3.1. Identification of Isolates
After purifying, 15 isolates were obtained and examined by morphological and biochemical identification. The preliminary assays were: Gram staining and catalase activity. Afterward, 8 g positive and catalase-negative strains were selected as potential LAB. The morphology of the isolates was examined by using a light microscope, and the cocci and bacillus-shaped isolates were selected for further assays. According to the results obtained from physiological, morphological, and biochemical assays, bacteria were classified based on their similarities and differences. Eight selected isolates were identified to genus level, based on biochemical assays: Group 1: Homofermentative bacilli that grew at 10 °C and pH = 4.4, but were unable to hydrolyze arginine, this group was identified as homofermentative lactobacilli; Group 2: Gram-positive, catalase-negative, homofermentative cocci with tetrad arrangement and unable to hydrolyze arginine. They grew at 10 °C, but not at 45 °C and pH = 9.6, so were considered pediococcus species; Group 3: The isolates of
Lactococcus that could grow at 10 °C and hydrolyze arginine; Group 4: Gram-positive, catalase-negative and homofermentative cocci which grew at both 10 and 45 °C, in the presence of 6.5% of NaCl at pH = 9.6 were identified as Enterococcus spp. (
Table 1) [
17]. Therefore, the identified isolates included Lactobacillus, Pediococcus, Lactococcus, and
Enterococcus species.
The results of 16S rRNA gene analysis showed that the bacterial isolates were
Enterococcus faecium KKP 3772 (KF1),
Enterococcus faecium C1 (KF2),
Pediococcus pentosaceus H11(KF3),
Lactococcus lactis Rsg (KK1),
Enterococcus faecalis P190052 (KK2),
Enterococcus mundtii CECT972T( KK3),
Pediococcus pentosaceus VNK-1 (KK4), and
Lpb. plantarum PM411 (KK5) (
Table 2). Among these bacteria, the first three isolates were obtained from Abadeh (Fars) kashk samples and the others were isolated from Kalat kashk samples (Razavi Khorasan).
Lactococcus lactis is an anaerobic homofermentative bacterium that produces large amounts of lactic acid (L
+) during growth and metabolic activity. This organism is also able to produce lactic acid (D
-) at a lower pH, and this ability is one of the reasons for the wide application of
Lactococcus lactis in dairy industries [
25]. Edalatian et al. (2012), stated that the Lighvan cheese microbiota comprises a consortium of Enterococci, including
Enterococcus faecalis and
Enterococcus faecium species as the dominant bacteria. The cause of the large population of Enterococcus bacteria in ripened Lighvan cheese is their tolerance to high salt concentrations and low pH. In addition to Enterococci,
Lpb. plantarum was found in high concentrations in Lighvan cheese [
17].
Enterococcus species are present in many fermented milk products. These bacteria have positive effects on the quality of Roquefort cheeses and increase the growth in other LAB. Specific strains of
Enterococcus have shown probiotic potential such as resistance to a wide range of pH and temperature, bacteriocin production, etc. [
26].
3.2. Cell Survival in Low pH Conditions, Simulated Digestive System, and Bile Salts
According to the definition by the World Health Organization (WHO), probiotics must be consumed in sufficient amounts (at least 10
7 CFU/mL) to show their beneficial effects. Conditions such as the acidic pH of the environment can hinder metabolism and decrease the growth and survival of LAB [
27]. Acids in the human stomach destroy biomolecules such as fatty acids, proteins, vitamins, and nucleic acids. Stomach cells secrete about 2 L of acidic gastric juices per day, which create strict conditions for the survival of microorganisms that pass through the stomach. The results of evaluating the resistance of lactic acid isolates to acidic conditions (pH 2 and 3) and gastric juices are presented in
Table 3.
Different trends were observed in the survival of the isolates in acidic pH. As shown in
Table 3, incubation at pH 2 for 3 h, reduced the bacterial growth by 3 to 6 logarithmic cycles. While the isolates remained unchanged at pH 3, except
Lactococcus lactis, which showed about 1.5 logarithmic cycle loss in the cell viability after 3 h of incubation. The survival of LAB strains in the range of pH 2 to 4 is an essential factor to perform as potential probiotics [
28,
29].
In agreement with our results, Ding and Shah (2007) investigated the effect of pH ≥ 2.5 on the reduction of 8 different species of Lactobacillus. The findings indicated the high sensitivity of bacterial cells to low pH [
30]. In another study, Parente et al. (2010), examined the activity of two enzymes, arginine deaminase, and glutamate decarboxylase, in
Lpb. plantarum, when exposed to different concentrations of acid. They found that the activity level of two enzymes increased with the increase in acid concentration; however, this increase in activity level had a downward trend at pH ≤ 3 [
31].
F
1F
0-ATPase is one of the main mechanisms to protect Gram-positive microorganisms against acidic conditions. Other factors, such as growth medium composition, cell membrane composition, nutritional compounds, and type of microorganism can affect pH resistance [
32]. Azat et al. investigated the pH resistance of LAB isolated from traditionally fermented Xinjiang cheese and stated that all tested isolates were tolerant to acidic and bile salt conditions, as the viable population of all strains was found to be more than 10
6 CFU/mL after incubation at pH 3 for 3 h [
33].
To investigate the probiotic potential of an isolate, it is essential to assess its tolerance to bile salts. Bile is a digestive secretion that breaks down large lipid droplets into smaller ones, causes emulsification, and finally the digestion of fat molecules. All bacteria have a cell wall containing fatty acids that can be destroyed by bile salts [
34]. The isolates that are resistant to high concentrations of bile salts, can survive and grow in the normal concentration of bile in the human gastrointestinal tract. The secretion of bile salts into the duodenum directly disrupts the growth of probiotic bacteria. The detergent properties of bile salts enable them to penetrate into bacterial cell membranes, disrupting their structure and changing cell homeostasis [
35]. The cow bile used in this study contained conjugated and non-conjugated bile salts such as taurocholic acid, glycolic acid (hydroxyacetic acid), lectin, and cholesterol. The results of the resistance of the isolates to different concentrations (0.3, 0.5, and 1%) of bile salts and non-conjugated bile salts are shown in
Table 4. KK3 and KF2 were the most susceptible isolates to the bile salts. In the 0.3% of bile salts, the cell viability of KK3 and KF2 was 7.90 ± 0.14% and 8.1 ± 0.47%, respectively, and at higher concentrations, isolates did not grow.
The highest cell viability at bile salts was observed in KK4 and KF3 isolates. The resistance and sensitivity of isolates to conjugated bile salts (TC: sodium taurocholate, TDC: sodium taurodeoxycholate, GC: sodium glycolate, GDC: Sodium glicodeoxycholate) were also investigated (
Table 4). The results showed that isolates of
Lactococcus lactis RSg (KK1) and
Enterococcus faecium KKP 3772 (KF1) were resistant against all non-conjugated bile salts and could grow. Exceptionally,
Enterococcus faecium C1 (KF2) had weak growth in TC.
Lpb. plantarum PM411 (KK5) and Pediococcus pentosaceus H11 (KF3) were resistant to the conjugated bile salts, except for GDC, but Pediococcus pentosaceus H11 (KF3) did not show deconjugation activity against TC and TDC. Enterococcus faecalis P190052 (KK2) was resistant to all of the deconjugated bile salts and showed the ability to deconjugate GDC and TDC. Enterococcus mundtii CECT972T (KK3) was susceptible to all compounds and showed only weak growth in the presence of GC. Lpb. plantarum PM411 (KK5) was resistant in the media containing TC, TDC, and GC bile salts, but weak growth was observed in media containing GDC. Generally, among all isolates, Enterococcus mundtii CECT972T (KK3) and Enterococcus faecium C1 (KF2) were the most sensitive isolates to bile salts. While Pediococcus pentosaceus VNK-1 (KK4) and Pediococcus pentosaceus H11 (KF3) showed the highest resistance.
3.3. Antibiotic Susceptibility
Table 5 displays the antibiotic resistance of bacterial isolates against 16 different antibiotics. Based on the results obtained, all isolates were resistant to tetracycline and also all of them were susceptible to chloramphenicol and Gentamicin. Most of the isolated LAB were susceptible to vancomycin and only
Lpb. plantarum PM411 (KK5) and
Pediococcus pentosaceus VNK-1 (KK4), showed moderate resistance. Among the tested isolates, only
Enterococcus faecium C1 (KF2) was resistant to cefixime and sulfamethoxazole.
Pediococcus pentosaceus VNK-1 (KK4) showed resistance only to cetracycline, cefepime, and clindamycin antibiotics. Generally, there are two main mechanisms for the antibiotic resistance of probiotics: (1) natural or intrinsic resistance, which is not transferable, (2) acquired resistance, usually caused by bacterial mutation, which might transfer plasmid encoding of antibiotic resistance genes and is potentially transferable to pathogenic bacteria or other commensals [
36].
The European Federation of Animal Science (EFAS) and the European Food Safety Authority (EFSA) recommend that the strains of LAB which are used in different medicines and foods, should not have transferable antibiotic resistance genes to be considered safe for human and animal consumption [
37]. Antibiotic resistance may have negative effects on probiotics. For instance, antibiotic resistance genes can be transferred to pathogenic or intestinal bacteria and make them resistant to antibiotic treatment [
38,
39]. Gupta and Sharma (2017) stated
Pediococcus acidilactici Ch-2 was susceptible to 11 out of 12 antibiotics. Therefore, this isolate can be considered a safe probiotic strain [
40].
3.4. Antibacterial Activity
Antimicrobial activity is one of the important characteristics to evaluate the probiotic potential of a microorganism. The antibacterial activity of probiotics can be due to the synthesis of H
2O
2, ethanol, phenols, diacetyl, proteins, and organic acids such as acetic and lactic acids that are produced during the growth of probiotics. These metabolites, together with help of a competitive exclusion mechanism, in which probiotics compete with harmful microorganisms for adhesive receptors and nutrients, can destroy and inhibit the colonization of pathogens in the body [
41]. In this study, the effect of pH, temperature, and enzyme on antimicrobial activity was determined. The results of the antimicrobial effect of the isolates are presented in
Table 6. The diameters of the inhibition zones on the culture medium of pathogen strains were measured as a criterion for measuring the antimicrobial activity of the isolates. Comparing the antimicrobial activity of different isolates showed that the antimicrobial activity of
Pediococcus pentosaceus VNK-1 (KK4) and
Pediococcus pentosaceus H11 (KF3) was higher than other strains and their inhibition zones were larger.
Enterococcus mundtii CECT972T (KK3) and
Lpb. plantarum PM411 (KK5) showed the smallest inhibition zone in different treatments.
Overall, the obtained results revealed that the isolates had different antimicrobial activities. Some isolates showed low, + (11–15 mm); while others had moderate, ++ (15–20 mm) and high +++, (>21 mm) inhibition zones. The treatment of isolates with catalase enzyme did not change, their inhibition rate except for Lpb. plantarum PM411 (KK5), which presented a decreased inhibition zone diameter and lower antibacterial activity. Heat treatment only decreased the antimicrobial activity of Enterococcus mundtii CECT972T (KK3) and Lactococcus lactis RSg (KK1). Comparison between untreated and treated samples in neutral pH conditions showed that the majority of the isolates did not exhibit antimicrobial activity, and only KK4, KF3, and KK5 isolates had poor inhibition activity against pathogen strains. KK4 isolate displayed antimicrobial activity against all of the examined pathogens. Generally, the highest antimicrobial activity of all isolates was observed against Staphylococcus aureus ATCC 25923 while the lowest activity was detected against Pseudomonas aeruginosa PTCC 1707.
3.5. Cell Surface Hydrophobicity and Auto-Aggregation
The cell surface hydrophobicity and auto-aggregation are phenotypic characteristics that are directly attributed to the adhesion ability of bacteria. Cell surface hydrophobicity is the nonspecific interaction between host and bacterial cells. Solvents such as xylene, chloroform, n-octane, n-hexadecane, and ethyl acetate are used to measure this feature [
18,
42]. This characteristic is supposed to be a major factor in determining the capability of LAB to adhere to the intestinal cells and their consequent proliferation [
43]. The adhesion depends on some factors, such as van der Waals force, Brownian motion, electric charge of the surface, and gravitational force. Additionally, surface hydrophobicity depends on the type of bacteria, so this characteristic should be exanimated separately for each strain. The s-layer, which consists of a single molecular layer of identical proteins or glycoproteins, plays a major role in the Brownian movement of LAB [
44].
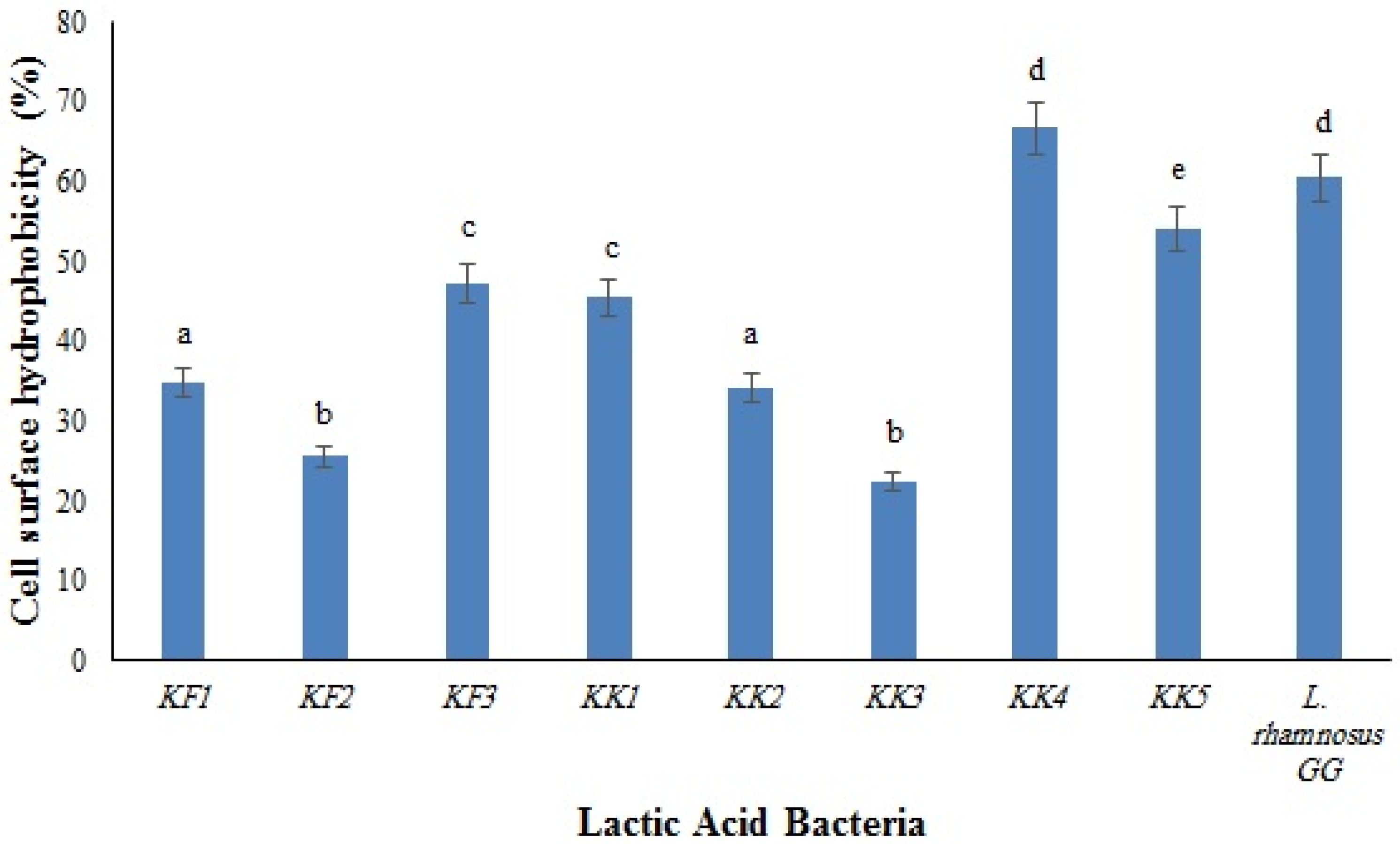
The results of the surface hydrophobicity of the isolates are shown in
Figure 1. The highest and the lowest hydrophobicity were observed in
Pediococcus pentosaceus VNK-1 (KK4) (66.7%) and in
Enterococcus mundtii CECT972T (KK3) (22.4%), respectively. The cell surface of microorganisms contains hydrophobic compounds such as proteins, teichoic acids, and lipids, which make them attach to the surface of the intestinal epithelium through covalent bonds. The differences between the cell surface hydrophobicity of bacteria are influenced by several parameters, such as the chemical composition and structural properties of bacteria (type of amino acids, composition of proteins, polysaccharides, and lipid compounds in the bacterial cells), the growth phase of bacteria, and environmental factors [
18].
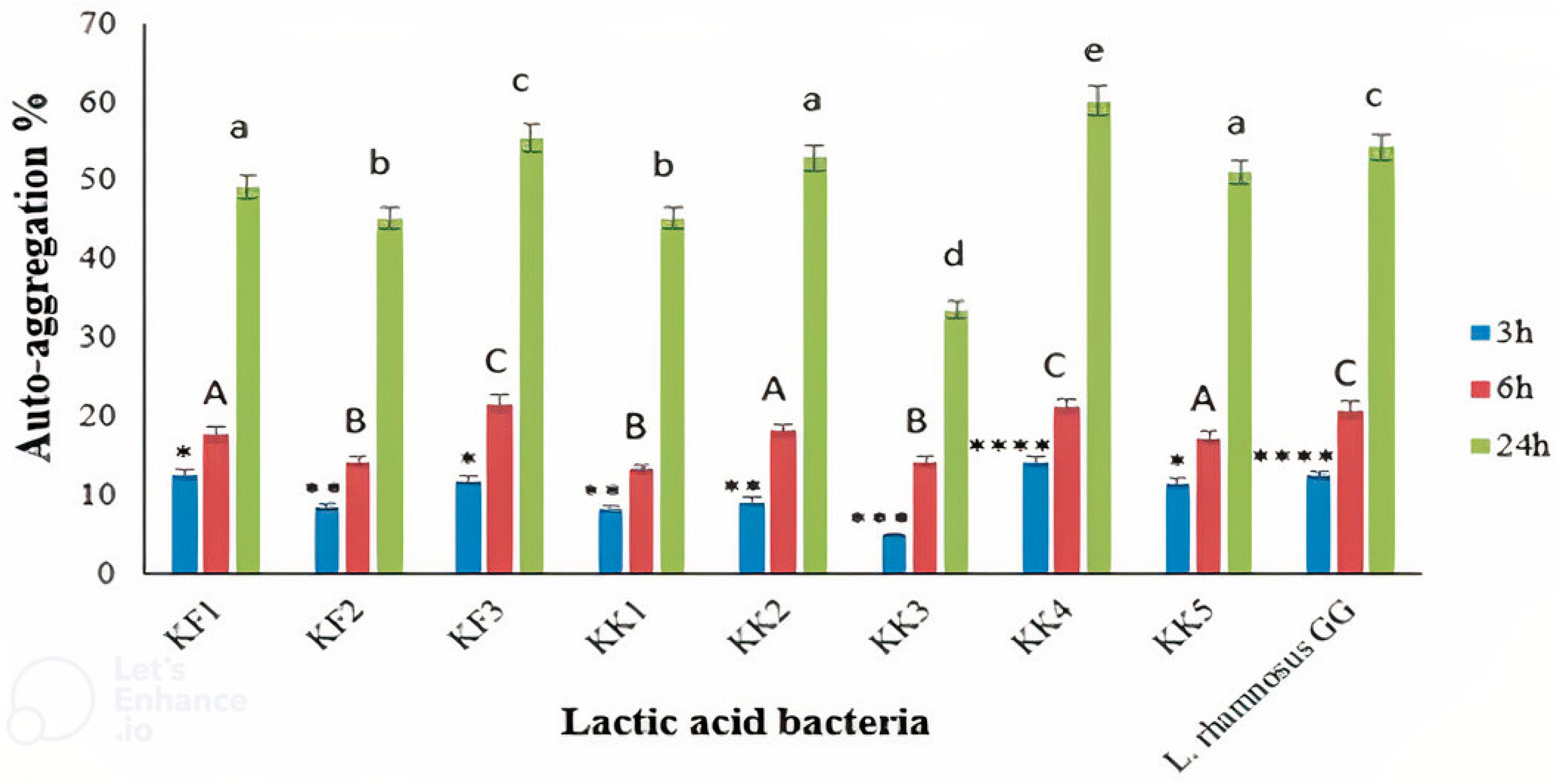
The results showed the auto-aggregation abilities of all the LAB isolates (
Figure 2). After 24 h of incubation, auto-aggregation of the LAB strains was between 33.60% and 60.20%. The highest and lowest auto-aggregation were detected in
Pediococcus pentosaceus VNK-1 (KK4) and
Enterococcus mundtii CECT972T (KK3), respectively. The auto-aggregation rate of
Pediococcus pentosaceus VNK-1 (KK4) (60.20%) was higher than that of
Lacticaseibacillus GG (54.3%), as the control strain. Indicating that this bacterium probably has better cell adhesion properties than
L. rhamnosus GG. The amount of auto-accumulation was progressively increased during 24 h of incubation. Our findings are in accordance with previous studies [
21,
45,
46].





{kind=link}
{kind=link}