
Effect of an Environment Friendly Heat and Relative Humidity Approach on γ-Aminobutyric Acid Accumulation in Different Highland Barley Cultivars

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Heat and Relative Humidity (HRH) Treatment
2.3. Determination of Germination Percentage, Kernel Weight, Starch and Protein
2.4. Determination of GABA
2.5. Determination of Free Amino Acid Composition
2.6. Determination of Polyamines
2.7. Assays of GAD, DAO and PAO
2.8. Statistical Analysis
3. Results and Discussion
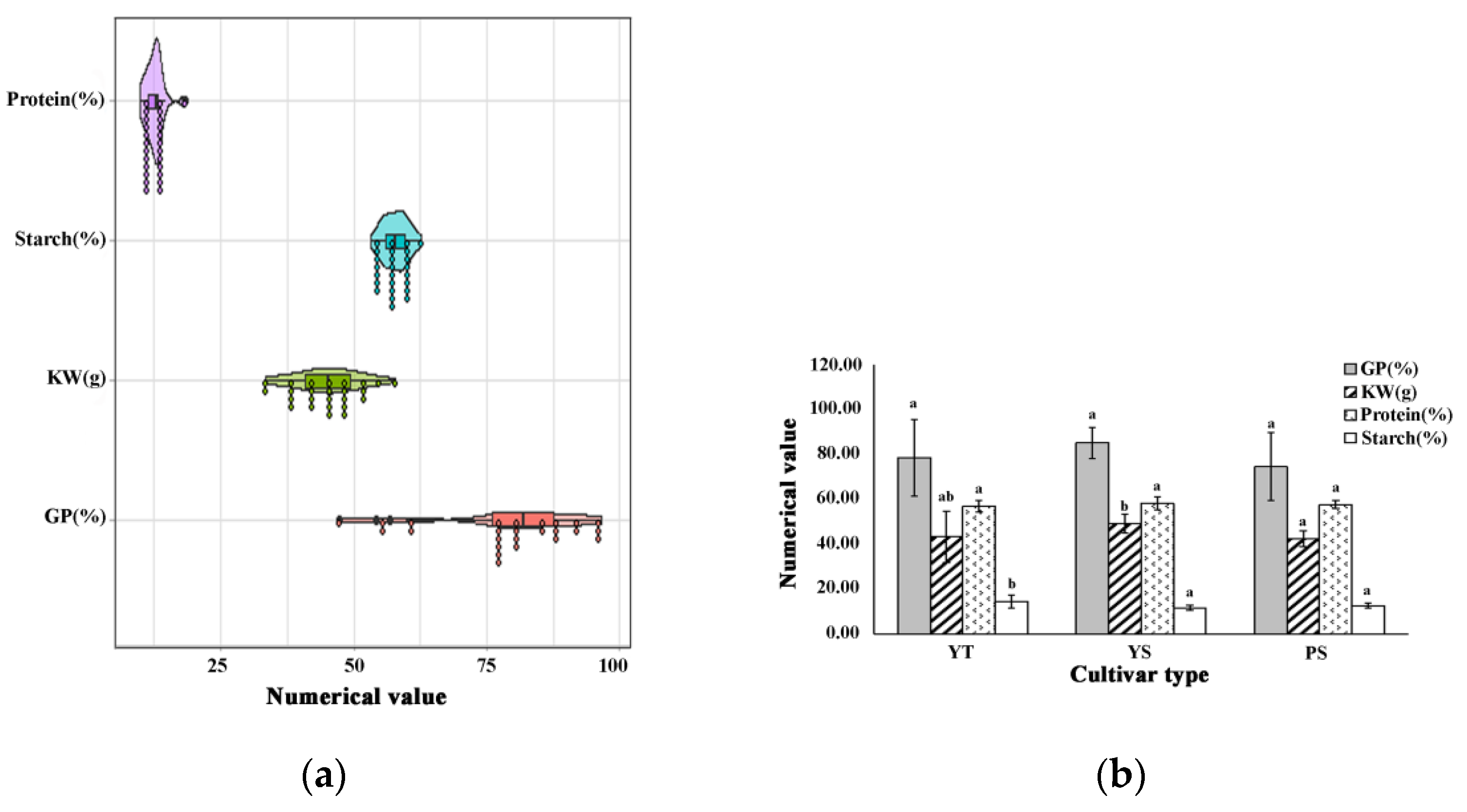
3.1. Cultivar Variation in Germination Percentage, Kernel Weight, Starch and Protein
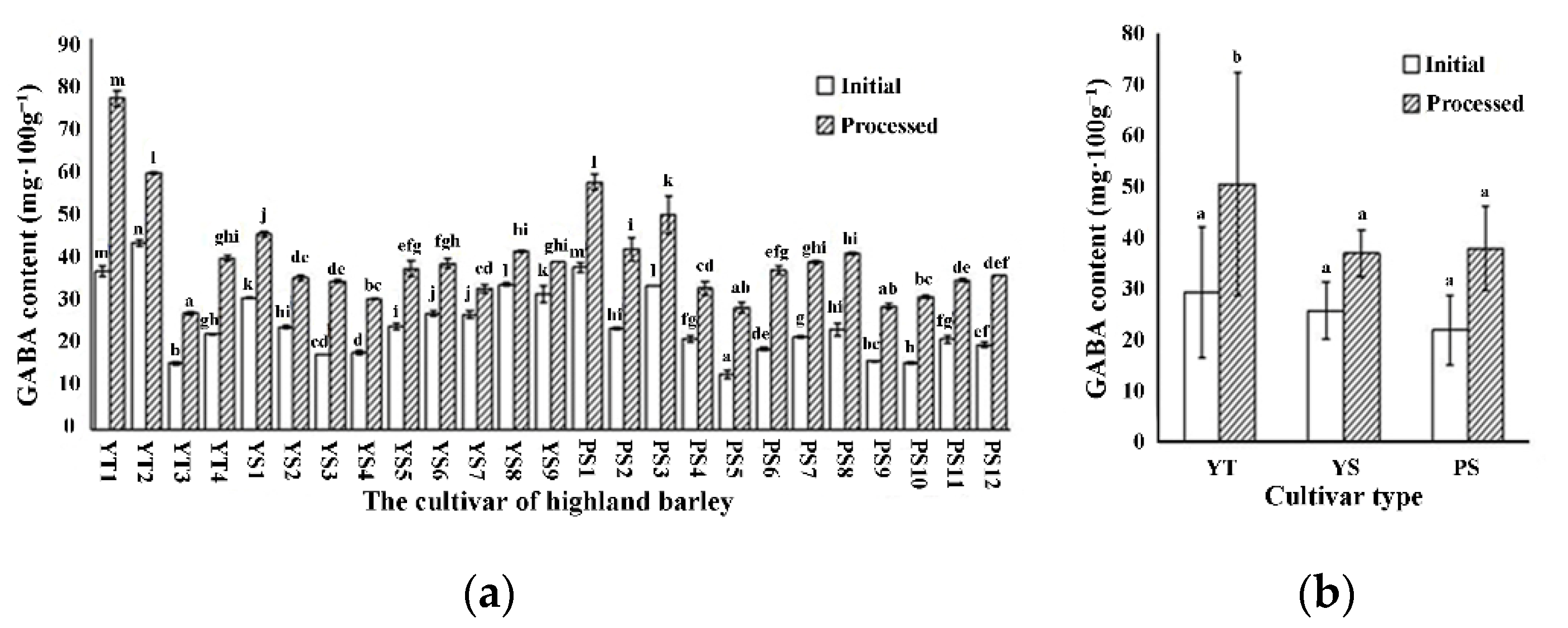
3.2. Effects of Cultivars on GABA Content
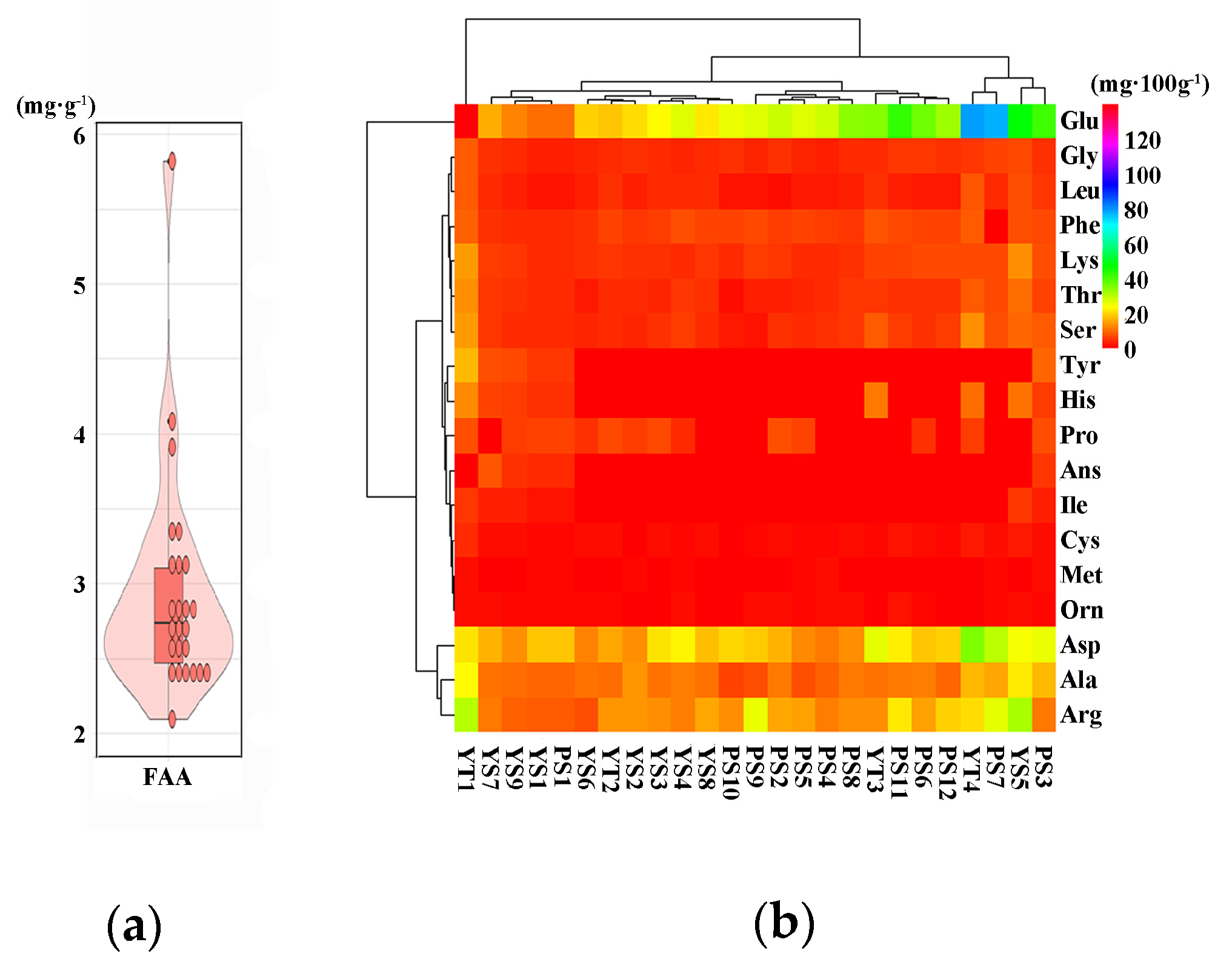
3.3. Cultivar Variation in Free Amino Acids Content and Composition
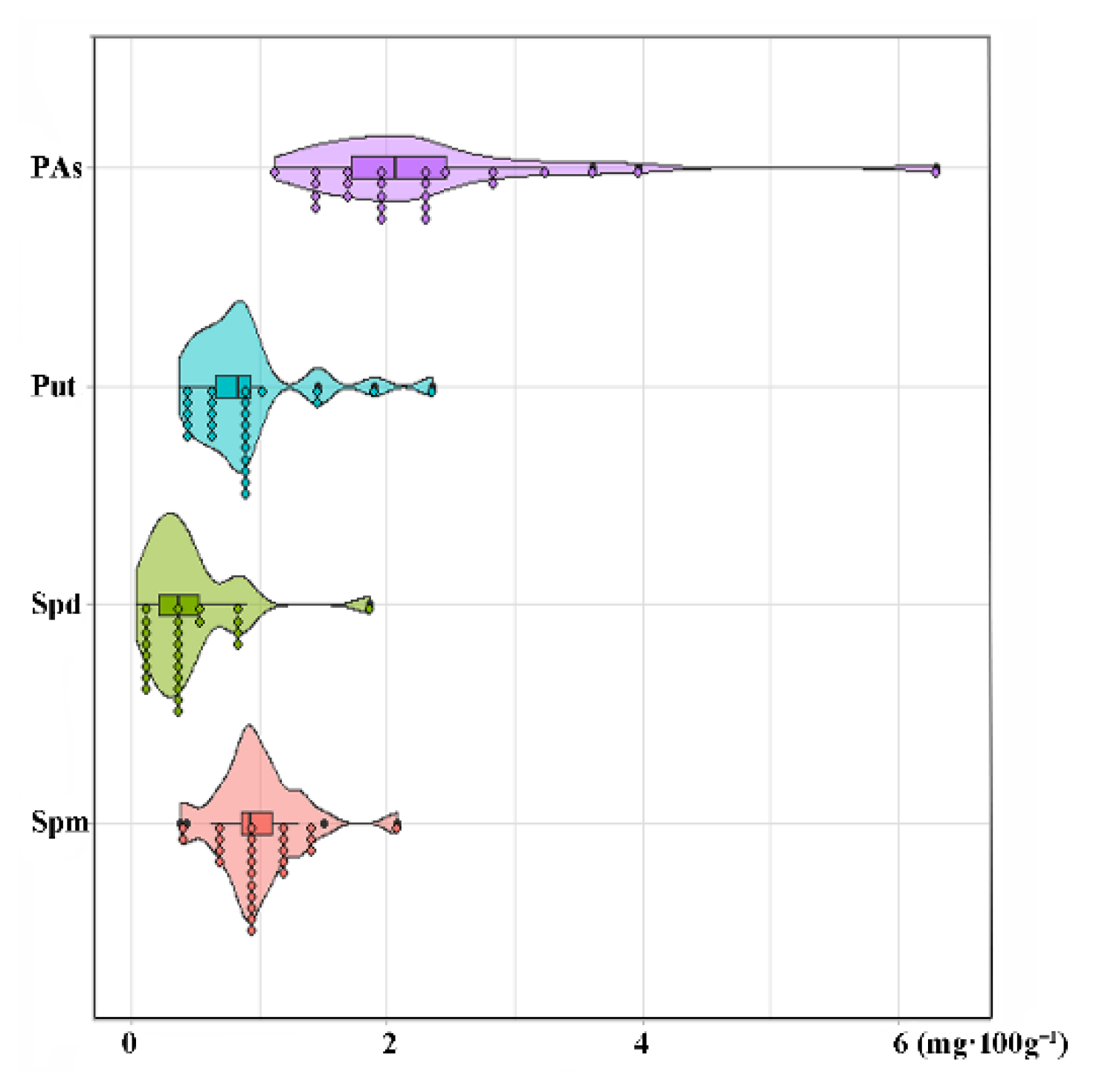
3.4. Cultivar Variation in Polyamine Content and Composition
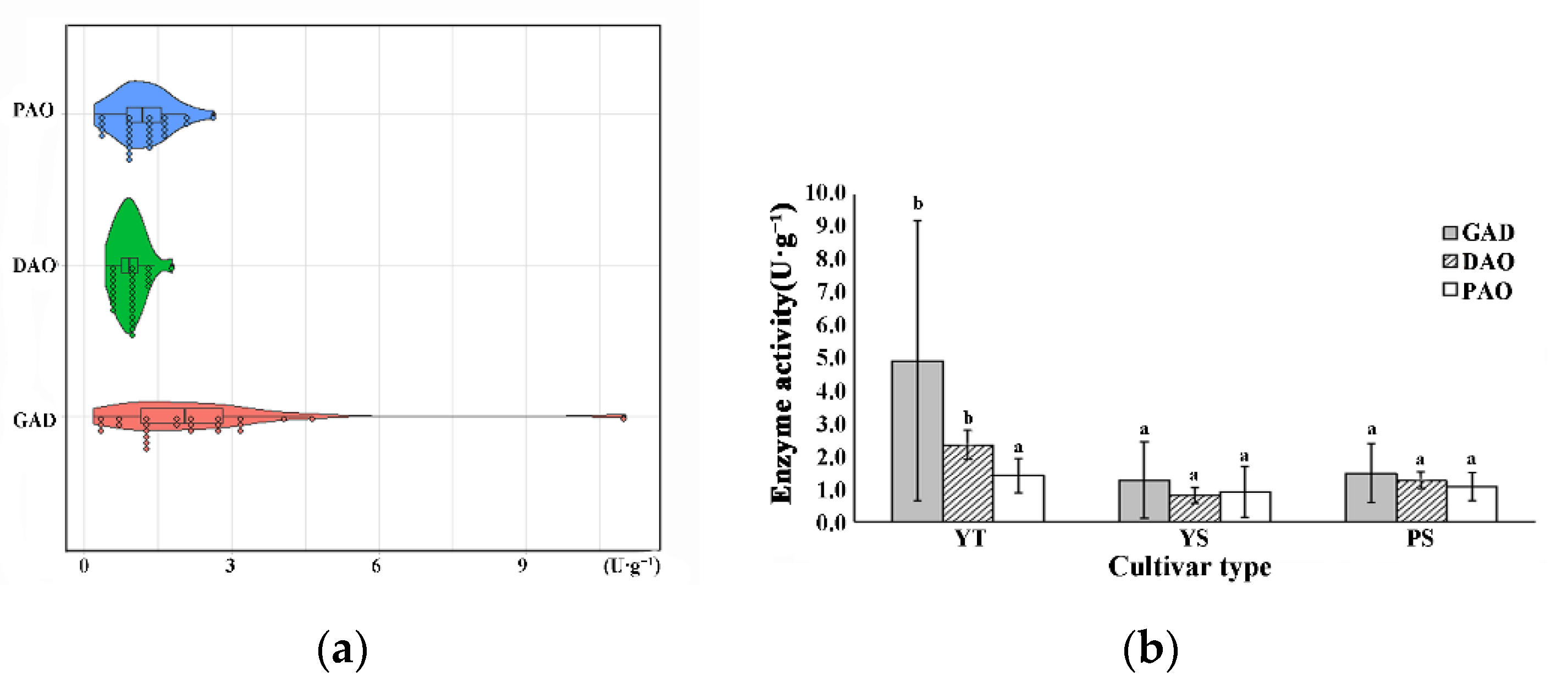
3.5. Cultivar Variation in GAD, DAO and PAO Activity
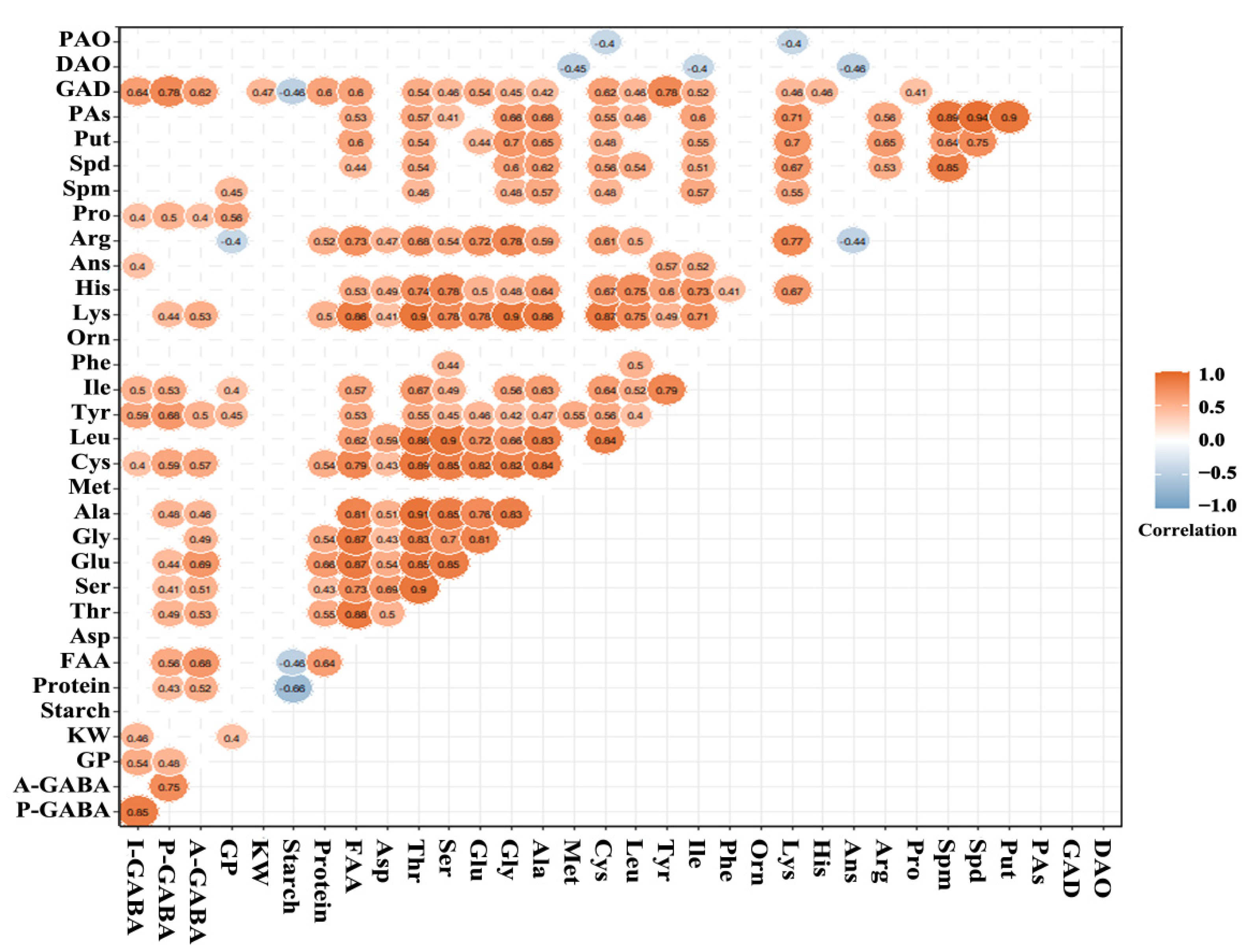
3.6. Correlations in Physical, Chemical and Biochemical Characteristics
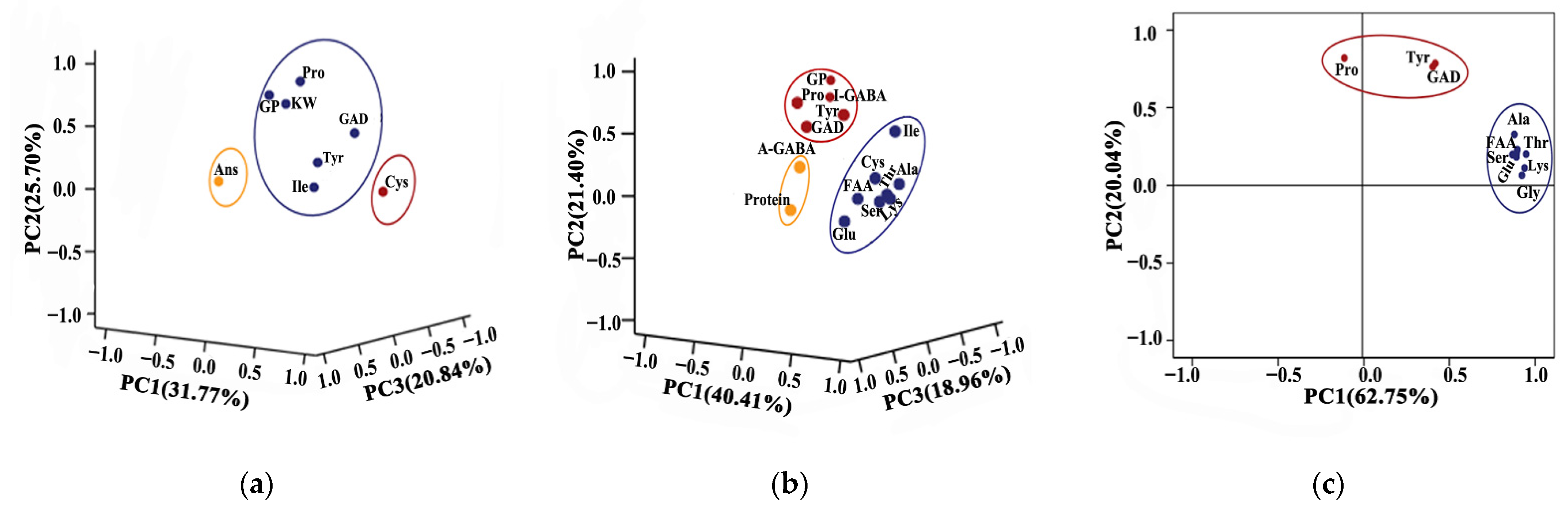
3.7. Principal Component Analysis (PCA)
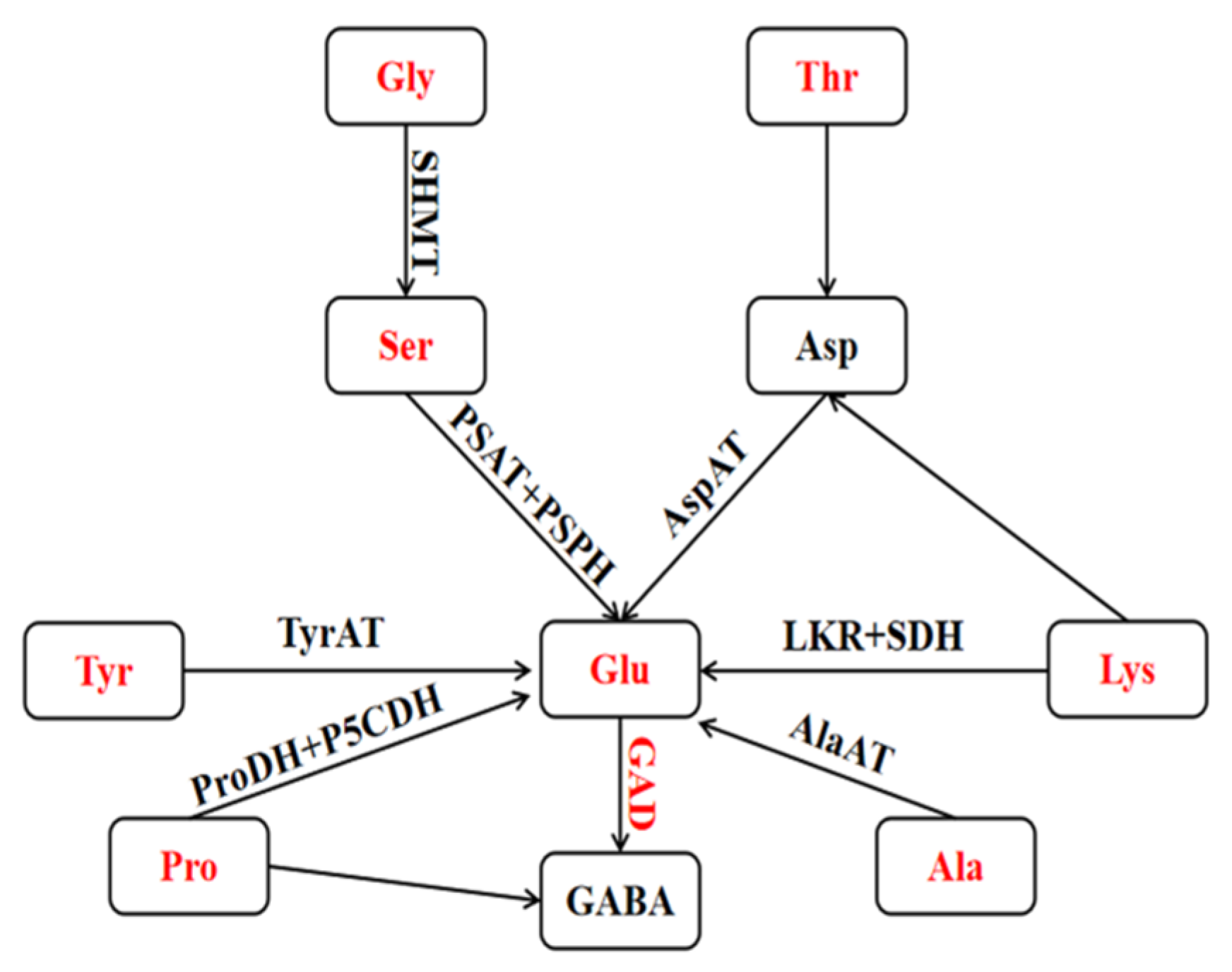
3.8. Mechanism of GABA-Shunt Pathway for GABA Accumulation in Highland Barley under HRH Treatment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, X.J.; Dang, B.; Fan, M.T. Free and bound phenolic compound content and antioxidant activity of different cultivated blue highland barley varieties from the Qinghai-Tibet Plateau. Molecules 2018, 23, 879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Zeng, X.; Lin, B.; Li, Z.; Yuan, H.; Wang, Y.; Zha, S.; Tashi, N. A microsatellite diversity analysis and the development of core-set germplasm in a large hulless barley (Hordeum vulgare L.) collection. BMC Genet. 2017, 18, 102. [Google Scholar] [CrossRef] [Green Version]
- Weng, C.J.; Yen, G.C. Chemopreventive effects of dietary phytochemicals against cancer invasion and metastasis: Phenolic acids, monophenol, polyphenol, and their derivatives. Cancer Treat. Rev. 2012, 38, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.J. Cereal β-glucans in diet and health. J. Cereal Sci. 2007, 46, 230–238. [Google Scholar] [CrossRef]
- Ra, J.; Woo, S.; Jin, H.; Lee, M.J.; Kim, H.Y.; Ham, H.; Chung, I.M.; Seo, W.D. Evaluation of antihypertensive polyphenols of barley (Hordeum vulgare L.) seedlings via their effects on angiotensin-converting enzyme (ACE) inhibition. Appl. Biol. Chem. 2020, 63, 38–46. [Google Scholar] [CrossRef]
- Seifikalhor, M.; Aliniaeifard, S.; Hassani, B.; Niknam, V.; Lastochkina, O. Diverse role of gamma-aminobutyric acid in dynamic plant cell responses. Plant Cell Rep. 2019, 38, 847–867. [Google Scholar] [CrossRef]
- Bettler, B.; Kaupmann, K.; Mosbacher, J.; Gassmann, M. Molecular structure and physiological functions of GABA-B receptors. Physiol. Rev. 2004, 84, 835–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poojary, M.M.; Dellarosa, N.; Roohinejad, S.; Koubaa, M.; Tylewicz, U.; Gómez-Galindo, F.; Saraiva, J.A.; Rosa, M.D.; Barba, F.J. Influence of innovative processing on γ-aminobutyric acid (GABA) contents in plant food materials. Compr. Rev. Food Sci. Food Saf. 2017, 16, 895–905. [Google Scholar] [CrossRef]
- Nikmaram, N.; Dar, B.N.; Roohinejad, S.; Koubaa, M.; Barba, F.J.; Greiner, R.; Johnson, S.K. Recent advances in gamma-aminobutyric acid (GABA) properties in pulses: An overview. J. Sci. Food Agric. 2017, 97, 2681–2689. [Google Scholar] [CrossRef] [Green Version]
- Bai, T.; Jin, Y.L.; Zhu, M.X.; Wang, B. Effect of altitude difference on quality of highland barley varieties. J. Chin. Oils Assoc. 2019, 34, 34–39. [Google Scholar]
- Jannoey, P.; Niamsup, H.; Lumyong, S.; Suzuki, T.; Katayama, T.; Chairote, G. Comparison of gamma-aminobutyric acid production in Thai rice grains. World J. Microbiol. Biotechnol. 2009, 26, 257–263. [Google Scholar] [CrossRef]
- Li, Y.; Bai, Q.; Jin, X.; Wen, H.; Gu, Z. Effects of cultivar and culture conditions on gamma-aminobutyric acid accumulation in germinated fava beans (Vicia faba L.). J. Sci. Food Agric. 2010, 90, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.G.; Hu, Q.P. Changes in γ-aminobutyric acid content and related enzyme activities in Jindou 25 soybean (Glycine max L.) seeds during germination. LWT-Food Sci. Technol. 2014, 55, 341–346. [Google Scholar] [CrossRef]
- Baranzelli, J.; Kringel, D.H.; Colussi, R.; Paiva, F.F.; Aranha, B.C.; Miranda, M.Z.D.; Zavareze, E.D.R.; Dias, A.R.G. Changes in enzymatic activity, technological quality and gamma-aminobutyric acid (GABA) content of wheat flour as affected by germination. LWT-Food Sci. Technol. 2018, 90, 483–490. [Google Scholar] [CrossRef]
- Zeng, Y.W.; Pu, X.Y.; Du, J.; Yang, X.M.; Li, X.; Mandal, M.S.N.; Yang, T.; Yang, J.Z. Molecular mechanism of functional ingredients in barley to combat human chronic diseases. Oxidative Med. Cell. Longev. 2020, 2020, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.C.; Xie, M.X.; Wang, Y.C.; Li, J.Y. Molecular mechanisms underlying gamma-aminobutyric acid (GABA) accumulation in giant embryo rice seeds. J. Agric. Food Chem. 2017, 65, 4883–4889. [Google Scholar] [CrossRef]
- Wang, C.; Fan, L.; Gao, H.; Wu, X.; Li, J.; Lv, G.; Gong, B. Polyamine biosynthesis and degradation are modulated by exogenous gamma-aminobutyric acid in root-zone hypoxia-stressed melon roots. Plant Physiol. Biochem. 2014, 82, 17–26. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bown, A.W.; McLean, M.D. Metabolism and functions of gamma-aminobutyric acid. Trends Plant Sci. 1999, 4, 446–452. [Google Scholar] [CrossRef]
- Ding, J.; Ulanov, A.V.; Dong, M.; Yang, T.; Nemzer, B.V.; Xiong, S.; Zhao, S.; Feng, H. Enhancement of gama-aminobutyric acid (GABA) and other health-related metabolites in germinated red rice (Oryza sativa L.) by ultrasonication. Ultrason. Sonochemistry 2018, 40, 791–797. [Google Scholar] [CrossRef]
- Kim, M.Y.; Lee, S.H.; Jang, G.Y.; Park, H.J.; Li, M.; Kim, S.; Lee, Y.R.; Noh, Y.H.; Lee, J.; Jeong, H.S. Effects of high hydrostatic pressure treatment on the enhancement of functional components of germinated rough rice (Oryza sativa L.). Food Chem. 2015, 166, 86–92. [Google Scholar] [CrossRef]
- Li, X.; Hao, J.; Liu, X.; Liu, H.; Ning, Y.; Cheng, R.; Tan, B.; Jia, Y. Effect of the treatment by slightly acidic electrolyzed water on the accumulation of gamma-aminobutyric acid in germinated brown millet. Food Chem. 2015, 186, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Fukumori, T.; Kanemoto, S.; Mizuno, H.; Wakabayashi, K.; Liu, H.Q.; Ochiai, S. Grain or Legume Having Increased Content of Functional Component and a Manufacturing Method Thereof. U.S. Patent 8,399,037 B2, 19 March 2013. Available online: https://www.freepatentsonline.com/8399037.html (accessed on 12 January 2022).
- Ma, Y.; Tong, L.; Li, J.; Ashraf, J.; Wang, S.; Zhao, B.; Liu, L.; Blecker, C.; Zhou, S. Comparison of γ-aminobutyric acid accumulation capability in different mung bean (Vigna radiata L.) varieties under heat and relative humidity treatment, and its correlation with endogenous amino acids and polyamines. Int. J. Food Sci. Technol. 2020, 56, 1562–1573. [Google Scholar] [CrossRef]
- Andersson, A.; Elfverson, C.; Andersson, R.; Regner, S.; Aman, P. Chemical and physical characteristics of different barley samples. J. Sci. Food Agric. 1999, 79, 979–986. [Google Scholar] [CrossRef]
- American Association of Cereal Chemistry (AACC). Crude Protein—Kjeldahl Method, Boric Acid Modification (46-12.01); American Association of Cereal Chemistry (AACC): St. Paul, MI, USA, 1999. [Google Scholar]
- Oh, S.J.; Kim, H.S.; Lim, S.T.; Reddy, C.K. Enhanced accumulation of gamma-aminobutyric acid in rice bran using anaerobic incubation with various additives. Food Chem. 2019, 271, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Petrivalsky, M.; Brauner, F.; Luhova, L.; Gagneul, D.; Sebela, M. Aminoaldehyde dehydrogenase activity during wound healing of mechanically injured pea seedlings. J. Plant Physiol. 2007, 164, 1410–1418. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.L.; Zhai, H.Q.; Wan, J.M. Accumulation of γ-aminobutyric acid in giant-embryo rice grain in relation to glutamate decarboxylase activity and its gene expression during water soaking. Cereal Chem. 2007, 82, 191–196. [Google Scholar] [CrossRef]
- Liao, J.; Wu, X.; Xing, Z.; Li, Q.; Duan, Y.; Fang, W.; Zhu, X. γ-Aminobutyric acid (GABA) accumulation in tea (Camellia sinensis L.) through the GABA shunt and polyamine degradation pathways under anoxia. J. Agric. Food Chem. 2017, 65, 3013–3018. [Google Scholar] [CrossRef]
- Bleidere, M.; Gaile, Z. Grain quality traits important in feed barley. Proc. Latv. Acad. Sci. Sect. B 2012, 66, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Yang, P. Studies on the molecular mechanisms of seed germination. Proteomics 2015, 15, 1671–1679. [Google Scholar] [CrossRef]
- Moza, J.; Gujral, H.S. Starch digestibility and bioactivity of high altitude hulless barley. Food Chem. 2016, 194, 561–568. [Google Scholar] [CrossRef]
- Jood, S.; Kalra, S. Chemical composition and nutritional characteristics of some hull less and hulled barley cultivars grown in India. Mol. Nutr. Food Res. 2001, 45, 35–39. [Google Scholar] [CrossRef]
- Wang, J.; Zhong, Z.; Feng, X.; Fu, G.; Hou, W.; Wang, G.; Da, C. Spatial distribution regulation of protein content of naked barley varieties and its relationships with environmental factors in Qinghai-Tibet Plateau. Sci. Agric. Sin. 2017, 50, 969–977. [Google Scholar] [CrossRef]
- Podlesakova, K.; Ugena, L.; Spichal, L.; Dolezal, K.; Diego, N.D. Phytohormones and polyamines regulate plant stress responses by altering GABA pathway. New Biotechnol. 2019, 48, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiang, J.; Zhang, L.; Zhu, X.; Evers, J.; Werf, W.; Duan, L. Optimizing soaking and germination conditions to improve gamma-aminobutyric acid content in japonica and indica germinated brown rice. J. Funct. Foods 2014, 10, 283–291. [Google Scholar] [CrossRef]
- Srivastava, B.I.S. Polyamine changes during senescence and tumorogenesis in plants. Mech. Ageing Dev. 1987, 40, 17–30. [Google Scholar] [CrossRef]
- Fait, A.; Fromm, H.; Walter, D.; Galili, G.; Fernie, A.R. Highway or byway: The metabolic role of the GABA shunt in plants. Trends Plant Sci. 2008, 13, 14–19. [Google Scholar] [CrossRef]
- Saeed, S.; Foad, M. Stress-induced changes in the free amino acid composition of two wheat cultivars with difference in drought resistance. Afr. J. Biotechnol. 2012, 11, 9559–9565. [Google Scholar] [CrossRef]
- Kamara, J.S.; Konishi, S.; Sasanuma, T.; Abe, T. Variation in free amino acid profile among some rice (Oryza sativa L.) cultivars. Breed. Sci. 2010, 60, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Guo, Q.; Gu, Z. GABA shunt and polyamine degradation pathway on gamma-aminobutyric acid accumulation in germinating fava bean (Vicia faba L.) under hypoxia. Food Chem. 2013, 136, 152–159. [Google Scholar] [CrossRef]
- Umbarger, H.E. Amino acid biosynthesis and its regulation. Annu. Rev. Biochem. 1978, 47, 532–606. [Google Scholar] [CrossRef]
- Chung, H.J.; Jang, S.H.; Cho, H.Y.; Lim, S.T. Effects of steeping and anaerobic treatment on GABA (γ-aminobutyric acid) content in germinated waxy hull-less barley. LWT-Food Sci. Technol. 2009, 42, 1712–1716. [Google Scholar] [CrossRef]
- Stepansky, A.; Less, H.; Angelovici, R.; Aharon, R.; Zhu, X.; Galili, G. Lysine catabolism, an effective versatile regulator of lysine level in plants. Amino Acids 2006, 30, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.; Dans, P.D.; Coitino, E.L.; Borsani, O.; Monza, J. Connecting proline and gamma-aminobutyric acid in stressed plants through non-enzymatic reactions. PLoS ONE 2015, 10, e0115349. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Zhou, S.; Wang, L.; Liu, X.; Ma, Y.; Tong, L.; Zhang, Y.; Wang, F. Effect of an Environment Friendly Heat and Relative Humidity Approach on γ-Aminobutyric Acid Accumulation in Different Highland Barley Cultivars. Foods 2022, 11, 691. https://doi.org/10.3390/foods11050691
Wang S, Zhou S, Wang L, Liu X, Ma Y, Tong L, Zhang Y, Wang F. Effect of an Environment Friendly Heat and Relative Humidity Approach on γ-Aminobutyric Acid Accumulation in Different Highland Barley Cultivars. Foods. 2022; 11(5):691. https://doi.org/10.3390/foods11050691
Chicago/Turabian StyleWang, Shanshan, Sumei Zhou, Lili Wang, Xiaojiao Liu, Yuling Ma, Litao Tong, Yuhong Zhang, and Fengzhong Wang. 2022. "Effect of an Environment Friendly Heat and Relative Humidity Approach on γ-Aminobutyric Acid Accumulation in Different Highland Barley Cultivars" Foods 11, no. 5: 691. https://doi.org/10.3390/foods11050691