
The Wheat Aleurone Layer: Optimisation of Its Benefits and Application to Bakery Products

Abstract
:1. Introduction
2. Aleurone Layer
2.1. Description: Histology and Functions
2.2. Composition
2.2.1. Cell Wall

{kind=link}
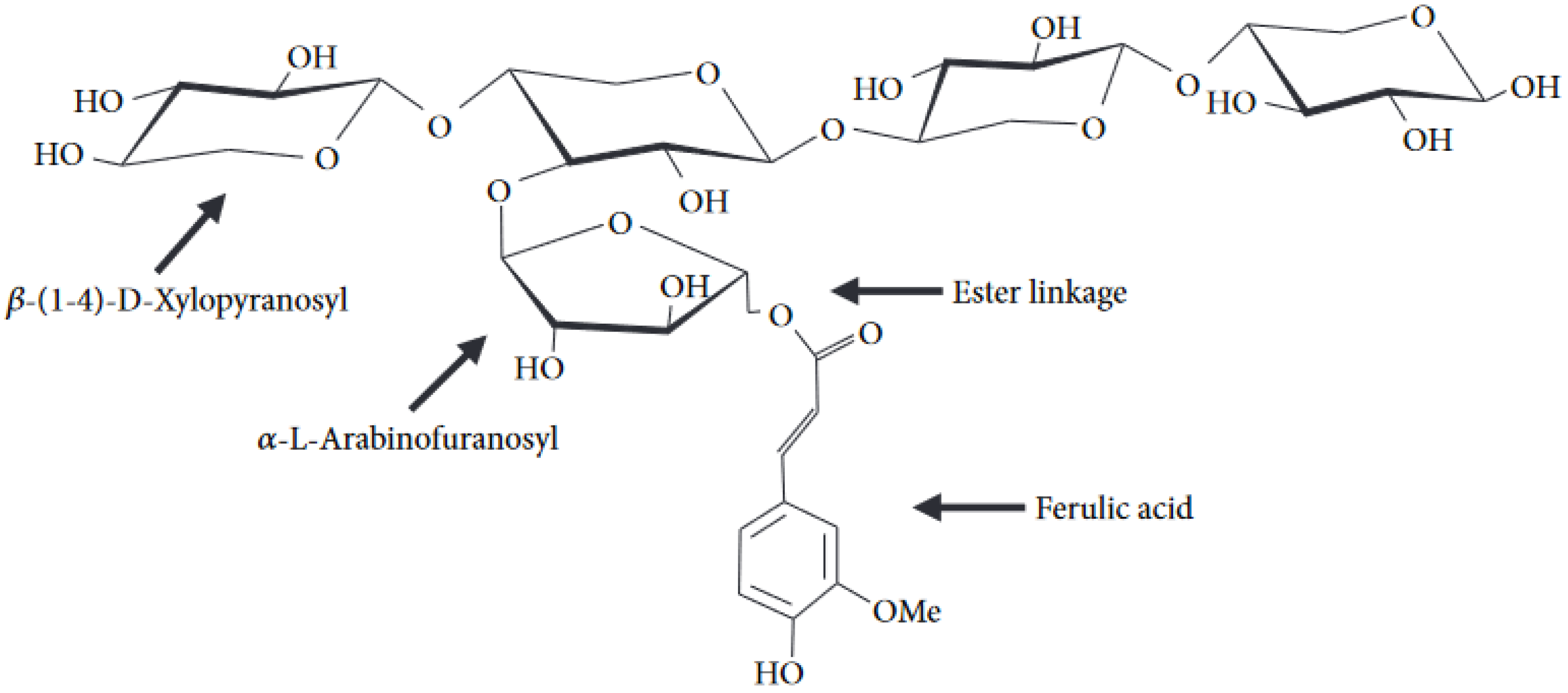{kind=link}
| Aleurone Layer * (mg/100 g dm) | Bran * (mg/100 g dm) | Whole Grain (mg/100 g dm) | |
|---|---|---|---|
| Ferulic acid Total Monomer Dimer Trimer | 628–817 [32,49] 798–814 [23] 31–107 [23,32,49] 2–15 [23,49,50] | 500–1500 [3,49] - 101 [49] - | - 86–87 [23] 14–15 [23] 3–4 [23,49] |
| p-Coumaric acid | 15–29 [23,49,50] | 13–16 [10] | 1–3 [10,23,49] |
| Sinapic acid | 6–44 [10,23,46,49] | 11–28 [10] | 4–8 [10,23] |
| p-hydroxybenzoic acid | 2.8 [10] | 1.9–2.2 [10,49] | 0.5 [10] |
| Vanillic acid | 2 [10] | 1.6–3.5 [10,49] | 0.5–2.1 [10] |
| Syringic acid | 9 [10] | 3.5–5.7 [10,49] | 1.3–1.8 [10] |
| Aleurone Layer * | Bran * | Whole Grain | |
|---|---|---|---|
| Protein (%, w/w) Part in tissue Part in cell-wall Total in grain | ≈30.0 [32,33,54,55,56] 1.0 [36,37] ≈15.0 [32,33,54,55,56] | 15.2–16.9 [3] 9.2 [36] 14.0 [3] | 10.0–15.0 [32,33,54] - - - |
| AMINO ACIDS | |||
| (in %) [54,56] | (in g/16 g N) [54] | ||
| Alanine | 5.9 | 4.9 | 3.5 |
| Arginine | 11.1 | 7.0 | 4.6 |
| Aspartic acid | 7.9 | 7.2 | 5.0 |
| Cysteine | - | 2.0 | 2.2 |
| Glutamic acid | 20.9 | 18.6 | 30.6 |
| Glycine | 5.8 | 7.1 | 3.9 |
| Histidine | 3.4 | 2.6 | 2.2 |
| Isoleucine | 3.6 | 3.5 | 3.8 |
| Leucine | 6.5 | 6.0 | 6.7 |
| Lysine | 4.8 | 4.0 | 2.7 |
| Methionine | 1.6 | 1.6 | 1.7 |
| Phenylalanine | 3.8 | 3.9 | 4.6 |
| Proline | 6.3 | 5.9 | 9.8 |
| Serine | 2.9 | 4.5 | 4.8 |
| Threonine | 2.9 | 3.3 | 2.9 |
| Tryptophan | 4.0 | 1.6 | 1.2 |
| Tyrosine | 3.3 | 2.8 | 3.1 |
| Valine | 5.3 | 5.0 | 4.7 |
2.2.2. Intracellular Medium
3. Health and Nutritional Benefits
3.1. Digestibility and Colonic Fermentation
3.2. Health Benefits of Aleurone Consumption
3.3. Antioxidant Capacity
3.4. Bioactive Components of Aleurone and Related Potential Nutritional and Health Benefits
4. Potential of the Aleurone Layer as an Ingredient in Bread- and Cereal-Based Products
4.1. Extraction of the Aleurone Layer and Its Challenges
| Origin of Products * | 1a | 1b | 2 | 3 | 4 |
|---|---|---|---|---|---|
| ASH (g/100 g) | 9.3–13.3 [12,124,125] | ≈ 10.0–13.3 [7,12,126,127] | 4.1 [85] | 7.2–7.34 [128,129] | 3.9–5.1 [130,131] |
| MOISTURE (g/100 g) | - | 8.0 [7] | 5.4 [85,87] | 8.14 [128] | 6.7–8.98 [130,131,132] |
| CARBOHYDRATES | |||||
| Arabinoxylans (g/100g) | - | 24.3 [127] | - | - | 14.39 [131] |
| A:X ratio | 0.62 [133] | 0.35–0.46 [7,127,133] | - | - | - |
| β-glucans (g/100 g) | 3.4 [133] | 3.91–4.5 [127,133] | - | - | 1.7 [132] |
| Cellulose (g/100 g) | 10.6 [133] | 6.0 [133] | - | - | - |
| Pentosans (g/100 g) | - | - | - | - | 20.2–21.6 [134] |
| Starch (g/100 g) | 1.9–5.8 [124,133] | 2.2–5.8 [126,127,133] | 36.5 [85,87] | 2.5–12.75 [128,129] | 33.46 [130] |
| Total Dietary Fibers (g/100 g) | 39.7–60.0 [12,124,125,133] | 39.7–49.2 [7,12,126,127,133] | 15.4 [85,87] | 43.36 [128] | 27.90–44.3 [130,131,132] |
| Soluble Dietary Fibers (g/100 g) | 4.1 [125] | - | - | 3.07 [128] | - |
| Insoluble Dietary Fibers (g/100 g) | 50.0 [125] | - | - | 40.15 [128] | - |
| MINERALS AND TRACE ELEMENTS | |||||
| Total (g/100 g) | 5.8–9.8 [124,125] | 7.0–9.8 [7,126,127] | 6.5 [85,87] | 4.5–6.33 [128,129] | 4.1–4.7 [134] |
| Calcium (Ca) (mg/100 g) | 76.2 [125] | 93 [10] | - | - | - |
| Copper (Cu) (mg/100 g) | - | - | - | 1.35 [128] | - |
| Iron (Fe) (mg/100 g) | 21.3 [125] | 26 [10] | - | 13.93 [128] | - |
| Magnesium (Mg) (mg/100 g) | 690–800 [12,125] | 850–1030 [10,12] | - | 770 [128] | - |
| Manganese (Mn) (mg/100 g) | - | - | - | 12.7 [128] | - |
| Sodium (Na) (mg/100 g) | 1.7 [125] | - | - | - | - |
| Phosphorus (P) (mg/100 g) | 1900 [125] | 2540 [10] | - | 1857 [128] | - |
| Potassium (K) (mg/100 g) | 1900 [125] | 2250 [10] | - | 1780 [128] | - |
| Zinc (Zn) (mg/100 g) | 11.4 [125] | 14.0 [10] | - | 12.05 [128] | - |
| PHENOLIC COMPOUNDS | |||||
| Total phenolic acids (mg/100 g) | - | - | - | - | 457 [132] |
| Total hydroxycinnamic acids Free (mg/100 g) Bound (mg/100 g) | 1.28 [135] 47.58 [135] | 1.22 [135] 60.65 [135] | - | - | - |
| p-Coumaric acid Total (mg/100 g) Free (mg/100 g) Bound (mg/100 g) Conjugated (mg/100 g) | - - 0.60 [135] - | 16.0 [126] 1.0–1.5 [126] 0.99–1.0 [126,135] 0 [126] | - | - | - |
| Sinapic acid—bound form (mg/100 g) | 0.46 [135] | 0.53 [135] | - | - | - |
| Alkylresorcinols (mg/100 g) | 1107 [135] | 993.24 [135] | - | - | 138 [132] |
| Flavonoids (mg/100 g) | 9.65 [135] | 8.95 [135] | - | - | - |
| Lignans (mg/100 g) | 6300 [133] | 4700 [133] | - | - | - |
| Phytic acid (mg/100 g) | 6900 [125] | - | 2360 [85,87] | - | - |
| PROTEINS | |||||
| Total (g/100 g) | 13.3–18.0 [12,124,125] | 21.0–22.2 [7,12,126,127] | 23.6 [85,87] | 15.2–17.46 [128,129] | 16.5–21 [130,131,132] |
| VITAMINS | |||||
| Total (mg/100 g) | >29.0 [125] | 30.0 [126] | - | - | - |
| Thiamin (B1) (mg/100 g) | 0.87–1.6 [12,125] | 1.1–1.4 [10,12,126] | - | 1.26 [128] | - |
| Riboflavin (B2) (mg/100 g) | 0.3 [125] | 0.2–0.3 [10,126] | - | 0.32 [128] | - |
| Niacin (B3) (mg/100 g) | 24.0 [125] | 21.0–32.9 [10,126] | - | 22.87 [128] | - |
| Pantothenic acid (B5) (mg/100 g) | - | 5.0 [126] | - | 1.87 [128] | - |
| Pyridoxin (B6) (mg/100 g) | 0.3 [125] | 1.3–1.4 [10,126] | - | 2.52 [128] | - |
| Folate (B9) (mg/100 g) | 0.8 [125] | 0.1–0.2 [10,126] | - | 0.2 [128] | - |
| Tocopherols and tocotrienols (E) (mg/100 g) | 2.0 [125] | 0.8–1.2 [10,126] | - | - | - |
4.2. Application to Breadmaking
4.2.1. Aleurone Bread Nutritional Profile
4.2.2. Aleurone Bread Dough Characteristics
4.2.3. End-Product: Aleurone Bread Characteristics
4.2.4. Underlying Mechanisms
4.3. Application to Other Food Products
5. Optimization of the Aleurone Layer’s Potential
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björck, I.; Östman, E.; Kristensen, M.; Mateo Anson, N.; Price, R.K.; Haenen, G.R.M.M.; Havenaar, R.; Bach Knudsen, K.E.; Frid, A.; Mykkänen, H.; et al. Cereal grains for nutrition and health benefits: Overview of results from in vitro, animal and human studies in the HEALTHGRAIN project. Trends Food Sci. Technol. 2012, 25, 87–100. [Google Scholar] [CrossRef]
- Chalamacharla, R.B.; Harsha, K.; Sheik, K.B.; Viswanatha, C.K. Wheat Bran-Composition and Nutritional Quality: A Review. Adv. Biotechnol. Microbiol. 2018, 9, 7. [Google Scholar]
- O’Dell, B.L.; De Boland, A.R.; Koirtyohann, S.R. Distribution of phytate and nutritionally important elements among the morphological components of cereal grains. J. Agric. Food Chem. 1972, 20, 718–723. [Google Scholar] [CrossRef]
- Curti, E.; Carini, E.; Bonacini, G.; Tribuzio, G.; Vittadini, E. Effect of the addition of bran fractions on bread properties. J. Cereal Sci. 2013, 57, 325–332. [Google Scholar] [CrossRef]
- Berger, M.; Ducroo, P. Arabinoxylanes et arabinoxylanases. Ind. Des Céréales 2004, 56, 3–14. [Google Scholar]
- Bonnand-Ducasse, M.; Della Valle, G.; Lefebvre, J.; Saulnier, L. Effect of wheat dietary fibres on bread dough development and rheological properties. J. Cereal Sci. 2010, 52, 200–206. [Google Scholar] [CrossRef]
- Arufe, S.; Chiron, H.; Doré, J.; Savary-Auzeloux, I.; Saulnier, L.; Della Valle, G. Processing & rheological properties of wheat flour dough and bread containing high levels of soluble dietary fibres blends. Food Res. Int. 2017, 97, 123–132. [Google Scholar] [CrossRef]
- Antoine, C.; Lullien, V.; Abécassis, J.; Rouau, X. Intérêt nutritionnel de la couche à aleurone du grain de blé. Sci. Des Aliment. 2002, 22, 545–556. [Google Scholar] [CrossRef]
- Brouns, F.; Hemery, Y.; Price, R.; Anson, N.M. Wheat aleurone: Separation, composition, health aspects, and potential food use. Crit. Rev. Food Sci. Nutr. 2012, 52, 553–568. [Google Scholar] [CrossRef] [Green Version]
- Meziani, S.; Nadaud, I.; Tasleem-Tahir, A.; Nurit, E.; Benguella, R.; Branlard, G. Wheat aleurone layer: A site enriched with nutrients and bioactive molecules with potential nutritional opportunities for breeding. J. Cereal Sci. 2021, 100, 103225. [Google Scholar] [CrossRef]
- Atwell, B.; Reding, W.; Earling, J.; Kanter, M.; Snow, K. Aleurone: Processing, Nutrition, Product Development, and Marketing. In Whole Grains and Health; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 123–136. ISBN 978-0-470-27760-7. [Google Scholar]
- Bagdi, A. Utilization of Wheat Aleurone-Rich Flour in Pasta and Bread Production and the Oxidative Modification of Its Arabinoxylan Fraction; Budapest University of Technology and Economic: Budapest, Hungary, 2016. [Google Scholar]
- Stone, B.A.; Minifie, J. Recovery of Aleurone Cells from Wheat Bran. U.S. Patent US4746073A, 24 May 1988. [Google Scholar]
- Bagdi, A.; Tóth, B.; Lőrincz, R.; Szendi, S.; Gere, A.; Kókai, Z.; Sipos, L.; Tömösközi, S. Effect of aleurone-rich flour on composition, baking, textural, and sensory properties of bread. LWT Food Sci. Technol. 2016, 65, 762–769. [Google Scholar] [CrossRef]
- Bartalné-Berceli, M.; Izsó, E.; Gergely, S.; Salgó, A. Development and application of novel additives in bread-making. Czech J. Food Sci. 2018, 36, 470–475. [Google Scholar] [CrossRef] [Green Version]
- Bartalné-Berceli, M.; Izsó, E.; Gergely, S.; Salgó, A. Bread Quality Improvement with Special Novel Additives. Int. J. Biol. Biomol. Agric. Food Biotechnol. Eng. 2015, 9, 5. [Google Scholar]
- Gómez, M.; Gutkoski, L.C.; Bravo-Núñez, Á. Understanding whole-wheat flour and its effect in breads: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3241–3265. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Sibakov, N.; Poutanen, K.; Micard, V. How does wheat grain, bran and aleurone structure impact their nutritional and technological properties? Trends Food Sci. Technol. 2015, 41, 118–134. [Google Scholar] [CrossRef]
- Laubin, B.B.; Lullien-Pellerin, V.V.; Nadaud, I.; Gaillard-Martinie, B.B.; Chambon, C.C.; Branlard, G.G. Isolation of the wheat aleurone layer for 2D electrophoresis and proteomics analysis. J. Cereal Sci. 2008, 48, 709–714. [Google Scholar] [CrossRef]
- Surget, A.; Barron, C. Histologie du grain de blé. Ind. Des Céréales 2005, 145, 3–7. [Google Scholar]
- Rhodes, D.I.; Sadek, M.; Stone, B.A. Hydroxycinnamic Acids in Walls of Wheat Aleurone Cells. J. Cereal Sci. 2002, 36, 67–81. [Google Scholar] [CrossRef]
- Barron, C.; Surget, A.; Rouau, X. Relative amounts of tissues in mature wheat (Triticum aestivum L.) grain and their carbohydrate and phenolic acid composition. J. Cereal Sci. 2007, 45, 88–96. [Google Scholar] [CrossRef]
- Xiong, F.; Yu, X.-R.; Zhou, L.; Wang, Z.; Wang, F.; Xiong, A.-S. Structural development of aleurone and its function in common wheat. Mol. Biol. Rep. 2013, 40, 6785–6792. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Fincher, G.B. Evolution and development of cell walls in cereal grains. Front. Plant Sci. 2014, 5, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becraft, P.W. Aleurone Cell Development. Plant Cell Monogr. 2007, 8, 45–56. [Google Scholar] [CrossRef]
- Bacic, A.; Stone, B.A. Isolation and Ultrastructure of Aleurone Cell Walls From Wheat and Barley. Funct. Plant Biol. 1981, 8, 453–474. [Google Scholar] [CrossRef]
- Meziani, S.; Nadaud, I.; Gaillard-Martinie, B.; Chambon, C.; Benali, M.; Branlard, G. Proteomic analysis of the mature kernel aleurone layer in common and durum wheat. J. Cereal Sci. 2012, 55, 323–330. [Google Scholar] [CrossRef]
- Saulnier, L.; Sado, P.-E.; Branlard, G.; Charmet, G.; Guillon, F. Wheat arabinoxylans: Exploiting variation in amount and composition to develop enhanced varieties. J. Cereal Sci. 2007, 46, 261–281. [Google Scholar] [CrossRef]
- Saulnier, L.; Guillon, F.; Chateigner-Boutin, A.-L. Cell wall deposition and metabolism in wheat grain. J. Cereal Sci. 2012, 56, 91–108. [Google Scholar] [CrossRef]
- Robert, P.; Jamme, F.; Barron, C.; Bouchet, B.; Saulnier, L.; Dumas, P.; Guillon, F. Change in wall composition of transfer and aleurone cells during wheat grain development. Planta 2011, 233, 393–406. [Google Scholar] [CrossRef]
- Feillet, P. Le Grain de blé: Composition et Utilisation; INRA: Paris, France, 2000; ISBN 2-7380-0896-8. [Google Scholar]
- Lopez, H.W.; Adam, A.; Leenhardt, F.; Scalbert, A.; Remesy, C. Maîtrise de la valeur nutritionnelle du pain. Ind. Des Céréales 2001, 124, 15–20. [Google Scholar]
- Andersson, R.; Andersson, A.; Åman, P. Chapter 16—Molecular Weight Distributions of Water-Extractable Β-Glucan and Arabinoxylan. In Healthgrain Methods; Shewry, P.R., Ward, J.L., Eds.; American Associate of Cereal Chemists International; AACC International Press: Washington, DC, USA, 2009; pp. 203–216. ISBN 978-1-891127-70-0. [Google Scholar]
- Zeng, H.; Lazarova, D.L.; Bordonaro, M. Mechanisms linking dietary fiber, gut microbiota and colon cancer prevention. World J. Gastrointest. Oncol. 2014, 6, 41–51. [Google Scholar] [CrossRef]
- Fincher, G.B.; Stone, B.A. CEREALS | Chemistry of Nonstarch Polysaccharides. In Encyclopedia of Grain Science; Wrigley, C., Ed.; Elsevier: Oxford, UK, 2004; pp. 206–223. ISBN 978-0-12-765490-4. [Google Scholar]
- Stone, B.; Morell, M.K. Chapter 9—Carbohydrates. In Wheat, 4th ed.; Khan, K., Shewry, P.R., Eds.; American Associate of Cereal Chemists International; AACC International Press: Washington, DC, USA, 2009; pp. 299–362. ISBN 978-1-891127-55-7. [Google Scholar]
- Mendez-Encinas, M.A.; Carvajal-Millan, E.; Rascon-Chu, A.; Astiazaran-Garcia, H.F.; Valencia-Rivera, D.E. Ferulated Arabinoxylans and Their Gels: Functional Properties and Potential Application as Antioxidant and Anticancer Agent. Oxidative Med. Cell. Longev. 2018, 2018, e2314759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtin, C.M.; Delcour, J.A. Arabinoxylans and Endoxylanases in Wheat Flour Bread-making. J. Cereal Sci. 2002, 35, 225–243. [Google Scholar] [CrossRef]
- Döring, C.; Jekle, M.; Becker, T. Technological and Analytical Methods for Arabinoxylan Quantification from Cereals. Crit. Rev. Food Sci. Nutr. 2016, 56, 999–1011. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.L.; Ng, A.; Waldron, K.W. The phenolic acid and polysaccharide composition of cell walls of bran layers of mature wheat (Triticum aestivum L. cv. Avalon) grains. J. Sci. Food Agric. 2005, 85, 2539–2547. [Google Scholar] [CrossRef]
- Jamme, F.; Robert, P.; Bouchet, B.; Saulnier, L.; Dumas, P.; Guillon, F. Aleurone cell walls of wheat grain: High spatial resolution investigation using synchrotron infrared microspectroscopy. Appl. Spectrosc. 2008, 62, 895–900. [Google Scholar] [CrossRef]
- Collins, H.M.; Burton, R.A.; Topping, D.L.; Liao, M.-L.; Bacic, A.; Fincher, G.B. REVIEW: Variability in Fine Structures of Noncellulosic Cell Wall Polysaccharides from Cereal Grains: Potential Importance in Human Health and Nutrition. Cereal Chem. 2010, 87, 272–282. [Google Scholar] [CrossRef]
- Antoine, C.; Peyron, S.; Mabille, F.; Lapierre, C.; Bouchet, B.; Abecassis, J.; Rouau, X. Individual Contribution of Grain Outer Layers and Their Cell Wall Structure to the Mechanical Properties of Wheat Bran. J. Agric. Food Chem. 2003, 51, 2026–2033. [Google Scholar] [CrossRef]
- Mateo Anson, N.; van den Berg, R.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Ferulic acid from aleurone determines the antioxidant potency of wheat grain (Triticum aestivum L.). J. Agric. Food Chem. 2008, 56, 5589–5594. [Google Scholar] [CrossRef]
- Antoine, C.; Peyron, S.; Lullien, V.; Abécassis, J.; Rouau, X. Wheat bran tissue fractionation using biochemical markers. J. Cereal Sci. 2004, 39, 387–393. [Google Scholar] [CrossRef]
- Van Hung, P.; Maeda, T.; Miyatake, K.; Morita, N. Total phenolic compounds and antioxidant capacity of wheat graded flours by polishing method. Food Res. Int. 2009, 42, 185–190. [Google Scholar] [CrossRef]
- Žilić, S. Phenolic Compounds of Wheat. Their Content, Antioxidant Capacity and Bioaccessibility. MOJ Food Process. Technol. 2016, 2, 37. [Google Scholar] [CrossRef]
- Piironen, V.; Lampi, A.-M.; Ekholm, P.; Salmenkallio-Marttila, M.; Liukkonen, K.-H. Chapter 7—Micronutrients and Phytochemicals in Wheat Grain. In Wheat, 4th ed.; Khan, K., Shewry, P.R., Eds.; American Associate of Cereal Chemists International; AACC International Press: Washington, DC, USA, 2009; pp. 179–222. ISBN 978-1-891127-55-7. [Google Scholar]
- Barron, C.; Samson, M.F.; Lullien-Pellerin, V.; Rouau, X. Wheat grain tissue proportions in milling fractions using biochemical marker measurements: Application to different wheat cultivars. J. Cereal Sci. 2011, 53, 306–311. [Google Scholar] [CrossRef]
- Jääskeläinen, A.-S.; Holopainen-Mantila, U.; Tamminen, T.; Vuorinen, T. Endosperm and aleurone cell structure in barley and wheat as studied by optical and Raman microscopy. J. Cereal Sci. 2013, 57, 543–550. [Google Scholar] [CrossRef]
- Rhodes, D.I.; Stone, B.A. Proteins in Walls of Wheat Aleurone Cells. J. Cereal Sci. 2002, 36, 83–101. [Google Scholar] [CrossRef]
- Bacic, A.; Stone, B.A. Chemistry and Organization of Aleurone Cell Wall Components From Wheat and Barley. Funct. Plant Biol. 1981, 8, 475–495. [Google Scholar] [CrossRef]
- Pomeranz, Y. Wheat: Chemistry and Technology, 3rd ed.; Pomeranz, Y., Ed.; AACC Monograph Series; American Association of Cereal Chemists: St. Paul, MN, USA, 1988; Volume I, ISBN 978-0-913250-65-5. [Google Scholar]
- Wieser, H.; Koehler, P.; Scherf, K.A. Chapter 2—Chemical Composition of Wheat Grains. In Wheat-An Exceptional Crop; Wieser, H., Koehler, P., Scherf, K.A., Eds.; Woodhead Publishing: Cambridge, UK, 2020; pp. 13–45. ISBN 978-0-12-821715-3. [Google Scholar]
- Shewry, P.R.; D’Ovidio, R.; Lafiandra, D.; Jenkins, J.A.; Mills, E.N.C.; Békés, F. Chapter 8—Wheat Grain Proteins. In Wheat, 4th ed.; Khan, K., Shewry, P.R., Eds.; American Associate of Cereal Chemists International; AACC International Press: Washington, DC, USA, 2009; pp. 223–298. ISBN 978-1-891127-55-7. [Google Scholar]
- Amrein, T.M.; Gränicher, P.; Arrigoni, E.; Amadò, R. In vitro digestibility and colonic fermentability of aleurone isolated from wheat bran. LWT-Food Sci. Technol. 2003, 36, 451–460. [Google Scholar] [CrossRef]
- Buttrose, M. Ultrastructure of the Developing Aleurone Cells of Wheat Grain. Aust. J. Biol. Sci. 1963, 16, 768. [Google Scholar] [CrossRef] [Green Version]
- Marion, D.; Dubreil, L.; Douliez, J.-P. Functionality of lipids and lipid-protein interactions in cereal-derived food products. Ol. Corps Gras Lipides 2003, 10, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Morrison, I.N.; Kuo, J.; O’Brien, T.P. Histochemistry and fine structure of developing wheat aleurone cells. Planta 1975, 123, 105–116. [Google Scholar] [CrossRef]
- Yan, X.; Ye, R.; Chen, Y. Blasting extrusion processing: The increase of soluble dietary fiber content and extraction of soluble-fiber polysaccharides from wheat bran. Food Chem. 2015, 180, 106–115. [Google Scholar] [CrossRef]
- Arte, E.; Huang, X.; Nordlund, E.; Katina, K. Biochemical characterization and technofunctional properties of bioprocessed wheat bran protein isolates. Food Chem. 2019, 289, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndolo, V.U.; Beta, T. Distribution of carotenoids in endosperm, germ, and aleurone fractions of cereal grain kernels. Food Chem. 2013, 139, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.F.; Hollis, J.H.; Migaud, M.; Browne, R.A. Analysis of Betaine and Choline Contents of Aleurone, Bran, and Flour Fractions of Wheat (Triticum aestivum L.) Using 1H Nuclear Magnetic Resonance (NMR) Spectroscopy. J. Agric. Food Chem. 2009, 57, 1948–1951. [Google Scholar] [CrossRef] [PubMed]
- Hemery, Y.; Lullien-Pellerin, V.; Rouau, X.; Abecassis, J.; Samson, M.-F.; Åman, P.; von Reding, W.; Spoerndli, C.; Barron, C. Biochemical markers: Efficient tools for the assessment of wheat grain tissue proportions in milling fractions. J. Cereal Sci. 2009, 49, 55–64. [Google Scholar] [CrossRef]
- Rosentrater, K.A.; Evers, A.D. Chapter 4—Chemical Components and Nutrition. In Kent’s Technology of Cereals, 5th ed.; Rosentrater, K.A., Evers, A.D., Eds.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Cambridge, UK, 2018; pp. 267–368. ISBN 978-0-08-100529-3. [Google Scholar]
- Landberg, R.; Kamal-Eldin, A.; Salmenkallio-Marttila, M.; Rouau, X.X.; Aman, P. Localization of alkylresorcinols in wheat, rye and barley kernels. J. Cereal Sci. 2008, 48, 401–406. [Google Scholar] [CrossRef]
- Chen, Z.; Zha, B.; Wang, L.; Wang, R.; Chen, Z.; Tian, Y. Dissociation of aleurone cell cluster from wheat bran by centrifugal impact milling. Food Res. Int. 2013, 54, 63–71. [Google Scholar] [CrossRef]
- Batifoulier, F.; Verny, M.-A.; Chanliaud, E.; Rémésy, C.; Demigné, C. Variability of B vitamin concentrations in wheat grain, milling fractions and bread products. Eur. J. Agron. 2006, 25, 163–169. [Google Scholar] [CrossRef]
- Saini, P.; Kumar, N.; Kumar, S.; Mwaurah, P.W.; Panghal, A.; Attkan, A.K.; Singh, V.K.; Garg, M.K.; Singh, V. Bioactive compounds, nutritional benefits and food applications of colored wheat: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2021, 61, 3197–3210. [Google Scholar] [CrossRef]
- Garg, M.; Kaur, S.; Sharma, A.; Kumari, A.; Tiwari, V.; Sharma, S.; Kapoor, P.; Sheoran, B.; Goyal, A.; Krishania, M. Rising Demand for Healthy Foods-Anthocyanin Biofortified Colored Wheat Is a New Research Trend. Front. Nutr. 2022, 9, 913. [Google Scholar] [CrossRef]
- Zheng, Q.; Li, B.; Li, H.; Li, Z. Utilization of blue-grained character in wheat breeding derived from Thinopyrum poticum. J. Genet. Genom. 2009, 36, 575–580. [Google Scholar] [CrossRef]
- Mateo Anson, N.; van den Berg, R.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Bioavailability of ferulic acid is determined by its bioaccessibility. J. Cereal Sci. 2009, 49, 296–300. [Google Scholar] [CrossRef]
- Bach Knudsen, K.E.; Steenfeldt, S.; Børsting, C.F.; Eggum, B.O. The nutritive value of decorticated mill fractions of wheat. 1. Chemical composition of raw and enzyme treated fractions and balance experiments with rats. Anim. Feed Sci. Technol. 1995, 52, 205–225. [Google Scholar] [CrossRef]
- D’hoe, K.; Conterno, L.; Fava, F.; Falony, G.; Vieira-Silva, S.; Vermeiren, J.; Tuohy, K.; Raes, J. Prebiotic Wheat Bran Fractions Induce Specific Microbiota Changes. Front. Microbiol. 2018, 9, 31. [Google Scholar] [CrossRef]
- Demuth, T.; Betschart, J.; Nyström, L. Structural modifications to water-soluble wheat bran arabinoxylan through milling and extrusion. Carbohydr. Polym. 2020, 240, 116328. [Google Scholar] [CrossRef] [PubMed]
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 2009, 53, S330–S375. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.J.; Ferguson, L.R. Dietary fibres may protect or enhance carcinogenesis. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1999, 443, 95–110. [Google Scholar] [CrossRef]
- Sagara, M.; Mori, M.; Mori, H.; Tsuchikura, S.; Yamori, Y. Effect of dietary wheat aleurone on blood pressure and blood glucose and its mechanisms in obese spontaneously hypertensive rats: Preliminary report on comparison with a soy diet (Clinical and Experimental Pharmacology and Physiology. Clin. Exp. Pharmacol. Physiol. 2007, 34, S37–S39. [Google Scholar] [CrossRef]
- Price, R.K.; Keaveney, E.M.; Hamill, L.L.; Wallace, J.M.W.; Ward, M.; Ueland, P.M.; McNulty, H.; Strain, J.J.; Parker, M.J.; Welch, R.W. Consumption of wheat aleurone-rich foods increases fasting plasma betaine and modestly decreases fasting homocysteine and LDL-cholesterol in adults. J. Nutr. 2010, 140, 2153–2157. [Google Scholar] [CrossRef] [Green Version]
- Borowicki, A.; Stein, K.; Scharlau, D.; Glei, M. Fermentation Supernatants of Wheat (Triticum aestivum L.) Aleurone Beneficially Modulate Cancer Progression in Human Colon Cells. J. Agric. Food Chem. 2010, 58, 2001–2007. [Google Scholar] [CrossRef]
- Borowicki, A.; Stein, K.; Scharlau, D.; Scheu, K.; Brenner-Weiss, G.; Obst, U.; Hollmann, J.; Lindhauer, M.; Wachter, N.; Glei, M. Fermented wheat aleurone inhibits growth and induces apoptosis in human HT29 colon adenocarcinoma cells. Br. J. Nutr. 2010, 103, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Borowicki, A.; Michelmann, A.; Stein, K.; Scharlau, D.; Scheu, K.; Obst, U.; Glei, M. Fermented Wheat Aleurone Enriched With Probiotic Strains LGG and Bb12 Modulates Markers of Tumor Progression in Human Colon Cells. Nutr. Cancer 2011, 63, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Stein, K.; Borowicki, A.; Scharlau, D.; Glei, M. Fermented wheat aleurone induces enzymes involved in detoxification of carcinogens and in antioxidative defence in human colon cells. Br. J. Nutr. 2010, 104, 1101–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenech, M.; Noakes, M.; Clifton, P.; Topping, D. Aleurone flour is a rich source of bioavailable folate in humans. Am. Soc. Nutr. Sci. 1999, 129, 1114–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenech, M.; Noakes, M.; Clifton, P.; Topping, D. Aleurone flour increases red-cell folate and lowers plasma homocyst(e)ine substantially in man. Br. J. Nutr. 2005, 93, 353–360. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, G.H.; Royle, P.J.; Pointing, G. Wheat Aleurone Flour Increases Cecal β-Glucuronidase Activity and Butyrate Concentration and Reduces Colon Adenoma Burden in Azoxymethane-Treated Rats. J. Nutr. 2001, 131, 127–131. [Google Scholar] [CrossRef]
- Keaveney, E.M.; Hamill, L.L.; Price, R.K.; Wallace, J.M.W.; McNulty, H.; Ward, M.; Strain, J.J.; Ueland, P.M.; Scott, J.M.; Molloy, A.M.; et al. Evaluation of the uptake of bioactive components from wheat-bran and wheat-aleurone fractions in healthy adults. Proc. Nutr. Soc. 2009, 67, E244. [Google Scholar] [CrossRef] [Green Version]
- Fava, F.; Ulaszewska, M.M.; Scholz, M.; Stanstrup, J.; Nissen, L.; Mattivi, F.; Vermeiren, J.; Bosscher, D.; Pedrolli, C.; Tuohy, K.M. Impact of wheat aleurone on biomarkers of cardiovascular disease, gut microbiota and metabolites in adults with high body mass index: A double-blind, placebo-controlled, randomized clinical trial. Eur. J. Nutr. 2022, 61, 2651–2671. [Google Scholar] [CrossRef]
- Mahdavi Sharif, P.; Jabbari, P.; Razi, S.; Keshavarz-Fathi, M.; Rezaei, N. Importance of TNF-alpha and its alterations in the development of cancers. Cytokine 2020, 130, 155066. [Google Scholar] [CrossRef]
- Mateo Anson, N.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Antioxidant and anti-inflammatory capacity of bioaccessible compounds from wheat fractions after gastrointestinal digestion. J. Cereal Sci. 2010, 51, 110–114. [Google Scholar] [CrossRef]
- Shrivastava, A.K.; Singh, H.V.; Raizada, A.; Singh, S.K. C-reactive protein, inflammation and coronary heart disease. Egypt. Heart J. 2015, 67, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Price, R.K.; Wallace, J.M.W.; Hamill, L.L.; Keaveney, E.M.; Strain, J.J.; Parker, M.J.; Welch, R.W. Evaluation of the effect of wheat aleurone-rich foods on markers of antioxidant status, inflammation and endothelial function in apparently healthy men and women. Br. J. Nutr. 2012, 108, 1644–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamill, L.L.; Keaveney, E.M.; Price, R.K.; Wallace, J.M.W.; Strain, J.J.; Welch, R.W. Absorption of ferulic acid in human subjects after consumption of wheat-bran and wheat-aleurone fractions. Proc. Nutr. Soc. 2008, 67, E255. [Google Scholar] [CrossRef] [Green Version]
- Bresciani, L.; Scazzina, F.; Leonardi, R.; Dall’Aglio, E.; Newell, M.; Dall’Asta, M.; Melegari, C.; Ray, S.; Brighenti, F.; Del Rio, D. Bioavailability and metabolism of phenolic compounds from wholegrain wheat and aleurone-rich wheat bread. Mol. Nutr. Food Res. 2016, 60, 2343–2354. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, M.; Bresciani, L.; Calani, L.; Cossu, M.; Martini, D.; Melegari, C.; Del Rio, D.; Pellegrini, N.; Brighenti, F.; Scazzina, F. In Vitro Bioaccessibility of Phenolic Acids from a Commercial Aleurone-Enriched Bread Compared to a Whole Grain Bread. Nutrients 2016, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graf, E. Antioxidant potential of ferulic acid. Free Radic. Biol. Med. 1992, 13, 435–448. [Google Scholar] [CrossRef]
- Toda, S.; Kimura, M.; Ohnishi, M. Effects of Phenolcarboxylic Acids on Superoxide Anion and Lipid Peroxidation Induced by Superoxide Anion. Planta Med. 1991, 57, 8–10. [Google Scholar] [CrossRef]
- Rietjens, S.J.; Bast, A.; de Vente, J.; Haenen, G.R.M.M. The olive oil antioxidant hydroxytyrosol efficiently protects against the oxidative stress-induced impairment of the NO• response of isolated rat aorta. Am. J. Physiol.-Heart Circ. Physiol. 2007, 292, H1931–H1936. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Moghadasian, M.H. Chemistry, natural sources, dietary intake and pharmacokinetic properties of ferulic acid: A review. Food Chem. 2008, 109, 691–702. [Google Scholar] [CrossRef]
- Castelluccio, C.; Bolwell, G.P.; Gerrish, C.; Rice-Evans, C. Differential distribution of ferulic acid to the major plasma constituents in relation to its potential as an antioxidant. Biochem. J. 1996, 316, 691–694. [Google Scholar] [CrossRef] [Green Version]
- Andreasen, M.F.; Kroon, P.A.; Williamson, G.; Garcia-Conesa, M.-T. Esterase Activity Able To Hydrolyze Dietary Antioxidant Hydroxycinnamates Is Distributed along the Intestine of Mammals. J. Agric. Food Chem. 2001, 49, 5679–5684. [Google Scholar] [CrossRef]
- Vitaglione, P.; Napolitano, A.; Fogliano, V. Cereal dietary fibre: A natural functional ingredient to deliver phenolic compounds into the gut. Trends Food Sci. Technol. 2008, 19, 451–463. [Google Scholar] [CrossRef]
- Mateo Anson, N.; Aura, A.; Selinheimo, E.; Mattila, I.; Poutanen, K.; van den Berg, R.; Havenaar, R.; Bast, A.; Haenen, G.R.M.M. Bioprocessing of Wheat Bran in Whole Wheat Bread Increases the Bioavailability of Phenolic Acids in Men and Exerts Antiinflammatory Effects ex Vivo. J. Nutr. 2011, 141, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinonen, S.; Nurmi, T.; Liukkonen, K.; Poutanen, K.; Wähälä, K.; Deyama, T.; Nishibe, S.; Adlercreutz, H. In Vitro Metabolism of Plant Lignans: New Precursors of Mammalian Lignans Enterolactone and Enterodiol. J. Agric. Food Chem. 2001, 49, 3178–3186. [Google Scholar] [CrossRef]
- Kamei, H.; Kojima, T.; Hasegawa, M.; Koide, T.; Umeda, T.; Yukawa, T.; Terabe, K. Suppression of Tumor Cell Growth by Anthocyanins In Vitro. Cancer Investig. 1995, 13, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Giusti, M.M.; Malik, M.; Moyer, M.P.; Magnuson, B.A. Effects of Commercial Anthocyanin-Rich Extracts on Colonic Cancer and Nontumorigenic Colonic Cell Growth. J. Agric. Food Chem. 2004, 52, 6122–6128. [Google Scholar] [CrossRef]
- Lampi, A.-M.; Nurmi, T.; Ollilainen, V.; Piironen, V. Tocopherols and Tocotrienols in Wheat Genotypes in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2008, 56, 9716–9721. [Google Scholar] [CrossRef] [PubMed]
- Mooney, S.; Leuendorf, J.-E.; Hendrickson, C.; Hellmann, H. Vitamin B6: A Long Known Compound of Surprising Complexity. Molecules 2009, 14, 329–351. [Google Scholar] [CrossRef] [Green Version]
- Ndolo, V.U.; Fulcher, R.G.; Beta, T. Application of LC–MS–MS to identify niacin in aleurone layers of yellow corn, barley and wheat kernels. J. Cereal Sci. 2015, 65, 88–95. [Google Scholar] [CrossRef]
- Bohm, A.; Buri, R. Composition Comprising Aleurone, Method of Administering the Same, and Method of Manufacturing the Same. U.S. Patent US20040258776A1, 26 May 2004. [Google Scholar]
- Bohm, A.; Kratzer, A. Method for Isolating Aleurone Particles. U.S. Patent US7780101B2, 24 August 2008. [Google Scholar]
- Bohm, A.; Bogoni, C.; Behrens, R.; Otto, T. Method for the Extraction of Aleurone from Bran. U.S. Patent US20030175384A1, 4 October 2011. [Google Scholar]
- Laux, J.; Von Reding, W.; Widmer, G. Aleurone Product and Corresponding Production Method. Patent WO2004073416A1, 13 January 2004. [Google Scholar]
- Barron, C.; Abecassis, J.; Chaurand, M.; Lullien-Pellerin, V.; Mabille, F.; Rouau, X.; Sadouti, A.; Samson, M.-F. Accès à des molécules d’intérêt par fractionnement par voie sèche. Innov. Agron. 2012, 19, 51–62. [Google Scholar]
- Hemery, Y.; Rouau, X.; Lullien-Pellerin, V.; Barron, C.; Abecassis, J. Dry processes to develop wheat fractions and products with enhanced nutritional quality. J. Cereal Sci. 2007, 46, 327–347. [Google Scholar] [CrossRef]
- Antoine, C.; Lullien-Pellerin, V.; Abecassis, J.; Rouau, X. Effect of wheat bran ball-milling on fragmentation and marker extractability of the aleurone layer. J. Cereal Sci. 2004, 40, 275–282. [Google Scholar] [CrossRef]
- Rosa, N.N.; Barron, C.; Gaiani, C.; Dufour, C.; Micard, V. Ultra-fine grinding increases the antioxidant capacity of wheat bran. J. Cereal Sci. 2013, 57, 84–90. [Google Scholar] [CrossRef]
- Hemery, Y.; Holopainen, U.; Lampi, A.-M.; Lehtinen, P.; Nurmi, T.; Piironen, V.; Edelmann, M.; Rouau, X. Potential of dry fractionation of wheat bran for the development of food ingredients, part II: Electrostatic separation of particles. J. Cereal Sci. 2011, 53, 9–18. [Google Scholar] [CrossRef]
- Tupper, R.J.; Pointing, G.; Westcott, S.J. Bran Products and Methods for Production Thereof. Patent WO2001021012A1, 15 September 2001. [Google Scholar]
- Kvist, S.; Carlsson, T.; Lawther, J.M.; DeCastro, F.B. Process for the Fractionation of Cereal Brans. U.S. Patent US7709033B2, 4 May 2010. [Google Scholar]
- Hemery, Y.; Rouau, X.; Dragan, C.; Bilici, M.; Beleca, R.; Dascalescu, L. Electrostatic properties of wheat bran and its constitutive layers: Influence of particle size, composition, and moisture content. J. Food Eng. 2009, 93, 114–124. [Google Scholar] [CrossRef]
- Andersson, A.A.M.; Kamal-Eldin, A.; Fraś, A.; Boros, D.; Åman, P. Alkylresorcinols in Wheat Varieties in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2008, 56, 9722–9725. [Google Scholar] [CrossRef] [PubMed]
- Pekkinen, J.; Rosa, N.N.; Savolainen, O.-I.; Keski-Rahkonen, P.; Mykkänen, H.; Poutanen, K.; Micard, V.; Hanhineva, K. Disintegration of wheat aleurone structure has an impact on the bioavailability of phenolic compounds and other phytochemicals as evidenced by altered urinary metabolite profile of diet-induced obese mice. Nutr. Metab. 2014, 11, 1. [Google Scholar] [CrossRef]
- Boshuizen, B.; Moreno de Vega, C.V.; De Maré, L.; de Meeûs, C.; de Oliveira, J.E.; Hosotani, G.; Gansemans, Y.; Deforce, D.; Van Nieuwerburgh, F.; Delesalle, C. Effects of Aleurone Supplementation on Glucose-Insulin Metabolism and Gut Microbiome in Untrained Healthy Horses. Front. Vet. Sci. 2021, 8, 642809. [Google Scholar] [CrossRef]
- Rosa, N.N.; Dufour, C.; Lullien-Pellerin, V.; Micard, V. Exposure or release of ferulic acid from wheat aleurone: Impact on its antioxidant capacity. Food Chem. 2013, 141, 2355–2362. [Google Scholar] [CrossRef]
- Rosa, N.N.; Aura, A.-M.; Saulnier, L.; Holopainen-Mantila, U.; Poutanen, K.; Micard, V. Effects of Disintegration on in Vitro Fermentation and Conversion Patterns of Wheat Aleurone in a Metabolical Colon Model. J. Agric. Food Chem. 2013, 61, 5805–5816. [Google Scholar] [CrossRef]
- Wellnitz, J. The Ingredient ‘Wheat Aleurone’ and Nutritional Benefits. In International Nutrition & Health Industry Expo, 3rd ed.; Whole Grain Forum: Beijing, China, 2011. [Google Scholar]
- Nkhata, L.; Izydorczyk, M.; Beta, T. Effect of Water-Extractable Arabinoxylans from Wheat Aleurone and Bran on Lipid Peroxidation and Factors Influencing their Antioxidant Capacity. Bioact. Carbohydr. Diet. Fibre 2017, 10, 20–26. [Google Scholar] [CrossRef]
- Yang, Y.-L.; Guan, E.-Q.; Li, M.; Li, N.; Bian, K.; Wang, T.; Lu, C.; Chen, M.; Xu, F. Effect of transglutaminase on the quality and protein characteristics of aleurone-riched fine dried noodles. LWT 2022, 154, 112584. [Google Scholar] [CrossRef]
- Li, X.; Hu, H.; Xu, F.; Liu, Z.; Zhang, L.; Zhang, H. Effects of aleurone-rich fraction on the hydration and rheological properties attributes of wheat dough. Int. J. Food Sci. Technol. 2018, 54, 1777–1786. [Google Scholar] [CrossRef]
- Xu, M.; Hou, G.G.; Ma, F.; Ding, J.; Deng, L.; Kahraman, O.; Niu, M.; Trivettea, K.; Lee, B.; Wu, L.; et al. Evaluation of aleurone flour on dough, textural, and nutritional properties of instant fried noodles. LWT 2020, 126, 109294. [Google Scholar] [CrossRef]
- Vangsøe, C.T.; Sørensen, J.F.; Bach Knudsen, K.E. Aleurone cells are the primary contributor to arabinoxylan oligosaccharide production from wheat bran after treatment with cell wall-degrading enzymes. Int. J. Food Sci. Technol. 2019, 54, 2847–2853. [Google Scholar] [CrossRef]
- Yang, M.; Li, N.; Wang, A.; Tong, L.; Wang, L.; Yue, Y.; Yao, J.; Zhou, S.; Liu, L. Evaluation of rheological properties, microstructure and water mobility in buns dough enriched in aleurone flour modified by enzyme combinations. Int. J. Food Sci. Technol. 2021, 56, 5913–5922. [Google Scholar] [CrossRef]
- Martín-García, B.; Gómez-Caravaca, A.M.; Marconi, E.; Verardo, V. Distribution of free and bound phenolic compounds, and alkylresorcinols in wheat aleurone enriched fractions. Food Res. Int. 2021, 140, 109816. [Google Scholar] [CrossRef]
- Bucsella, B.; Molnár, D.; Harasztos, A.H.; Tömösközi, S. Comparison of the rheological and end-product properties of an industrial aleurone-rich wheat flour, whole grain wheat and rye flour. J. Cereal Sci. 2016, 69, 40–48. [Google Scholar] [CrossRef]
- Hemdane, S.; Leys, S.; Jacobs, P.J.; Dornez, E.; Delcour, J.A.; Courtin, C.M. Wheat milling by-products and their impact on bread making. Food Chem. 2015, 187, 280–289. [Google Scholar] [CrossRef]
- Bagdi, A.; Szabó, F.; Gere, A.; Kókai, Z.; Sipos, L.; Tömösközi, S. Effect of aleurone-rich flour on composition, cooking, textural, and sensory properties of pasta. LWT-Food Sci. Technol. 2014, 59, 996–1002. [Google Scholar] [CrossRef]
- Noort, M.W.J.; van Haaster, D.; Hemery, Y.; Schols, H.A.; Hamer, R.J. The effect of particle size of wheat bran fractions on bread quality–Evidence for fibre–protein interactions. J. Cereal Sci. 2010, 52, 59–64. [Google Scholar] [CrossRef]
- Tian, B.; Zhou, C.; Li, D.; Pei, J.; Guo, A.; Liu, S.; Li, H. Monitoring the Effects of Hemicellulase on the Different Proofing Stages of Wheat Aleurone-Rich Bread Dough and Bread Quality. Foods 2021, 10, 2427. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Dhital, S.; Zhao, C.; Ye, F.; Chen, J.; Zhao, G. Dietary fiber-gluten protein interaction in wheat flour dough: Analysis, consequences and proposed mechanisms. Food Hydrocoll. 2021, 111, 106203. [Google Scholar] [CrossRef]
- Sivam, A.S.; Sun-Waterhouse, D.; Quek, S.; Perera, C.O. Properties of Bread Dough with Added Fiber Polysaccharides and Phenolic Antioxidants: A Review. J. Food Sci. 2010, 75, R163–R174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudha, M.L.; Vetrimani, R.; Leelavathi, K. Influence of fibre from different cereals on the rheological characteristics of wheat flour dough and on biscuit quality. Food Chem. 2007, 100, 1365–1370. [Google Scholar] [CrossRef]
- Wang, M.; Hamer, R.J.; van Vliet, T.; Oudgenoeg, G. Interaction of Water Extractable Pentosans with Gluten Protein: Effect on Dough Properties and Gluten Quality. J. Cereal Sci. 2002, 36, 25–37. [Google Scholar] [CrossRef]
- Anil, M. Using of hazelnut testa as a source of dietary fiber in breadmaking. J. Food Eng. 2007, 80, 61–67. [Google Scholar] [CrossRef]
- Wang, M.; Hamer, R.J.; van Vliet, T.; Gruppen, H.; Marseille, H.; Weegels, P.L. Effect of Water Unextractable Solids on Gluten Formation and Properties: Mechanistic Considerations. J. Cereal Sci. 2003, 37, 55–64. [Google Scholar] [CrossRef]
- Wang, M.; Oudgenoeg, G.; van Vliet, T.; Hamer, R.J. Interaction of water unextractable solids with gluten protein: Effect on dough properties and gluten quality. J. Cereal Sci. 2003, 38, 95–104. [Google Scholar] [CrossRef]
- Wang, M.; Van Vliet, T.; Hamer, R.J. Evidence that pentosans and xylanase affect the re-agglomeration of the gluten network. J. Cereal Sci. 2004, 39, 341–349. [Google Scholar] [CrossRef]
- Wang, M.; van Vliet, T.; Hamer, R.J. How gluten properties are affected by pentosans. J. Cereal Sci. 2004, 39, 395–402. [Google Scholar] [CrossRef]
- Collar, C.; Santos, E.; Rosell, C.M. Assessment of the rheological profile of fibre-enriched bread doughs by response surface methodology. J. Food Eng. 2007, 78, 820–826. [Google Scholar] [CrossRef]
- Frederix, S.A.; Van Hoeymissen, K.E.; Courtin, C.M.; Delcour, J.A. Water-Extractable and Water-Unextractable Arabinoxylans Affect Gluten Agglomeration Behavior during Wheat Flour Gluten−Starch Separation. J. Agric. Food Chem. 2004, 52, 7950–7956. [Google Scholar] [CrossRef] [PubMed]
- Biliaderis, C.G.; Izydorczyk, M.S.; Rattan, O. Effect of arabinoxylans on bread-making quality of wheat flours. Food Chem. 1995, 53, 165–171. [Google Scholar] [CrossRef]
- Autio, K. Effects of cell wall components on the functionality of wheat gluten. Biotechnol. Adv. 2006, 24, 633–635. [Google Scholar] [CrossRef] [PubMed]
- Courtin, C.M.; Delcour, J.A. Relative Activity of Endoxylanases Towards Water-extractable and Water-unextractable Arabinoxylan. J. Cereal Sci. 2001, 33, 301–312. [Google Scholar] [CrossRef]
- Byrne, J. Baked Goods with Aleurone Fractions may Boost Heart Health. Available online: https://www.bakeryandsnacks.com/Article/2010/05/07/Baked-goods-with-aleurone-fractions-may-boost-heart-health (accessed on 5 April 2022).
- Shewry, P.R.; Wan, Y.; Hawkesford, M.J.; Tosi, P. Spatial distribution of functional components in the starchy endosperm of wheat grains. J. Cereal Sci. 2020, 91, 102869. [Google Scholar] [CrossRef]
- Mert, I.D. The applications of microfluidization in cereals and cereal-based products: An overview. Crit. Rev. Food Sci. Nutr. 2020, 60, 1007–1024. [Google Scholar] [CrossRef]
- Aslam, M.F.; Ellis, P.R.; Berry, S.E.; Latunde-Dada, G.O.; Sharp, P.A. Enhancing mineral bioavailability from cereals: Current strategies and future perspectives. Nutr. Bull. 2018, 43, 184–188. [Google Scholar] [CrossRef] [Green Version]
- Hemery, Y.M.; Anson, N.M.; Havenaar, R.; Haenen, G.R.M.M.; Noort, M.W.J.; Rouau, X. Dry-fractionation of wheat bran increases the bioaccessibility of phenolic acids in breads made from processed bran fractions. Food Res. Int. 2010, 43, 1429–1438. [Google Scholar] [CrossRef]
- Gan, J.; Xie, L.; Peng, G.; Xie, J.; Chen, Y.; Yu, Q. Systematic review on modification methods of dietary fiber. Food Hydrocoll. 2021, 119, 106872. [Google Scholar] [CrossRef]
- Arzami, A.N.; Ho, T.M.; Mikkonen, K.S. Valorization of cereal by-product hemicelluloses: Fractionation and purity considerations. Food Res. Int. 2021, 151, 110818. [Google Scholar] [CrossRef] [PubMed]
- Bagdi, A.; Tömösközi, S.; Nyström, L. Structural and functional characterization of oxidized feruloylated arabinoxylan from wheat. Food Hydrocoll. 2017, 63, 219–225. [Google Scholar] [CrossRef]
- Zhang, M.-Y.; Liao, A.-M.; Thakur, K.; Huang, J.-H.; Zhang, J.-G.; Wei, Z.-J. Modification of wheat bran insoluble dietary fiber with carboxymethylation, complex enzymatic hydrolysis and ultrafine comminution. Food Chem. 2019, 297, 124983. [Google Scholar] [CrossRef] [PubMed]


| Aleurone Layer * | Bran * (%, w/w) | Whole Grain (%, w/w) | ||
|---|---|---|---|---|
| Part in Tissue (%, w/w) | Total in Grain (%, w/w) | |||
| Arabinoxylan | 65 [25,36,37] | - | 60–65 [32] | 4–9 [36] |
| β-glucans | 29 [36,37] | - | 6 [36] | 0.5–2.3 [36] |
| Cellulose | 2–3 [32,33,36,37] | ≈8 [32,33] | 25–30 [32] | 2–4 [32] |
| Glucomannan | 2 [36,37] | - | - | <1 [37] |
| Pentosans | 46 [32,33] | ≈44 [32,33] | 70–80 [32] | 8–10 [32] |
| Starch | 0 [32,33] | 0 [32] | 9–25 [3] | 67–71 [32] |
| Total Dietary Fibre | 43–48 [12] | - | 40–53 [3] | 13 [1] |
| Aleurone Layer * | Bran * (mg/100 g dm) | Whole Grain (mg/100 g dm) | ||
|---|---|---|---|---|
| Part in Tissue (mg/100 g dm) | Total in Grain (%, w/w) | |||
| Calcium | 73.0 [4,10] | 53.3 [4] | 100 [32] | 30.0–70.0 [32] |
| Copper | 12.4 [4] | 67.3 [4] | 0.8–1.6 [32,54] | 0.4–1.5 [4,32,55] |
| Iron | 19.0–34.0 [4,10,54] | 78.6 [4] | 5.0–15.0 [3,32,54] | 1.8–8.0 [4,32,54] |
| Magnesium | 600 [4,10] | 86.7 [4] | 500–700 [32] | 100–200 [4,32] |
| Manganese | 8.0–13.0 [4,10,54] | 53.2 [4] | 7.2–14.4 [3,54] | 2.4–5.6 [4,54,55] |
| Sodium | - | - | 5.0–30.0 [32] | 3.0 [32] |
| Phosphorus | 1400–3170 [4,10,54] | 76.2 [4] | 900–1500 [1] | 218–792 [1] |
| Potassium | 1100 [4,10] | 67.6 [4] | 1000–1500 [32] | 350–600 [32] |
| Zinc | 12.0 [4,10] | 68.2 [4] | 5.6–50 [3,32,54] | 2.1–12 [4,32,54] |
| Total (%) | 12.0 [32,33] | ≈44.0 [32,33] | 3.39 [1] | 1.1–2.5 [1,32] |
| Aleurone Layer * | Bran * (mg/100 g dm) | Whole Grain (mg) | ||
|---|---|---|---|---|
| Part in Tissue (mg/100 g dm) | Total in Grain (%, w/w) | |||
| Thiamin (B1) | 1.6 [36,56,65] | 32 [3,49,54,66] | 0.54 [25] | 0.4–0.8 [32] |
| Riboflavin (B2) | 1.0 [36,56,65] | 37 [54,66] | 0.39–0.75 [25] | 0.1–0.2 [32] |
| Niacin (B3) | 61.3–90.2 [36,56,65] | ≈80 [3,49,54,66] | 14.0–18.0 [25] | 4.0–6.0 [32] |
| Pantothenic acid (B5) | 4.51 [36,56] | 41 [54,66] | 2.2–3.9 [25] | 1.0–2.0 [32] |
| Pyridoxin (B6) | 3.6 [36,56,65] | ≈60 [3,49,54,66] | 1.0–1.3 [25] | 0.5–1.0 [32] |
| Folate (B9) | 0.2–0.8 [10] | - | 0.079–0.2 [25] | 0.035–0.055 [32] |
| Vitamin E | 1.2–2.0 [10] | - | 1.4 [25] | 2.0–6.0 [32] |
| Aleurone Layer * | Bran * | Whole Grain | ||
|---|---|---|---|---|
| Part in Tissue | Total in Grain | |||
| Alkylresorcinols (mg/100 g) | ≈3.0 [65,67,68] | 20–400 [67] | 220–400 [1,3] | 42–70 [1,50,65] |
| Phytic acid (g/100 g) | 8.4–15.6 [50,65,68] | 2.2–5.2 [3] | 1.2–1.4 [50,65] | 1.18–1.38 [50,65] |
| Lutein (µg/100 g) | 7.2–42.5 [63,68] | - | 97–140 [3] | 81.9 [63] |
| Zeaxanthin (µg/100 g) | 21.2–77.6 [63,68] | - | 25–219 [1] | 9.0–43.8 [1,63] |
| Choline (mg/100 g) | 1260 [54] | - | 112 [1] | 112 [1] |
| Aleurone-Rich Flour Used | Enrichment Level (%, w/w) | Basic Flour Type and Protein Content (%) | Bread Formulation Process | Specificities | Reference | |
|---|---|---|---|---|---|---|
| Table 7, 1a/1b | 20 | White flour | 11–13 | Straight dough | Addition of vital wheat gluten, high fructose corn syrup, and dough conditioner | [12,128] |
| Sponge dough | Addition of vital wheat gluten, high fructose corn syrup, ascorbic acid, dough conditioners, mono- and diglycerides | |||||
| Table 7, 1a/1b | 40 and 75 | Wheat flour | 11.86 | ICC Standard Method 131 | - | [13,15] |
| Table 7, 1a/1b | 25 and 50 | Wheat flour | 12.9 | ICC Standard Method 131 | - | [16,17] |
| Table 7, 4 | 10, 20, 30 and 40 | Wheat flour | 14.16 | - | - | [131] |
| Table 7, 1a/1b | 15, 40, 75 and 100 | Conventional bread wheat flour | 15.24 | Sourdough (MSZ method, 1989) | - | [136] |
| 54.11% purity | 18 | Wheat flour | - | GB/T 358969-2018 method | Hemicellulase addition (0–60 mg/kg of flour) | [140] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebert, L.; Buche, F.; Sorin, A.; Aussenac, T. The Wheat Aleurone Layer: Optimisation of Its Benefits and Application to Bakery Products. Foods 2022, 11, 3552. https://doi.org/10.3390/foods11223552
Lebert L, Buche F, Sorin A, Aussenac T. The Wheat Aleurone Layer: Optimisation of Its Benefits and Application to Bakery Products. Foods. 2022; 11(22):3552. https://doi.org/10.3390/foods11223552
Chicago/Turabian StyleLebert, Lucie, François Buche, Arnaud Sorin, and Thierry Aussenac. 2022. "The Wheat Aleurone Layer: Optimisation of Its Benefits and Application to Bakery Products" Foods 11, no. 22: 3552. https://doi.org/10.3390/foods11223552