
Anti-Aging Effect of Agrocybe aegerita Polysaccharide through Regulation of Oxidative Stress and Gut Microbiota
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Fly Strains and Diet
2.3. Lifespan Assay
2.4. Hydrogen Peroxide (H2O2) Challenge Assay
2.5. Animals and Treatment
2.6. Biochemical Analysis
2.7. Intestinal Microbiota Analysis
2.8. Data Analysis
3. Results
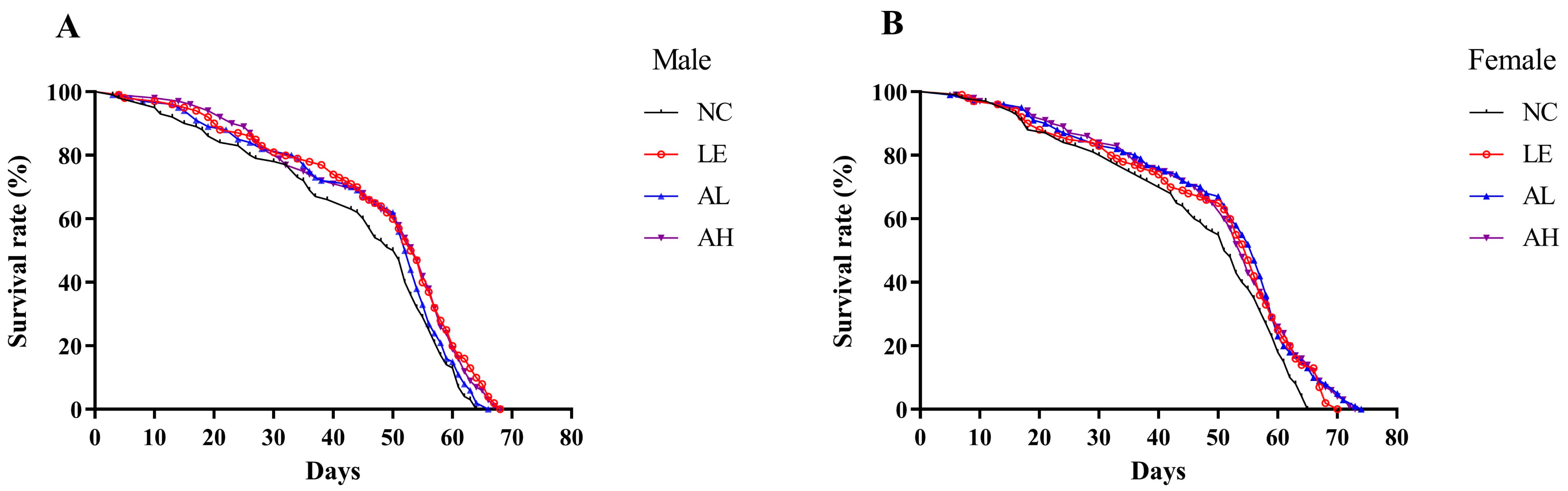
3.1. Effects of AAPS on the Lifespan of Drosophila
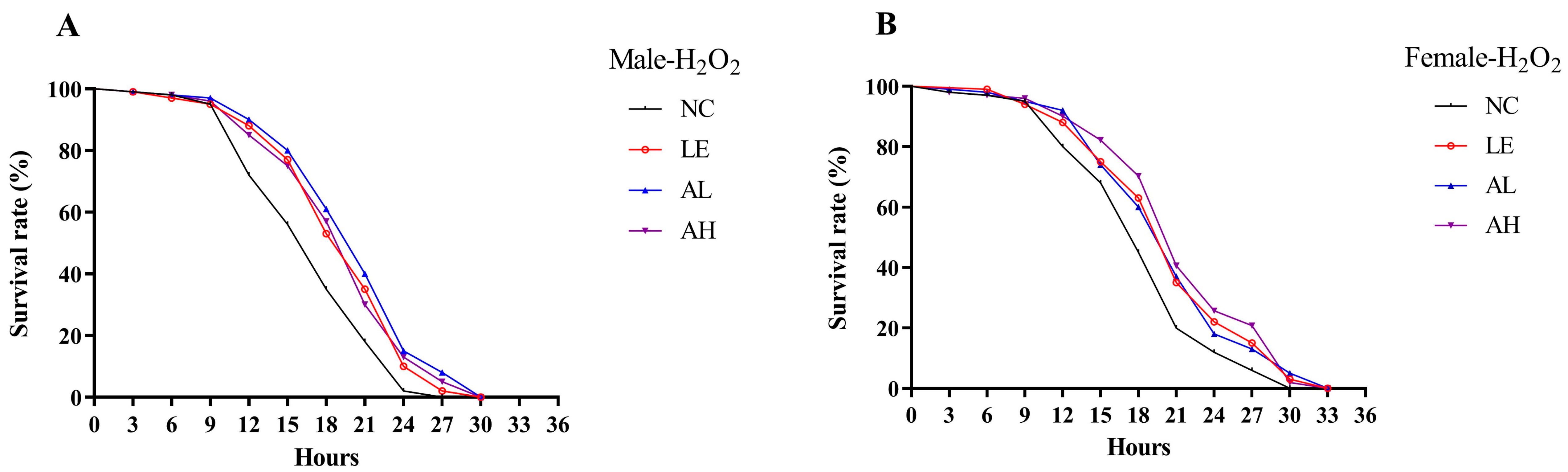
3.2. Effects of AAPS on H2O2-Induced Oxidative Stress in Drosophila
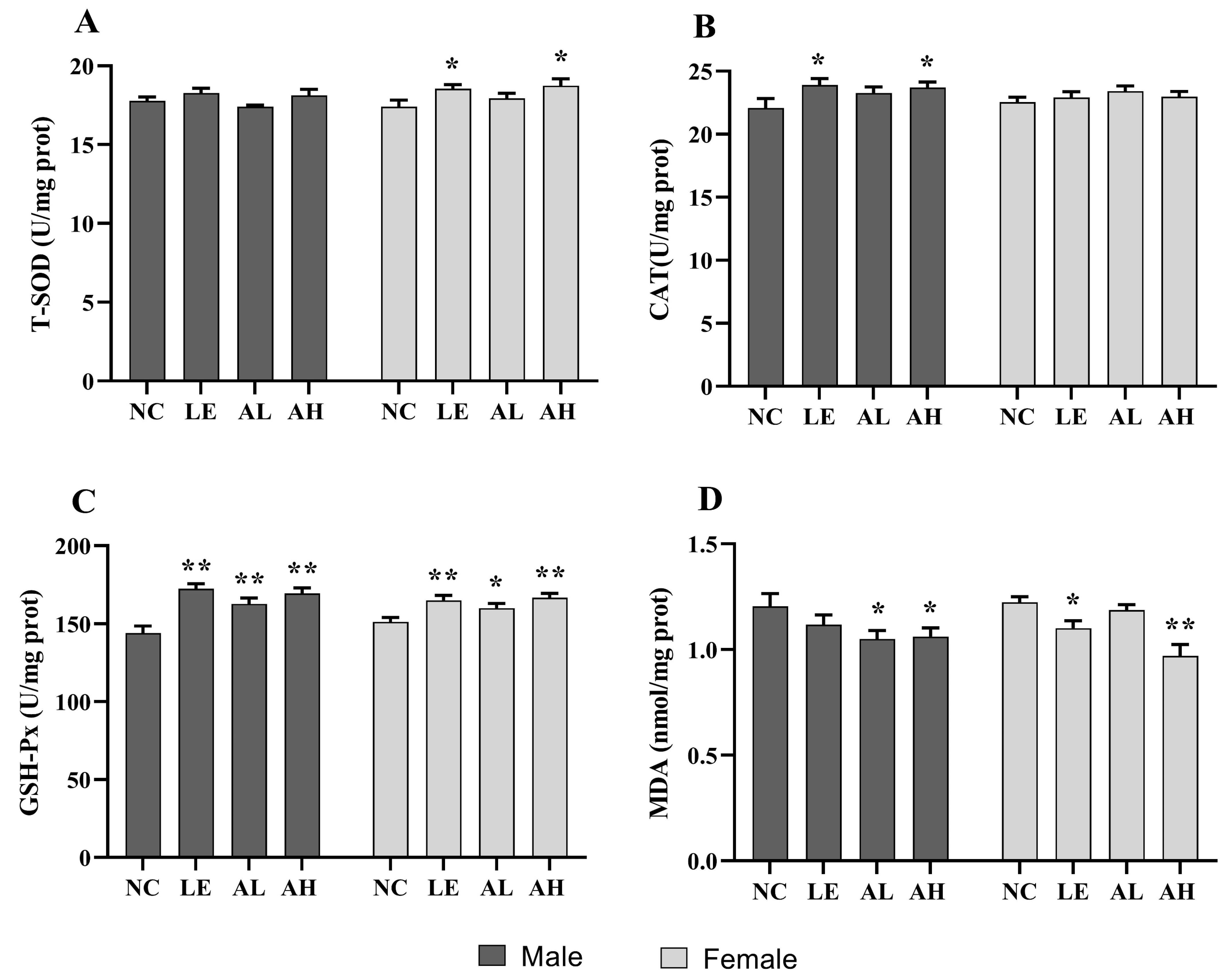
3.3. Effects of AAPS on the Antioxidant Capacity of Drosophila
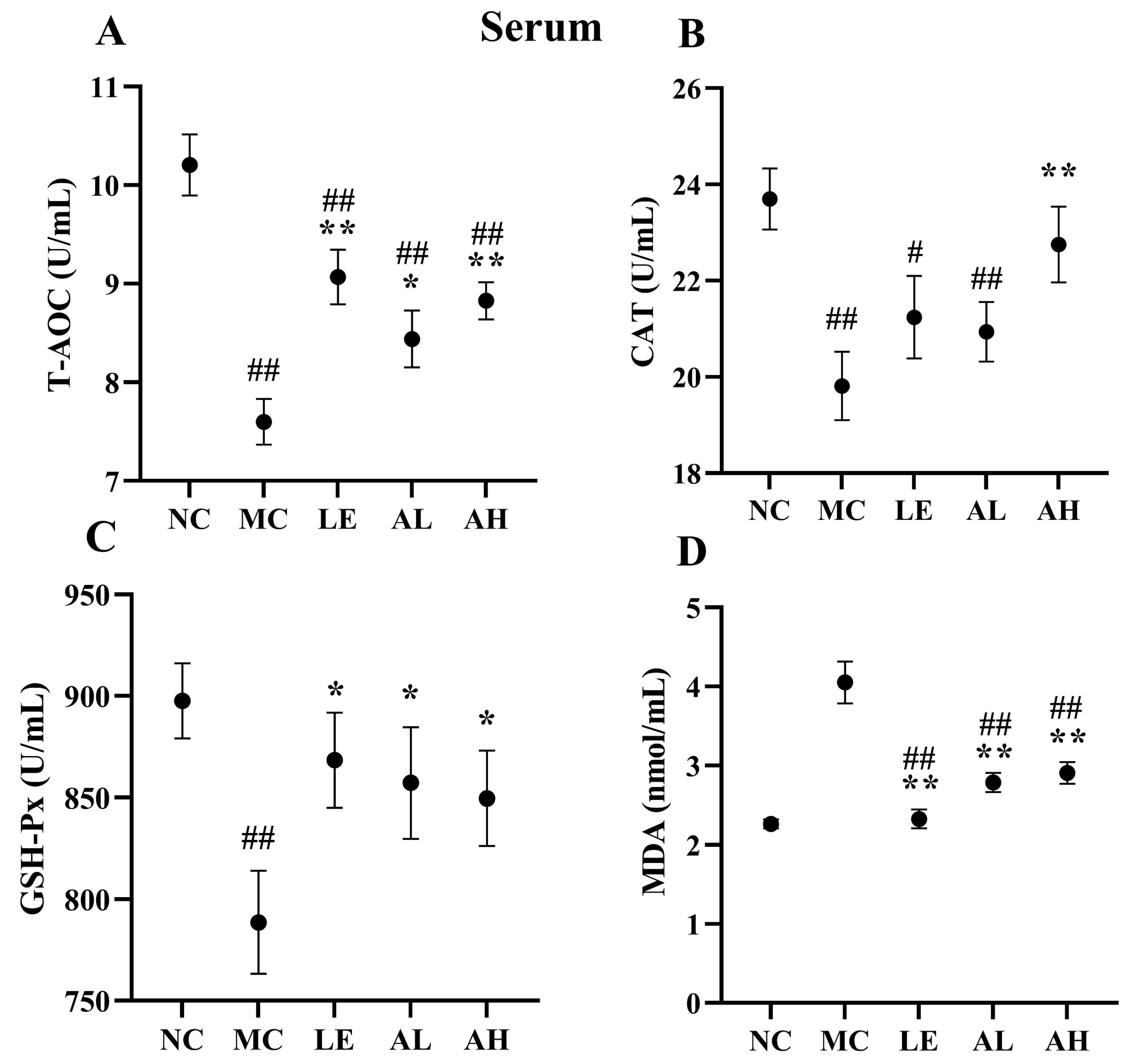
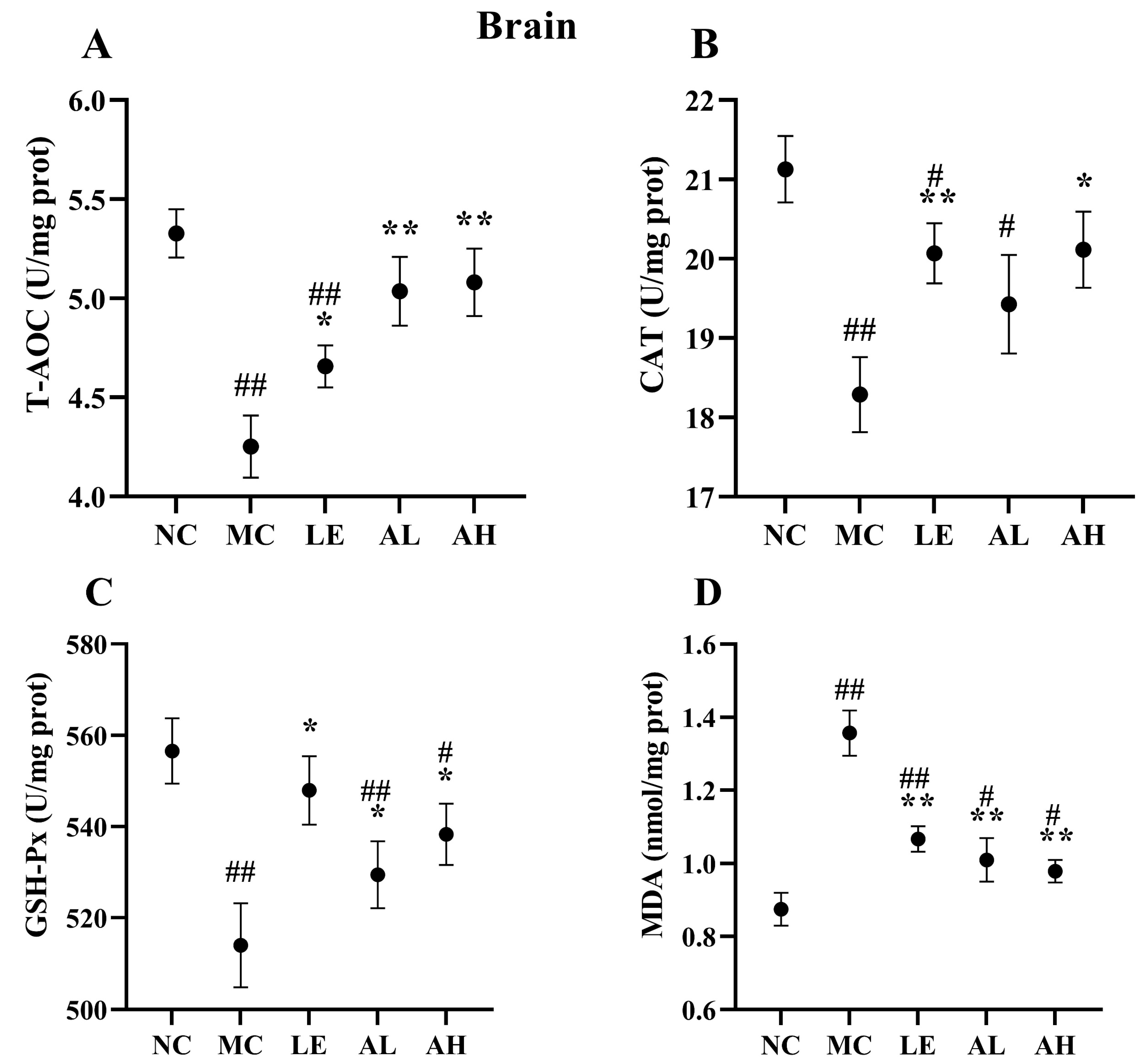
3.4. Effects of AAPS on the Antioxidant Capacity of Mice
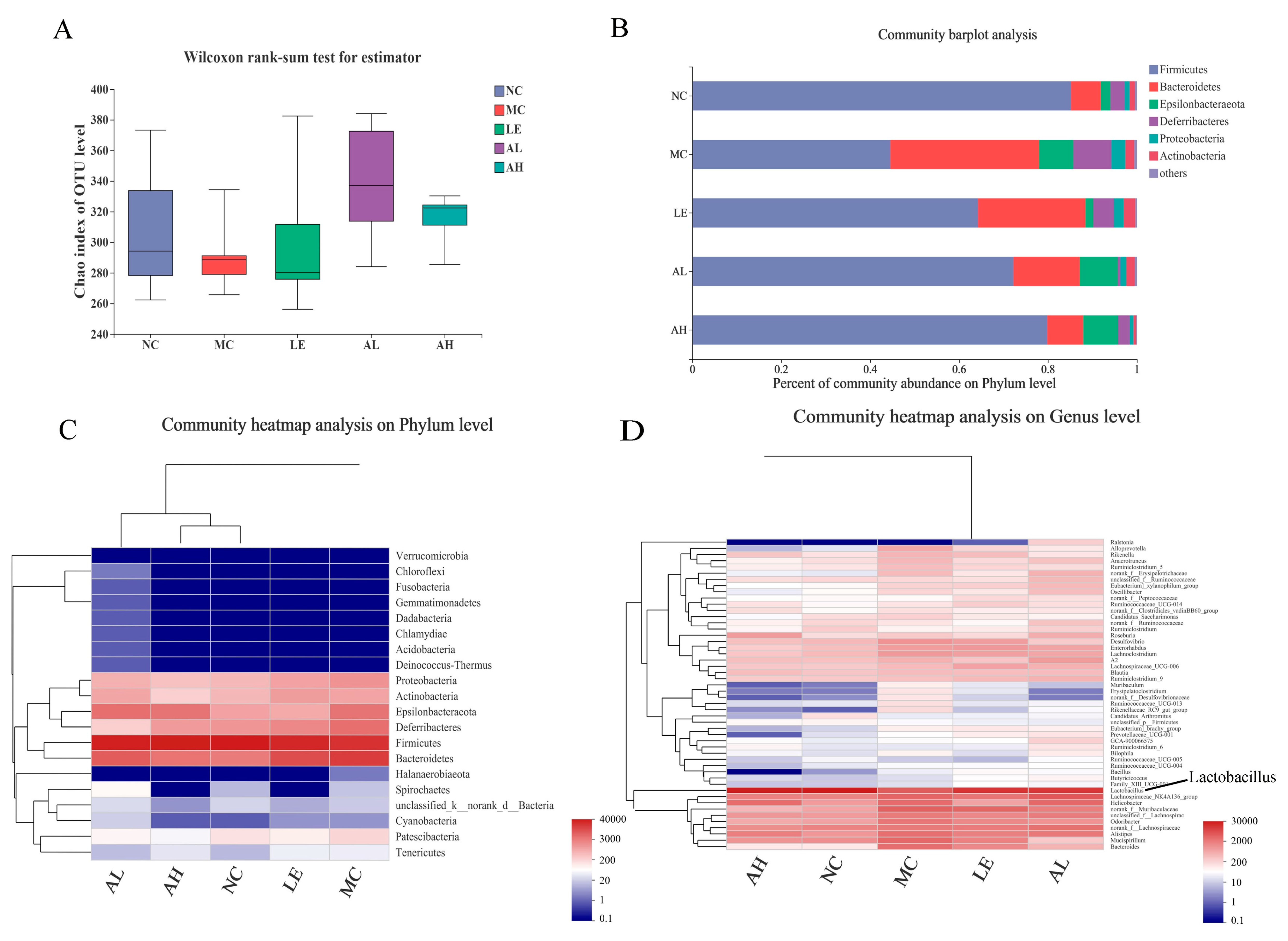
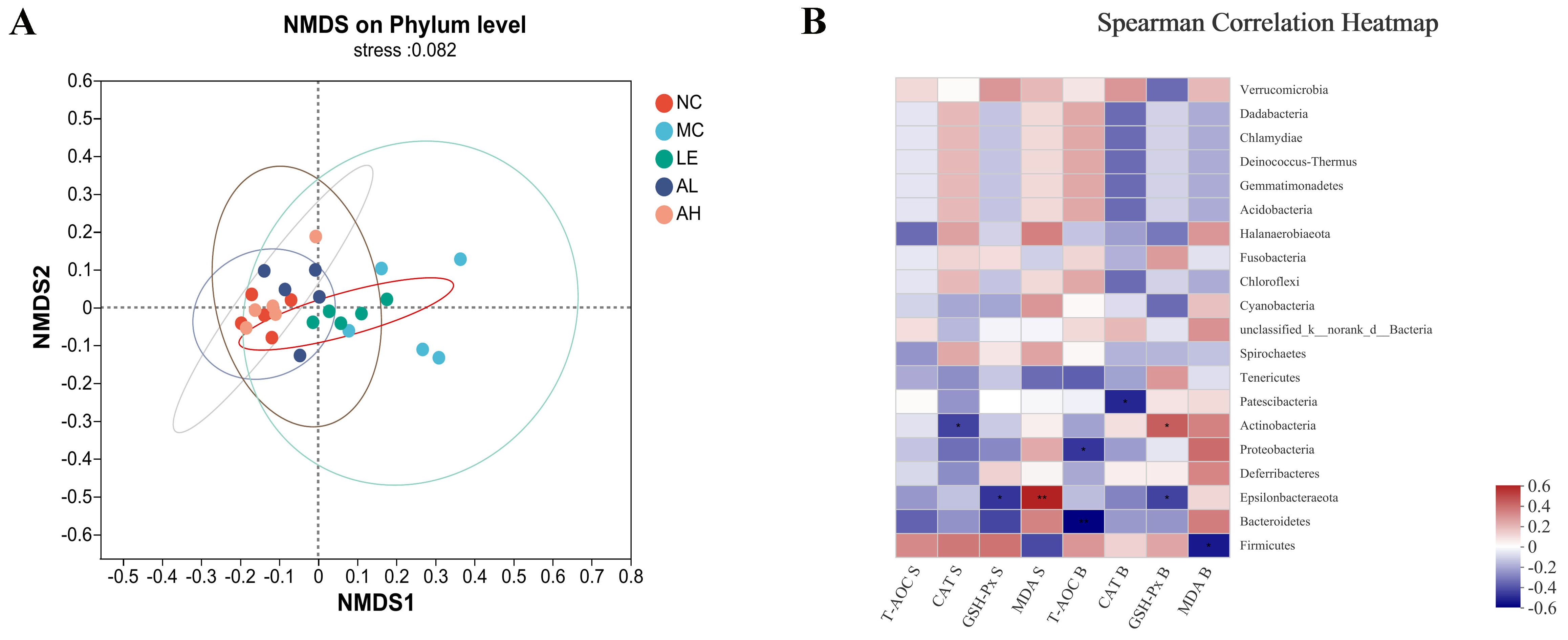
3.5. Effects of AAPS on the Composition of the Gut Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Martemucci, G.; Portincasa, P.; Ciaula, A.D.; Mariano, M.; Centonze, V.; D’Alessandro, A.G. Oxidative stress, aging, antioxidant supplementation and their impact on human health: An overview. Mech. Ageing Dev. 2022, 206, 111707. [Google Scholar] [CrossRef]
- Denham, H. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar]
- Wojtunik-Kulesza, K.A.; Oniszczuk, A.; Oniszczuk, T.; Waksmundzka-Hajnos, M. The influence of common free radicals and antioxidants on development of Alzheimer’s disease. Biomed. Pharmacother. 2016, 78, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J. Redox signaling: An evolution from free radicals to aging. Free Radic. Biol. Med. 2016, 97, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Ionescu-Tucker, A.; Cotman, C.W. Emerging roles of oxidative stress in brain aging and Alzheimer’s disease. Neurobiol. Aging. 2021, 107, 86–95. [Google Scholar] [CrossRef]
- Azman, K.F.; Zakaria, R. D-Galactose-induced accelerated aging model: An overview. Biogerontology 2019, 20, 763–782. [Google Scholar] [CrossRef] [PubMed]
- Shwe, T.; Pratchayasakul, W.; Chattipakorn, N.; Chattipakorn, S.C. Role of D-galactose-induced brain aging and its potential used for therapeutic interventions. Exp. Gerontol. 2018, 101, 13–36. [Google Scholar] [CrossRef]
- Lee, S.H.; Min, K.J. Drosophila melanogaster as a model system in the study of pharmacological interventions in aging. Transl. Med. Aging. 2019, 3, 98–103. [Google Scholar] [CrossRef]
- Taormina, G.; Ferrante, F.; Vieni, S.; Grassi, N.; Russo, A.; Mirisola, M.G. Longevity: Lesson from model organisms. Genes 2019, 10, 518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.J.; Yun, I.J.; Kim, K.H.; Lim, S.H.; Ham, H.J.; Eum, W.S.; Joo, J.H. Amino acid and fatty acid compositions of Agrocybe chaxingu, an edible mushroom. J. Food Compos. Anal. 2011, 24, 175–178. [Google Scholar] [CrossRef]
- Jing, H.; Li, J.; Zhang, J.; Wang, W.; Li, S.; Ren, Z.; Gao, Z.; Song, X.; Wang, X.; Le, J. The antioxidative and anti-aging effects of acidic- and alkalic-extractable mycelium polysaccharides by Agrocybe aegerita (Brig.) Sing. Int. J. Biol. Macromol. 2018, 106, 1270–1278. [Google Scholar] [CrossRef]
- El-Maradny, Y.A.; El-Fakharany, E.M.; Abu-Serie, M.M.; Hashish, M.H.; Selim, H.S. Lectins purified from medicinal and edible mushrooms: Insights into their antiviral activity against pathogenic viruses. Int. J. Biol. Macromol. 2021, 179, 239–258. [Google Scholar] [CrossRef]
- Lin, S.; Ching, L.T.; Lam, K.; Cheung, P.C.K. Anti-angiogenic effect of water extract from the fruiting body of Agrocybe aegerita. LWT Food Sci. Technol. 2017, 75, 155–163. [Google Scholar] [CrossRef]
- Li, G.; Liu, X.; Cong, S.; Deng, Y.; Zheng, X. A novel serine protease with anticoagulant and fibrinolytic activities from the fruiting bodies of mushroom Agrocybe aegerita. Int. J. Biol. Macromol. 2021, 168, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, D.; Chen, Y.; Zhong, R.; Gao, L.; Yang, C.; Ai, C.; El-Seedi, H.R.; Zhao, C. Physicochemical characterization of a polysaccharide from Agrocybe aegirita and its anti-ageing activity. Carbohydr. Polym. 2020, 236, 116056. [Google Scholar] [CrossRef]
- Wu, L.; Liu, X.; Hu, R.; Chen, Y.; Xiao, M.; Liu, B.; Zeng, F. Prebiotic Agrocybe cylindracea crude polysaccharides combined with Lactobacillus rhamnosus GG postpone aging-related oxidative stress in mice. Food Funct. 2022, 13, 1218–1231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, X.; Pan, J.; Zhao, Q.; Li, Y.; Gao, W.; Zhang, Z. Anti-aging effect of brown black wolfberry on Drosophila melanogaster and d-galactose-induced aging mice. J. Funct. Foods 2020, 65, 103724. [Google Scholar] [CrossRef]
- Cai, X.; Chen, S.; Liang, J.; Tang, M.; Wang, S. Protective effects of crimson snapper scales peptides against oxidative stress on Drosophila melanogaster and the action mechanism. Food Chem. Toxicol. 2021, 148, 111965. [Google Scholar] [CrossRef] [PubMed]
- Kemoun, P.H.; Ader, I.; Planat-Benard, V.; Dray, C.; Fazilleau, N.; Monsarrat, P.; Cousin, B.; Paupert, J.; Ousset, M.; Lorsignol, A.; et al. A gerophysiology perspective on healthy ageing. Ageing Res. Rev. 2022, 73, 101537. [Google Scholar] [CrossRef]
- Conway, J.; Duggal, N.A. Ageing of the gut microbiome: Potential influences on immune senescence and inflammageing. Ageing Res. Rev. 2021, 68, 101323. [Google Scholar] [CrossRef] [PubMed]
- Barone, M.; D’Amico, F.; Rampelli, S.; Brigidi, P.; Turroni, S. Age-related diseases, therapies and gut microbiome: A new frontier for healthy aging. Mech. Ageing Dev. 2022, 206, 111711. [Google Scholar] [CrossRef] [PubMed]
- Travier, L.; Singh, R.; Fernández, D.S.; Deczkowska, A. Microbial and immune factors regulate brain maintenance and aging. Curr. Opin. Neurobiol. 2022, 76, 102607. [Google Scholar] [CrossRef] [PubMed]
- Warraich, U.A.; Hussain, F.; Kayani, H.R. Aging—Oxidative stress, antioxidants and computational modeling. Heliyon 2020, 6, e04107. [Google Scholar] [CrossRef] [PubMed]
- Demirci-Çekiç, S.; Özkan, G.; Avan, A.N.; Uzunboy, S.; Çapanoğlu, E.; Apak, R. Biomarkers of oxidative stress and antioxidant defense. J. Pharm. Biomed. 2022, 209, 114477. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative stress mitigation by antioxidants—An overview on their chemistry and influences on health status. Eur. J. Med. Chem. 2021, 209, 112891. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.J.; Jana, C.K.; Das, N. Role of catalase in oxidative stress-and age-associated degenerative diseases. Oxid. Med. Cell Longev. 2019, 2019, 9613090. [Google Scholar] [CrossRef] [Green Version]
- Mas-Bargues, C.; Escrivá, C.; Dromant, M.; Borrás, C.; Viña, J. Lipid peroxidation as measured by chromatographic determination of malondialdehyde. Human plasma reference values in health and disease. Free Radic. Biol. Med. 2021, 709, 108941. [Google Scholar] [CrossRef] [PubMed]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Vaiserman, A.M.; Koliada, A.K.; Marotta, F. Gut microbiota: A player in aging and a target for anti-aging intervention. Ageing Res. Rev. 2017, 35, 36–45. [Google Scholar] [CrossRef]
- Coman, V.; Vodnar, D.C. Gut microbiota and old age: Modulating factors and interventions for healthy longevity. Exp. Gerontol. 2020, 141, 111095. [Google Scholar] [CrossRef]
- Tiihonen, K.; Ouwehand, A.C.; Rautonen, N. Human intestinal microbiota and healthy ageing. Ageing Res. Rev. 2010, 9, 107–116. [Google Scholar] [CrossRef]
- Biagi, E.; Franceschi, C.; Rampelli, S.; Severgnini, M.; Ostan, R.; Turroni, S.; Consolandi, C.; Quercia, S.; Scurti, M.; Monti, D.; et al. Gut microbiota and extreme longevity. Curr. Biol. 2016, 26, 1480–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, F.; Hua, Y.; Zeng, B.; Ning, R.; Li, Y.; Zhao, J. Gut microbiota signatures of longevity. Curr. Biol. 2016, 26, 832–833. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Babaei, P.; Ji, B.; Nielsen, J. Human gut microbiota and healthy aging: Recent developments and future prospective. Nutr. Health Aging 2016, 4, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagi, H.; Shibuya, S.; Kato, T.; Nakanishi, Y.; Tsuboi, A.; Moriya, S.; Ohno, H.; Miyamoto, H.; Kodama, H.; Shimizu, T. SOD1 deficiency alters gastrointestinal microbiota and metabolites in mice. Exp. Gerontol. 2020, 130, 110795. [Google Scholar] [CrossRef]
- Li, Q.; Gänzle, M.G. Host-adapted lactobacilli in food fermentations: Impact of metabolic traits of host adapted lactobacilli on food quality and human health. Curr. Opin. Food Sci. 2020, 31, 71–80. [Google Scholar] [CrossRef]
- Remus, D.M.; Kleerebezem, M.; Bron, P.A. An intimate tête-à-tête—How probiotic lactobacilli communicate with the host. Eur. J. Pharmacol. 2011, 668, S33–S42. [Google Scholar] [CrossRef]
- Li, B.; Evivie, S.E.; Lu, J.; Jiao, Y.; Wang, C.; Li, Z.; Liu, F.; Huo, G. Lactobacillus helveticus KLDS1.8701 alleviates d-galactose-induced aging by regulating Nrf-2 and gut microbiota in mice. Food Funct. 2018, 9, 6586–6598. [Google Scholar] [CrossRef]
- Kong, Y.; Olejar, K.J.; On, S.L.W.; Chelikani, V. The potential of lactobacillus spp. for modulating oxidative stress in the gastrointestinal tract. Antioxidants 2020, 9, 610. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Mean Lifespan (d) | Maximum Lifespan (d) | Median Survival (d) | Prolongation of Mean Life Span (%) | |
|---|---|---|---|---|---|
| Male | NC | 42.74 ± 1.66 | 61.70 ± 0.38 | 50.5 | - |
| LE | 47.00 ± 1.60 * | 65.40 ± 0.32 ** | 53.5 | 9.97 | |
| AL | 45.57 ± 1.56 | 62.80 ± 0.42 * | 52.5 | 6.62 | |
| AH | 46.85 ± 1.53 * | 64.60 ± 0.47 ** | 54 | 9.62 | |
| Female | NC | 45.14 ± 1.61 | 63.20 ± 0.24 | 51.5 | - |
| LE | 47.91 ± 1.67 | 67.10 ± 0.33 ** | 55 | 6.14 | |
| AL | 49.12 ± 1.64 * | 69.70 ± 0.66 ** | 56 | 8.82 | |
| AH | 48.79 ± 1.61 | 69.00 ± 0.60 ** | 54 | 8.09 |
| Group | Mean Lifespan (h) | Maximum Lifespan (h) | Median Survival (h) | Prolongation of Mean Life Span (%) | |
|---|---|---|---|---|---|
| Male | NC | 17.28 ± 0.48 | 24.60 ± 0.38 | 18 | - |
| LE | 19.68 ± 0.52 ** | 27.60 ± 0.38 ** | 21 | 13.89 | |
| AL | 20.61 ± 0.53 ** | 29.10 ± 0.43 ** | 21 | 19.27 | |
| AH | 19.77 ± 0.53 ** | 28.50 ± 0.47 ** | 21 | 14.41 | |
| Female | NC | 18.63 ± 0.57 | 28.80 ± 0.46 | 18 | - |
| LE | 20.82 ± 0.61 ** | 30.90 ± 0.43 ** | 21 | 11.76 | |
| AL | 20.73 ± 0.61 ** | 31.50 ± 0.47 ** | 21 | 11.27 | |
| AH | 21.60 ± 0.63 ** | 30.60 ± 0.38 ** | 21 | 15.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Wu, L.; Tong, A.; Zhen, H.; Han, D.; Yuan, H.; Li, F.; Wang, C.; Fan, G. Anti-Aging Effect of Agrocybe aegerita Polysaccharide through Regulation of Oxidative Stress and Gut Microbiota. Foods 2022, 11, 3783. https://doi.org/10.3390/foods11233783
Liu X, Wu L, Tong A, Zhen H, Han D, Yuan H, Li F, Wang C, Fan G. Anti-Aging Effect of Agrocybe aegerita Polysaccharide through Regulation of Oxidative Stress and Gut Microbiota. Foods. 2022; 11(23):3783. https://doi.org/10.3390/foods11233783
Chicago/Turabian StyleLiu, Xiaoyan, Linxiu Wu, Aijun Tong, Hongmin Zhen, Dong Han, Hongyang Yuan, Fannian Li, Chengtao Wang, and Guangsen Fan. 2022. "Anti-Aging Effect of Agrocybe aegerita Polysaccharide through Regulation of Oxidative Stress and Gut Microbiota" Foods 11, no. 23: 3783. https://doi.org/10.3390/foods11233783