
The Effects of Starch Molecular Fine Structure on Thermal and Digestion Properties of Rice Starch

Abstract
:1. Introduction
2. Overview of Rice Starch Structural Levels and Their Characterization
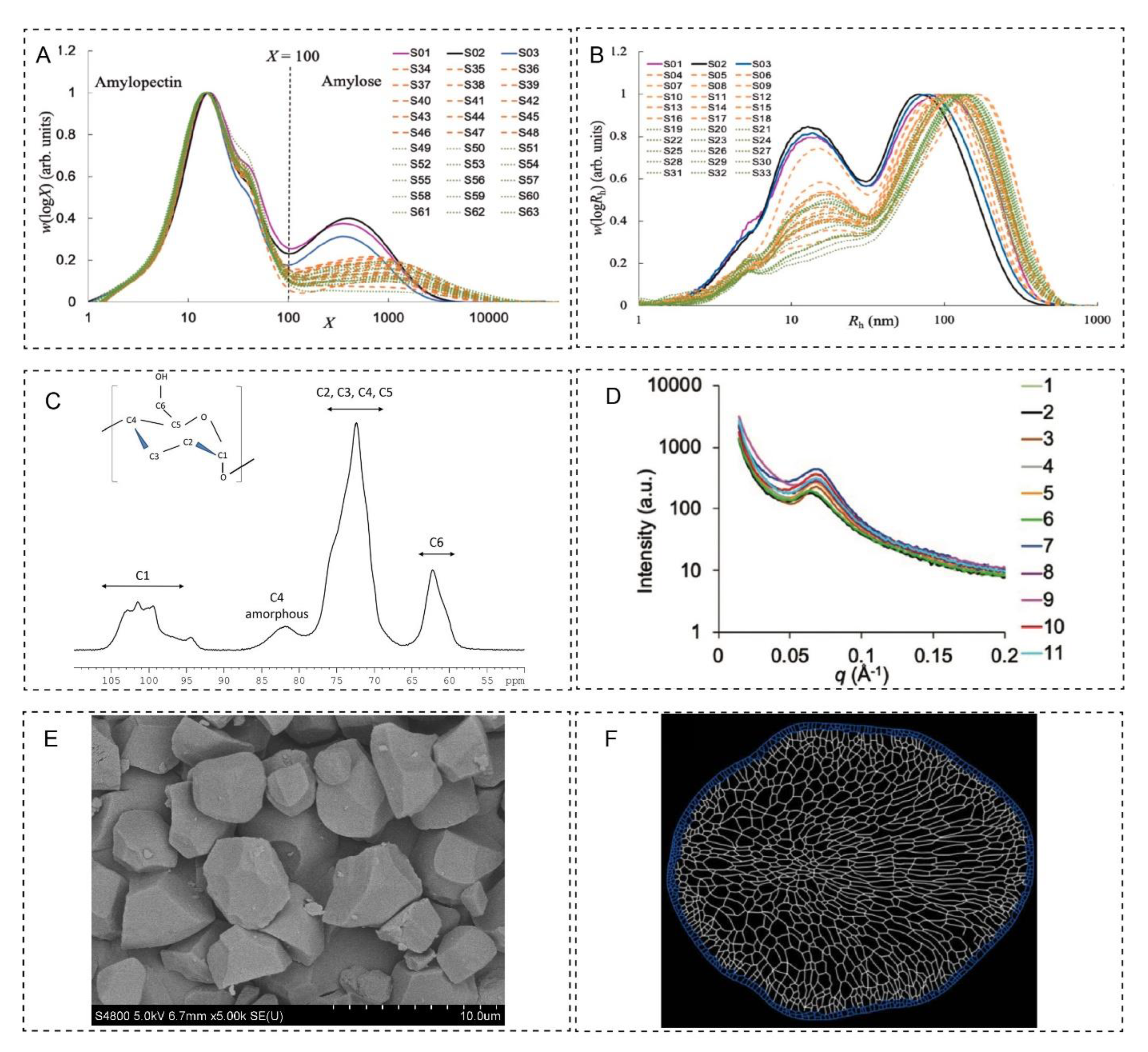
2.1. First Level—Chain-Length Distribution

2.2. Second Level—Molecular Size Distribution
2.3. Third Level—Nanocrystals
2.4. Fourth Level—Growth Rings
2.5. Fifth Level—Granules
2.6. Sixth Level—Cellular Structure
3. Starch Order-Disorder Structural Changes during Conventional Cooking of Whole Rice
3.1. Starch Fine Molecular Structure and Gelatinization during Cooking
3.2. Importance of Starch Fine Molecular Structures for Starch Retrogradation during Cooling of Cooked Dehulled Rice Grains
4. Digestibility Measurements and Starch Digestibility in Cooked Whole Rice
4.1. Digestion Kinetics

{kind=link}
| Kinetics Model | Equation | Parameters | Advantages and Applications | Disadvantages and Limitations | Reference |
|---|---|---|---|---|---|
| Single first-order kinetics (SK) | C: starch digested amount at infinite time; k is a pseudo-first-order rate constant. | Being able to quantify the whole digestogram into a single rate constant. | Less capable of fitting to multi-phase starch digestograms. | [81] | |
| Logarithm-of-slope plot (LOS) | ln | Parameters are the same as those of the SK model. | Being able to identify the number of starch digestible fractions and fit digestograms with a sequential digestion pattern. | Only suitable for fitting to multi-phase starch digestograms with a sequential digestion pattern. | [84] |
| Parallel first-order kinetics (PK) | C0 is the starch digested amount at t = 0; C1∞ and C2∞ are the maximum starch digested amount at very long time for digestible fractions 1 and 2, respectively; k1 and k2 are rate constants for the two digestible fractions, respectively. | Being able to fit digestograms with a parallel digestion pattern. | Only suitable for fitting to multi-phase starch digestograms with a parallel digestion pattern. | [87] | |
| Combination of parallel and sequential kinetics (CPS) | The parameters C0, C1∞, C2∞, k1 and k2 are similar to that of PK model. t2start is the time when the digestion of slowly digestible fraction commences. | Being able to identify the digestion pattern of different starch digestible fractions. | Less capable of identifying the number of starch digestible fractions | [65] | |
| Consecutive reactions kinetics (CRK) | For the starch substrate: CSS(t) = exp(−kbt) CDP(∞). | is the starch digested ratio after infinite digestion time; kb and kcat are the rate constants for the formation of enzyme-substrate complex and of digestion product, respectively. | Being able to differentiate the enzymatic binding and catalysis step in starch digestograms. | n.a. | [88] |
| For the enzyme-starch complex: | |||||
| For the digested starch product: |
4.2. The Importance of Starch Molecular Fine Structure for Starch Digestibility of Cooked Whole Rice
5. Future Directions
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, G.; Hamaker, B.R. Slowly digestible starch: Concept, mechanism, and proposed extended glycemic index. Crit. Rev. Food Sci. Nutr. 2009, 49, 852–867. [Google Scholar] [CrossRef] [PubMed]
- DeMartino, P.; Cockburn, D.W. Resistant starch: Impact on the gut microbiome and health. Curr. Opin. Biotechnol. 2019, 61, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Bede, D.; Lou, Z.X. Recent developments in resistant starch as a functional food. Starch-Starke 2021, 73, 2000139. [Google Scholar] [CrossRef]
- Esquivel-Fajardo, E.A.; Martinez-Ascencio, E.U.; Oseguera-Toledo, M.E.; Londono-Restrepo, S.M.; Rodriguez-Garcia, M.E. Influence of physicochemical changes of the avocado starch throughout its pasting profile: Combined extraction. Carbohydr. Polym. 2022, 281, 119048. [Google Scholar] [CrossRef]
- Li, C.; Hu, Y.; Gu, F.; Gong, B. Causal relations among starch fine molecular structure, lamellar/crystalline structure and in vitro digestion kinetics of native rice starch. Food Funct. 2021, 12, 682–695. [Google Scholar] [CrossRef]
- Van Hung, P.; Chau, H.T.; Phi, N.T. In vitro digestibility and in vivo glucose response of native and physically modified rice starches varying amylose contents. Food Chem. 2016, 191, 74–80. [Google Scholar] [CrossRef]
- Slaughter, S.L.; Ellis, P.R.; Butterworth, P.J. An investigation of the action of porcine pancreatic alpha-amylase on native and gelatinised starches. Biochim. Biophys. Acta 2001, 1525, 29–36. [Google Scholar] [CrossRef]
- Sivakamasundari, S.K.; Priyanga, S.; Moses, J.A.; Anandharamakrishnan, C. Impact of processing techniques on the glycemic index of rice. Crit. Rev. Food Sci. Nutr. 2021, 62, 3323–3344. [Google Scholar] [CrossRef]
- Sun, J.; Wang, Y.; Zhang, X.Q.; Rasmussen, S.K.; Jiang, X.T.; Song, W.J.; Wu, D.X.; Shu, X.L. Dependence of physiochemical, functional and textural properties of high-resistant starch rice on endogenous nonstarch polysaccharides. Int. J. Food Sci. Tech. 2018, 53, 1079–1086. [Google Scholar] [CrossRef]
- Li, C.; Hamaker, B.R. Effects of different storage temperatures on the intra- and intermolecular retrogradation and digestibility of sago starch. Int. J. Biol. Macromol. 2021, 182, 65–71. [Google Scholar] [CrossRef]
- Li, C.; Hu, Y. Antagonistic effects of amylopectin and amylose molecules on the starch inter- and intramolecular interactions during retrogradation. LWT 2021, 148, 111942. [Google Scholar] [CrossRef]
- Gong, B.; Cheng, L.; Gilbert, R.G.; Li, C. Distribution of short to medium amylose chains are major controllers of in vitro digestion of retrograded rice starch. Food Hydrocolloid 2019, 96, 634–643. [Google Scholar] [CrossRef]
- Klostermann, C.E.; Buwalda, P.L.; Leemhuis, H.; Vos, P.d.; Schols, H.A.; Bitter, J.H. Digestibility of resistant starch type 3 is affected by crystal type, molecular weight and molecular weight distribution. Carbohydr. Polym. 2021, 265, 118069. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Zaragoza, E.; Sanchez-Zapata, E.; Sendra, E.; Sayas, E.; Navarro, C.; Fernandez-Lopez, J.; Perez-Alvarez, J.A. Resistant starch as prebiotic: A review. Starch-Starke 2011, 63, 406–415. [Google Scholar] [CrossRef]
- Li, C.; Wu, A.; Yu, W.; Hu, Y.; Li, E.; Zhang, C.; Liu, Q. Parameterizing starch chain-length distributions for structure-property relations. Carbohydr. Polym. 2020, 241, 116390. [Google Scholar] [CrossRef] [PubMed]
- Nada, S.S.; Zou, W.; Li, C.F.; Gilbert, R.G. Parameterizing amylose chain-length distributions for biosynthesis-structure-property relations. Anal. Bioanal. Chem. 2017, 409, 6813–6819. [Google Scholar] [CrossRef]
- Wu, A.C.; Morell, M.K.; Gilbert, R.G. A parameterized model of amylopectin synthesis provides key insights into the synthesis of granular starch. PLoS One 2013, 8, e65768. [Google Scholar] [CrossRef]
- Yi, X.; Li, E.; Yu, S.; Zhang, X.; Yang, C.; Shao, S.; Gilbert, R.G.; Li, C. Combined effects of starch fine molecular structures and water content on starch digestibility of cooked white rice. Int. J. Biol. Macromol. 2022, 215, 192–202. [Google Scholar] [CrossRef]
- Li, C. Consecutive reaction kinetics model reveals the nature of long-term rice amylopectin retrogradation characteristics. Food Chem. 2022, 369, 131000. [Google Scholar] [CrossRef]
- Martinez, M.M.; Li, C.; Okoniewska, M.; Mukherjee, I.; Vellucci, D.; Hamaker, B. Slowly digestible starch in fully gelatinized material is structurally driven by molecular size and A and B1 chain lengths. Carbohydr. Polym. 2018, 197, 531–539. [Google Scholar] [CrossRef]
- Bertoft, E.; Annor, G.A.; Shen, X.; Rumpagaporn, P.; Seetharaman, K.; Hamaker, B.R. Small differences in amylopectin fine structure may explain large functional differences of starch. Carbohydr. Polym. 2016, 140, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Zhang, B.; Tan, C.P.; Fu, X.; Huang, Q. Effects of limited moisture content and storing temperature on retrogradation of rice starch. Int. J. Biol. Macromol. 2019, 137, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, S.; Ren, X.; Lu, Y.; Liu, D.; Cai, X.; Li, Q.; Gao, J.; Liu, Q. Molecular structure and physicochemical properties of starches from rice with different amylose contents resulting from modification of OsGBSSI activity. J. Agric. Food Chem. 2017, 65, 2222–2232. [Google Scholar] [CrossRef]
- Li, C.; Gong, B. Relations between rice starch fine molecular and lamellar/ crystalline structures. Food Chem. 2021, 353, 129467. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hu, Y.; Zhang, B. Plant cellular architecture and chemical composition as important regulator of starch functionality in whole foods. Food Hydrocolloid 2021, 117, 106744. [Google Scholar] [CrossRef]
- O’Shea, M.G.; Samuel, M.S.; Konik, C.M.; Morell, M.K. Fluorophore-assisted carbohydrate electrophoresis (FACE) of oligosaccharides—Efficiency of labelling and high-resolution separation. Carbohydr. Res. 1998, 307, 1–12. [Google Scholar] [CrossRef]
- Broberg, S.; Koch, K.; Andersson, R.; Kenne, L. A comparison between MALDI-TOF mass spectrometry and HPAEC-PAD analysis of debranched starch. Carbohydr. Polym. 2000, 43, 285–289. [Google Scholar] [CrossRef]
- Jones, R.G.; Kahovec, J.; Stepto, R.; Wilks, E.S.; Hess, M.; Kitayama, T.; Metanomski, W.V. Compendium of Polymer Terminology and Nomenclature. IUPAC Recommendations 2008; Royal Society of Chemistry: Cambridge, UK, 2009. [Google Scholar] [CrossRef] [Green Version]
- Wu, A.C.; Li, E.; Gilbert, R.G. Exploring extraction/dissolution procedures for analysis of starch chain-length distributions. Carbohydr. Polym. 2014, 114, 36–42. [Google Scholar] [CrossRef]
- Rohrer, J. Chapter 8—A practical guide for carbohydrate determinations by high-performance anion-exchange chromatography with pulsed amperometric detection (HPAE-PAD). In Separation Science and Technology; Pohl, C., Avdalovic, N., Srinivasan, K., Eds.; Academic Press: Cambridge, MA, USA, 2021; Volume 13, pp. 233–257. [Google Scholar]
- Wong, K.S.; Jane, J. Effects of pushing agents on the separation and detection of debranched amylopectin by high-performance anion-exchange chromatography with pulsed amperometric detection. J. Liq. Chromatogr. 1995, 18, 63–80. [Google Scholar] [CrossRef]
- Vilaplana, F.; Hasjim, J.; Gilbert, R.G. Amylose content in starches: Toward optimal definition and validating experimental methods. Carbohydr. Polym. 2012, 88, 103–111. [Google Scholar] [CrossRef]
- Fitzgerald, M.A.; Bergman, C.J.; Resurreccion, A.P.; Muller, J.; Jimenez, R.; Reinke, R.F.; Martin, M.; Blanco, P.; Molina, F.; Chen, M.-H.; et al. Addressing the dilemmas of measuring amylose in rice. Cereal Chem. 2009, 86, 492–498. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Gilbert, R.G. Starch molecular structure: The basis for an improved understanding of cooked rice texture. Carbohydr. Polym. 2018, 195, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yu, W.; Zhang, C.; Zhu, Y.; Xu, J.; Li, E.; Gilbert, R.G.; Liu, Q. New insights into amylose and amylopectin biosynthesis in rice endosperm. Carbohydr. Polym. 2020, 230, 115656. [Google Scholar] [CrossRef]
- Yu, W.; Li, H.; Zou, W.; Tao, K.; Zhu, J.; Gilbert, R.G. Using starch molecular fine structure to understand biosynthesis-structure-property relations. Trends in Food Science & Technology 2019, 86, 530–536. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Henry, R.J.; Gilbert, R.G. Starch structure-property relations in Australian wild rices compared to domesticated rices. Carbohydr Polym 2021, 271, 118412. [Google Scholar] [CrossRef] [PubMed]
- Katoh, E.; Murata, K.; Fujita, N. (13)C CP/MAS NMR can discriminate genetic backgrounds of rice starch. ACS Omega 2020, 5, 24592–24600. [Google Scholar] [CrossRef]
- Zhao, L.; Cai, C.; Wei, C. An image processing method for investigating the morphology of cereal endosperm cells. Biotech. Histochem. 2019, 95, 249–261. [Google Scholar] [CrossRef]
- Runyon, J.R.; Ulmius, M.; Nilsson, L. A perspective on the characterization of colloids and macromolecules using asymmetrical flow field-flow fractionation. Colloids Surf. A Physicochem. Eng. Asp. 2014, 442, 25–33. [Google Scholar] [CrossRef]
- Cave, R.A.; Seabrook, S.A.; Gidley, M.J.; Gilbert, R.G. Characterization of starch by size-exclusion chromatography: The limitations imposed by shear scission. Biomacromolecules 2009, 10, 2245–2253. [Google Scholar] [CrossRef]
- Vilaplana, F.; Gilbert, R.G. Two-dimensional size/branch length distributions of a branched polymer. Macromolecules 2010, 43, 7321–7329. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, K.; Hasjim, J.; Li, E.; Flanagan, B.M.; Gidley, M.J.; Dhital, S. Freeze-drying changes the structure and digestibility of B-polymorphic starches. J. Agric. Food Chem. 2014, 62, 1482–1491. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Zhao, S.; Zhang, B.; Tan, M.; Niu, M.; Jia, C.; Huang, Q. Understanding the supramolecular structures and pasting features of adlay seed starches. Food Hydrocolloid 2018, 83, 411–418. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, M.E.; Hernandez-Landaverde, M.A.; MiguelDelgado, J.; Ramirez-Gutierrez, C.F.; Ramirez-Cardona, M.; Millan-Malo, B.M.; Londoño-Restrepo, S.M. Crystalline structures of the main components of starch. Curr. Opin. Food Sci. 2021, 37, 107–111. [Google Scholar] [CrossRef]
- Witt, T.; Doutch, J.; Gilbert, E.P.; Gilbert, R.G. Relations between molecular, crystalline, and lamellar structures of amylopectin. Biomacromolecules 2012, 13, 4273–4282. [Google Scholar] [CrossRef]
- Doutch, J.; Gilbert, E.P. Characterisation of large scale structures in starch granules via small-angle neutron and X-ray scattering. Carbohydr. Polym. 2013, 91, 444–451. [Google Scholar] [CrossRef]
- Zhang, B.; Gilbert, E.P.; Qiao, D.; Xie, F.; Wang, D.K.; Zhao, S.; Jiang, F. A further study on supramolecular structure changes of waxy maize starch subjected to alkaline treatment by extended-q small-angle neutron scattering. Food Hydrocolloid 2019, 95, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Chi, C.; Wu, C. How to calculate starch lamellar features with improved accuracy by small angle X-ray scattering. Int. J. Biol. Macromol. 2019, 141, 622–625. [Google Scholar] [CrossRef]
- Gong, B.; Liu, W.; Tan, H.; Yu, D.; Song, Z.; Lucia, L.A. Understanding shape and morphology of unusual tubular starch nanocrystals. Carbohydr. Polym. 2016, 151, 666–675. [Google Scholar] [CrossRef]
- Zhang, Z.; Fan, X.; Yang, X.; Li, C.; Gilbert, R.G.; Li, E. Effects of amylose and amylopectin fine structure on sugar-snap cookie dough rheology and cookie quality. Carbohydr. Polym. 2020, 241, 116371. [Google Scholar] [CrossRef]
- Waigh, T.A.; Gidley, M.J.; Komanshek, B.U.; Donald, A.M. The phase transformations in starch during gelatinisation: A liquid crystalline approach. Carbohydr. Res. 2000, 328, 165–176. [Google Scholar] [CrossRef]
- Jenkins, P.J.; Donald, A.M. Gelatinisation of starch: A combined SAXS/ WAXS/DSC and SANS study. Carbohydr. Res. 1998, 308, 133–147. [Google Scholar] [CrossRef]
- Wani, A.A.; Singh, P.; Shah, M.A.; Schweiggert-Weisz, U.; Gul, K.; Wani, I.A. Rice starch diversity: Effects on structural, morphological, thermal, and physicochemical properties—A Review. Compr. Rev. Food Sci. Food Saf. 2012, 11, 417–436. [Google Scholar] [CrossRef]
- Hanashiro, I.; Abe, J.-i.; Hizukuri, S. A periodic distribution of the chain length of amylopectin as revealed by high-performance anion-exchange chromatography. Carbohydr. Res. 1996, 283, 151–159. [Google Scholar] [CrossRef]
- Tao, K.Y.; Li, C.; Yu, W.W.; Gilbert, R.G.; Li, E.P. How amylose molecular fine structure of rice starch affects functional properties. Carbohydr. Polym. 2019, 204, 24–31. [Google Scholar] [CrossRef]
- Kong, X.; Bertoft, E.; Bao, J.; Corke, H. Molecular structure of amylopectin from Amaranth starch and its effect on physicochemical properties. Int. J. Biol. Macromol. 2008, 43, 377–382. [Google Scholar] [CrossRef]
- Zhu, F. Relationships between amylopectin internal molecular structure and physicochemical properties of starch. Trends Food Sci. Technol. 2018, 78, 234–242. [Google Scholar] [CrossRef]
- Zhu, F.; Corke, H.; Bertoft, E. Amylopectin internal molecular structure in relation to physical properties of sweetpotato starch. Carbohydr. Polym. 2011, 84, 907–918. [Google Scholar] [CrossRef]
- Noda, T.; Isono, N.; Krivandin, A.V.; Shatalova, O.V.; Błaszczak, W.; Yuryevc, V. Origin of defects in assembled supramolecular structures of sweet potato starches with different amylopectin chain-length distribution. Carbohydr. Polym. 2009, 76, 400–409. [Google Scholar] [CrossRef]
- Nakamura, Y.; Sakurai, A.; Inaba, Y.; Kimura, K.; Iwasawa, N.; Nagamine, T. The fine structure of amylopectin in endosperm from Asian cultivated rice can be largely classified into two classes. Starch-Starke 2002, 54, 117–131. [Google Scholar] [CrossRef]
- Li, C.; Gong, B. Insights into chain-length distributions of amylopectin and amylose molecules on the gelatinization property of rice starches. Int. J. Biol. Macromol. 2020, 155, 721–729. [Google Scholar] [CrossRef]
- Vamadevan, V.; Bertoft, E.; Seetharaman, K. On the importance of organization of glucan chains on thermal properties of starch. Carbohydr. Polym. 2013, 92, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Cao, P.; Cao, W.; Li, C. Relations between starch fine molecular structures with gelatinization property under different moisture content. Carbohydrate Polymers 2022, 278, 118955. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hu, Y. Combination of parallel and sequential digestion kinetics reveals the nature of digestive characteristics of short-term retrograded rice starches. Food Hydrocolloid 2020, 108, 106071. [Google Scholar] [CrossRef]
- Li, C.; Gong, B.; Huang, T.; Yu, W.W. In vitro digestion rate of fully gelatinized rice starches is driven by molecular size and amylopectin medium-long chains. Carbohydr. Polym. 2021, 254, 117275. [Google Scholar] [CrossRef]
- Cooke, D.; Gidley, M.J. Loss of crystalline and molecular order during starch gelatinisation: Origin of the enthalpic transition. Carbohydr. Res. 1992, 227, 103–112. [Google Scholar] [CrossRef]
- Ball, S.G.; van de Wal, M.H.B.J.; Visser, R.G.F. Progress in understanding the biosynthesis of amylose. Trends Plant Sci. 1998, 3, 462–467. [Google Scholar] [CrossRef]
- Zhong, Y.; Liu, L.; Qu, J.; Blennow, A.; Hansen, A.R.; Wu, Y.; Guo, D.; Liu, X. Amylose content and specific fine structures affect lamellar structure and digestibility of maize starches. Food Hydrocolloid 2020, 108, 105994. [Google Scholar] [CrossRef]
- Wang, S.J.; Li, C.L.; Copeland, L.; Niu, Q.; Wang, S. Starch retrogradation: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 568–585. [Google Scholar] [CrossRef]
- Li, C.; Luo, J.; Zhang, C.; Yu, W. Causal relations among starch chain-length distributions, short-term retrogradation and cooked rice texture. Food Hydrocolloid 2020, 108, 106064. [Google Scholar] [CrossRef]
- Perdon, A.A.; Siebenmorgen, T.J.; Buescher, R.W.; Gbur, E.E. Starch retrogradation and texture of cooked milled rice during storage. J. Food Sci. 1999, 64, 828–832. [Google Scholar] [CrossRef]
- Cui, R.; Oates, C.G. The effect of retrogradation on enzyme susceptibility of sago starch. Carbohydr. Polym. 1997, 32, 65–72. [Google Scholar] [CrossRef]
- Zhang, M.; Sun, C.; Wang, X.; Wang, N.; Zhou, Y. Effect of rice protein hydrolysates on the short-term and long-term retrogradation of wheat starch. Int. J. Biol. Macromol. 2020, 155, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Mclver, R.G.; Axford, D.W.E.; Colwell, K.H.; Elton, G.A.H. Kinetic study of the retrogradation of gelatinised starch. J. Sci. Food Agric. 1968, 19, 560–563. [Google Scholar] [CrossRef]
- Jankowski, T.; Rha, C.K. Retrogradation of Starch in Cooked Wheat. Starch-Starke 1986, 38, 6–9. [Google Scholar] [CrossRef]
- Klucinec, J.D.; Thompson, D.B. Amylose and amylopectin interact in retrogradation of dispersed high-amylose starches. Cereal Chem. 1999, 76, 282–291. [Google Scholar] [CrossRef]
- Ottenhof, M.A.; Farhat, I.A. Starch retrogradation. Biotechnol. Genet. Eng. Rev. 2004, 21, 215–228. [Google Scholar] [CrossRef] [Green Version]
- Han, K.T.; Kim, H.R.; Moon, T.W.; Choi, S.J. Isothermal and temperature-cycling retrogradation of high-amylose corn starch: Impact of sonication on its structural and retrogradation properties. Ultrason. Sonochem. 2021, 76, 105650. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Sopade, P.A. Modeling starch digestograms: Computational characteristics of kinetic models for in vitro starch digestion in food research. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1422–1445. [Google Scholar] [CrossRef] [Green Version]
- Goñi, I.; Garcia-Alonso, A.; Saura-Calixto, F. A starch hydrolysis procedure to estimate glycemic index. Nutr. Res. 1997, 17, 427–437. [Google Scholar] [CrossRef]
- Dhital, S.; Shrestha, A.K.; Gidley, M.J. Relationship between granule size and in vitro digestibility of maize and potato starches. Carbohydr. Polym. 2010, 82, 480–488. [Google Scholar] [CrossRef]
- Zhang, B.; Dhital, S.; Gidley, M.J. Synergistic and Antagonistic Effects of alpha-Amylase and Amyloglucosidase on Starch Digestion. Biomacromolecules 2013, 14, 1945–1954. [Google Scholar] [CrossRef] [PubMed]
- Butterworth, P.J.; Warren, F.J.; Grassby, T.; Patel, H.; Ellis, P.R. Analysis of starch amylolysis using plots for first-order kinetics. Carbohydrate Polymers 2012, 87, 2189–2197. [Google Scholar] [CrossRef]
- Yu, W.; Zhou, X.; Li, C. Application of first-order kinetics modeling to reveal the nature of starch digestion characteristics. Food Funct. 2021, 12, 6652–6663. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Zou, W.; Dhital, S.; Wu, P.; Gidley, M.J.; Fox, G.P.; Gilbert, R.G. The adsorption of alpha-amylase on barley proteins affects the in vitro digestion of starch in barley flour. Food Chem. 2018, 241, 493–501. [Google Scholar] [CrossRef]
- Li, H.; Dhital, S.; Gidley, M.J.; Gilbert, R.G. A more general approach to fitting digestion kinetics of starch in food. Carbohydr. Polym. 2019, 225, 115244. [Google Scholar] [CrossRef]
- Li, C.; Hu, Y. Modeling of in vitro digestogram by consecutive reaction kinetics model reveals the nature of starch digestive characteristics. Food Hydrocolloid 2022, 124, 107203. [Google Scholar] [CrossRef]
- Chung, H.J.; Lim, H.S.; Lim, S.T. Effect of partial gelatinization and retrogradation on the enzymatic digestion of waxy rice starch. J. Cereal Sci. 2006, 43, 353–359. [Google Scholar] [CrossRef]
- Martinez, M.M.; Roman, L.; Gomez, M. Implications of hydration depletion in the in vitro starch digestibility of white bread crumb and crust. Food Chem. 2018, 239, 295–303. [Google Scholar] [CrossRef]
- Pletsch, E.A.; Hamaker, B.R. Brown rice compared to white rice slows gastric emptying in humans. Eur. J. Clin. Nutr. 2018, 72, 367–373. [Google Scholar] [CrossRef]
- Gu, F.; Li, C.; Hamaker, B.R.; Gilbert, R.G.; Zhang, X. Fecal microbiota responses to rice RS3 are specific to amylose molecular structure. Carbohydr. Polym. 2020, 243, 116475. [Google Scholar] [CrossRef]
- Patel, H.; Royall, P.G.; Gaisford, S.; Williams, G.R.; Edwards, C.H.; Warren, F.J.; Flanagan, B.M.; Ellis, P.R.; Butterworth, P.J. Structural and enzyme kinetic studies of retrograded starch: Inhibition of alpha-amylase and consequences for intestinal digestion of starch. Carbohydr. Polym. 2017, 164, 154–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, X.; Li, C. Main controllers for improving the resistant starch content in cooked white rice. Food Hydrocolloid 2022, 122, 107083. [Google Scholar] [CrossRef]
- Li, C.; Cao, P.; Wu, P.; Yu, W.; Gilbert, R.G.; Li, E. Effects of endogenous proteins on rice digestion during small intestine (in vitro) digestion. Food Chem. 2021, 344, 128687. [Google Scholar] [CrossRef]
- Quek, R.; Henry, C.J. Influence of polyphenols from lingonberry, cranberry, and red grape on in vitro digestibility of rice. Int. J. Food Sci. Nutr. 2015, 66, 378–382. [Google Scholar] [CrossRef] [PubMed]
| Level | Type of Structure | Main Parameters | Range | Reference |
|---|---|---|---|---|
| 1st | Individual chains | Amylopectin CLD | DP ≤ 100 | [12] |
| Amylose CLD | DP > 100 | |||
| 2nd | Whole starch molecules | Amylopectin Rh | Average Rh 66.1–83.8 nm | [12] |
| Amylose Rh | Average Rh 13.4–17.6 nm | |||
| 3rd | Double helices | FTIR 1045 cm−1/1022 cm−1 | 0.59 to 0.67 | [23] |
| 4th | Growth rings | d | 8.62–9.11 nm | [24] |
| da | 3.63–3.75 nm | |||
| dc | 4.99–5.35 nm | |||
| 5th | Granules | Granule size | 2–8 μm | [23] |
| 6th | Cells and grains | Spatial distribution of cellular components. | [25] | |
| Cell wall architecture and chemical compositions; | n.a. | |||
| Protein matrix; | ||||
| Lipids and polyphenols |
| Property | Relations to Starch Fine Molecular Structures | Proposed Mechanisms | References |
|---|---|---|---|
| Gelatinization | Longer amylopectin A and B1 chains result in higher gelatinization temperatures. | Longer amylopectin A and B1 chains form longer and more stable double helices. | [56,57] |
| The amount of amylopectin Acltr chains is positively associated with starch gelatinization temperature. | Amylopectin Acltr chains preferentially form double helices. | [61] | |
| Gelatinization conclusion temperature is associated with the amount of amylopectin trans-lamellar chains (DP ~37–69). | Amylopectin trans-lamellar chains can form long amylopectin double helices. | [62] | |
| Amylopectin inter-block chain length is positively correlated with starch gelatinization onset temperature. | Longer amylopectin internal chain segments allow a more ordered alignment of amylopectin double helices. | [63] | |
| AC has an approximately parabolic relationship with rice starch gelatinization temperatures. | Either a low or high AC can promote the formation of more ordered amylopectin double helices. | [64] | |
| Longer amylose intermediate chains can increase the gelatinization temperature range of rice starches. | Longer amylose intermediate chains can interact with amylopectin molecules into double helices. | [24,62] | |
| Retrogradation | AC > 20 % can increase the nucleation rate for long-term amylopectin retrogradation. | Crystallites formed by amylose provide crystal seeds for amylopectin long-term retrogradation. | [19] |
| Amylose with a larger molecular size promotes the nucleation rate for short-term rice starch retrogradation. | Larger amylose molecules have a higher mobility and denser nucleation sites. | [65] | |
| Longer amylopectin external chains and smaller amylopectin molecules can have both inter- and intramolecular interactions. | Longer amylopectin external chains and smaller amylopectin molecules have higher flexibility. | [20] | |
| Digestion | Larger rice amylopectin molecules have a faster digestion rate at the fully gelatinized state. | Larger amylopectin molecules have shorter branches, supplying more non-reducing ends for amyloglucosidase. | [66] |
| Rice starches with more short to intermediate amylose chains have a slower starch digestion rate in the retrograded state. | Amylose short to intermediate chains are prone to retrograde. | [12] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Yu, W.; Gilbert, R.G. The Effects of Starch Molecular Fine Structure on Thermal and Digestion Properties of Rice Starch. Foods 2022, 11, 4012. https://doi.org/10.3390/foods11244012
Li C, Yu W, Gilbert RG. The Effects of Starch Molecular Fine Structure on Thermal and Digestion Properties of Rice Starch. Foods. 2022; 11(24):4012. https://doi.org/10.3390/foods11244012
Chicago/Turabian StyleLi, Cheng, Wenwen Yu, and Robert G. Gilbert. 2022. "The Effects of Starch Molecular Fine Structure on Thermal and Digestion Properties of Rice Starch" Foods 11, no. 24: 4012. https://doi.org/10.3390/foods11244012