3.1. Total Coliform Bacteria and Enterococci in Flour, Powder and Dough
Twenty-nine flour samples and 31 samples of plant or insect powders and 13 samples of dough were subjected to microbiological analysis. In 87% of samples of flour and powder were determined the total number of aerobic bacteria (
Table 5). Fecal indicators (coliform bacteria or enterococci) were detected in 53% of samples of flour and powder. A total of 85% of samples of dough were determined total aerobic (
Table 6). Coliform bacteria and enterococci were presented in 38% of all samples of dough, where total enterococci were observed only in one sample, namely sample of strudel dough in number of 4.1 log CFU/g.
The majority of detected numbers of total coliform bacteria (TCB) has ranged between 2.3 and 4.1 log CFU/g. These counts are comparable to Batool et al. (2012), who determined coliform bacteria counts in Pakistani flour in the range of 3.0–4.0 log CFU/g [
33]. Twelve powder samples contained coliforms ranging from 2.3 to 5.3 log CFU/g. We recorded the most numerous growths in the case of chestnut powder (M3) originating from Italy. The microbiological quality of flour and other mill products is defined in the second part of the Codex Alimentarius of the Slovak Republic, which regulates the microbial requirements for food and packaging. This section sets out the requirements for coliform bacteria, fungi, and
Bacillus cereus. As for coliform bacteria, these can be found in flours in the number of 103, the limit value being 104 in two samples out of five examined samples [
34]. Microbiological criteria for powders are not currently available. Four samples of pizza and puff dough contained coliform bacteria (
Table 6). In March 2022, the consumption of frozen pizza was connected to the
E. coli outbreak in France [
35].
Escherichia coli was detected only in one sample of pumpkin powder.
Compared to coliform bacteria, the number of samples with enterococci was much lower. We detected enterococci only in six samples of powder and one sample of strudel pastry. In cricket powder, we observed the highest count of enterococci. The genus
Enterococcus spp. is part of the healthy intestinal microbiota of humans and animals, and some strains are used as starter cultures or probiotics. Due to their preferred intestinal habitat, their wide occurrence and ease of cultivation serve as indicators of fecal contamination and are part of hygienic standards for water and food products [
36,
37].
3.2. Antibiotic Resistant Coliform Bacteria and Enterococci in Flour, Powder and Dough
Besides TCB and TEC (total enterococci), we also detected their possible antibiotic resistance. When quantifying resistant coliform bacteria and enterococci, we applied the concentrations of individual antibiotics in accordance with EUCAST guidelines. In case of coliform bacteria, we applied the antibiotics ampicillin (penicillin), gentamicin (aminoglycoside), chloramphenicol (amphenicol), ciprofloxacin (fluoroquinolone), and tetracycline (tetracycline), which represent different classes of antibiotics. For tetracycline, we used the concentration specified by CLSI guidelines, while there are no breakpoints given by EUCAST. The antibiotics ampicillin (penicillin), gentamicin (aminoglycoside), ciprofloxacin (fluoroquinolone), and vancomycin (glycopeptide) were applied in the determination of resistant enterococci.
In 45% of flour samples, we detected antibiotic-resistant coliforms (
Table 7). The majority was observed only in the case of ampicillin. Only in one sample of rye flour did we also observe gentamicin-resistant coliforms. In 39% of powder samples, we observed ampicillin resistance. We recorded the highest number in a sample of chestnut powder (M3). Significantly higher resistance to ampicillin was expected because most coliforms have intrinsic resistance to this antibiotic, except for the genus
Escherichia. In five powder samples, we detected gentamicin resistance in the range of 2.7–4.3 log CFU/g. We recorded chloramphenicol and tetracycline resistance only in chestnut powder (M3). We did not detect resistance to ciprofloxacin in any sample. We have observed ampicillin-resistant enterococci only in six powder samples. In samples of chestnut (M3) and coconut powder (M19, M30), we also detected vancomycin-resistant enterococci. The only sample of insect flour-cricket flour (M49) contained ampicillin (penicillins) and gentamicin (aminoglycosides)-resistant coliform bacteria. Ampicillin-resistant enterococci were also present in this flour sample. In dough samples, we did not register any antibiotic-resistant bacteria.
The presence of total and antibiotic-resistant coliforms and enterococci in powders from chickpeas, rice, banana, almond, and coconut is of considerable concern, while these powders can be used in cold cuisine as an addition to drinks, spreads, salads, yogurt, smoothies, etc. [
18].
In the United States in 2018, voluntary recalls of approximately 6300 packages of organic coconut powder were reported, due to the possible presence of
Salmonella, which was detected in one package during routine testing. In the same year,
Salmonella was the cause of the further withdrawal of flour from the market, namely 2099 packages of organic amaranth powder. Fortunately, none of these withdrawals have reported cases of disease due to flour consumption [
38,
39].
3.3. Identification of Antibiotic-Resistant Strains
Isolated colonies of antibiotic-resistant coliforms and enterococci were identified using a MALDI-TOF mass spectrometer, which can be used to analyze biological macromolecules by determining the weight of molecules after mixing with a matrix and laser beam ionization. Identification of microorganisms is performed by comparing the obtained mass spectrum with the spectra of reference strains in the relevant database. This method provides fast, well reproducible and accurate identification of microorganisms at the genus and species level [
40].
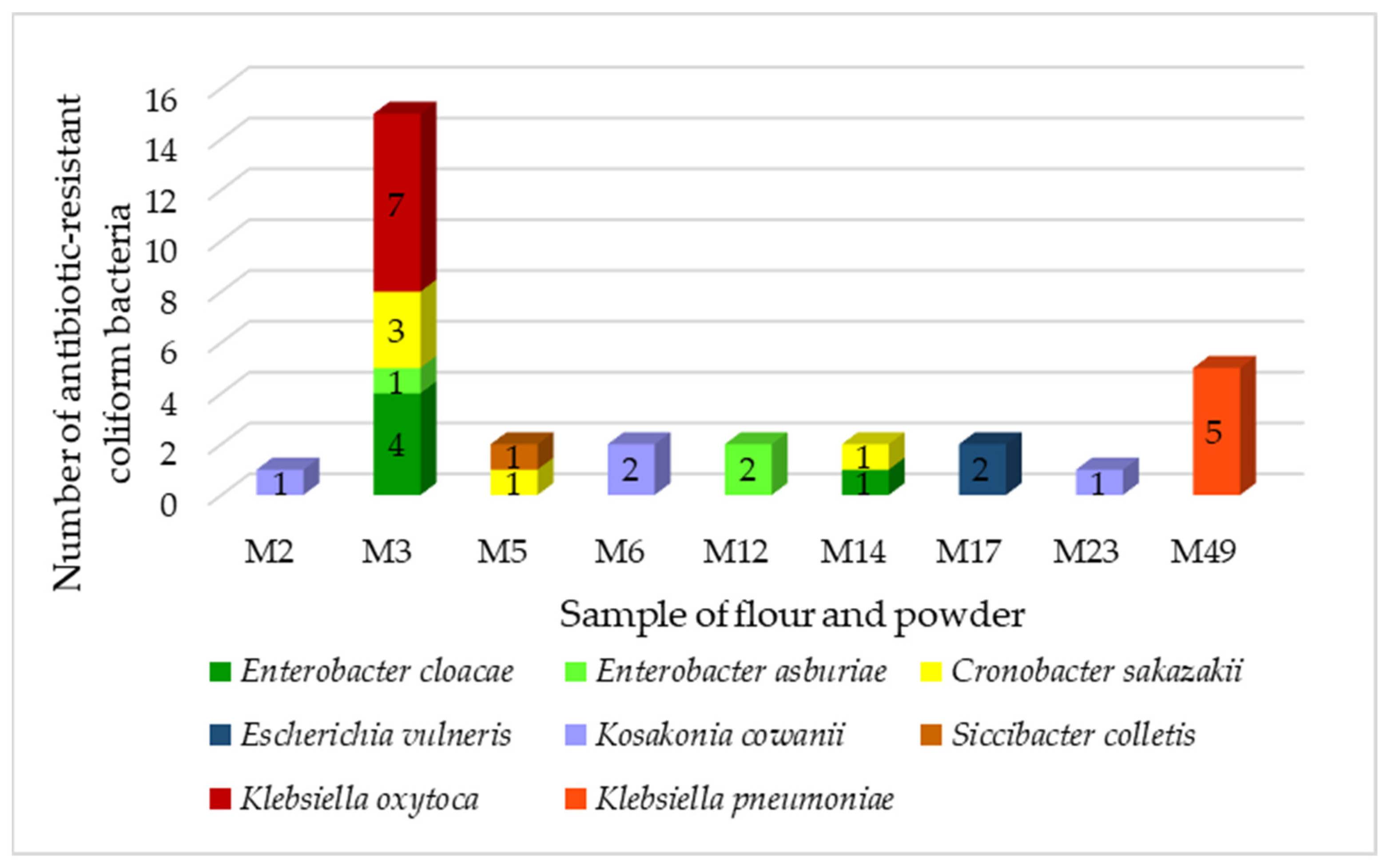
In total, we identified 32 resistant coliform isolates belonging to six genera (
Figure 1).
The majority was from the
Klebsiella family (37%). Bacteria of the genus
Klebsiella are widespread in nature. They are associated with several pathological processes in humans and animals but can also enter the food chain.
Klebsiella is not considered to be a major intestinal or foodborne pathogen, but its participation in intestinal diseases cannot be ruled out. Multidrug-resistant
Klebsiella spp. producing broad-spectrum β-lactamases that can be transported through food [
41]. We identified two species of the genus
Klebsiella,
K. oxytoca and
K. pneumoniae. These two species are the most common pathogens causing nosocomial infections in humans and are of great concern regarding the development of resistance to multiple antibiotics [
42].
Klebsiella pneumoniae is a common cause of community-acquired and nosocomial infections, such as urinary tract infections, lower respiratory tract infections, and liver abscesses. Although not traditionally considered a foodborne pathogen, there have been reports of
K. pneumoniae infections preceded by intestinal colonization, supporting the theory of food as a possible vector for the transmission of these pathogens [
43]. All strains of
K. pneumoniae were isolated from cricket powder.
Strains of
Cronobacter sakazakii were isolated from chestnut and amaranth powder.
Cronobacter sakazakii is an opportunistic pathogen associated with life-threatening neonatal infections that can lead to severe manifestations such as brain abscesses, meningitis, necrotizing enterocolitis, and systemic sepsis. Most cases are associated with the consumption of powdered infant formula contaminated with these microorganisms [
44].
Kosakonia cowanii was isolated from teff and rye flour as well as from rice powder.
Kosakonua cowanii is a recently reclassified bacterial species formerly known as
Enterobacter cowanii. The bacterium is thought to be primarily a plant pathogen, but cases of human infections have been reported in which
K. cowanii has been identified as the cause of rhabdomyolysis and bacteremia associated with rose thorns and as the cause of acute cholecystitis [
45,
46]. Up to 63% of
Enterobacter were isolated from chestnut powder. These bacteria are ubiquitous and have been found in a wide variety of foods, including fruits and vegetables, meat, fish, eggs, tea, herbs, legumes, spices, dry animal feed, dairy products, powdered baby food, cereals, nuts, seeds, flour, pasta, chocolate, drinks, and water.
Enterobacter cloacae is the most isolated species of the genus
Enterobacter from humans and animals. It is referred to as an opportunistic pathogen identified in nosocomial infections [
47].
Escherichia vulneris and
Siccibacter colletis were isolated from chickpea powder.
Enterobacter vulneris is an opportunistic human pathogen that has been identified in clinical cases of urosepsis, osteomyelitis, intravenous catheter-related bacteremia, meningitis, and dialysis-related peritonitis [
48].
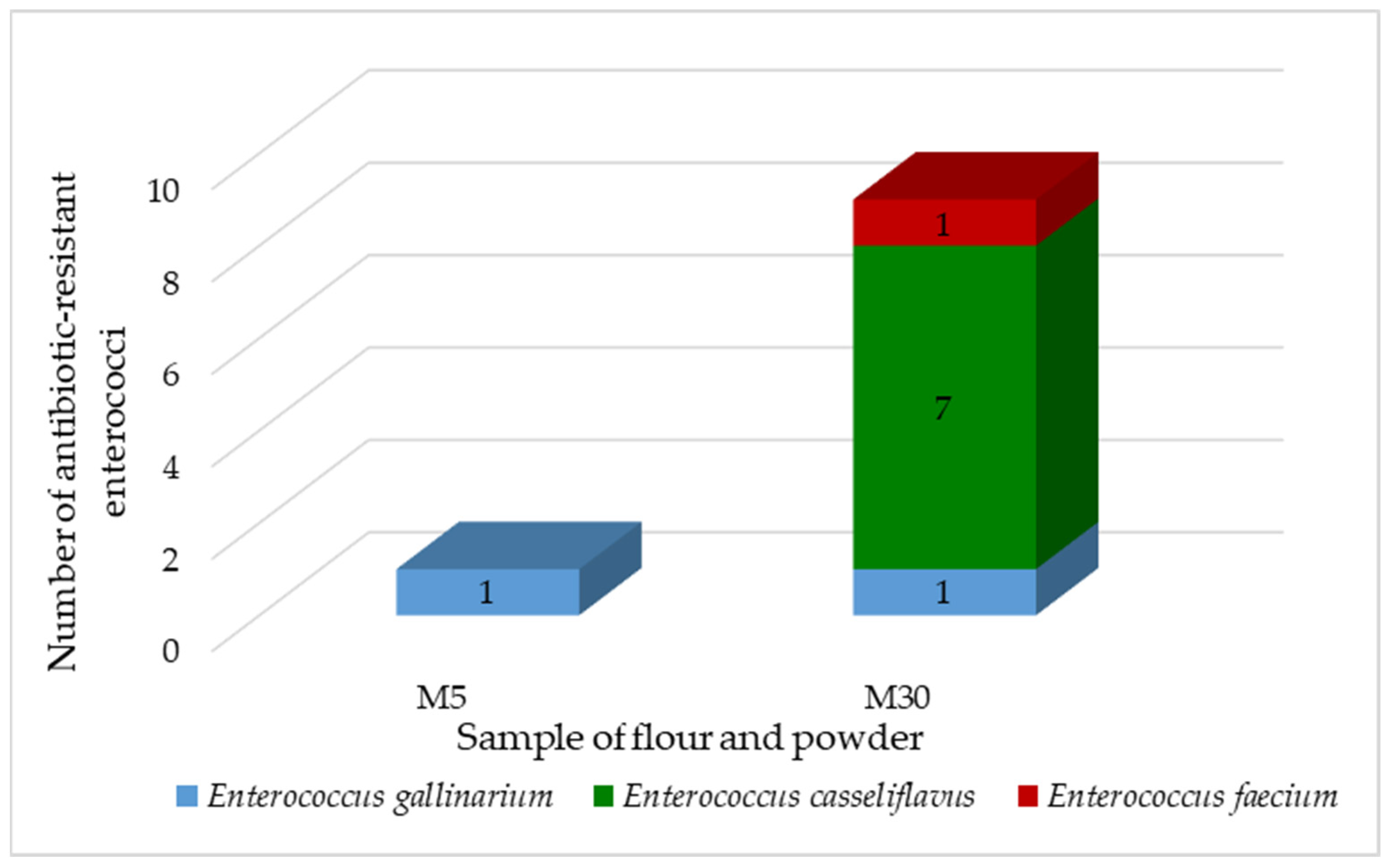
We have isolated and identified 10 antibiotic-resistant enterococci. The majority belonged to
E. casseliflavus (70%), followed by
E.gallinarium (20%) and
E. faecium (10%) (
Figure 2).
Enterococcus faecium is the second most common species of
Enterococcus, which is the cause of nosocomial diseases. It often possesses vancomycin resistance genes, making it a priority pathogen for the development of new antibiotics [
49]. Most enterococci were isolated from coconut powder, only one isolate was derived from amaranth powder.
3.4. Characterization of Antibiotic-Resistant Strains
In almost all isolates we have detected ampicillin resistance (
Table 8). Only one isolate of
K. cowani from rye flour showed susceptibility to this antibiotic. The majority of coliform bacteria are intrinsically resistant to ampicillin, in the case of the genus
Escherichia it is an acquired mechanism of resistance [
24]. We did not observe resistance to gentamicin, ciprofloxacin and meropenem in any of the isolates. In seven isolates, we detected chloramphenicol resistance. In five
K. oxytoca isolates from chestnut powder, we observed tetracycline resistance.
Enterococci have intrinsic and acquired resistance to most antibiotics used in humans, leading to treatment complications. Enterococci are naturally resistant to β-lactams, carbapenems and cephalosporins. They may also acquire resistance to tetracyclines, quinolones, macrolides, erythromycin, streptogramin, polymixins, clindamycin, and glycopeptides (vancomycin) [
50]. Of the ten isolates tested, only three showed resistance to ampicillin at CLSI concentrations (
Table 9). Ampicillin and penicillin, belonging to β-lactam ATBs, inhibit the synthesis of peptidoglycan, a critical component required for bacterial viability [
50]. Higher levels of ampicillin resistance in enterococci are associated with the presence of the species-specific chromosomal gene pbp5, which encodes class B penicillin-binding proteins (PBPs) with binding affinity for ampicillin and cephalosporins [
51]. All isolates were vancomycin-resistant. Vancomycin-resistant enterococci found in food can temporarily colonize the human gut and confer multidrug resistance to human
E. faecalis via a conjugation plasmid [
52].
Multidrug resistance, defined as resistance to three or more antimicrobials, was not detected in any of the isolates. The high level of antibiotic resistance is due to the widespread use of antibiotics in medical practice, especially in developing countries [
42].
Efflux causes a decrease in intracellular drug concentrations, leading to subtoxic levels, which may ultimately induce phenotypic resistance [
53]. All isolates of coliform bacteria and enterococci showed normal efflux. In contrast to our results, Krahulcova et al. recorded in their work the overproduction of efflux pumps in up to 19% of coliforms obtained from smoothie drinks [
25].
About 1300 types of β-lactamases are currently known, of which 200 are types of broad-spectrum β-lactamases (ESBLs) [
54]. ESBLs are commonly present in
Klebsiella and
E. coli species but have also been detected in other members of the
Enterobacterales family, such as
Salmonella,
Enterobacter,
Citrobacter,
Serratia and
Proteus. These microorganisms are the cause of septicemia, pneumonia, intra-abdominal abscess as well as various infections, e.g., urinary tract infections [
55]. ESBL-related infections are on the rise and have been observed worldwide in hospitals and communities [
56].
Coliform isolates, in which we confirmed ampicillin resistance were subjected to the detection of the
blaTEM,
blaSHV and
blaOXA genes. We did not detect the presence of the
blaTEM and
blaOXA genes in any of the isolates. We confirmed the presence of the
blaSHV-1 gene in 13% of ampicillin-resistant coliform bacteria, of which all isolates belonged to the
K. oxytoca species (
Table 10).
Klebsiella species exhibit resistance to a broad spectrum of antibiotics, with resistance to penicillins, particularly ampicillin and carbenicillin, being mediated by the enzyme SHV-1 [
57]. In the remaining 87% of isolates, ampicillin resistance may be due to other mechanisms or genes.
We also determined the presence of
tetA and
tetE genes in coliform isolates (
Table 10). These genes mediate tetracycline resistance through membrane-associated proteins that export the drug from the cell and thereby reduce its intracellular concentration [
58]. Although we did not detect excessive production of efflux pumps in any of the isolates, we recorded the presence of the
tetA and
tetE genes in eight isolates. It follows that the isolates have genes encoding specific efflux pumps, but they are not expressed. In addition to overproduction of efflux pumps, resistance to tetracycline antibiotics is mediated by two other mechanisms, namely the protection of ribosomes by proteins that prevent the efficient binding of tetracycline or the production of enzymes that inhibit the ATB. Each of these mechanisms is encoded by a wide variety of genes [
58].
In the case of enterococci, we focused on the detection of the
vanA gene, which is the most common gene encoding glycopeptide resistance [
59]. Worldwide, the
vanA gene is frequently detected in
E. faecium and
E. faecalis strains isolated from humans and animals but is less common in other species, such as
E. gallinarum,
E. hirae, and
E. durans [
60]. Although all isolates showed vancomycin resistance, we did not detect the
vanA gene in any of the isolates. However, vancomycin resistance is mediated by several genes referred to as
vanA -B, -C, -D, -E, -G. Of these, the
vanA and
vanB genes located on plasmids are most frequently detected, followed by chromosomally localized
vanD and
vanC genes. These genes encode changes in the amino acid sequence of the peptidoglycan precursor protein, thereby preventing vancomycin binding [
61]. The relationship of food enterococci with clinical infections has not yet been clearly elucidated [
62]. Foodborne enterococci are not known to be a direct source of resistant enterococci in humans but could pose a risk of transmitting resistance determinants to strains present in the human intestinal microbiota [
60].
Biofilms can be found in food, medical and natural environments. Pathogenic bacteria that enter food production areas can remain there in the form of a biofilm covering the surfaces of machinery and equipment. Biofilms formed on these surfaces are a major cause of contamination of the final product, which ultimately leads to disease transmission, reduced food shelf life and quality, and thus huge economic losses [
63]. Studies suggest that the biofilm is important for the transfer of conjugation plasmids due to the high proximity of cells in this structure. The resistance of a biofilm to ATB depends on various factors, such as physical, physiological, and genetic factors [
63]. In a mature biofilm, bacterial cells can tolerate antibiotics at concentrations 10–1000-fold higher than those required to kill planktonic cells [
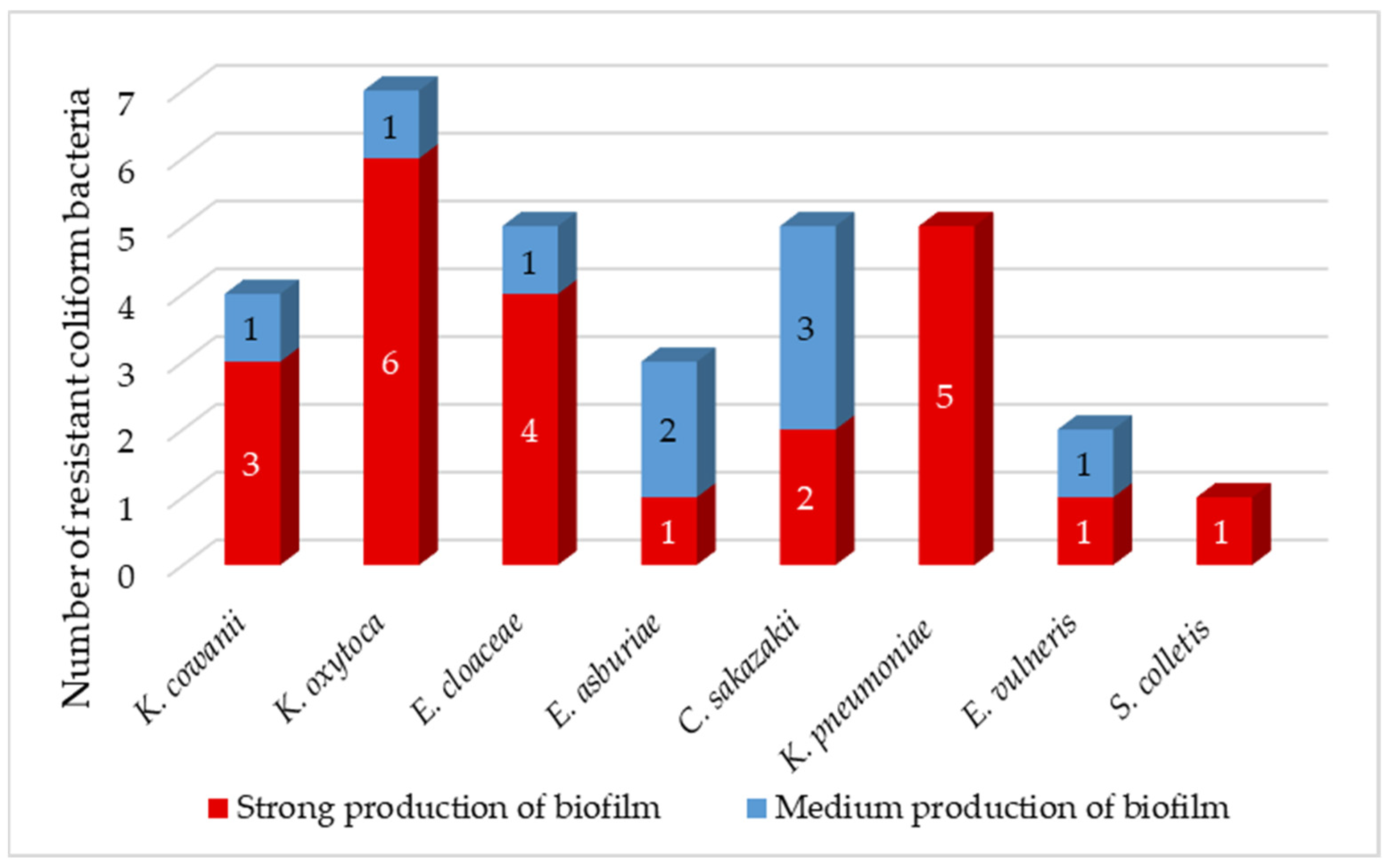
64]. The majority of isolated coliforms (72%) were strong producers of biofilm (
Figure 3), which correlates with the results of Krahulcová, who evaluated 73% of coliform isolates from raw milk and smoothie as strong biofilm producers [
25]. The remaining 28% of isolates were medium-strength biofilm producers. We identified 86% of
K. oxytoca as a strong biofilm producer. All
K. pneumoniae isolates were classified as strong biofilm producers. Biofilm-forming
K. pneumoniae is a major source of nosocomial infections associated with high morbidity and mortality due to limited treatment options [
63]. Two isolates of
C. sakazakii showed strong production, while the remaining three isolates were intermediate biofilm producers. The survival and persistence of
C. sakazakii in powdered infant formula require the body’s ability to adapt to harsh osmotic and dry conditions. Studies have shown that
C. sakazakii cells in biofilms were more persistent compared to plankton cells when exposed to low humidity conditions [
65].
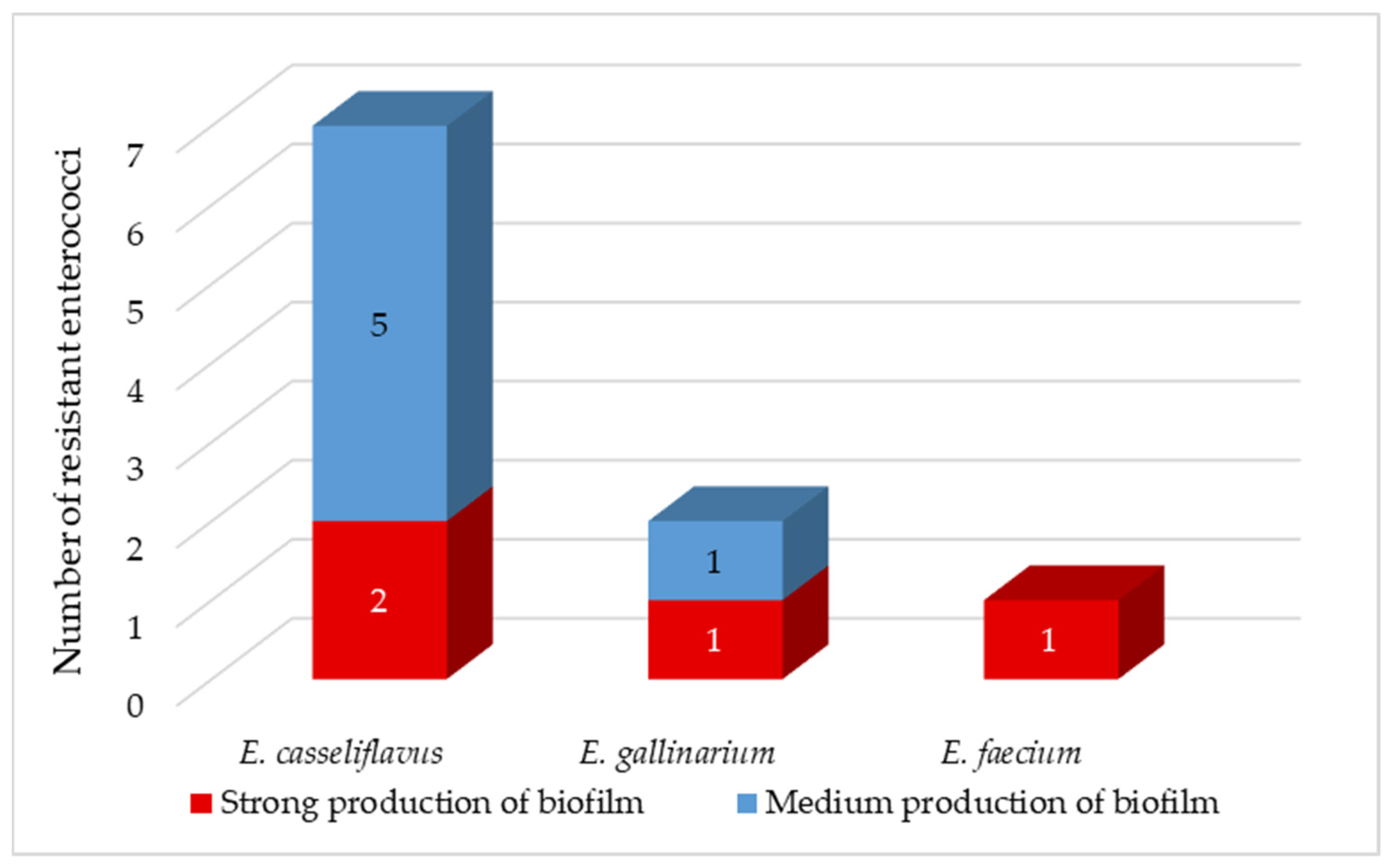
Enterococci are known for their ability to form biofilms [
66]. Enterococcal biofilms have been reported in many infections, including urinary tract, wounds, gastrointestinal tract, and endocarditis. Biofilm-associated enterococcal infections are not only difficult to eradicate but also serve as a reservoir for antibiotic resistance genes. Low penetration of antibiotics through the biofilm matrix and the presence of persistent cells contribute to antibiotic tolerance of biofilms, leading to persistent infections [
49]. Approximately 80% of persistent bacterial infections in the United States have been found to be associated with biofilms [
63]. We observed medium biofilm production in isolates of
E. casseliflavus and
E. gallinarium (
Figure 4). In the remaining 40% of isolates, including
E. faecium isolate, we have detected strong production of biofilm. We did not observe very strong biofilm production in any of the coliform bacteria nor enterococci isolates (
Figure 4).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}