
Functional Fermented Milk with Fruit Pulp Modulates the In Vitro Intestinal Microbiota

 ,
,  ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lactic Acid Bacteria (LAB) Cultures
2.2. Passion Fruit and Buriti Pulps Preparation
2.3. Preparation of the Cultures and Fermented Milk
2.4. Lactic Acid Bacteria Viability in Fermented Milk
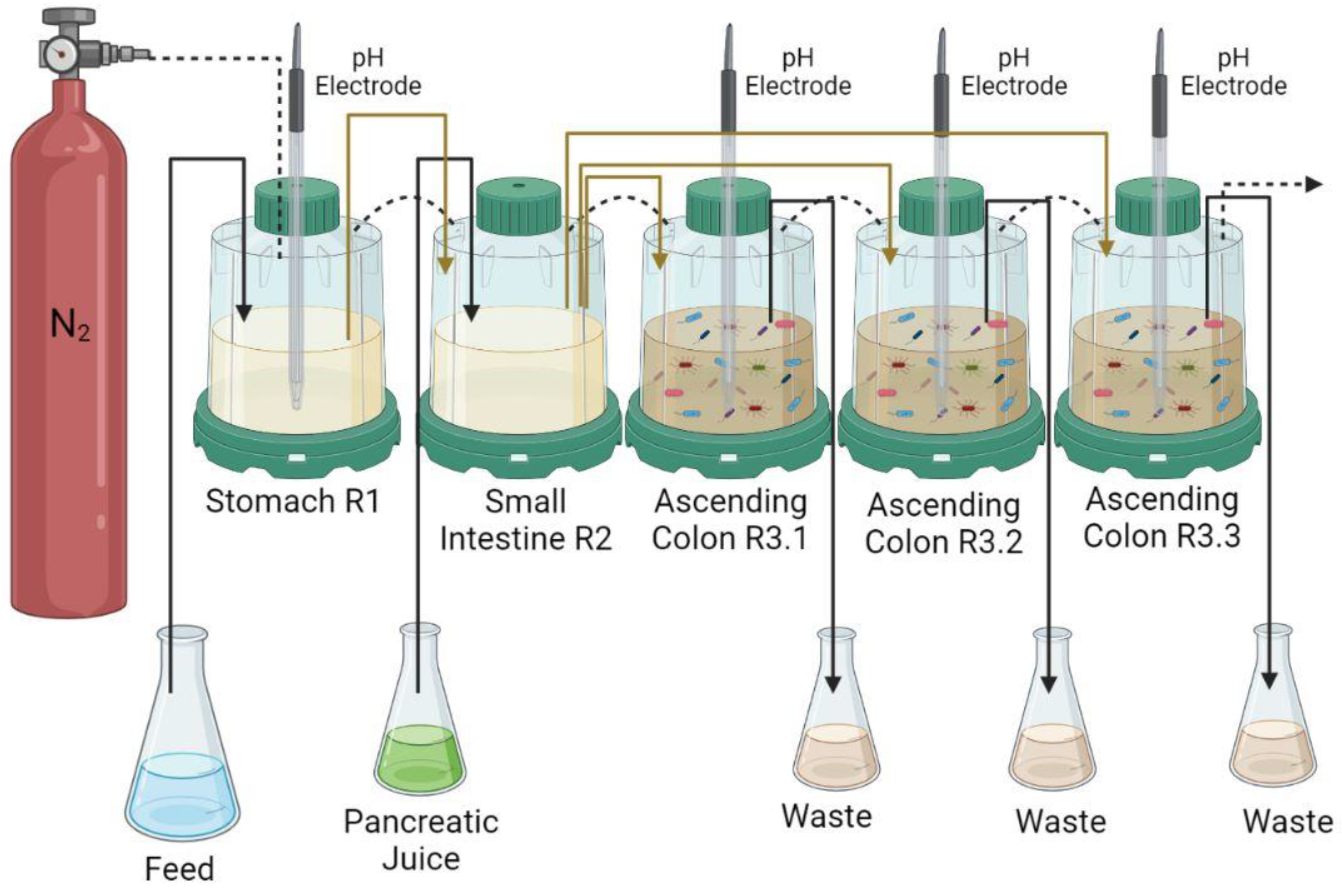
2.5. Simulator of Human Intestinal Microbial Ecosystem (SHIME®)
2.5.1. Composition of the SHIME® Feed Medium
2.5.2. Microbiota Colonization
2.5.3. Experimental Protocol
2.6. Evaluation of Metabolites Production in SHIME®
2.7. Microbial 16S rRNA Gene-Based Sequencing and Bioinformatic Analysis
2.8. Statistical Analysis
3. Results and Discussion
3.1. LAB Viability in Fermented Milk
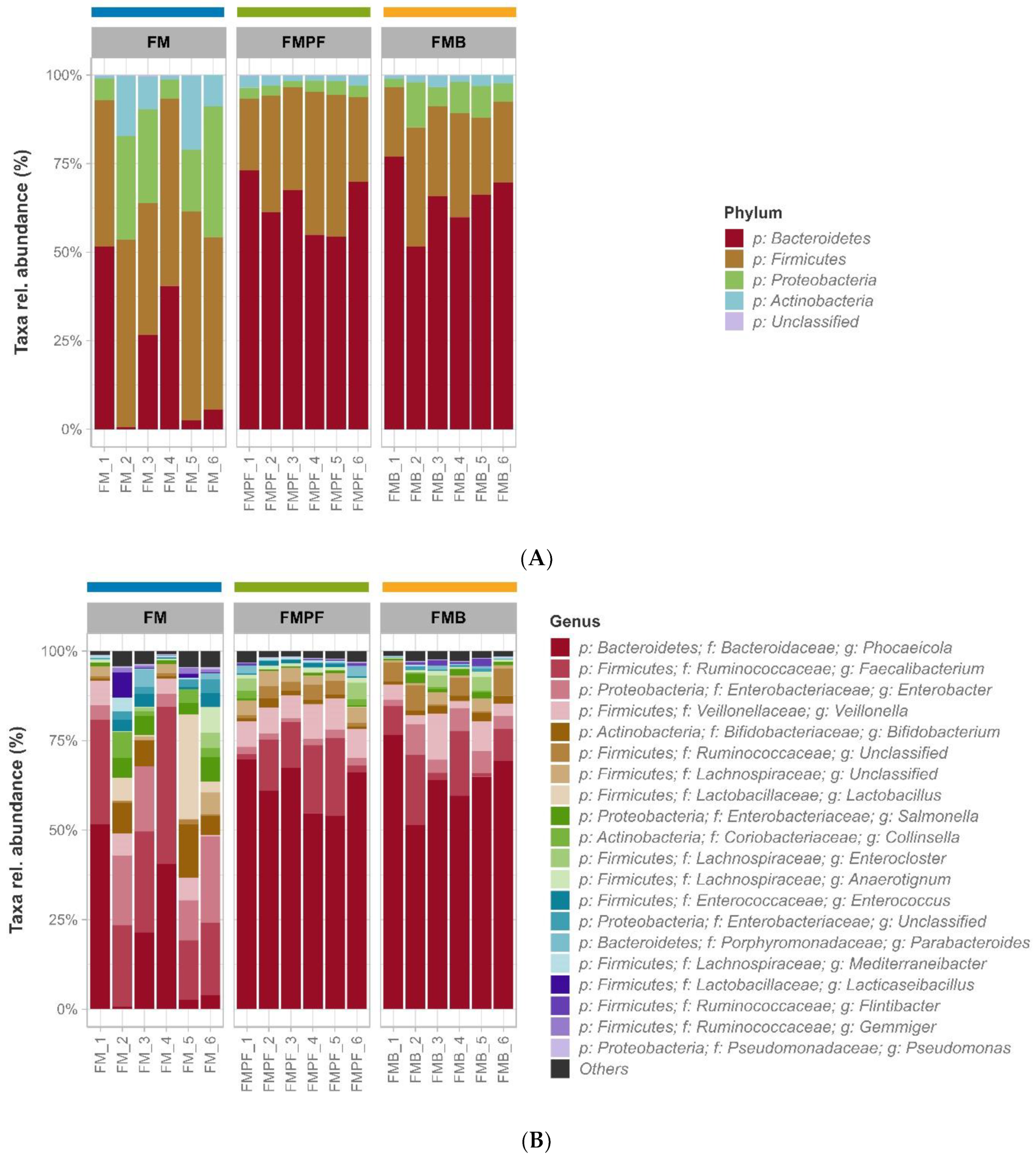
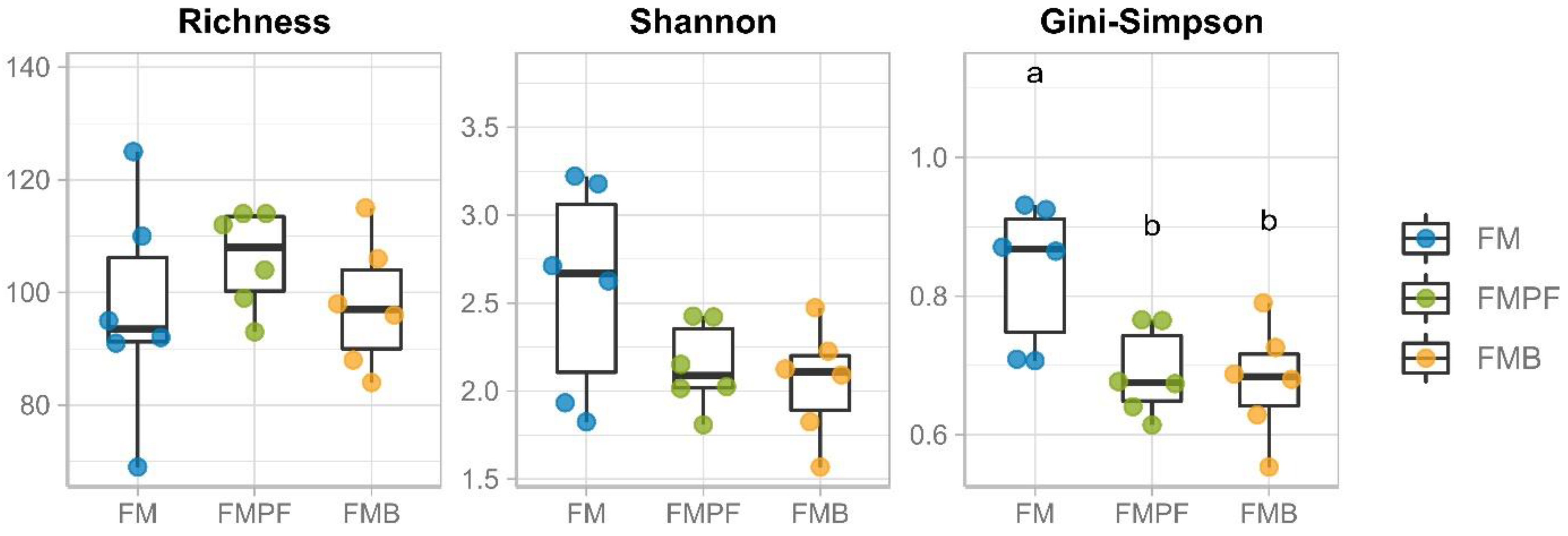
3.2. Microbiota Composition Evaluation by Gene Sequencing
3.2.1. Data Processing
3.2.2. Taxonomic Profile and Ecology
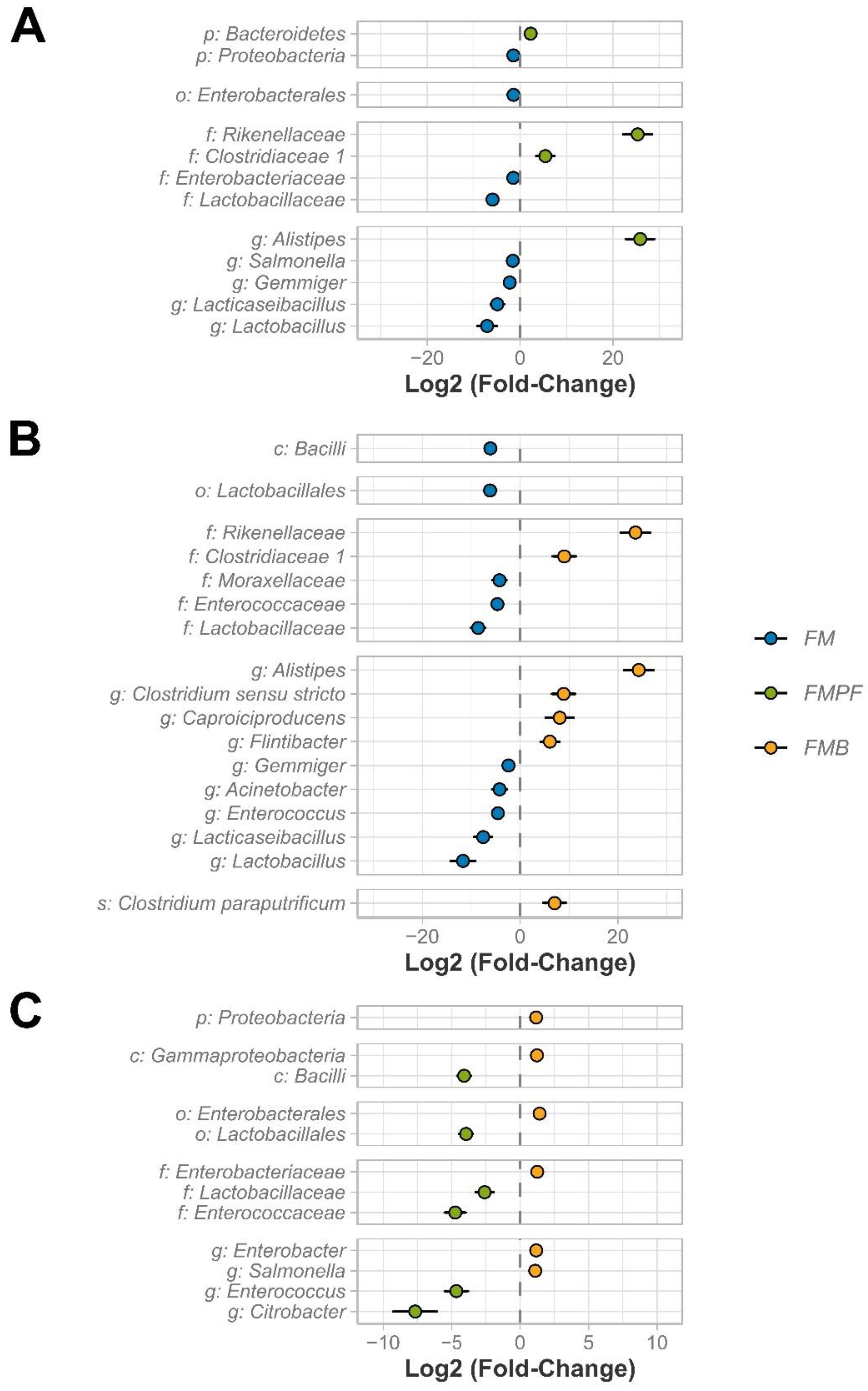
3.2.3. Differential Abundance
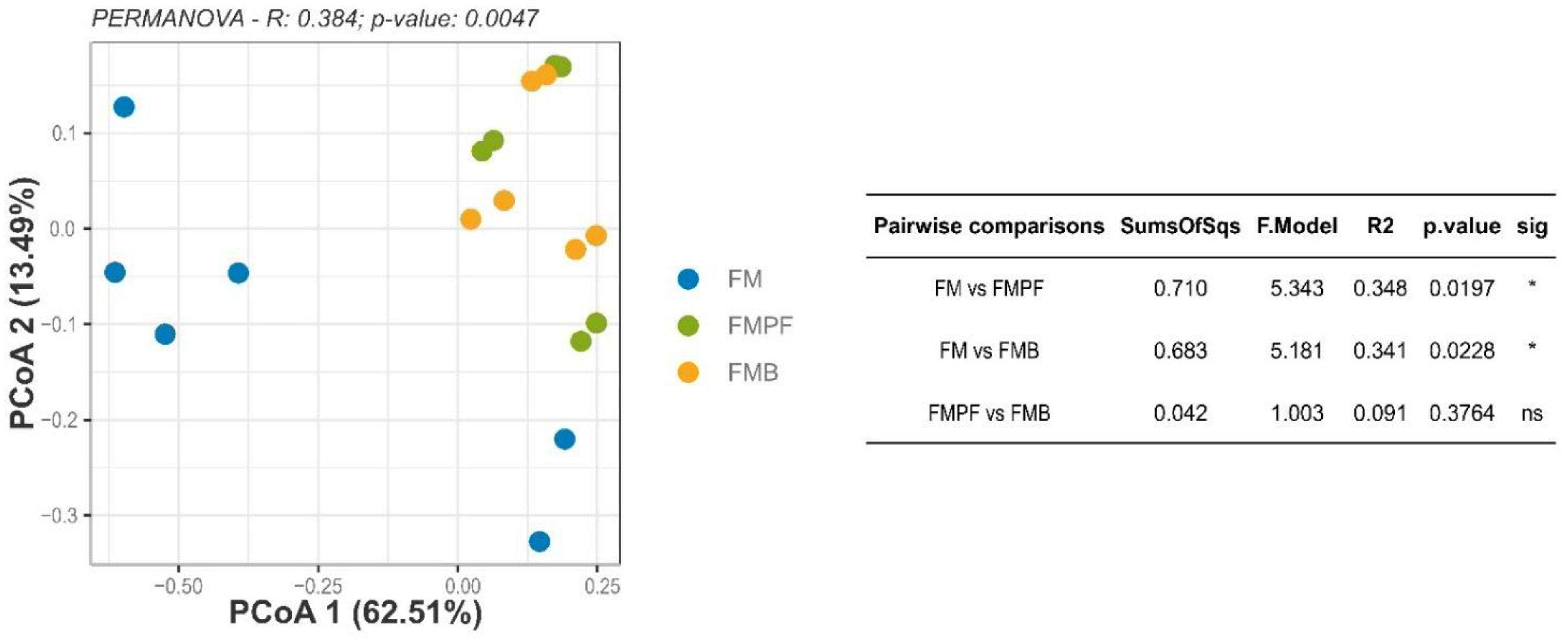
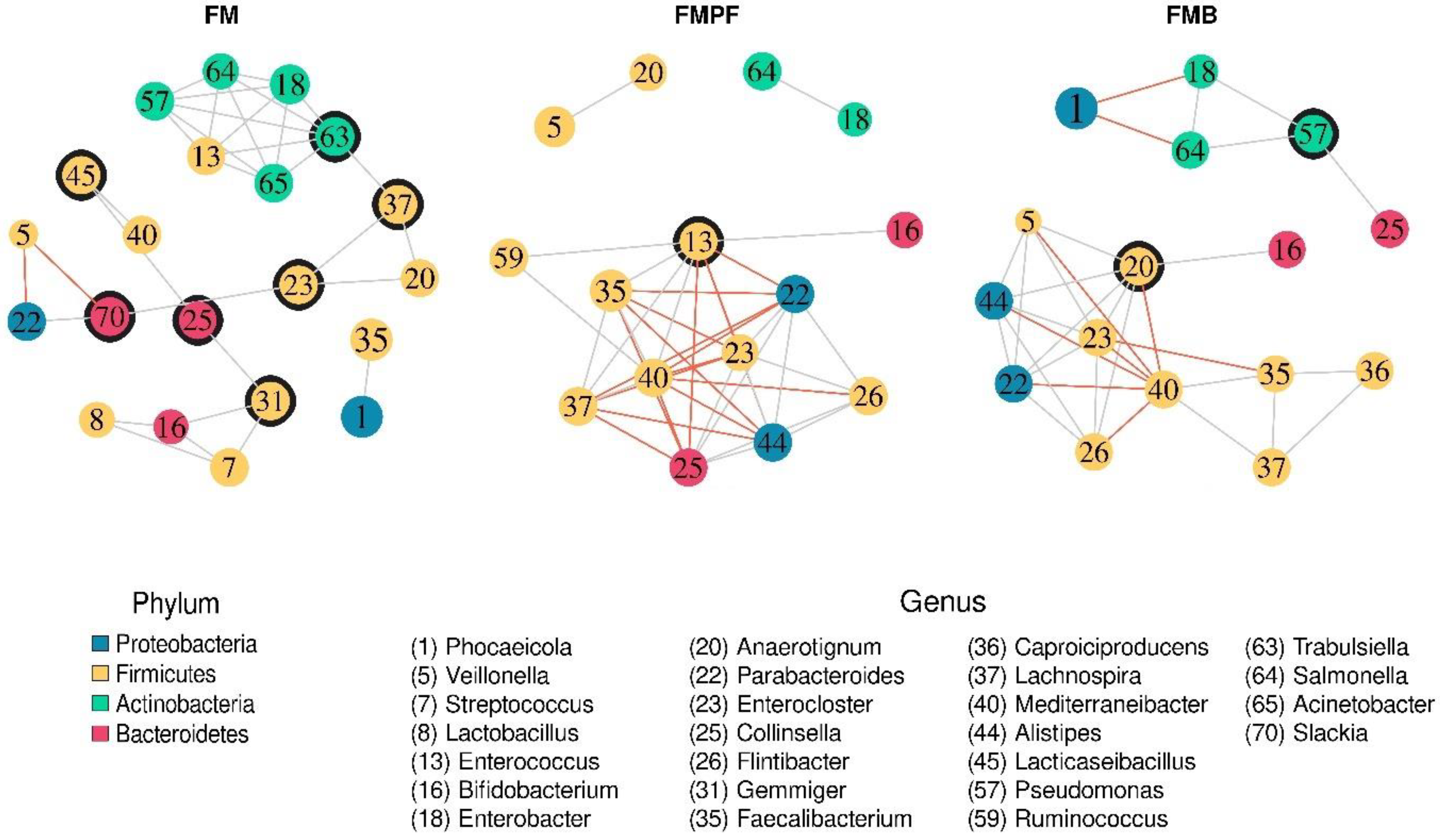
3.2.4. Structure of the Microbial Communities
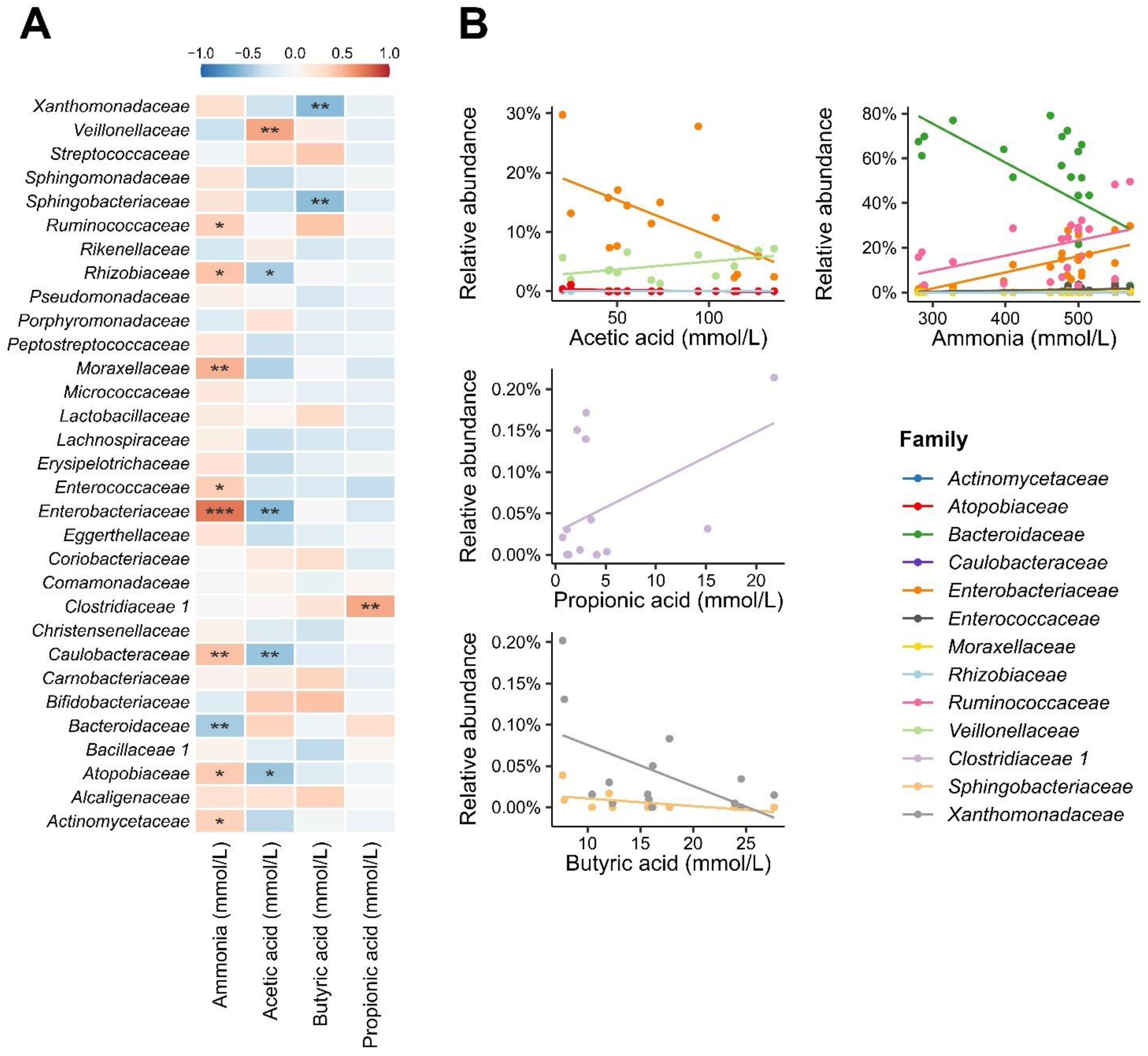
3.3. Short-Chain Fatty Acids and Ammonium Ions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Banwo, K.; Olojede, A.O.; Adesulu-Dahunsi, T.; Verma, D.K.; Thakur, M.; Tripathy, S.; Singh, S.; Patel, A.R.; Gupta, A.K.; Aguilar, C.N.; et al. Functional importance of bioactive compounds of foods with potential health benefits: A review on recent trends. Food Biosci. 2021, 43, 101320. [Google Scholar] [CrossRef]
- Teixeira, J.L.P.; Baptista, D.P.; Orlando, E.A.; Gigante, M.L.; Pallone, J.A.L. Effect of processing on the bioaccessibility of essential minerals in goat and cow milk and dairy products assessed by different static in vitro digestion models. Food Chem. 2022, 374, 131739. [Google Scholar] [CrossRef] [PubMed]
- Cha, K.H.; Lee, E.H.; Yoon, H.S.; Lee, J.H.; Kim, J.Y.; Kang, K.; Soo, P.J.; Jin, J.B.; Ko, G.; Pan, C.H. Effects of fermented milk treatment on microbial population and metabolomic outcomes in a three-stage semi-continuous culture system. Food Chem. 2018, 263, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Casarotti, S.N.; Borgonovi, T.F.; Tieghi, T.M.; Sivieri, K.; Penna, A.L.B. Probiotic low-fat fermented goat milk with passion fruit by-product: In vitro effect on obese individuals’ microbiota and on metabolites production. Food Res. Int. 2020, 136, 109453. [Google Scholar] [CrossRef] [PubMed]
- Barat, A.; Ozcan, T. Growth of probiotic bacteria and characteristics of fermented milk containing fruit matrices. Int. J. Dairy Technol. 2018, 71, 120–129. [Google Scholar] [CrossRef]
- Borgonovi, T.F.; Casarotti, S.N.; Penna, A.L.B. Lacticaseibacillus casei SJRP38 and buriti pulp increased bioactive compounds and probiotic potential of fermented milk. LWT Food Sci. Technol. 2021, 143, 111124. [Google Scholar] [CrossRef]
- Maqsood, S.; Adiamoa, O.; Ahmada, M.; Mudgil, P. Bioactive compounds from date fruit and seed as potential nutraceutical and functional food ingredients. Food Chem. 2020, 308, 125522. [Google Scholar] [CrossRef]
- Coman, V.; Teleky, B.E.; Mitrea, L.; Martău, G.A.; Szabo, K.; Călinoiu, L.F.; Vodnar, D.C. Bioactive potential of fruit and vegetable wastes. Adv. Food Nutr. Res. 2019, 91, 157–225. [Google Scholar] [CrossRef]
- Tabashsum, Z.; Peng, M.; Kahan, E.; Rahaman, S.O.; Biswas, D. Effect of conjugated linoleic acid overproducing Lactobacillus with berry pomace phenolic extracts on Campylobacter jejuni pathogenesis. Food Funct. 2019, 10, 296–303. [Google Scholar] [CrossRef]
- Ahmad, I.; Khalique, A.; Junaid, M.; Shahid, M.Q.; Imran, M.; Rashid, A.A. Effect of polyphenol from apple peel extract on the survival of probiotics in yoghurt ice cream. Food Sci. Technol. Int. 2020, 55, 2580–2588. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, M.; Sun, X.; Liu, X.; Choueiry, F.; Xu, R.; Shi, H.; Zhu, J. Black raspberry extract shifted gut microbe diversity and their metabolic landscape in a human colonic model. J. Chromatogr. B 2022, 1188, 123027. [Google Scholar] [CrossRef]
- Cheng, Y.; Wu, T.; Chu, X.; Tang, S.; Cao, W.; Liang, F.; Fang, Y.; Pan, S.; Xu, X. Fermented blueberry pomace with antioxidant properties improves fecal microbiota community structure and short chain fatty acids production in an in vitro mode. LWT Food Sci. Technol. 2020, 125, 109260. [Google Scholar] [CrossRef]
- Espín, J.C.; González-Sarrías, A.; Tomás-Barberán, F.A. The gut microbiota: A key factor in the therapeutic effects of (poly)phenols. Biochem. Pharmacol. 2017, 139, 82–93. [Google Scholar] [CrossRef]
- Piekarska-Radzik, L.; Klewicka, E. Mutual influence of polyphenols and Lactobacillus spp. bacteria in food: A review. Eur. Food Res. Technol. 2021, 247, 9–24. [Google Scholar] [CrossRef]
- Alkushi, A.G.; Abdelfattah-Hassan, A.; Eldoumani, H.; Elazab, S.T.; Mohamed, S.A.M.; Metwally, A.S.; El-Shetry, E.S.; Saleh, A.A.; Elsawy, N.A.; Ibrahim, D. Probiotics-loaded nanoparticles attenuated colon inflammation, oxidative stress, and apoptosis in colitis. Sci. Rep. 2022, 12, 5116. [Google Scholar] [CrossRef]
- Gebrayel, P.; Nicco, C.; Khodor, S.A.; Bilinski, J.; Caselli, E.; Comelli, E.M.; Egert, M.; Giaroni, C.; Karpinski, T.M.; Loniewski, I.; et al. Microbiota medicine: Towards clinical revolution. J. Transl. Med. 2022, 20, 111. [Google Scholar] [CrossRef]
- Ho, S.W.; El-Nezami, H.; Corke, H.; Ho, C.S.; Shah, N.P. L-citrulline enriched fermented milk with Lactobacillus helveticus attenuates dextran sulfate sodium (DSS) induced colitis in mice. J. Nutr. Biochem. 2022, 99, 108858. [Google Scholar] [CrossRef]
- Ren, S.; Chen, A.; Tian, Y.; Bai, Z.; Wang, C. Lactobacillus paracasei from koumiss ameliorates diarrhea in mice via tight junctions modulation. Nutrition 2022, 98, 111584. [Google Scholar] [CrossRef]
- Le Roy, C.I.; Kurilshikov, A.; Leeming, E.R.; Visconti, A.; Bowyer, R.C.E.; Menni, C.; Falchi, M.; Koutnikova, H.; Veiga, P.; Zhernakova, A.; et al. Yoghurt consumption is associated with changes in the composition of the human gut microbiome and metabolome. BMC Microbiol. 2022, 22, 39. [Google Scholar] [CrossRef]
- Salotti-Souza, B.M.; Borgonovi, T.F.; Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Lactobacillus casei and Lactobacillus fermentum strains isolated from mozzarella cheese: Probiotic potential, safety, acidifying kinetic parameters and viability under gastrointestinal tract conditions. Probiotics Antimicrob. Proteins 2019, 11, 382–396. [Google Scholar] [CrossRef]
- Todorov, S.D.; Perin, L.M.; Carneiro, B.M.; Rahal, P.; Holzapfel, W.; Nero, L.A. Safety of Lactobacillus plantarum ST8Sh and its bacteriocin. Probiotics Antimicrob. Proteins 2017, 9, 334–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura-Filho, J.M. Buriti (Mauritia flexuosa L.) Preparation: Production, Characterization and Application in Fermented Milk. Ph.D. Thesis, University of Sao Paulo, Sao Paulo, Brazil, 29 May 2017. Available online: https://repositorio.unesp.br/handle/11449/151139 (accessed on 5 September 2022).
- Solieri, L.; Bianchi, A.; Mottolese, G.; Lemmetti, F.; Giudici, P. Tailoring the probiotic potential of non-starter Lactobacillus strains from ripened Parmigiano Reggiano cheese by in vitro screening and principal component analysis. Food Microbiol. 2014, 38, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Molly, K.; Woestyne, M.V.; Smet, I.; Verstraete, W. Validation of the simulator of the human intestinal microbial ecosystem (SHIME®) reactor using microorganism associated activities. Microb. Ecol. Health Dis. 1994, 7, 191–200. [Google Scholar] [CrossRef]
- Salgaço, M.K.; Perina, N.P.; Tomé, T.M.; Mosquera, E.M.B.; Lazarini, T.; Sartoratto, A.; Sivieri, K. Probiotic infant cereal improves children’s gut microbiota: Insights using the Simulator of Human Intestinal Microbial Ecosystem (SHIME®). Food Res. Int. 2021, 143, 110292. [Google Scholar] [CrossRef] [PubMed]
- Possemiers, S.; Verthé, K.; Uyttendaele, S.; Verstraete, W. PCR-DGGE-based quantification of stability of the microbial community in a simulator of the human intestinal microbial ecosystem. FEMS Microbiol. Ecol. 2004, 49, 495–507. [Google Scholar] [CrossRef]
- Duque, A.L.R.F.; Monteiro, M.; Adorno, M.A.T.; Sakamoto, I.K.; Sivieri, K. An exploratory study on the influence of orange juice on gut microbiota using a dynamic colonic model. Food Res. Int. 2016, 84, 160–169. [Google Scholar] [CrossRef]
- Van De Wiele, T.; Boon, N.; Possemiers, S.; Jacobs, H.; Verstraete, W. Prebiotic effects of chicory inulin in the simulator of the human intestinal microbial ecosystem. FEMS Microbiol. Ecol. 2004, 51, 143–153. [Google Scholar] [CrossRef]
- Espírito Santo, A.P.; Cartolano, N.S.; Silva, T.F.; Soares, F.A.; Gioielli, L.A.; Perego, P.; Converti, A.; Oliveira, M.N. Fibers from fruit by-products enhance probiotic viability and fatty acid profile and increase CLA content in yoghurts. Int. J. Food Microbiol. 2012, 154, 135–144. [Google Scholar] [CrossRef]
- Adorno, M.A.T.; Hirasawa, J.S.; Varesche, M.B.A. Development and validation of two methods to quantify volatile acids (C2–C6) by GC/FID: Headspace (automatic and manual) and liquid–liquid extraction (LLE). Am. J. Anal. Chem. 2014, 5, 406–414. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, F.; Rossi, E.A.; Sakamoto, I.K.; Adorno, M.A.T.; Van De Wiele, T.; Sivieri, K. Beneficial effects of fermented vegetal beverages on human gastrointestinal microbial ecosystem in a simulator. Food Res. Int. 2014, 64, 43–52. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Didion, J.P.; Martin, M.; Collins, F.S. Atropos: Specific, sensitive, and speedy trimming of sequencing reads. PeerJ 2017, 5, e3720. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R. The Ribosomal Database Project (RDP-II): Sequences and tools for high-throughput rRNA analysis. Nucleic Acids Res. 2004, 33, D294–D296. [Google Scholar] [CrossRef] [Green Version]
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. Ampvis2: An R package to analyse and visualise 16S rRNA amplicon data. BioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Lahti, L.; Shetty, S. Microbiome R Package, 2012. (Introduction to the microbiome R package [Internet]). Available online: https://microbiome.github.io/tutorials/ (accessed on 10 November 2022).
- Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. Version 1.3-5. 2021. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 8 October 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 20 October 2022).
- Arbizu, P.M. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis. 2019. Available online: https://rdrr.io/github/gauravsk/ranacapa/man/pairwise_adonis.html (accessed on 12 August 2022).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A. ggpubr: “ggplot2” Based Publication Ready Plots. Version 0.4.0. 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 12 August 2022).
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; Version 2.2.9; Northwestern University: Evanston, IL, USA, 2022; Available online: https://CRAN.R-project.org/package=psych (accessed on 12 August 2022).
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Complex Systems. Inter J. 2006, 1695, 1–9. Available online: https://www.bibsonomy.org/bibtex/bb49a4a77b42229a427fec316e9fe515 (accessed on 15 August 2022).
- Ozcan, T.; Yilmaz-Ersan, L.; Akpinar-Bayizit, A.; Delikanli, B.; Balati, A. Survival of Lactobacillus spp. in fruit based fermented dairy beverages. Int. J. Food Eng. 2015, 1, 44–49. [Google Scholar] [CrossRef]
- Casarotti, S.N.; Borgonovi, T.F.; Batista, C.L.F.M.; Penna, A.L.B. Guava, orange and passion fruit by-products: Characterization and its impacts on kinetics of acidification and properties of probiotic fermented products. LWT Food Sci. Technol. 2018, 98, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Beisner, J.; Gonzalez-Granda, A.; Basrai, M.; Damms-Machado, A.; Bischoff, S.C. Fructose-induced intestinal microbiota shift following two types of short term high-fructose dietary phases. Nutrients 2020, 12, 3444. [Google Scholar] [CrossRef] [PubMed]
- Purchiaroni, F.; Tortora, A.; Gabrielli, M.; Bertucci, F.; Gigante, G.; Ianiro, G.; Ojetti, V.; Scarpellini, E.; Gasbarrini, A. The role of intestinal microbiota and the immune system. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 323–333. Available online: https://www.europeanreview.org/wp/wp-content/uploads/323-333.pdf (accessed on 25 October 2022). [PubMed]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A relevant minority for the maintenance of gut homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Azimirad, M.; Jo, Y.; Kim, M.S.; Jeong, M.; Shahrokh, S.; Aghdaei, H.A.; Zali, M.R.; Lee, S.; Yadegar, A.; Shin, J.H. Alterations and prediction of functional profiles of gut microbiota after fecal microbiota transplantation for Iranian recurrent Clostridioides difficile infection with underlying inflammatory bowel disease: A pilot study. J. Inflamm. Res. 2022, 15, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Rettedal, E.A.; Altermann, E.; Roy, N.C.; Dalziel, J.E. The effects of unfermented and fermented cow and sheep milk on the gut microbiota. Front. Microbiol. 2019, 10, 458. [Google Scholar] [CrossRef] [Green Version]
- Oddi, S.; Huber, P.; Duque, A.L.R.F.; Vinderola, G.; Sivieri, K. Breast-milk derived potential probiotics as strategy for the management of childhood obesity. Food Res. Int. 2020, 137, 109673. [Google Scholar] [CrossRef]
- Cani, P.D.; Moens De Hase, E.; Van Hul, M. Gut microbiota and host metabolism: From proof of concept to therapeutic intervention. Microorganisms 2021, 9, 1302. [Google Scholar] [CrossRef]
- Martín, R.; Miquel, S.; Benevides, L.; Bridonneau, C.; Robert, V.; Hudault, S.; Florian, C.; Berteau, O.; Azevedo, V.; Châtel, J.M.; et al. Functional characterization of novel Faecalibacterium prausnitzii strains isolated from healthy volunteers: A step forward in the use of F. Prausnitzii as a next-generation probiotic. Front. Microbiol. 2017, 8, 1226. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C. The startup bugs. Nat. Biotechnol. 2013, 31, 279–281. [Google Scholar] [CrossRef]
- Cobo, F.; Pérez-Carrasco, V.; Rodríguez-Guerrero, E.; Sampedro, A.; Rodríguez-Granger, J.; García-Salcedo, J.A.; Navarro-Mari, J.M. Misidentification of Phocaeicola (Bacteroides) dorei in two patients with bacteremia. Anaerobe 2022, 75, 102544. [Google Scholar] [CrossRef]
- Wang, C.; Li, S.; Zhang, Z.; Yu, Z.; Yu, L.; Tian, F.; Chen, W.; Zhai, Q. Phocaeicola faecalis sp. nov., a strictly anaerobic bacterial strain adapted to the human gut ecosystem. Antonie Van Leeuwenhoek 2021, 114, 1225–1235. [Google Scholar] [CrossRef]
- Lück, R.; Deppenmeier, U. Genetic tools for the redirection of the central carbon flow towards the production of lactate in the human gut bacterium Phocaeicola (Bacteroides) vulgatus. Appl. Microbiol. Biotechnol. 2022, 106, 1211–1225. [Google Scholar] [CrossRef]
- Wexler, A.G.; Goodman, A.L. An insider’s perspective: Bacteroides as a window into the microbiome. Nat. Microbiol. 2017, 25, 17026. [Google Scholar] [CrossRef] [Green Version]
- Flint, H.J.; Bayer, E.A.; Rincon, M.T.; Lamed, R.; White, B.A. Polysaccharide utilization by gut bacteria: Potential for new insights from genomic analysis. Nat. Rev. Microbiol. 2008, 6, 121–131. [Google Scholar] [CrossRef]
- Glover, J.S.; Ticer, T.D.; Engevik, M.A. Characterizing the mucin-degrading capacity of the human gut microbiota. Sci. Rep. 2022, 19, 8456. [Google Scholar] [CrossRef]
- Makki, K.; Deehan, E.; Walter, J.; Bäckhed, F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe 2018, 13, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; Reyes-Gavilán, C.G.; Salazar, N. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Russell, W.R.; Duncan, S.H.; Scobbie, L.; Duncan, G.; Cantlay, L.; Calder, A.G.; Anderson, S.E.; Flint, H. Major phenylpropanoid-derived metabolites in the human gut can arise from microbial fermentation of protein. Mol. Nutr. Food Res. 2013, 57, 523–535. [Google Scholar] [CrossRef]
- Benevides, L.; Burman, S.; Martin, R.; Robert, V.; Thomas, M.; Miquel, S.; Chain, F.; Sokol, H.; Bermudez-Humaran, L.G.; Morrison, M.; et al. New insights into the diversity of the genus Faecalibacterium. Front. Microbiol. 2017, 8, 1790. [Google Scholar] [CrossRef]
- Walker, A.W.; Ince, J.; Duncan, S.H.; Webster, L.M.; Holtrop, G.; Ze, X.; Brown, D.; Stares, M.D.; Scott, P.; Bergerat, A.; et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011, 5, 220–230. [Google Scholar] [CrossRef]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar] [CrossRef] [Green Version]
- Miquel, S.; Martín, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Halder, C.V.; Faria, A.V.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Zouiouich, S.; Loftfield, E.; Huybrechts, I.; Viallon, V.; Louca, P.; Vogtmann, E.; Wells, P.M.; Steves, C.J.; Herzig, K.H.; Menni, C.; et al. Markers of metabolic health and gut microbiome diversity: Findings from two population-based cohort studies. Diabetologia 2021, 64, 1749–1759. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.B.; Jonathan, M.; Saad, S.M.I.; Schols, H.A.; Venema, K. Degradation of fibres from fruit by-products allows selective modulation of the gut bacteria in an in vitro model of the proximal colon. J. Funct. Foods 2019, 1, 275–285. [Google Scholar] [CrossRef]
- Borgonovi, T.F.; Virgolin, L.B.; Janzantti, N.S.; Casarotti, S.N.; Penna, A.L.B. Fruit bioactive compounds: Effect on lactic acid bacteria and on intestinal microbiota. Food Res. Int. 2022, 161, 111809. [Google Scholar] [CrossRef]
- Rodríguez, L.G.R.; Gasga, V.M.Z.; Pescuma, M.; Nieuwenhove, C.V.; Mozzi, F.; Burgos, J.A.S. Fruits and fruit by-products as sources of bioactive compounds. Benefits and trends of lactic acid fermentation in the development of novel fruit-based functional beverages. Food Res. Int. 2021, 140, 109854. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and vegetables, as a source of nutritional compounds and phytochemicals: Changes in bioactive compounds during lactic fermentation. Food Res. Int. 2018, 104, 86–99. [Google Scholar] [CrossRef]
- Dou, Z.; Chen, C.; Fu, X. Bioaccessibility, antioxidant activity and modulation effect on gut microbiota of bioactive compounds from Moringa oleifera Lam. leaves during digestion and fermentation in vitro. Food Funct. 2019, 1, 5070–5079. [Google Scholar] [CrossRef]
- Luca, S.V.; Macovei, I.; Bujor, A.; Miron, A.; Skalicka-Woźniak, K.; Aprotosoaie, A.C.; Trifan, A. Bioactivity of dietary polyphenols: The role of metabolites. Crit. Rev. Food Sci. Nutr. 2020, 60, 626–659. [Google Scholar] [CrossRef]
- Molinari, R.; Merendino, N.; Costantini, L. Polyphenols as modulators of pre-established gut microbiota dysbiosis: State-of-the-art. Biofactors 2022, 48, 255–273. [Google Scholar] [CrossRef]
- Belizário, J.E.; Faintuch, J.; Garay-Malpartida, M. Gut Microbiome Dysbiosis and Immunometabolism: New Frontiers for Treatment of Metabolic Diseases. Mediat. Inflamm. 2018, 2018, 2037838. [Google Scholar] [CrossRef]
- Chidambaram, S.B.; Essa, M.M.; Rathipriya, A.G.; Bishir, M.; Ray, B.; Mahalakshmi, A.M.; Tousif, A.H.; Sakharkar, M.K.; Kashyap, R.S.; Friedland, R.P.; et al. Gut dysbiosis, defective autophagy and altered immune responses in neurodegenerative diseases: Tales of a vicious cycle. Pharmacol. Ther. 2022, 231, 107988. [Google Scholar] [CrossRef]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. LWT Food Sci. Technol. 2018, 4, 308–325. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.H.W.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ. Res. 2017, 31, 1183–1196. [Google Scholar] [CrossRef] [Green Version]
- Wiegel, J.; Tanner, R.; Rainey, F.A. An introduction to the family Clostridiaceae. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 4, pp. 654–678. [Google Scholar] [CrossRef]
- Jessen, J.E.; Orlygsson, J. Production of ethanol from sugars and lignocellulosic biomass by Thermoanaerobacter J1 isolated from a hot spring in Iceland. J. Biomed. Biotechnol. 2012, 2012, 186982. [Google Scholar] [CrossRef] [Green Version]
- Kong, C.; Gao, R.; Yan, X.; Huang, L.; Qin, H. Probiotics improve gut microbiota dysbiosis in obese mice fed a high-fat or high-sucrose diet. Nutrition 2019, 60, 175–184. [Google Scholar] [CrossRef]
- Graf, J. The Family Rikenellaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 857–859. [Google Scholar] [CrossRef]
- Shkoporov, A.N.; Chaplin, A.V.; Khokhlova, E.V.; Shcherbakova, V.A.; Motuzova, O.V.; Bozhenko, V.K.; Kafarskaia, L.L.; Efimov, B.A. Alistipes inops sp. nov. and Coprobacter secundus sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2015, 65, 4580–4588. [Google Scholar] [CrossRef]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut bacteria with emerging implications to inflammation, cancer, and mental health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef]
- Felis, G.E.; Pot, B. The family Lactobacillaceae. In Lactic Acid Bacteria; Holzapfel, W.H., Wood, B.J.B., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 245–247. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/9781118655252.part4 (accessed on 15 September 2022).
- Widyastuti, Y.; Febrisiantosa, A.; Tidona, F. Health-promoting properties of lactobacilli in fermented dairy products. Front. Microbiol. 2021, 21, 673890. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Pimentel, T.C.; Brandão, L.R.; Oliveira, M.P.; Costa, W.K.A.; Magnani, M. Health benefits and technological effects of Lacticaseibacillus casei-01: An overview of the scientific literature. Trends Food Sci. Technol. 2021, 1, 722–737. [Google Scholar] [CrossRef]
- De Filippis, F.; Pasolli, E.; Ercolini, D. The food-gut axis: Lactic acid bacteria and their link to food, the gut microbiome and human health. FEMS Microbiol. Rev. 2020, 1, 454–489. [Google Scholar] [CrossRef]
- Martins, A.R.; Burkert, C.A.V. Galactooligosaccharides (GOS) and their prebiotic and bifidogenic effects. Braz. J. Food Technol. 2009, 12, 230–240. [Google Scholar] [CrossRef]
- Diener, C.; Gibbons, S.M.; Resendis-Antonio, O. MICOM: Metagenome-scale modeling to infer metabolic interactions in the gut microbiota. mSystems 2020, 5, e00606-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, S.A.; Kuipers, B.; Atashgahi, S.; Aalvink, S.; Smidt, H.; De Vos, W.M. Inter-species metabolic interactions in an in-vitro minimal human gut microbiome of core bacteria. NPJ Biofilms Microbiomes 2022, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Chousalkar, K.K. Short-term feeding of probiotics and synbiotics modulates caecal microbiota during Salmonella Typhimurium infection but does not reduce shedding and invasion in chickens. Appl. Microbiol. Biotechnol. 2020, 104, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, ethnicity or subsistence-specific variations in human microbiome composition and diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, H.; Zhong, C.; Yun, Y.H.; Chen, W.; Chen, W. Determination of microbial diversity and community composition in unfermented and fermented washing rice water by high-throughput sequencing. Curr. Microbiol. 2021, 78, 1730–1740. [Google Scholar] [CrossRef]
- Wongkuna, S.; Ghimire, S.; Chankhamhaengdecha, S.; Janvilisri, T.; Scaria, J. Mediterraneibacter catenae SW178 sp. nov., an intestinal bacterium of feral chicken. PeerJ 2021, 9, e11050. [Google Scholar] [CrossRef]
- Ruiz-Rico, M.; Renwick, S.; Vancuren, S.J.; Robinson, A.V.; Gianetto-Hill, C.; Allen-Vercoe, E.; Barat, J.M. Influence of free and immobilized chitosan on a defined human gut microbial ecosystem. Food Res. Int. 2022, 161, 111890. [Google Scholar] [CrossRef]
- Fagundes, R.R.; Bourgonje, A.R.; Saeed, A.; Vila, A.V.; Plomp, N.; Blokzijl, T.; Sadabad, M.S.; von Martels, J.Z.H.; van Leeuwen, S.S.; Weersma, R.K.; et al. Inulin-grown Faecalibacterium prausnitzii cross-feeds fructose to the human intestinal epithelium. Gut Microbes 2021, 13, 1993582. [Google Scholar] [CrossRef]
- Wan, M.; Li, Q.; Lei, Q.; Zhou, D.; Wang, S. Polyphenols and polysaccharides from morus alba l. fruit attenuate high-fat diet-induced metabolic syndrome modifying the gut microbiota and metabolite profile. Foods 2022, 11, 1818. [Google Scholar] [CrossRef]
- Li, J.; Pang, B.; Yan, X.; Shang, X.; Hu, X.; Shi, J. Prebiotic properties of different polysaccharide fractions from Artemisia sphaerocephala Krasch seeds evaluated by simulated digestion and in vitro fermentation by human fecal microbiota. Int. J. Biol. Macromol. 2020, 162, 414–424. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Pukall, R.; Abt, B.; Foesel, B.U.; Meier-Kolthoff, J.P.; Kumar, N.; Bresciani, A.; Martines, I.; Just, S.; Ziegler, C.; et al. The Mouse Intestinal Bacterial Collection (miBC) provides host-specific insight into cultured diversity and functional potential of the gut microbiota. Nat. Microbiol. 2016, 1, 16131. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.; Kim, Y.; Lee, C.; Kwon, B.; Noh, J.; Jee, J.J.; Yoon, S.S.; Koh, H.; Sowon, P. The effect of formula-based nutritional treatment on colitis in a murine model. J. Korean Med. Sci. 2021, 27, e342. [Google Scholar] [CrossRef]
- Guzior, D.V.; Quinn, R.A. Review: Microbial transformations of human bile acids. Microbiome 2021, 9, 140. [Google Scholar] [CrossRef]
- Baquero, M.R.; Del Campo, R.; Bravo-Vázquez, D.A. Microcins in Enterobacteriaceae: Peptide antimicrobials in the eco-active intestinal chemosphere. Front. Microbiol. 2019, 10, 2261. [Google Scholar] [CrossRef]
- Sibinelli-Sousa, S.; Araújo-Silva, A.L.; Hespanhol, J.T.; Bayer-Santos, E. Revisiting the steps of Salmonella gut infection with a focus on antagonistic interbacterial interactions. FEBS J. 2022, 289, 4192–4211. [Google Scholar] [CrossRef]
- Feng, W.; Ao, H.; Peng, C. Gut microbiota, short-chain fatty acids, and herbal medicines. Front. Pharmacol. 2018, 9, 1354. [Google Scholar] [CrossRef] [PubMed]
- Ban, Q.; Cheng, J.; Sun, X.; Jiang, Y.; Zhao, S.; Song, X.; Guo, M. Effects of a synbiotic yogurt using monk fruit extract as sweetener on glucose regulation and gut microbiota in rats with type 2 diabetes mellitus. J. Dairy Sci. 2020, 103, 2956–2968. [Google Scholar] [CrossRef] [PubMed]
- Khuropakhonphong, R.; Whanmek, K.; Purttiponthanee, S.; Chathiran, W.; Srichamnong, W.; Santivarangkna, C.; Trachootham, D. Bulgarian yogurt relieved symptoms and distress and increased fecal short-chain fatty acids in healthy constipated women: A randomized, blinded crossover-controlled trial. NFS J. 2021, 22, 20–31. [Google Scholar] [CrossRef]
- He, X.; Parenti, M.; Grip, T.; Lönnerdal, B.; Timby, N.; Domellof, M.; Hernell, O.; Slupsky, C. Fecal microbiome and metabolome of infants fed bovine MFGM supplemented formula or standard formula with breast-fed infants as reference: A randomized controlled trial. Sci. Rep. 2019, 9, 11589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davila, A.M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.H.; Sanz, Y.; Tomé, D. Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host. Pharmacol. Res. 2013, 68, 95–107. [Google Scholar] [CrossRef]
- Rodrigues, V.C.C.; Duque, A.L.R.F.; Fino, L.C.; Simabuco, F.M.; Sartoratto, A.; Cabral, L.; Noronha, M.F.; Sivieri, K.; Antunes, A.E.C. Modulation of the intestinal microbiota and the metabolites produced by the administration of ice cream and a dietary supplement containing the same probiotics. Br. J. Nutr. 2020, 12, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, F.; Larsen, N.; De Mello Tieghi, T.; Adorno, M.A.T.; Kot, W.; Saad, S.M.I.; Jespersen, L.; Sivieri, K. Modulation of gut microbiota from obese individuals by in vitro fermentation of citrus pectin in combination with Bifidobacterium longum BB-46. Appl. Microbiol. Biotechnol. 2018, 102, 8827–8840. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, V.P.B.; Salgaço, M.K.; Pimentel, T.C.; Silva, T.C.R.; Sartoratto, A.; Lima, M.S.; Sassi, C.F.C.; Mesa, V.; Magnani, M.; Sivieri, K. Spirulina platensis biomass enhances the proliferation rate of Lactobacillus acidophilus 5 (La-5) and combined with La-5 impact the gut microbiota of medium-age healthy individuals through an in vitro gut microbiome model. Food Res. Int. 2022, 154, 110880. [Google Scholar] [CrossRef]
- Gil-Sánchez, I.; Cueva, C.; Tamargo, A.; Quintela, J.C.; de la Fuente, E.; Walker, A.W.; Moreno-Arribas, M.V.; Bartolomé, B. Application of the dynamic gastrointestinal simulator (simgi®) to assess the impact of probiotic supplementation in the metabolism of grape polyphenols. Food Res. Int. 2020, 129, 108790. [Google Scholar] [CrossRef]
- Freire, F.C.; Adorno, M.A.T.; Sakamoto, I.K.; Antoniassi, R.; Chaves, A.C.S.D.; Santos, K.M.O.; Sivieri, K. Impact of multi-functional fermented goat milk beverage on gut microbiota in a dynamic colon model. Food Res. Int. 2017, 99, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Van der Wielen, P.W.J.J.; Biesterveld, S.; Notermans, S.; Hofstra, H.; Urlings, B.A.P.; van Knapen, F. Role of volatile fatty acids in development of the cecal microflora in broiler chickens during growth. Appl. Environ. Microbiol. 2000, 66, 2536–2540. [Google Scholar] [CrossRef] [Green Version]
- Guillemard, E.; Poirel, M.; Schafer, F.; Quinquis, L.; Rossoni, C.; Keicher, C.; Wagner, F.; Szajewska, H.; Barbut, F.; Derrien, M.; et al. A Randomised, controlled trial: Effect of a multi-strain fermented milk on the gut microbiota recovery after Helicobacter pylori therapy. Nutrients 2021, 13, 3171. [Google Scholar] [CrossRef]
- Vallès, Y.; Artacho, A.; Pascual-García, A.; Ferrús, M.L.; Gosalbes, M.J.; Abellán, J.J.; Francino, M.P. Microbial succession in the gut: Directional trends of taxonomic and functional change in a birth cohort of Spanish infants. PLoS Genet. 2014, 10, e1004406. [Google Scholar] [CrossRef] [Green Version]
- Van Den Abbeele, P.; Sprenger, N.; Ghyselinck, J.; Marsaux, B.; Marzorati, M.; Rochat, F. A Comparison of the in vitro effects of 2’fucosyllactose and lactose on the composition and activity of gut microbiota from infants and toddlers. Nutrients 2021, 13, 726. [Google Scholar] [CrossRef]
- Wang, M.; Li, M.; Wu, S.; Lebrilla, C.B.; Chapkin, R.S.; Ivanov, I.; Donovan, S.M. Fecal microbiota composition of breast-fed infants is correlated with human milk oligosaccharides consumed. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 825–833. [Google Scholar] [CrossRef] [Green Version]
- Gamage, H.K.A.H.; Tetu, S.G.; Chong, R.W.W.; Ashton, J.; Packer, N.H.; Paulsen, I.T. Cereal products derived from wheat, sorghum, rice and oats alter the infant gut microbiota in vitro. Sci. Rep. 2017, 7, 14312. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Atiyeh, H.K.; Stevenson, B.S.; Tanner, R.S.; Wilkins, M.R.; Huhnke, R.L. Mixed culture syngas fermentation and conversion of carboxylic acids into alcohols. Bioresour. Technol. 2014, 152, 337–346. [Google Scholar] [CrossRef]
- Buckel, W. Unusual enzymes involved in five pathways of glutamate fermentation. Appl. Microbiol. Biotechnol. 2001, 57, 263–273. [Google Scholar] [CrossRef]
- Catto, C.; Garuglieri, E.; Borruso, L.; Erba, D.; Casiraghi, M.C.; Cappitelli, F.; Villa, F.; Zecchin, S.; Zanchi, R. Impacts of dietary silver nanoparticles and probiotic administration on the microbiota of an in-vitro gut model. Environ. Pollut. 2019, 245, 754–763. [Google Scholar] [CrossRef]
- Chaikham, P.; Apichartsrangkoon, A.; Worametrachanon, S.; van de Wiele, T. Impact of encapsulated Lactobacillus casei 01 along with pasteurized purple-rice drinks on modulating colon microbiome using a digestive model. Int. J. Food Engineer. 2016, 12, 637–646. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, R.; Li, X.; Li, J.; Ji, W.; Zeng, X.; Bao, J. Exposure to the environmental pollutant ammonia causes changes in gut microbiota and inflammatory markers in fattening pigs. Ecotoxicol. Environ. Saf. 2021, 208, 111564. [Google Scholar] [CrossRef]
- Mansilla, F.I.; Ficoseco, C.A.; Miranda, M.H.; Puglisi, E.; Nader-Macías, M.E.F.; Vignolo, G.M.; Fontana, C.A. Administration of probiotic lactic acid bacteria to modulate fecal microbiome in feedlot cattle. Nature 2022, 12, 12957. [Google Scholar] [CrossRef]
- Han, X.; Zhou, Q.; Gao, Z.; Lin, X.; Zhou, K.; Cheng, X.; Chitrakar, B.; Chen, H.; Zhao, W. In vitro digestion and fecal fermentation behaviors of polysaccharides from Ziziphus Jujuba cv. Pozao and its interaction with human gut microbiota. Food Res. Int. 2022, 162, 112022. [Google Scholar] [CrossRef]
- Fang, C.; Chen, G.; Kan, J. Characterization and in vitro simulated gastrointestinal digestion and fermentation of Mentha haplocalyx polysaccharide. Int. J. Biol. Macromol. 2022, 222, 360–372. [Google Scholar] [CrossRef]
- Terpend, K.; Possemiers, S.; Daguet, D.; Marzorati, M. Arabinogalactan and fructo-oligosaccharides have a different fermentation profile in the Simulator of the Human Intestinal Microbial Ecosystem (SHIME®). Environ. Microbiol. Rep. 2013, 5, 595–603. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Polic, I.I.; Matheyambath, A.C.; Lepointe, G. Modulation of human gut microbiota composition and metabolites by arabinogalactan and Bifidobacterium longum subsp. longum BB536 in the Simulator of the Human Intestinal Microbial Ecosystem (SHIME®). J. Funct. Foods 2021, 87, 104820. [Google Scholar] [CrossRef]
- Świątecka, D.; Dominika, Ś.; Narbad, A.; Arjan, N.; Ridgway, K.P.; Karyn, R.P. The study on the impact of glycated pea proteins on human intestinal bacteria. Int. J. Food Microbiol. 2011, 31, 267–272. [Google Scholar] [CrossRef]
- Raveschot, C.; Cudennec, B.; Coutte, F.; Flahaut, C.; Fremont, M.; Drider, D.; Dhulster, P. Production of bioactive peptides by Lactobacillus species: From gene to application. Front. Microbiol. 2018, 9, 2354. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Log CFU/mL | L. casei | L. plantarum | S. thermophilus |
|---|---|---|---|
| FMC | 9.95 ± 0.00 A | 9.87 ± 0.04 A | 3.45 ± 0.21 A |
| FMPF | 9.59 ± 0.02 B | 9.43 ± 0.09 B | 3.74 ± 0.06 A |
| FMB | 8.69 ± 0.01 C | 9.56 ± 0.03 B | 3.78 ± 0.00 A |
| Attribute | FM | FMPF | FMB |
|---|---|---|---|
| N. of nodes | 21 | 15 | 16 |
| N. of edges (+/−) | 32 (30/2) | 38 (21/17) | 33 (24/9) |
| N. of modules (sizes) | 3 (12, 7, 2) | 3 (11, 2, 2) | 2 (11, 5) |
| Modularity | 0.42 | 0.101 | 0.298 |
| Clustering coefficient | 0.809 | 0.848 | 0.75 |
| Mean degree | 3.048 | 5.067 | 4.125 |
| Max. degree | 6 (ID: 91) | 9 (IDs: 13, 40) | 8 (ID: 40) |
| Mean betweenness | 6.095 | 1.333 | 2.625 |
| Max. betweenness | 30 (IDs: 37, 63) | 12 (ID: 13) | 13.7 (ID: 40) |
| Main hubs | ID: 63 | IDs: 40 | ID: 40 |
| Periods | Acetic Acid | Propionic Acid | Butyric Acid | |
|---|---|---|---|---|
| Control | 22.35 ± 2.26 D | 3.21 ± 0.24 B | 14.03 ± 1.66 B | 557.22 ± 9.74 A |
| FMC | 110.67 ± 16.44 A | 1.74 ± 0.79 B | 24.56 ± 0.00 A | 492.11 ± 34.36 B |
| Washout FMC | 47.05 ± 3.31 CD | 1.01 ± 0.30 B | 11.24 ± 0.81 BC | 506.56 ± 29.43 AB |
| FMPF | 125.66 ± 10.07 A | 1.15 ± 0.00 B | 15.96 ± 0.16 B | 284.89 ± 6.04 D |
| Washout FMPF | 52.65 ± 2.79 CD | 3.79 ± 1.33 B | 7.72 ± 0.08 C | 496.57 ± 7.98 B |
| FMB | 108.90 ± 5.06 AB | 18.47 ± 3.32 A | 25.80 ± 1.87 A | 378.67 ± 6.15 C |
| Washout FMB | 71.04 ± 2.37 BC | 3.13 ± 0.98 B | 16.97 ± 0.79 B | 472.11 ± 6.27 B |
| Periods | Acetic Acid | Propionic Acid | Butyric Acid | |
|---|---|---|---|---|
| Control | 22.35 ± 2.26 D | 3.21 ± 0.24 B | 14.03 ± 1.66 B | 557.22 ± 9.74 A |
| FMC | 110.67 ± 16.44 A | 1.74 ± 0.79 B | 24.56 ± 0.00 A | 492.11 ± 34.36 B |
| Washout FMC | 47.05 ± 3.31 CD | 1.01 ± 0.30 B | 11.24 ± 0.81 BC | 506.56 ± 29.43 AB |
| FMPF | 125.66 ± 10.07 A | 1.15 ± 0.00 B | 15.96 ± 0.16 B | 284.89 ± 6.04 D |
| Washout FMPF | 52.65 ± 2.79 CD | 3.79 ± 1.33 B | 7.72 ± 0.08 C | 496.57 ± 7.98 B |
| FMB | 108.90 ± 5.06 AB | 18.47 ± 3.32 A | 25.80 ± 1.87 A | 378.67 ± 6.15 C |
| Washout FMB | 71.04 ± 2.37 BC | 3.13 ± 0.98 B | 16.97 ± 0.79 B | 472.11 ± 6.27 B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borgonovi, T.F.; Salgaço, M.K.; Oliveira, G.L.V.d.; Carvalho, L.A.L.d.; Pinheiro, D.G.; Todorov, S.D.; Sivieri, K.; Casarotti, S.N.; Penna, A.L.B. Functional Fermented Milk with Fruit Pulp Modulates the In Vitro Intestinal Microbiota. Foods 2022, 11, 4113. https://doi.org/10.3390/foods11244113
Borgonovi TF, Salgaço MK, Oliveira GLVd, Carvalho LALd, Pinheiro DG, Todorov SD, Sivieri K, Casarotti SN, Penna ALB. Functional Fermented Milk with Fruit Pulp Modulates the In Vitro Intestinal Microbiota. Foods. 2022; 11(24):4113. https://doi.org/10.3390/foods11244113
Chicago/Turabian StyleBorgonovi, Tais Fernanda, Mateus Kawata Salgaço, Gislane Lelis Vilela de Oliveira, Lucas Amoroso Lopes de Carvalho, Daniel Guariz Pinheiro, Svetoslav Dimitrov Todorov, Kátia Sivieri, Sabrina Neves Casarotti, and Ana Lúcia Barretto Penna. 2022. "Functional Fermented Milk with Fruit Pulp Modulates the In Vitro Intestinal Microbiota" Foods 11, no. 24: 4113. https://doi.org/10.3390/foods11244113