.T.; Chang, Y.N. Application of statistical experimental methods to optimize production of poly (γ-glutamic acid) by Bacillus licheniformis CCRC 12826. Enzyme Microb. Technol. 2002, 31, 213–220. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, D.J.; Jia, J.; Yang, H.Q. Fishmeal wastewater as a low-cost nitrogen source for γ-polyglutamic acid production using Bacillus subtilis. Waste Biomass. Valorization 2019, 10, 789–795. [Google Scholar] [CrossRef]
- Chen, X.; Chen, S.; Sun, M.; Yu, Z. High yield of poly-y-glutamic acid from Bacillus subtilis by solid-state fermentation using swine manure as the basis of a solid substrate. Bioresour. Technol. 2005, 96, 1872–1879. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.M.; Jang, W.J.; Park, H.D.; Kim, Y.O.; Kim, C.H.; Kong, I.S. Efficient production of poly γ-d-glutamic acid from the bloom-forming green macroalgae, Ulva sp., by Bacillus sp. SJ-10. Biotechnol. Bioeng. 2019, 116, 1594–1603. [Google Scholar] [CrossRef]
- Altun, M. Bioproduction of γ-Poly (glutamic acid) using feather hydrolysate as a fermentation substrate. Trak. Univ. J. Nat. Sci. 2019, 20, 27–34. [Google Scholar] [CrossRef]
- Scheel, R.A.; Fusi, A.D.; Min, B.C.; Thomas, C.M.; Ramarao, B.V.; Nomura, C.T. Increased production of the value-added biopolymers poly (γ-3-hydroxyalkanoate) and poly (γ-glutamic acid) from hydrolyzed paper recycling waste fines. Front. Bioeng. Biotechnol. 2019, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wu, D.; Ren, H. Economical production of agricultural γ-polyglutamic acid using industrial wastes by Bacillus subtilis. Biochem. Eng. J. 2019, 146, 117–123. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, Z.; Xu, H.; Feng, X.; Li, S.; Cai, H.; Wei, Y.; Ouyang, P. Improvement of poly (gamma-glutamic acid) biosynthesis and quantitative metabolic flux analysis of a two-stage strategy for agitation speed control in the culture of Bacillus subtilis NX-2. Biotechnol. Bioproc. Eng. 2011, 16, 1144–1151. [Google Scholar] [CrossRef]
- Zhang, D.; Feng, X.; Li, S.; Chen, F.; Xu, H. Effects of oxygen vectors on the synthesis and molecular weight of poly (gamma-glutamic acid) and the metabolic characterization of Bacillus subtilis NX-2. Process. Biochem. 2012, 47, 2103–2109. [Google Scholar] [CrossRef]
- Jeong, J.H.; Kim, J.N.; Wee, Y.J.; Ryu, H.W. The statistically optimized production of poly (γ-glutamic acid) by batch fermentation of a newly isolated Bacillus subtilis RKY3. Bioresour. Technol. 2010, 101, 4533–4539. [Google Scholar] [CrossRef]
- Goto, A.; Kunioka, M. Biosynthesis and hydrolysis of poly (gamma-glutamic acid) from Bacillus subtilis IF03335. Biosci. Biotechnol. Biochem. 1992, 56, 1031–1035. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.D.; Mack, M.; Contiero, J. Glycerol: A promising and abundant carbon source for industrial microbiology. Biotechnol. Adv. 2009, 27, 30–39. [Google Scholar] [CrossRef]
- Zeng, W.; Liang, Z.; Li, Z.; Bian, Y.; Li, Z.; Tang, Z.; Chen, G. Regulation of poly-γ-glutamic acid production in Bacillus subtilis GXA-28 by potassium. J. Taiwan Inst. Chem. Eng. 2016, 61, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Cromwick, A.M.; Birrer, G.A.; Gross, R.A. Effects of pH and aeration on γ-poly (glutamic acid) formation by Bacillus licheniformis in controlled batch fermentor cultures. Biotechnol. Bioeng. 1996, 50, 222–227. [Google Scholar] [CrossRef]
- Yoon, S.H.; Hwan Do, J.; Yup Lee, S.; Nam Chang, H. Production of poly-γ-glutamic acid by fed-batch culture of Bacillus licheniformis. Biotechnol. Lett. 2000, 22, 585–588. [Google Scholar] [CrossRef]
- Wang, J.; Yuan, H.; Wei, X.; Chen, J.; Chen, S. Enhancement of poly-γ-glutamic acid production by alkaline pH stress treatment in Bacillus licheniformis WX-02. J. Chem. Technol. Biotechnol. 2015, 91, 2399–2403. [Google Scholar] [CrossRef]
- Wu, Q.; Xu, H.; Ying, H.J.; Ouyang, P.K. Kinetic analysis and pH-shift control strategy for poly (gamma-glutamic acid) production with Bacillus subtilis CGMCC 0833. Biochem. Eng. J. 2010, 50, 24–28. [Google Scholar] [CrossRef]
- Feng, J.; Gu, Y.; Quan, Y.; Gao, W.; Dang, Y.; Cao, M.; Lu, X.; Wang, Y.; Song, C.; Wang, S. Construction of energy-conserving sucrose utilization pathways for improvingpoly-y-glutamic acid production in Bacillus amyloliquefaciens. Microb. Cell Factoriesi. 2017, 16, 98. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.; Chen, G.; Guo, Y.; Zhang, B.; Dong, M.; Wu, Y.; Wang, J.; Che, Z.; Liang, Z. Production of poly-γ-glutamic acid by a thermotolerant glutamate-independent strain and comparative analysis of the glutamate dependent difference. AMB Express 2017, 7, 213. [Google Scholar] [CrossRef] [Green Version]
- Zeng, W.; Chen, G.; Wu, Y.; Dong, M.; Zhang, B.; Liang, Z. Nonsterilized fermentative production of poly-γ-glutamic acid from cassava starch and corn steep powder by a thermophilic Bacillus subtilis. J. Chem. Technol. Biotechnol. 2018, 93, 2917–2924. [Google Scholar] [CrossRef]
- Meissner, L.; Kauffmann, K.; Wengeler, T.; Mitsunaga, H.; Fukusaki, E.; Büchs, J. Influence of nitrogen source and pH value on undesired poly (γ-glutamic acid) formation of a protease producing Bacillus licheniformis strain. J. Ind. Microbiol. Biotechnol. 2015, 42, 1203–1215. [Google Scholar] [CrossRef]
- Hirasawa, T.; Shimizu, H. Recent advances in amino acid production by microbial cells. Curr. Opin. Biotechnol. 2016, 42, 133–146. [Google Scholar] [CrossRef]
- Jose, A.A.; Anusree, G.; Pandey, A.; Binod, P. Production optimization of poly-γ-glutamic acid by Bacillus amyloliquefaciens under solid-state fermentation using soy hull as substrate. Indian J. Biotechnol. 2018, 17, 44–49. [Google Scholar]
- Bajaj, I.B.; Singhal, R.S. Flocculation properties of poly (γ-glutamic acid) produced from Bacillus subtilis isolate. Food Bioprocess. Technol. 2011, 4, 745–752. [Google Scholar] [CrossRef]
- Do, J.H.; Chang, H.N.; Lee, S.Y. Efficient recovery of gamma-poly (glutamic acid) from highly viscous culture broth. Biotechnol. Bioeng. 2001, 76, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Pal, P. Fermentative production of poly (γ-glutamic acid) from renewable carbon source and downstream purification through a continuous membrane-integrated hybrid process. Bioresour. Technol. 2015, 177, 141–148. [Google Scholar] [CrossRef]
- McLean, R.J.; Beauchemin, D.; Clapham, L.; Beveridge, T.J. Metal-binding characteristics of the gamma-glutamyl capsular polymer of Bacillus licheniformis ATCC 9945. Appl. Environ. Microbiol. 1990, 56, 3671–3677. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Liu, H.; Wang, L.; Zhang, Y.; Tian, H.; Ma, X. Effect of poly-γ-glutamic acid on the stability of set yoghurts. J. Food Sci. Technol. 2018, 55, 4634–4641. [Google Scholar] [CrossRef]
- Siaterlis, A.; Deepika, G.; Charalampopoulos, D. Effect of culture medium and cryoprotectants on the growth and survival of probiotic lactobacilli during freeze drying. Lett. Appl. Microbiol. 2009, 48, 295–301. [Google Scholar] [CrossRef]
- Tanimoto, H.; Fox, T.; Eagles, J.; Satoh, H.; Nozawa, H.; Okiyama, A.; Morinaga, Y.; Fairweather-Tait, S. Acute effect of poly-gamma-glutamic acid on calcium absorption in post-menopausal women. J. Am. Coll. Nutr. 2007, 26, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chen, S.; Zhang, J.; Sun, M.; Liu, Z.; Yu, Z. Co-producing lipopeptides and poly-gamma-glutamic acid by solid-state fermentation of Bacillus subtilis using soybean and sweet potato residues and its biocontrol and fertilizer synergistic effects. Bioresour. Technol. 2008, 99, 3318–3323. [Google Scholar] [CrossRef]
- Lee, W.; Kim, M.; Lee, S.H.; Jung, H.G.; Oh, J.W. Prophylactic efficacy of orally administered Bacillus poly-γ-glutamic acid, a non-LPS TLR4 ligand, against norovirus infection in mice. Sci. Rep. 2018, 8, 8667. [Google Scholar] [CrossRef]
- Pang, X.; Lei, P.; Feng, X.; Xu, Z.; Xu, H.; Liu, K. Poly-γ-glutamic acid, a bio-chelator, alleviates the toxicity of Cd and Pb in the soil and promotes the establishment of healthy Cucumis sativus L. seedling. Environ. Sci. Pollut. Res. 2018, 25, 19975–19988. [Google Scholar] [CrossRef]
- Singer, J.W. Paclitaxel poliglumex (XYOTAX (TM), CT-2103): A macromolecular taxane. J. Control. Release 2005, 109, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.T.; Kwak, C.; Lee, W.S.; Kim, J.; Jeong, J.; Sung, M.H.; Yang, J.; Poo, H. Poly-γ-Glutamic Acid Complexed with Alum Induces Cross-Protective Immunity of Pandemic H1N1 Vaccine. Front. Immunol. 2019, 10, 1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csikós, Z.; Fazekas, E.; Rózsa, D.; Borbély, J.; Kerekes, K. Crosslinked poly-γ-glutamic acid based nanosystem for drug delivery. J. Drug Deliv. Sci. Technol. 2018, 48, 478–489. [Google Scholar] [CrossRef]
- Shih, I.L.; Van, Y.T.; Yeh, L.C.; Lin, H.G.; Chang, Y.N. Production of a biopolymer flocculant from Bacillus licheniformis and its flocculation properties. Bioresour. Technol. 2001, 78, 267–272. [Google Scholar] [CrossRef]




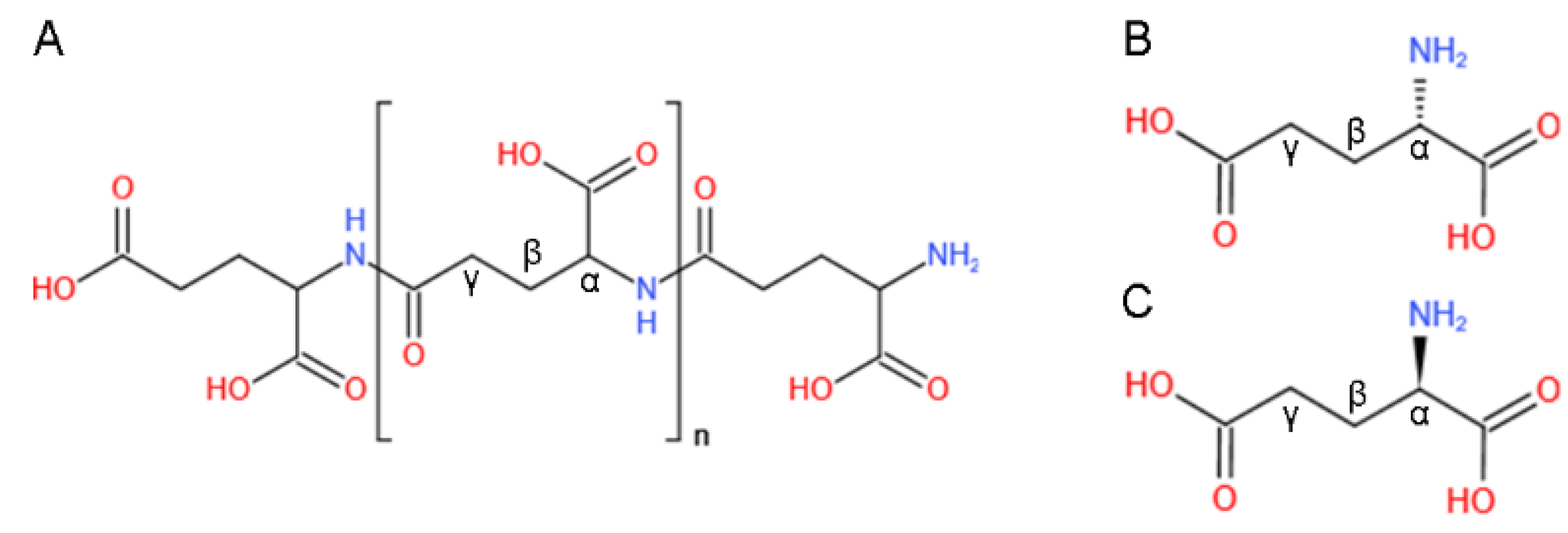{kind=link}
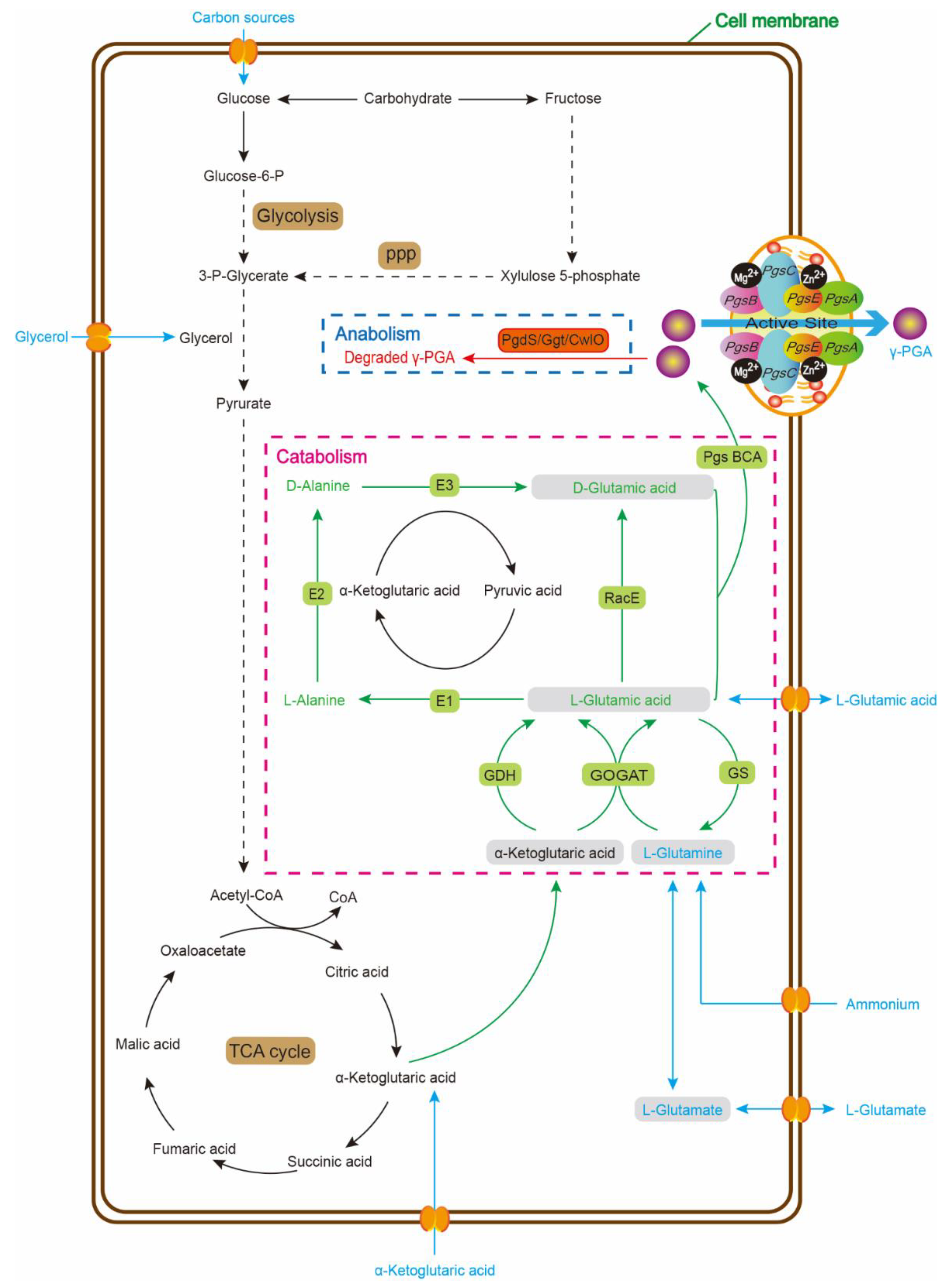{kind=link}
{kind=link}
| Strains | Source | Main Medium Components | Cultural Conditions | Final Yield (g/L) | Ref. |
|---|---|---|---|---|---|
| B. subtilis ZJU-7 | Isolated from fermented bean curd | Glucose, L-glutamic, yeast extract, NaCl, Ca2+, Mg2+, Mn2+ | Bioreactor, pH 6.5, 37 °C | 101.1 | [72] |
| B. subtilis NX-2 | Isolated from soil samples | Glucose, glutamic, (NH4)2SO4, yeast extract, K2HPO4, Mg2+, Mn2+ | APFB (aerobic plant fibrous-bed bioreactor) immobilized cell fermentation, pH 7.0, 32 °C | 71.21 | [73] |
| Isolated from soil samples | Cane molasses, monosodium glutamate liquid waste | Bioreactor, pH 7.0, 32 °C | 52.1 | [74] | |
| B. subtilis MJ80 | Isolated from soil samples | Glutamic acid, starch, urea, citric acid, glycerol, NaCl, K2HPO4, Mg2+, Mn2+ | Fermenter for immobilized cell fermentation, pH 7.0, 37 °C | 68.7 | [75] |
| B. subtilis HB-1 | Isolated from soil samples | Glutamate, xylose, corncob fibers hydrolysate, yeast extract, NaCl | Bioreactor, pH 6.5, 37 °C | 28.15 | [76] |
| B. methyotrophicus SK 19.001 | Isolated from soil samples | Glucose, yeast extract, K2HPO4, Mg2+, Mn2+ | Flask, pH 7.2, 37 °C | 35.34 | [27] |
| B. licheniformis P-104 | Isolated from Chinese soybean paste | Glucose, glutamate, citric acid, (NH4)2SO4, K2HPO4, Mg2+, Mn2+ | Bioreactor, pH 7.0, 37 °C | 41.6 | [77] |
| Strains | Engineering Methods | Main Medium Components | Final Yield (g/L) | Ref. |
|---|---|---|---|---|
| B. subtilis ISW1214 | Carrying the plasmid of γ-PGA synthetic system | Sucrose, xylose, NaCl, NaHPO4, KH2PO4, Mg2+ | 9.0 | [54] |
| B. subtilis PB5249 | Deletion of genes (pgdS and ggt) | Glucose, L-glutamic acid, citric acid, NH4Cl, K2HPO4, Mg2+, Mn2, Ca2+, Fe2+ | 40 | [78] |
| B. licheniformis WX-02 | Expression of glr gene for encoding glutamate recemase | Glucose, L-glutamic acid, citric acid, NH4Cl, NaCl, K2HPO4, Mg2+, Mn2+, Ca2+, Zn2+ | 14.38 | [79] |
| Enhanced expression of pgdS gene | Glucose, glutamate, citric acid, NH4Cl, K2HPO4, Mg2+, Mn2, Ca2+, Zn2+ | 20.16 | [80] | |
| Substituted by the native glpFK promoter with the constitutive promoter (P43), ytzE promoter (PytzE), and bacABC operon promoter (PbacA) | Sodium glutamate, citric acid, glycerol, | 17.65 | [81] | |
| Over-expression of glpK, glpX, zwf, and tkt1 promoters | Sodium citrate, glycerol, NaNO3, NH4Cl | 12.83 | [81] | |
| B. amyloliquefaciens | Deletion of gene (epsA-O, sac, lps, pta, pgdS, cwlO, luxS, and rocG), expression of synthetic small synthetic regulator RNAs (repressed the expression of rocG and glnA gene) | Sucrose, (NH4)2SO4, K2HPO4, KH2PO4, Mg2+ | 20.3 | [82] |
| B. amyloliquefaciens LL3 | Double knockout of gene (pgdS and cwlO) | Sucrose, (NH4)2SO4, NaCl, K2HPO4, KH2PO4 | 7.12 | [68] |
| Gene knockout of rocR, rocG, gudB, and odhA | 5.68 | [83] | ||
| Gene knockout of fadR, lysC, aspB, pckA, proAB, rocG, and gudB | Tryptone, xylose, yeast extract, NaCl, ampicillin, chloramphenicol, or tetracycline | 4.84 | [84] | |
| Enhancing NADPH level by inserting a strong promoter PC2up | 6.46 | [84] |
| Strains | Genetic Engineering | Main Medium Components | Final Yield (g/L) | Ref. |
|---|---|---|---|---|
| C. glutamicum ATCC 13869 | Cloning and expressing pgsBCA of B. licheniformis TKPG011 | Glucose, (NH4)2SO4, soy protein hydrolysate, thiamine hydrochloride, KH2PO4, Mg2+, Mn2, Fe2+, Ca2+ | 18 | [92] |
| C. glutamicum ATCC 13032 | Cloning and expressing pgsABC from B. licheniformis NK-03 | Glucose, tryptone, yeast extract | 0.7 | [93] |
| E. coli BL21 | Cloning and overexpressing γ-PGA biosynthesis genes | Glucose, L-glutamic acid, yeast extract, (NH4)2SO4 | 3.7 | [91] |
| E. coli LRP | Expressing pgsBCA and race from B. amyloliquefaciens LL3 | Glucose, yeast extract, NaCl, | 0.7 | [1] |
| E. coli JM 109 | Cloning pgsBCA and racE from both B. licheniformis NK-03 and B. amyloliquefaciens LL3 and co-expression | Glucose, L-glutamic acid, yeast extract, NaCl, (NH4)2SO4, K2HPO4, KH2PO4, Mg2+, | 0.65 | [55] |
| B. subtilis PB5249 | ∆pgdS∆ggt deletion mutants | glucose, L-glutamate, citric acid, NH4Cl, K2HPO4, MgSO4·7H2O, MnSO4·H2O, FeCl3·6H2O, CaCl2·2H2O | 40 | [78] |
| B. subtilis WB600 | Overexpressing pWB980-pgsBCA | Glucose, sodium glutamate, MgSO4, (NH4)2SO4, K2HPO4 | 1.74 | [94] |
| B. subtilis ISW1214 | Overexpressing pWH1520-PxylA-pgsBCA | Sucrose, xylose, NaCl, MgSO4, KH2PO4, NaHPO4 | 9.0 | [54] |
| B. licheniformis WX-02 | Overexpressing pHY300PLK-P43-glr | Sucrose, (NH4)2SO4, MgSO4, KH2PO4, K2HPO4 | 14.38 | [79] |
| Overexpressing pHY300PLK-PpgdS-pgdS | Glucose, sodium glutamate, sodium citrate, NH4Cl, MgSO4, ZnSO4, MnSO4, CaCl2, K2HPO4 | 20.16 | [80] | |
| B. amyloliquefaciens LL3 | ∆pgdS∆cwlO deletion mutants | Sucrose, (NH4)2SO4, MgSO4, KH2PO4, K2HPO4 | 7.12 | [68] |
| ∆rocR∆rocG∆gudB∆odhA deletion mutants | Sucrose, (NH4)2SO4, MgSO4, KH2PO4, K2HPO4 | 5.68 | [83] | |
| B. amyloliquefaciens | ∆cwlO∆epsA-Ovgb deletion mutants | Sucrose, (NH4)2SO4, MgSO4, FeSO4·4H2O, CaCl2·2H2O, MnSO4·4H2O, ZnCl2, KH2PO4, K2HPO4, | 5.12 | [68] |
| Repressed both rocG and glnA genes | Sucrose, (NH4)2SO4, MgSO4, KH2PO4, K2HPO4 | 20.3 | [82] |
| Strains | Cultural Methods | Final Yield (g/L) | Ref. |
|---|---|---|---|
| B. subtilis NX-2 | Using a two-stage strategy for agitation speed control | 40.5 | [109] |
| Adding hydrogen peroxide | 33.9 | [110] | |
| B. subtilis CGMCC 0833 | Applying pH-shift control strategy | 27.7 | [111] |
| B. subtilis F-2-01 | Adding more carbon sources (L-glutamic acid and glycerol) | 45.5 | [43] |
| B. subtilis BL53 | Adding some precursors | 25.2 | [112] |
| B. subtilis C10 | Abundant supply of organic acid | 27.7 | [47] |
| B. subtilis GXA-28 | Addition of KCl | 25.62 | [113] |
| B. licheniformis TISTR 1010 | Different feeding strategies (glucose, citric acid, NH4Cl, NaCl, Mg2+, Mn2, Ca2+, K2HPO4, Tween-80) | 27.5 | [114] |
| B. licheniformis ATCC 9945A | Different feeding strategies (L-glutamic acid, citric acid, glycerol, NH4Cl) | 23.0 | [98] |
| 35.0 | [115] | ||
| 57.5 | [116] | ||
| B. licheniformis WBL-3 | Optimized culture medium (L-glutamic acid, citric acid, glycerol, NH4Cl) | 22.8 | [102] |
| Optimized culture medium (glutamic acid, citric acid, glycerol) | 19.3 | [102] | |
| B. licheniformis P-104 | Optimized culture medium (glutamate, glucose, citric acid, (NH4)2SO4) | 41.6 | [77] |
| B. licheniformis NCIM 2324 | Addition of metabolic precursors (α-ketoglutaric acid) | 35.75 | [18] |
| Optimized culture medium (L-glutamic acid, citric acid, glycerol, (NH4)2SO4, K2HPO4, Mg2+, Mn2+) | 26.12 | [18] | |
| Optimization effecting factors at a time (L-glutamic acid, sugarcane juice, citric acid, NH4Cl) | 35.75 | [18] | |
| B. licheniformis WX-02 | Adding pH stress treatment | 36.26 | [101] |
| B. velezensis NRRL B-23189 | Optimized culture medium (molasses, citric acid, NH4Cl) | 4.82 | [117] |
| B. methylotrophicus SK 19.001 | Optimized culture medium (glucose, citric acid, NH4Cl) | 35.3 | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Hou, L.; Gao, Y.; Tian, Z.; Fan, B.; Wang, F.; Li, S. Recent Advances in Microbial Synthesis of Poly-γ-Glutamic Acid: A Review. Foods 2022, 11, 739. https://doi.org/10.3390/foods11050739
Li D, Hou L, Gao Y, Tian Z, Fan B, Wang F, Li S. Recent Advances in Microbial Synthesis of Poly-γ-Glutamic Acid: A Review. Foods. 2022; 11(5):739. https://doi.org/10.3390/foods11050739
Chicago/Turabian StyleLi, Danfeng, Lizhen Hou, Yaxin Gao, Zhiliang Tian, Bei Fan, Fengzhong Wang, and Shuying Li. 2022. "Recent Advances in Microbial Synthesis of Poly-γ-Glutamic Acid: A Review" Foods 11, no. 5: 739. https://doi.org/10.3390/foods11050739