

Influence of Dietary Inulin on Fecal Microbiota, Cardiometabolic Risk Factors, Eicosanoids, and Oxidative Stress in Rats Fed a High-Fat Diet
 , , , , and
, , , , and
Abstract
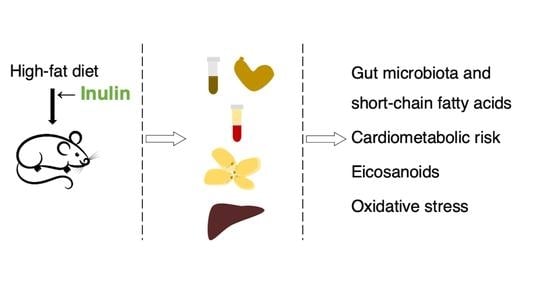
:
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Experimental Design
2.3. Feed Intake, Biometric Data, and Feces and Urine Collection
2.4. Sample Processing
2.5. Measurements of Biochemical Parameters
2.6. Histological Analysis of Perigonadal Adipose Tissue and Liver Samples
2.7. Analysis of Fecal Microbiota
2.8. Measurement of Short-Chain Fatty Acids in Feces and Cecal Content
2.9. Measurement of Eicosanoids in Liver
2.10. Measurements of Biomarkers of Oxidative Stress
2.10.1. Plasma Antioxidant Capacity
2.10.2. Antioxidant Enzymes and Glutathione
2.10.3. End Products of Lipid Peroxidation
2.11. Statistical Analysis
3. Results
3.1. Feed Intake, Biometric Data, and Feces and Urine
3.2. Biochemical Parameters
3.3. Histological Analysis of Perigonadal Adipose Tissue and Liver Samples
3.4. Analysis of Fecal Microbiota
3.5. Short-Chain Fatty Acids in Feces and Cecal Content
3.6. Eicosanoids in Liver
3.7. Biomarkers of Oxidative Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fabiani, R.; Naldini, G.; Chiavarini, M. Dietary patterns and metabolic syndrome in adult subjects: A systematic review and meta-analysis. Nutrients 2019, 11, 2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imamura, F.; Micha, R.; Wu, J.H.Y.; de Oliveira Otto, M.C.; Otite, F.O.; Abioye, A.I.; Mozaffarian, D. Effects of Saturated Fat, Polyunsaturated Fat, Monounsaturated Fat, and Carbohydrate on Glucose-Insulin Homeostasis: A Systematic Review and Meta-analysis of Randomised Controlled Feeding Trials. PLoS Med. 2016, 13, e1002087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Després, J.-P.; Lemieu, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Chassaing, B.; Singh, V.; Pellizzon, M.; Ricci, M.; Fythe, M.D.; Kumar, M.V.; Gewirtz, A.T. Fiber-Mediated Nourishment of Gut Microbiota Protects against Diet-Induced Obesity by Restoring IL-22-Mediated Colonic Health. Cell Host Microbe 2018, 23, 41–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Romero, S.; Hereu, M.; Atienza, L.; Casas, J.; Jáuregui, O.; Amézqueta, S.; Dasilva, G.; Medina, I.; Nogués, M.R.; Romeu, M.; et al. Mechanistically different effects of fat and sugar on insulin resistance, hypertension and gut microbiota in rats. Am. J. Physiol. Metab. 2018, 314, E552–E563. [Google Scholar] [CrossRef] [Green Version]
- Portune, K.J.; Benítez-Páez, A.; Del Pulgar, E.M.G.; Cerrudo, V.; Sanz, Y. Gut microbiota, diet, and obesity-related disorders—The good, the bad, and the future challenges. Mol. Nutr. Food Res. 2017, 61, 1600252. [Google Scholar] [CrossRef] [Green Version]
- Chassaing, B.; Miles-Brown, J.; Pellizzon, M.; Ulman, E.; Ricci, M.; Zhang, L.; Patterson, A.D.; Vijay-Kumar, M.; Gewirtz, A.T. Lack of soluble fiber drives diet-induced adiposity in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G528–G541. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Tuohy, F.F.; Chabo, K.M.C.; Waget, A.; et al. Metabolic Endotoxemia Initiates Obesity and Insulin Resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Nishiokada, A.; Miyoshi, M.; Fujiwara, M.; Aoyama-Ishikawa, M.; Maeshige, N.; Takahashi, M.; Hamada, Y.; Usami, Y.; Honda, M.; Arita, M.; et al. Changes of hepatic lipid mediators associated with intake of high-fat diet for 12 weeks in endotoxemic rats using LC-ESI-MS/MS. Clin. Nutr. 2015, 34, 685–693. [Google Scholar] [CrossRef]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot. Essent. Fat. Acids 2018, 132, 41–48. [Google Scholar] [CrossRef]
- Boyer, C.; Bannenberg, G.; Neve, E.; Ryrfeldt, A.; Moldéus, P. Evidence for the activation of the signal-responsive phospholipase A2 by exogenous hydrogen peroxide. Biochem. Pharmacol. 1995, 50, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Kaul, N.; Forman, H. Activation of NF kappa B by the respiratory burst of macrophages. Free Radic. Biol. Med. 1996, 21, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.N.O.; Makishima, M.; Matsuda, M.S.I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Jakubiak, G.K.; Osadnik, K.; Lejawa, M.; Osadnik, T.; Goławski, M.; Lewandowski, P.; Pawlas, N. “Obesity and insulin resistance” is the component of the metabolic syndrome most strongly associated with oxidative stress. Antioxidants 2022, 11, 79. [Google Scholar] [CrossRef]
- Anderson, E.J.; Lustig, M.E.; Boyle, K.E.; Woodlief, T.L.; Kane, D.A.; Lin, C.T.; Price, J.W.; Kang, L.; Rabinovitch, P.S.; Szeto, H.H.; et al. Mitochondrial H2O2 emission and cellular redox state link excess fat intake to insulin resistance in both rodents and humans. J. Clin. Investig. 2009, 119, 573–581. [Google Scholar] [CrossRef]
- Mozaffarian, D. Dietary and Policy Priorities for Cardiovascular Disease, Diabetes, and Obesity. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef]
- Ferrario, C.; Statello, R.; Carnevali, L.; Mancabelli, L.; Milani, C.; Mangifesta, M.; Duranti, S.; Lugli, G.A.; Jimenez, B.; Lodge, S.; et al. How to feed the Mammalian gut microbiota: Bacterial and metabolic modulation by dietary fibers. Front. Microbiol. 2017, 8, 1749. [Google Scholar] [CrossRef]
- Roberfroid, M.B. Introducing inulin-type fructans. Br. J. Nutr. 2005, 93, S13–S25. [Google Scholar] [CrossRef]
- Ahmed, W.; Rashid, S. Functional and therapeutic potential of inulin: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1–13. [Google Scholar] [CrossRef]
- Roberfroid, M.B. Caloric Value of Inulin and Oligofructose. J. Nutr. 1999, 129, 1436S–1437S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Beek, C.M.; Canfora, E.E.; Kip, A.M.; Gorissen, S.H.M.; Olde Damink, S.W.M.; van Eijk, H.M.; Holst, J.J.; Blaak, E.E.; Dejong, C.H.C.; Lenaerts, K. The prebiotic inulin improves substrate metabolism and promotes short-chain fatty acid production in overweight to obese men. Metabolism 2018, 87, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Weitkunat, K.; Stuhlmann, C.; Postel, A.; Rumberger, S.; Fankhänel, M.; Woting, A.; Petzke, K.J.; Gohlke, S.; Schulz, T.J.; Blaut, M.; et al. Short-chain fatty acids and inulin, but not guar gum, prevent diet-induced obesity and insulin resistance through differential mechanisms in mice. Sci. Rep. 2017, 7, 6109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Den Besten, G.; Bleeker, A.; Gerding, A.; Van Eunen, K.; Havinga, R.; Van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.J.; et al. Short-chain fatty acids protect against high-fat diet-induced obesity via a pparg-dependent switch from lipogenesis to fat oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitkunat, K.; Schumann, S.; Petzke, K.J.; Blaut, M.; Loh, G.; Klaus, S. Effects of dietary inulin on bacterial growth, short-chain fatty acid production and hepatic lipid metabolism in gnotobiotic mice. J. Nutr. Biochem. 2015, 26, 929–937. [Google Scholar] [CrossRef]
- González-Bosch, C.; Boorman, E.; Zunszain, P.A.; Mann, G.E. Short-chain fatty acids as modulators of redox signaling in health and disease. Redox Biol. 2021, 47, 102165. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Drabkin, D.L.; Austin, J.H. Spectrophotometric studies: II. Preparations from washed blood cells; nitric oxide hemoglobin and sulfhemoglobin. J. Biol. Chem. 1935, 112, 51–65. [Google Scholar] [CrossRef]
- Hernáez, Á.; Fernández-Castillejo, S.; Farràs, M.; Catalán, Ú.; Subirana, I.; Montes, R.; Solà, R.; Muñoz-Aguayo, D.; Gelabert-Gorgues, A.; Díaz-Gil, Ó.; et al. Olive oil polyphenols enhance high-density lipoprotein function in humans: A randomized controlled trial. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2115–2119. [Google Scholar] [CrossRef]
- Fortuño-Mar, À.; Pasquali, P. Cryobiopsy, Cryoanesthesia, and Cryoanalgesia. In Cryosurgery. A Practica Manual; Pasquali, P., Ed.; Springer: Heidelberg, Germany, 2015; pp. 85–91. [Google Scholar]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahti, L.; Shetty, S. Tools for Microbiome Analysis in R. Available online: http://microbiome.github.io/microbiome (accessed on 17 September 2021).
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. Available online: https://cran.r-project.org/package=vegan (accessed on 17 September 2021).
- Strassburg, K.; Mallet, C.; Duesterloh, A.; Bendik, I.; Hankemeier, T.; Langridge, J.; Vreeken, R.J.; Astarita, G. Targeted Lipidomics of Oxylipins (Oxygenated Fatty Acids). Waters Appl. Note 2015, 147, 20–30. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Cohen, G.; Dembiec, D.M.J. Measurement of catalase activity in tissue extracts. Anal. Biochem. 1970, 34, 30–38. [Google Scholar] [CrossRef]
- Wheeler, C.R.; Salzman, J.A.; Elsayed, N.M.; Omaye, S.T.; Korte, D.W., Jr. Automated assays for superoxide dismutase, catalase, glutathione peroxidase, and glutathione reductase activity. Anal. Biochem. 1990, 184, 193–199. [Google Scholar] [CrossRef]
- Hissin, P.J.; Hilf, R. A fluorometric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Buege, J.; Aust, S. Microsomal lipid peroxidation. Methods Enzym. 1978, 52, 302–310. [Google Scholar] [CrossRef]
- Richard, M.J.; Portal, B.; Meo, J.; Coudray, C.; Hadjian, A.; Favier, A. Malondialdehyde kit evaluated for determining plasma and lipoprotein fractions that react with thiobarbituric acid. Clin. Chem. 1992, 38, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Gérard-Monnier, D.; Erdelmeier, I.; Régnard, K.; Moze-Henry, N.; Yadan, J.C.; Chaudière, J. Reactions of 1-methyl-2-phenylindole with malondialdehyde and 4-hydroxyalkenals. Analytical applications to a colorimetric assay of lipid peroxidation. Chem. Res. Toxicol. 1998, 11, 1176–1183. [Google Scholar] [CrossRef]
- Jangra, S.; Raja, S.K.; Sharma, R.K.; Pothuraju, R.; Mohanty, A.K. Ameliorative effect of fermentable fibres on adiposity and insulin resistance in C57BL/6 mice fed a high-fat and sucrose diet. Food Funct. 2019, 10, 3696–3705. [Google Scholar] [CrossRef]
- Albouery, M.; Bretin, A.; Buteau, B.; Grégoire, S.; Martine, L.; Gambert, S.; Bron, A.M.; Acar, N.; Chassaing, B.; Bringer, M.A. Soluble fiber inulin consumption limits alterations of the gut microbiota and hepatic fatty acid metabolism caused by high-fat diet. Nutrients 2021, 13, 37. [Google Scholar] [CrossRef]
- Tan, S.; Caparros-Martin, J.A.; Matthews, V.B.; Koch, H.; O’Gara, F.; Croft, K.D.; Ward, N.C. Isoquercetin and inulin synergistically modulate the gut microbiome to prevent development of the metabolic syndrome in mice fed a high fat diet. Sci. Rep. 2018, 8, 10100. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Zapata, R.C.; Pezeshki, A.; Reidelberger, R.D.; Chelikani, P.K. Inulin fiber dose-dependently modulates energy balance, glucose tolerance, gut microbiota, hormones and diet preference in high-fat-fed male rats. J. Nutr. Biochem. 2018, 59, 142–152. [Google Scholar] [CrossRef]
- Lyon, M.R.; Kacinik, V. Is There a Place for Dietary Fiber Supplements in Weight Management? Curr. Obes. Rep. 2012, 1, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, J.D.; Yanckello, L.M.; Chlipala, G.; Hammond, T.C.; McCulloch, S.D.; Parikh, I.; Sun, S.; Morganti, J.M.; Green, S.J.; Lin, A.L. Dietary inulin alters the gut microbiome, enhances systemic metabolism and reduces neuroinflammation in an APOE4 mouse model. PLoS ONE 2019, 14, e0221828. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, K.; Maiorano, R.; Welch, R.M.; Miller, D.D.; Lei, X.G. Cecum is the major degradation site of ingested inulin in young pigs. J. Nutr. 2007, 137, 2399–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.W.; Cephas, K.D.; Holscher, H.D.; Kerr, K.R.; Mangian, H.F.; Tappenden, K.A.; Swanson, K.S. Nondigestible fructans alter gastrointestinal barrier function, gene expression, histomorphology, and themicrobiota profiles of diet-induced obese C57BL/6J Mice. J. Nutr. 2016, 146, 949–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, H.; Zhao, A.; Wang, Q.; Yang, X.; Ren, D. Supplementation of Inulin with Various Degree of Polymerization Ameliorates Liver Injury and Gut Microbiota Dysbiosis in High Fat-Fed Obese Mice. J. Agric. Food Chem. 2020, 68, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Li, D.; Zhao, J.; Liu, X.; Gu, Z.; Chen, Y.Q.; Zhang, H.; Chen, W. Metagenomic insights into the effects of fructo-oligosaccharides (FOS) on the composition of fecal microbiota in mice. J. Agric. Food Chem. 2015, 63, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Pachikian, B.; Taminiau, B.; Daube, G.; Frédérick, R.; Cani, P.D.; Bindels, L.B.; Delzenne, N.M. Intestinal sucrase as a novel target contributing to the regulation of glycemia by prebiotics. PLoS ONE 2016, 11, e0160488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozato, N.; Saito, S.; Yamaguchi, T.; Katashima, M.; Tokuda, I.; Sawada, K.; Katsuragi, Y.; Kakuta, M.; Imoto, S.; Ihara, K.; et al. Blautia genus associated with visceral fat accumulation in adults 20–76 years of age. NPJ Biofilm. Microbiomes 2019, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiel, S.; Gianfrancesco, M.A.; Rodriguez, J.; Portheault, D.; Leyrolle, Q.; Bindels, L.B.; da Silveira Cauduro, C.G.; Mulders, M.D.G.H.; Zamariola, G.; Azzi, A.S.; et al. Link between gut microbiota and health outcomes in inulin-treated obese patients: Lessons from the Food4Gut multicenter randomized placebo-controlled trial. Clin. Nutr. 2020, 39, 3618–3628. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; Havinga, R.; Bleeker, A.; Rao, S.; Gerding, A.; Van Eunen, K.; Groen, A.K.; Reijngoud, D.J.; Bakker, B.M. The short-chain fatty acid uptake fluxes by mice on a guar gum supplemented diet associate with amelioration of major biomarkers of the metabolic syndrome. PLoS ONE 2014, 9, e107392. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Brooks, L.; Viardot, A.; Tsakmaki, A.; Stolarczyk, E.; Howard, J.K.; Cani, P.D.; Everard, A.; Sleeth, M.L.; Psichas, A.; Anastasovskaj, J.; et al. Fermentable carbohydrate stimulates FFAR2-dependent colonic PYY cell expansion to increase satiety. Mol. Metab. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Molinar-Toribio, E.; Pérez-Jiménez, J.; Ramos-Romero, S.; Gómez, L.; Taltavull, N.; Nogués, M.R.; Adeva, A.; Jáuregui, O.; Joglar, J.; Clapés, P.; et al. D-Fagomine attenuates metabolic alterations induced by a high-energy-dense diet in rats. Food Funct. 2015, 6, 2614–2619. [Google Scholar] [CrossRef] [PubMed]
- Feillet-Coudray, C.; Sutra, T.; Fouret, G.; Ramos, J.; Wrutniak-Cabello, C.; Cabello, G.; Cristol, J.P.; Coudray, C. Oxidative stress in rats fed a high-fat high-sucrose diet and preventive effect of polyphenols: Involvement of mitochondrial and NAD(P)H oxidase systems. Free Radic. Biol. Med. 2009, 46, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, M.K.; Hallahan, N.L.; Brown, S.H.; Liu, M.; Mitchell, T.W.; Cooney, G.J.; Turner, N. Mouse strain-dependent variation in obesity and glucose homeostasis in response to high-fat feeding. Diabetologia 2013, 56, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Fabbrini, E.; Magkos, F.; Mohammed, B.S.; Pietka, T.; Abumrad, N.A.; Patterson, B.W.; Okunade, A.; Klein, S. Intrahepatic fat, not visceral fat, is linked with metabolic complications of obesity. Proc. Natl. Acad. Sci. USA 2009, 106, 15430–15435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasilva, G.; Lois, S.; Méndez, L.; Miralles-Pérez, B.; Romeu, M.; Ramos-Romero, S.; Torres, J.L.; Medina, I. Fish Oil Improves Pathway-Oriented Profiling of Lipid Mediators for Maintaining Metabolic Homeostasis in Adipose Tissue of Prediabetic Rats. Front. Immunol. 2021, 12, 608875. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, S.; Méndez, L.; Dasilva, G.; Torres, J.L.; Ramos-Romero, S.; Romeu, M.; Nogués, M.R.; Medina, I. Targeting hepatic protein carbonylation and oxidative stress occurring on diet-induced metabolic diseases through the supplementation with fish oils. Mar. Drugs 2018, 16, 353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Romero, S.; Hereu, M.; Atienza, L.; Casas, J.; Taltavull, N.; Romeu, M.; Amézqueta, S.; Dasilva, G.; Medina, I.; Torres, J.L. Functional Effects of the Buckwheat Iminosugar d-Fagomine on Rats with Diet-Induced Prediabetes. Mol. Nutr. Food Res. 2018, 62, 1800373. [Google Scholar] [CrossRef]
- Fischer, R.; Konkel, A.; Mehling, H.; Blossey, K.; Gapelyuk, A.; Wessel, N.; Von Schacky, C.; Dechend, R.; Muller, D.N.; Rothe, M.; et al. Dietary omega-3 fatty acids modulate the eicosanoid profi le in man primarily via the CYP-epoxygenase pathway. J. Lipid Res. 2014, 55, 1150–1164. [Google Scholar] [CrossRef] [Green Version]
- Jurado-Fasoli, L.; Di, X.; Kohler, I.; Osuna-Prieto, F.J.; Hankemeier, T.; Krekels, E.; Harms, A.C.; Yang, W.; Garcia-Lario, J.V.; Fernández-Veledo, S.; et al. Omega-6 and omega-3 oxylipins as potential markers of cardiometabolic risk in young adults. Obesity 2022, 30, 50–61. [Google Scholar] [CrossRef]
- Pickens, C.A.; Sordillo, L.M.; Zhang, C.; Fenton, J.I. Obesity is positively associated with arachidonic acid-derived 5- and 11-hydroxyeicosatetraenoic acid (HETE). Metabolism 2017, 70, 177–191. [Google Scholar] [CrossRef]
- Schmelzer, K.R.; Kubala, L.; Newman, J.W.; Kim, I.H.; Eiserich, J.P.; Hammock, B.D. Soluble epoxide hydrolase is a therapeutic target for acute inflammation. Proc. Natl. Acad. Sci. USA 2005, 102, 9772–9777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozak, W.; Aronoff, D.M.; Boutaud, O.; Kozak, A. 11,12-Epoxyeicosatrienoic acid attenuates synthesis of prostaglandin E2 in rat monocytes stimulated with lipopolysaccharide. Exp. Biol. Med. 2003, 228, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa-Nagata, N.; Takamura, T.; Ando, H.; Nakamura, S.; Kurita, S.; Misu, H.; Ota, T.; Yokoyama, M.; Honda, M.; Miyamoto, K.I.; et al. Increased oxidative stress precedes the onset of high-fat diet-induced insulin resistance and obesity. Metabolism 2008, 57, 1071–1077. [Google Scholar] [CrossRef] [Green Version]
- Rindler, P.M.; Plafker, S.M.; Szweda, L.I.; Kinter, M. High dietary fat selectively increases catalase expression within cardiac mitochondria. J. Biol. Chem. 2013, 288, 1979–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taltavull, N.; Miralles-Pérez, B.; Nogués, M.R.; Ramos-Romero, S.; Méndez, L.; Medina, I.; Torres, J.L.; Romeu, M. Effects of fish oil and grape seed extract combination on hepatic endogenous antioxidants and bioactive lipids in diet-induced early stages of insulin resistance in rats. Mar. Drugs 2020, 18, 318. [Google Scholar] [CrossRef]
- Renaud, H.J.; Cui, J.Y.; Lu, H.; Klaassen, C.D. Effect of diet on expression of genes involved in lipid metabolism, oxidative stress, and inflammation in mouse liver-insights into mechanisms of hepatic steatosis. PLoS ONE 2014, 9, e88584. [Google Scholar] [CrossRef]
- Huang, J.; Tabbi-Anneni, I.; Gunda, V.; Wang, L. Transcription factor Nrf2 regulates SHP and lipogenic gene expression in hepatic lipid metabolism. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Sohn, I.; Ahn, J.I.; Lee, K.H.; Lee, Y.S.; Lee, Y.S. Hepatic gene expression profiles in a long-term high-fat diet-induced obesity mouse model. Gene 2004, 340, 99–109. [Google Scholar] [CrossRef]
- Méndez, L.; Pazos, M.; Molinar-Toribio, E.; Sánchez-Martos, V.; Gallardo, J.M.; Rosa Nogués, M.; Torres, J.L.; Medina, I. Protein carbonylation associated to high-fat, high-sucrose diet and its metabolic effects. J. Nutr. Biochem. 2014, 25, 1243–1253. [Google Scholar] [CrossRef] [Green Version]
- Morshedi, M.; Valenlia, K.B.; Hosseinifard, E.S.; Shahabi, P.; Abbasi, M.M.; Ghorbani, M.; Barzegari, A.; Sadigh-Eteghad, S.; Saghafi-Asl, M. Beneficial psychological effects of novel psychobiotics in diabetic rats: The interaction among the gut, blood, and amygdala. J. Nutr. Biochem. 2018, 57, 145–152. [Google Scholar] [CrossRef]
- Jakubiak, G.K.; Osadnik, K.; Lejawa, M.; Kasperczyk, S.; Osadnik, T.; Pawlas, N. Oxidative Stress in Association with Metabolic Health and Obesity in Young Adults. Oxid. Med. Cell Longev. 2021, 2021, 9987352. [Google Scholar] [CrossRef] [PubMed]
- Maciejczyk, M.; Żebrowska, E.; Zalewska, A.; Chabowski, A. Redox balance, antioxidant defense, and oxidative damage in the hypothalamus and cerebral cortex of rats with high fat diet-induced insulin resistance. Oxid. Med. Cell Longev. 2018, 2018, 6940515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, G.; Prior, R.L. Comparison of different analytical methods for assessing total antioxidant capacity of human serum. Clin. Chem. 1998, 44, 1309–1315. [Google Scholar] [CrossRef] [PubMed]
- Adeyanju, O.A.; Badejogbin, O.C.; Areola, D.E.; Olaniyi, K.S.; Dibia, C.; Soetan, O.A.; Oniyide, A.A.; Michael, O.S.; Olatunji, L.A.; Soladoye, A.O. Sodium butyrate arrests pancreato-hepatic synchronous uric acid and lipid dysmetabolism in high fat diet fed Wistar rats. Biomed. Pharmacother. 2021, 133, 110994. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| STD | HF | HF + Inulin | |
|---|---|---|---|
| Feces | |||
| Week 5 | |||
| Acetic acid (mmol/kg) | 177 ± 111 | 63 ± 28 # | 144 ± 95 * |
| Propionic acid (mmol/kg) | 18 ± 11 | 4.2 ± 3.1 # | 34 ± 32 * |
| Butyric acid (mmol/kg) | 16 ± 14 | 3.8 ± 2.8 # | 26 ± 26 * |
| Isobutyric acid (mmol/kg) | 1.7 ± 1.0 | 0.72 ± 0.84 # | 0.26 ± 0.14 |
| Valeric acid (mmol/kg) | 2.0 ± 1.4 | 0.75 ± 0.64 # | 1.2 ± 1.2 |
| Isovaleric acid (mmol/kg) | 2.3 ± 1.4 | 1.2 ± 1.0 # | 0.80 ± 0.69 |
| Total SCFAs | 215 ± 130 | 56 ± 40 # | 189 ± 144 * |
| Week 9 | |||
| Acetic acid (mmol/kg) | 133 ± 57 | 29 ± 9 # | 38 ± 17 |
| Propionic acid (mmol/kg) | 14 ± 8 | 2.2 ± 1.6 # | 5.7 ± 3.6 * |
| Butyric acid (mmol/kg) | 28 ± 24 | 1.6 ± 1.8 # | 0.81 ± 0.75 |
| Isobutyric acid (mmol/kg) | 1.3 ± 0.7 | 0.23 ± 0.21 # | 0.15 ± 0.15 |
| Valeric acid (mmol/kg) | 1.4 ± 0.7 | 0.15 ± 0.07 # | 0.09 ± 0.06 |
| Isovaleric acid (mmol/kg) | 1.2 ± 0.5 | 0.28 ± 0.24 # | 0.32 ± 0.30 |
| Total SCFAs | 178 ± 84 | 31 ± 14 # | 40 ± 19 |
| Cecal content | |||
| Acetic acid (mmol/kg) | 110 ± 26 | 99 ± 35 | 100 ± 36 |
| Propionic acid (mmol/kg) | 32 ± 6 | 28 ± 8 | 28 ± 9 |
| Butyric acid (mmol/kg) | 20 ± 7 | 18 ± 9 | 18 ± 6 |
| Isobutyric acid (mmol/kg) | 3.0 ± 1.1 | 2.8 ± 0.8 | 3.3 ± 0.6 † |
| Valeric acid (mmol/kg) | 2.7 ± 1.4 | 2.4 ± 1.2 | 2.7 ± 1.4 |
| Isovaleric acid (mmol/kg) | 3.0 ± 1.3 | 2.6 ± 1.1 | 3.2 ± 0.7 |
| Total SCFAs | 171 ± 35 | 152 ± 53 | 156 ± 51 |
| STD | HF | HF + Inulin | |
|---|---|---|---|
| 5-HEPE (nmol/g tissue) | 0.150 ± 0.056 | 0.085 ± 0.030 # | 0.086 ± 0.059 |
| 11-HEPE (nmol/g tissue) | 0.30 ± 0.13 | 0.13 ± 0.05 # | 0.12 ± 0.06 |
| 5-HETE (nmol/g tissue) | 2.0 ± 0.6 | 1.2 ± 0.4 # | 1.0 ± 0.7 † |
| 20-HETE (nmol/g tissue) | 0.14 ± 0.02 | 0.08 ± 0.03 # | 0.08 ± 0.03 |
| 11(12)-EET (nmol/g tissue) | 0.22 ± 0.06 | 0.11 ± 0.03 # | 0.07 ± 0.02 * |
| 12-HETE (nmol/g tissue) | 2.0 ± 0.9 | 1.0 ± 0.5 # | 0.8 ± 0.6 |
| 15-HETE (nmol/g tissue) | 0.80 ± 0.28 | 0.33 ± 0.14 # | 0.29 ± 0.20 |
| 15-HETrE (nmol/g tissue) | 0.41 ± 0.11 | 0.41 ± 0.08 | 0.32 ± 0.08 * |
| PGD2 (nmol/g tissue) | 1.1 ± 0.4 | 0.32 ± 0.18 # | 0.33 ± 0.13 |
| PGE2 (nmol/g tissue) | 0.63 ± 0.28 | 0.23 ± 0.14 # | 0.31 ± 0.08 †† |
| Sum of HEPEs | 0.45 ± 0.18 | 0.22 ± 0.07 # | 0.21 ± 0.11 |
| Sum of HETEs | 5.0 ± 1.7 | 2.6 ± 1.0 # | 2.2 ± 1.6 † |
| 5-HEPE/5-HETE ratio | 0.076 ± 0.018 | 0.072 ± 0.013 | 0.084 ± 0.017 ††† |
| Sum of PGs | 1.71 ± 0.67 | 0.55 ± 0.31 # | 0.64 ± 0.19 |
| STD | HF | HF + Inulin | |
|---|---|---|---|
| Plasma | |||
| ORAC (µmol TE/mL) | 15 ± 6 | 18 ± 6 # | 15 ± 5 |
| FRAP (mmol TE/L) | 0.10 ± 0.02 | 0.10 ± 0.01 | 0.10 ± 0.01 † |
| GSH (nmol/mL) | 13 ± 3 | 17 ± 4 # | 20 ± 4 * |
| GSSG (nmol/mL) | 19 ± 5 | 22 ± 4 $ | 26 ± 4 †† |
| GSSG/GSH ratio | 1.5 ± 0.4 | 1.4 ± 0.4 | 1.3 ± 0.4 |
| TBARS (nmol MDA Eq/mL) | 2.2 ± 0.5 | 2.2 ± 0.4 | 1.8 ± 0.3 * |
| Erythrocytes | |||
| SOD (U/g Hb) | 2930 ± 630 | 3220 ± 440 | 2830 ± 350 ††† |
| CAT (mmol H2O2/min/g Hb) | 29 ± 4 | 32 ± 4 $$ | 34 ± 4 |
| GPx (U/g Hb) | 72 ± 12 | 82 ± 12 $$$ | 92 ± 16 |
| GR (U/g Hb) | 1.2 ± 0.2 | 0.9 ± 0.2 # | 0.8 ± 0.2 |
| TBARS (nmol MDA Eq/g Hb) | 0.46 ± 0.16 | 0.63 ± 0.06 # | 0.63 ± 0.28 |
| STD | HF | HF + Inulin | |
|---|---|---|---|
| Perigonadal adipose tissue | |||
| SOD (U/g tissue) | 64 ± 18 | 55 ± 20 | 45 ± 20 |
| CAT (nmol H2O2/min/g tissue) | 24 ± 14 | 25 ± 9 | 30 ± 7 |
| GPx (U/g tissue) | 0.69 ± 0.36 | 0.60 ± 0.24 | 0.67 ± 0.14 |
| GR (U/g tissue) | 0.34 ± 0.10 | 0.24 ± 0.07 # | 0.28 ± 0.05 |
| GSH (nmol/g tissue) | 7.0 ± 2.7 | 7.7 ± 3.3 | 8.1 ± 3.8 |
| GSSG (nmol/g tissue) | 28 ± 7 | 28 ± 8 | 28 ± 10 |
| GSSG/GSH ratio | 4.4 ± 1.6 | 4.1 ± 1.9 | 3.8 ± 3.0 |
| TBARS (nmol MDA Eq/g tissue) | 3.0 ± 1.5 | 1.2 ± 0.5 # | 0.9 ± 0.6 |
| Liver | |||
| SOD (U/g tissue) | 5970 ± 1460 | 6090 ± 1270 | 5200 ± 1510 |
| CAT (mmol H2O2/min/g tissue) | 7.3 ± 1.6 | 7.6 ± 1.0 | 8.2 ± 1.0 |
| GPx (U/g tissue) | 42 ± 3 | 59 ± 5 # | 57 ± 6 |
| GR (U/g tissue) | 8.6 ± 2.4 | 7.5 ± 2.0 | 8.5 ± 1.8 |
| GSH (µmol/g tissue) | 1.8 ± 0.5 | 1.6 ± 0.2 $ | 1.6 ± 0.4 |
| GSSG (µmol/g tissue) | 1.2 ± 0.3 | 1.2 ± 0.3 | 1.3 ± 0.4 |
| GSSG/GSH ratio | 0.75 ± 0.36 | 0.79 ± 0.27 | 0.88 ± 0.54 |
| MDA + 4-HAE (nmol MDA Eq/g tissue) | 29 ± 6 | 26 ± 4 # | 29 ± 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miralles-Pérez, B.; Nogués, M.R.; Sánchez-Martos, V.; Fortuño-Mar, À.; Ramos-Romero, S.; Torres, J.L.; Ponomarenko, J.; Amézqueta, S.; Zhang, X.; Romeu, M. Influence of Dietary Inulin on Fecal Microbiota, Cardiometabolic Risk Factors, Eicosanoids, and Oxidative Stress in Rats Fed a High-Fat Diet. Foods 2022, 11, 4072. https://doi.org/10.3390/foods11244072
Miralles-Pérez B, Nogués MR, Sánchez-Martos V, Fortuño-Mar À, Ramos-Romero S, Torres JL, Ponomarenko J, Amézqueta S, Zhang X, Romeu M. Influence of Dietary Inulin on Fecal Microbiota, Cardiometabolic Risk Factors, Eicosanoids, and Oxidative Stress in Rats Fed a High-Fat Diet. Foods. 2022; 11(24):4072. https://doi.org/10.3390/foods11244072
Chicago/Turabian StyleMiralles-Pérez, Bernat, Maria Rosa Nogués, Vanessa Sánchez-Martos, Àngels Fortuño-Mar, Sara Ramos-Romero, Josep L. Torres, Julia Ponomarenko, Susana Amézqueta, Xiang Zhang, and Marta Romeu. 2022. "Influence of Dietary Inulin on Fecal Microbiota, Cardiometabolic Risk Factors, Eicosanoids, and Oxidative Stress in Rats Fed a High-Fat Diet" Foods 11, no. 24: 4072. https://doi.org/10.3390/foods11244072