
Storage Drives Alterations of Proteomic and Protein Structural Properties in Rice (Oryza sativa L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Regents, Material, and Storage Condition
2.2. Structural Characteristics
2.2.1. Extraction of Rice Protein
2.2.2. Surface Hydrophobicity (H0)
2.2.3. Free Sulfhydryl and Disulfide Bond
2.2.4. FTIR
2.3. Structural Characteristics
2.3.1. Isolation of and Protein Extraction
2.3.2. Digestion and TMT Labeling
2.3.3. Separation of Peptide Fractions
2.3.4. LC-MS/MS Analysis
2.3.5. Identification and Quantitation of Protein
2.3.6. Differentially Expressed Proteins and Functional Analysis of Proteins
2.4. Statistical Analysis
3. Results and Discussion
3.1. Structural Properties
3.1.1. Sulfhydryl and Disulfide Bonds Content
3.1.2. Surface Hydrophobicity H0
3.1.3. FTIR
3.2. TMT-Labeled Proteomics
3.2.1. Protein Identification in Both Rice Varieties
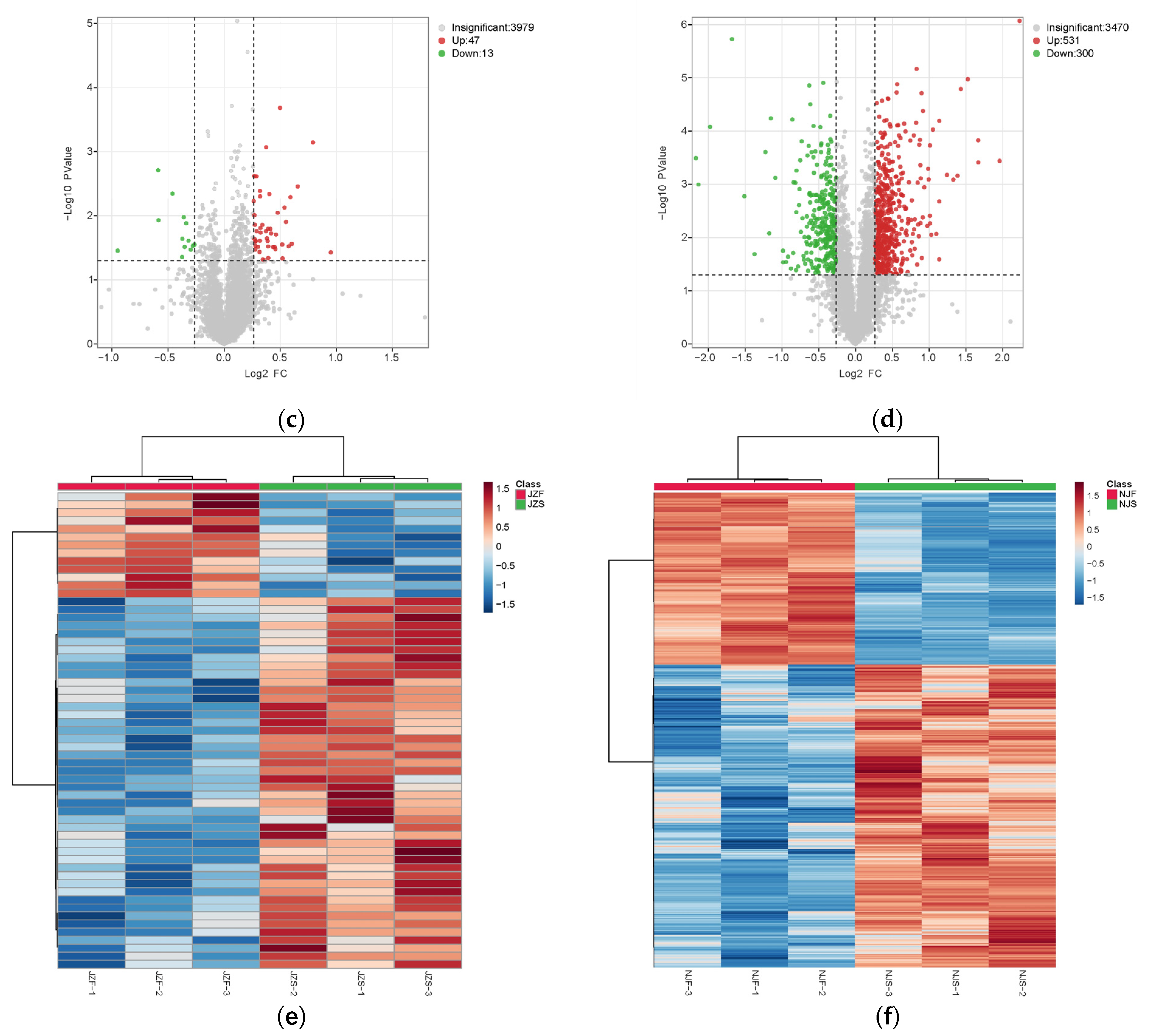
3.2.2. Identification of DEPs in Both Rice Varieties
3.2.3. GO Analysis
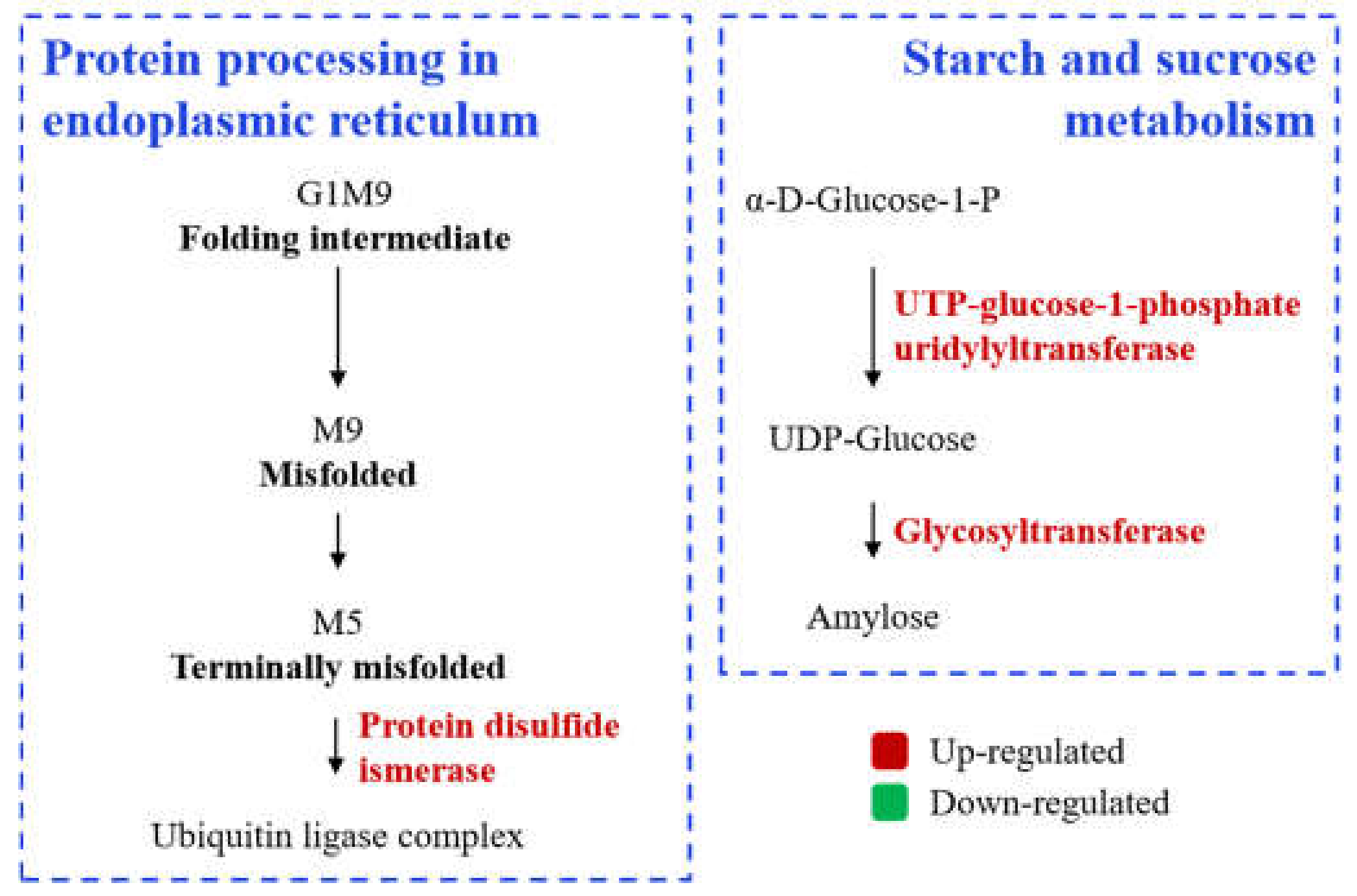
3.2.4. Comparative KEGG Pathway Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, Z.; Robards, K.; Helliwell, S.; Blanchard, C. Ageing of Stored Rice: Changes in Chemical and Physical Attributes. J. Cereal Sci. 2002, 35, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Saikrishna, A.; Dutta, S.; Subramanian, V.; Moses, J.A.; Anandharamakrishnan, C. Ageing of rice: A review. J. Cereal Sci. 2018, 81, 161–170. [Google Scholar] [CrossRef]
- Xu, J.; Liu, K.; Zhang, C. Electronic nose for volatile organic compounds analysis in rice aging. Trends Food Sci. Technol. 2021, 109, 83–93. [Google Scholar] [CrossRef]
- Chen, Z.; Li, P.; Du, Y.; Jiang, Y.; Cai, M.; Cao, C. Dry cultivation and cultivar affect starch synthesis and traits to define rice grain quality in various panicle parts. Carbohydr. Polym. 2021, 269, 118336. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Zhao, L.; Lin, L.; Wang, J.; Liu, Q.; Wei, C. Changes in kernel morphology and starch properties of high-amylose brown rice during the cooking process. Food Hydrocoll. 2017, 66, 227–236. [Google Scholar] [CrossRef]
- Ong, M.H.; Blanshard, J.M.V. Texture determinants of cooked, parboiled rice. II: Physicochemical properties and leaching behaviour of rice. J. Cereal Sci. 1995, 21, 261–269. [Google Scholar] [CrossRef]
- Zhao, Q.; Lin, J.; Wang, C.; Yousaf, L.; Xue, Y.; Shen, Q. Protein structural properties and proteomic analysis of rice during storage at different temperatures. Food Chem. 2021, 361, 130028. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, X.; Sun, H.; Cao, X.; Liu, J.; Zhang, Z. Relationship of grain protein content with cooking and eating quality as affected by nitrogen fertilizer at late growth stage for different types of rice varieties. Chin. J. Rice Sci. 2019, 33, 541–552. [Google Scholar] [CrossRef]
- Chen, Y.; Jiang, W.; Jiang, Z.; Chen, X.; Cao, J.; Dong, W.; Dai, B. Changes in Physicochemical, Structural, and Sensory Properties of Irradiated Brown Japonica Rice during Storage. J. Agric. Food Chem. 2015, 63, 4361–4369. [Google Scholar] [CrossRef]
- Guo, Y.; Cai, W.; Tu, K.; Tu, S.; Wang, S.; Zhu, X.; Zhang, W. Infrared and Raman Spectroscopic Characterization of Structural Changes in Albumin, Globulin, Glutelin, and Prolamin during Rice Aging. J. Agric. Food Chem. 2012, 61, 185–192. [Google Scholar] [CrossRef]
- Itze-Mayrhofer, C.; Brem, G. Quantitative proteomic strategies to study reproduction in farm animals: Female reproductive fluids. J. Proteom. 2020, 225, 103884. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, J.; Wang, A.; Wang, R.; Sun, H.; Strappe, P.; Zhou, Z. Physiological and proteomic analyses provide insights into the rice yellowing. J. Cereal Sci. 2020, 95, 103048. [Google Scholar] [CrossRef]
- Xiao, R.; Li, L.; Ma, Y. A label-free proteomic approach differentiates between conventional and organic rice. J. Food Compos. Anal. 2019, 80, 51–61. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, D.; Zhao, L.; Liu, J.; Shang, B.; Yang, W.; Duan, X.; Sun, H. Metabolomic Analysis Reveals Insights into Deterioration of Rice Quality during Storage. Foods 2022, 11, 1729. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Duan, X.; Shang, B.; Hong, Y.; Sun, H. Analysis of lipidomics profile of rice and changes during storage by UPLC-Q-extractive orbitrap mass spectrometry. Food Res. Int. 2021, 142, 110214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhao, L.; Wang, W.; Wang, Q.; Liu, J.; Wang, Y.; Liu, H.; Shang, B.; Duan, X.; Sun, H. Lipidomics reveals the changes in non-starch and starch lipids of rice (Oryza sativa L.) during storage. J. Food Compos. Anal. 2022, 105, 104205. [Google Scholar] [CrossRef]
- Wu, X.; Li, F.; Wu, W. Effects of oxidative modification by 13-hydroperoxyoctadecadienoic acid on the structure and functional properties of rice protein. Food Res. Int. 2020, 132, 109096. [Google Scholar] [CrossRef]
- Kachuk, C.; Stephen, K.; Doucette, A. Comparison of sodium dodecyl sulfate depletion techniques for proteome analysis by mass spectrometry. J. Chromatogr. A 2015, 1418, 158–166. [Google Scholar] [CrossRef]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Schöneich, C. Thiyl radicals and induction of protein degradation. Free Radic. Res. 2016, 50, 143–149. [Google Scholar] [CrossRef]
- Sun, W.; Zhou, F.; Sun, D.-W.; Zhao, M. Effect of Oxidation on the Emulsifying Properties of Myofibrillar Proteins. Food Bioprocess Technol. 2013, 6, 1703–1712. [Google Scholar] [CrossRef]
- Wang, C.; Wang, J.; Zhu, D.; Hu, S.; Kang, Z.; Ma, H. Effect of dynamic ultra-high pressure homogenization on the structure and functional properties of whey protein. J. Food Sci. Technol. 2020, 57, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Azizi, R.; Capuano, E.; Nasirpour, A.; Pellegrini, N.; Golmakani, M.-T.; Hosseini, S.M.H.; Farahnaky, A. Varietal differences in the effect of rice ageing on starch digestion. Food Hydrocoll. 2019, 95, 358–366. [Google Scholar] [CrossRef]
- Hamaker, B.R.; Griffin, V.K. Effect of disulfide bond-containing protein on rice starch gelatinization and pasting. Cereal Chem. 1993, 70, 377–380. [Google Scholar] [CrossRef]
- Ren, J.; Li, S. Effects of thermal treatment on structure and surface hydrophobicity of sunflower seed protein isolate. China Oils Fats 2016, 41, 24–27. [Google Scholar]
- Zhou, X.; Liu, Y.; Li, X.; Yu, J. Effects of protein-glutaminase on the molecular structure and functional properties of rice glutelin. J. Chin. Inst. Food Sci. Technol. 2010, 10, 98–105. [Google Scholar] [CrossRef]
- Zhang, B.; Li, H.; Li, F.; Zhou, Q.; Wu, X.; Wu, W. Effects of rice bran phenolics on the structure of rice bran protein under different degrees of rancidity. LWT 2022, 167, 113812. [Google Scholar] [CrossRef]
- Hou, F.; Ding, W.; Qu, W.; Oladejo, A.O.; Xiong, F.; Zhang, W.; He, R.; Ma, H. Alkali solution extraction of rice residue protein isolates: Influence of alkali concentration on protein functional, structural properties and lysinoalanine formation. Food Chem. 2017, 218, 207–215. [Google Scholar] [CrossRef]
- Singh, T.P.; Sogi, D.S. Comparative study of structural and functional characterization of bran protein concentrates from superfine, fine and coarse rice cultivars. Int. J. Biol. Macromol. 2018, 111, 281–288. [Google Scholar] [CrossRef]
- Gao, J.; Fu, H.; Zhou, X.; Chen, Z.; Luo, Y.; Cui, B.; Chen, G.; Liu, J. Comparative proteomic analysis of seed embryo proteins associated with seed storability in rice (Oryza sativa L) during natural aging. Plant Physiol. Biochem. 2016, 103, 31–44. [Google Scholar] [CrossRef]
- Hu, M. Study on the Quality Deterioration Mechanism of Tibetan Hulless Barley under Different Storage Conditions; Central South University of Forestry and Technology: Changsha, China, 2017. [Google Scholar]
- Onda, Y.; Kobori, Y. Differential activity of rice protein disulfide isomerase family members for disulfide bond formation and reduction. FEBS Open Bio 2014, 4, 730–734. [Google Scholar] [CrossRef]
- Cao, Z. Effects of High Temperature on Rice (Oryza sativa L.) Floral Injury and Grain Quality in Relation to Carbon and Nitrogen Metabolism; Zhejiang University: Zhejiang, China, 2014. [Google Scholar]
- Houston, N.L.; Fan, C.; Xiang, Q.-Y.; Schulze, J.-M.; Jung, R.; Boston, R.S. Phylogenetic Analyses Identify 10 Classes of the Protein Disulfide Isomerase Family in Plants, Including Single-Domain Protein Disulfide Isomerase-Related Proteins. Plant Physiol. 2005, 137, 762–778. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; He, D.; Teng, H.; Chen, L.; Song, H.; Huang, Q. Physiological and proteomic analyses of coix seed aging during storage. Food Chem. 2018, 260, 82–89. [Google Scholar] [CrossRef]
- Cao, Q.; Lv, W.; Jiang, H.; Chen, X.; Wang, X.; Wang, Y. Genome-wide identification of glutathione S-transferase gene family members in tea plant (Camellia sinensis) and their response to environmental stress. Int. J. Biol. Macromol. 2022, 205, 749–760. [Google Scholar] [CrossRef]
- Bhattacharya, K.R. 5—Ageing of Rice. In Rice Quality: A Guide to Rice Properties and Analysis; Bhattacharya, K.R., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 116–163. [Google Scholar]
- Liu, J.; Hu, Z.; Liu, D.; Zheng, A.; Ma, Q. Glutathione metabolism-mediated ferroptosis reduces water-holding capacity in beef during cold storage. Food Chem. 2023, 398, 133903. [Google Scholar] [CrossRef]
- Ajadi, A.A.; Cisse, A.; Ahmad, S.; Yifeng, W.; Yazhou, S.; Shufan, L.; Xixi, L.; Bello, B.K.; Tajo, S.M.; Xiaohong, T.; et al. Protein Phosphorylation and Phosphoproteome: An Overview of Rice. Rice Sci. 2020, 27, 184–200. [Google Scholar] [CrossRef]
- Li, X.; Deng, X.; Guo, X.; Wei, Y.; Zhao, Y.; Guo, X.; Zhu, X.; Zhang, J.; Hu, L. Two-dimensional gel analysis to investigate the effect of hydroxyl radical oxidation on freshness indicator protein of Coregonus peled during 4 °C storage. LWT 2022, 158, 113147. [Google Scholar] [CrossRef]
- Huang, Y.-C.; Lai, H.-M. Characteristics of the starch fine structure and pasting properties of waxy rice during storage. Food Chem. 2014, 152, 432–439. [Google Scholar] [CrossRef]
- Wu, P.; Li, C.; Bai, Y.; Yu, S.; Zhang, X. A starch molecular basis for aging-induced changes in pasting and textural properties of waxy rice. Food Chem. 2019, 284, 270–278. [Google Scholar] [CrossRef]
- Der Agopian, R.G.; Peroni-Okita, F.H.G.; Soares, C.A.; Mainardi, J.A.; do Nascimento, J.R.O.; Cordenunsi, B.R.; Lajolo, F.M.; Purgatto, E. Low temperature induced changes in activity and protein levels of the enzymes associated to conversion of starch to sucrose in banana fruit. Postharvest Biol. Technol. 2011, 62, 133–140. [Google Scholar] [CrossRef]
- Huan, C.; Du, X.; Wang, L.; Kebbeh, M.; Li, H.; Yang, X.; Shen, S.; Zheng, X. Transcriptome analysis reveals the metabolisms of starch degradation and ethanol fermentation involved in alcoholic off-flavour development in kiwifruit during ambient storage. Postharvest Biol. Technol. 2021, 180, 111621. [Google Scholar] [CrossRef]
- Amagliani, L.; O’Regan, J.; Kelly, A.L.; O’Mahony, J.A. The composition, extraction, functionality and applications of rice proteins: A review. Trends Food Sci. Technol. 2017, 64, 1–12. [Google Scholar] [CrossRef]
- Tang, S.; Chen, W.; Liu, W.; Zhou, Q.; Zhang, H.; Wang, S.; Ding, Y. Open-field warming regulates the morphological structure, protein synthesis of grain and affects the appearance quality of rice. J. Cereal Sci. 2018, 84, 20–29. [Google Scholar] [CrossRef]
- Zhao, Q.; Xue, Y.; Shen, Q. Changes in the major aroma-active compounds and taste components of Jasmine rice during storage. Food Res. Int. 2020, 133, 109160. [Google Scholar] [CrossRef]
- Paulo, C.C.; Carlos, H.P.F.; Jean, C.H. Adjustment of mathematical models and quality of soybean grains in the drying with high temperatures. Rev. Bras. Eng. Agrícola Ambient. 2016, 20, 385–392. [Google Scholar]
- Müller, A.; Nunes, M.T.; Maldaner, V.; Coradi, P.C.; de Moraes, R.S.; Martens, S.; Leal, A.F.; Pereira, V.F.; Marin, C.K. Rice Drying, Storage and Processing: Effects of Post-Harvest Operations on Grain Quality. Rice Sci. 2022, 29, 16–30. [Google Scholar] [CrossRef]
- Qiu, S.; Kawamura, S.; Fujikawa, S.; Doi, T. Long-term storability of rough rice and brown rice under different storage conditions. Eng. Agric. Environ. Food 2014, 7, 40–45. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | α-Helix/% | β-Sheet/% | β-Turn/% | Random Coil/% |
|---|---|---|---|---|
| NJF | 20.6 ± 0.2 | 37.3 ± 0.2 | 11.2 ± 0.2 | 30.8 ± 0.3 |
| NJS | 17.8 ± 0.2 * | 21.6 ± 0.5 * | 13.1 ± 0.2 * | 47.5 ± 0.4 * |
| JZF | 32.4 ± 0.7 | 17.1 ± 0.5 | 12.9 ± 0.7 | 37.6 ± 0.5 |
| JZS | 30.7 ± 0.5 * | 19.9 ± 0.8 * | 11.6 ± 0.1 * | 37.8 ± 0.3 |
| No. | Protein ID | NJS vs. NJF | JZS vs. JZF | KEGG Pathway Name | ||||
|---|---|---|---|---|---|---|---|---|
| FC | p-Value | Regulated | FC | p-Value | Regulated | |||
| 1 | Q67VS7 | 2.013 | 0.004 | up | 1.313 | 0.046 | up | |
| 2 | C7J9I2 | 1.653 | 0.001 | up | 1.518 | 0.028 | up | / |
| 3 | Q0J8Q8 | 1.502 | 0.000 | up | 1.322 | 0.005 | up | / |
| 4 | Q6AUV3 | 1.467 | 0.008 | up | 1.249 | 0.005 | up | / |
| 5 | Q6ATY6 | 1.409 | 0.011 | up | 1.248 | 0.025 | up | Purine metabolism Pyrimidine metabolism Metabolic pathways RNA polymerase |
| 6 | B9EZ14 | 1.387 | 0.003 | up | 1.266 | 0.014 | up | / |
| 7 | Q7XQF0 | 1.376 | 0.007 | up | 1.252 | 0.018 | up | Ubiquitin-mediated proteolysis |
| 8 | B9F2Z5 | 1.321 | 0.005 | up | 1.212 | 0.034 | up | / |
| 9 | B9G0M2 | 1.282 | 0.001 | up | 1.212 | 0.025 | up | Ubiquitin-mediated proteolysis |
| 10 | Q6ZJ18 | 1.248 | 0.001 | up | 1.233 | 0.031 | up | / |
| 11 | Q2QPW3 | 0.809 | 0.001 | down | 0.802 | 0.025 | down | / |
| 12 | B9G505 | 0.649 | 0.001 | down | 0.771 | 0.044 | down | Protein export |
| 13 | B9ESY3 | 0.776 | 0.000 | down | 1.435 | 0.046 | up | Spliceosome |
| 14 | A0A0P0Y2R7 | 2.199 | 0.000 | up | 0.667 | 0.012 | down | |
| 15 | Q8S7K1 | 1.302 | 0.000 | up | 0.791 | 0.013 | down | / |
| No. | Protein ID | Description | Gene | NJS vs. NJF | JZS vs. JZF | ||||
|---|---|---|---|---|---|---|---|---|---|
| FC | p-Value | Regulated | FC | p-Value | Regulated | ||||
| Redox Homeostasis/Stress-Related | |||||||||
| 1 | Q655N4 | Putative 70 kDa heat-shock protein | OsJ_22372 | 1.344 | 0.006 | up | / | / | / |
| 2 | Q6K7E9 | 18.6 kDa class III heat shock protein | HSP18.6 | 1.322 | 0.006 | up | 1.110 | 0.102 | / |
| 3 | Q6AUW3 | 22.3 kDa class VI heat shock protein | HSP22.3 | 1.228 | 0.030 | up | 1.177 | 0.334 | / |
| 4 | Q943E6 | 16.9 kDa class I heat shock protein 2 | HSP16.9B | 1.221 | 0.047 | up | 0.856 | 0.292 | / |
| 5 | Q7XRB5 | Protein disulfide isomerase-like 1-2 | PDIL1-2 | 2.197 | 0.026 | up | 1.038 | 0.787 | / |
| 6 | K4FHN8 | Protein disulfide-isomerase | Unknown | 1.030 | 0.604 | / | 1.732 | 0.001 | up |
| 7 | Q945W9 | Glutathione S-transferase GSTU6 | LOC_Os10g38590 | 1.396 | 0.003 | up | 1.037 | 0.401 | / |
| Oxidative phosphorylation in mitochondria | |||||||||
| 8 | Q0JKB4 | F-type H+-transporting ATPase subunit β | Os01g0685800 | 0.659 | 0.019 | down | 0.856 | 0.018 | / |
| 9 | Q8GTK7 | F-type H+-transporting ATPase subunit δ | Os07g0495200 | 0.673 | 0.000 | down | 0.972 | 0.735 | / |
| 10 | Q7XXS0 | F-type H+-transporting ATPase subunit d | Os08g0478200 | 0.759 | 0.010 | down | 1.062 | 0.585 | / |
| 11 | Q8RU25 | F-type H+-transporting ATPase subunit γ | OSJNAa0053D03.3 | 0.831 | 0.045 | down | 0.874 | 0.083 | / |
| 12 | B9F0H4 | V-type proton ATPase proteolipid subunit | Os02g0550100 | 0.798 | 0.020 | down | / | / | / |
| 13 | Q2F948 | NADH-ubiquinone oxidoreductase chain 5 | nad5 | 0.668 | 0.007 | down | / | / | / |
| 14 | Q8HCQ0 | NADH-ubiquinone oxidoreductase chain 3 | nad3 | 0.780 | 0.039 | down | 1.158 | 0.300 | / |
| 15 | Q8HCM1 | NADH dehydrogenase subunit 2 | nad2 | 0.807 | 0.019 | down | 1.052 | 0.524 | / |
| 16 | Q6K9W2 | NADH dehydrogenase [ubiquinone] 1 α subcomplex subunit 9 | Os02g0816800 | 0.667 | 0.002 | down | 0.992 | 0.729 | / |
| 17 | A0A5S6RA53 | NADH dehydrogenase [ubiquinone] 1 α subcomplex subunit 12 | Os10g0579300 | 0.725 | 0.000 | down | 0.974 | 0.643 | / |
| 18 | Q8GS72 | NADH dehydrogenase [ubiquinone] 1 α subcomplex subunit 6 | Os07g0640100 | 0.774 | 0.002 | down | 1.028 | 0.692 | / |
| 19 | A0A5S6RAL2 | NADH dehydrogenase [ubiquinone] 1 α subcomplex subunit 6 | Os04g0310500 | 0.783 | 0.002 | down | 1.004 | 0.939 | / |
| 20 | Q0J7V0 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 6 | P0577B11.122 | 0.748 | 0.006 | down | 0.982 | 0.658 | / |
| 21 | Q6ZJ19 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 5-B | Os08g0556600 | 0.756 | 0.014 | down | 1.126 | 0.086 | / |
| Fatty acid metabolism | |||||||||
| 22 | B7F6I0 | 3-oxoacyl-[acyl-carrier-protein] synthase | Os04g0445700 | 1.563 | 0.002 | up | 1.078 | 0.161 | / |
| 23 | Q6H5J0 | Enoyl-[acyl-carrier-protein] reductase [NADH] 2 | Os09g0277800 | 1.492 | 0.000 | up | 1.012 | 0.680 | / |
| 24 | A3CDM0 | Acyl-[acyl-carrier-protein] hydrolase | Os11g0659500 | 1.247 | 0.010 | up | 1.041 | 0.147 | / |
| 25 | A3C125 | Acyl carrier protein | OsJ_30173 | 0.770 | 0.022 | down | 1.096 | 0.300 | / |
| 26 | Q6ZJI9 | Acyl carrier protein | Os08g0549300 | 0.642 | 0.010 | down | 1.064 | 0.541 | / |
| Energy/carbohydrate metabolism | |||||||||
| 27 | A0A0P0UZU8 | ATP-dependent 6-phosphofructokinase | PFK | 1.289 | 0.009 | up | 1.063 | 0.088 | / |
| 28 | A0A0P0W2E0 | Glycosyltransferase | Os03g0693600 | 1.768 | 0.000 | up | 1.135 | 0.178 | / |
| 29 | Q40677 | Fructose-bisphosphate aldolase | ALDO | 1.495 | 0.000 | up | 0.934 | 0.086 | / |
| 30 | Q7XSK0 | β-glucosidase 18 | BGLU18 | 1.457 | 0.001 | up | 0.934 | 0.305 | / |
| 31 | B7FA07 | Phosphopyruvate hydratase | Os09g0375000 | 1.340 | 0.001 | up | 0.954 | 0.387 | / |
| 32 | Q8S7N6 | Pyruvate kinase | LOC_Os10g42100 | 1.332 | 0.017 | up | 0.925 | 0.717 | / |
| 33 | Q0ILJ3 | Sucrose transport protein SUT2 | SUT2 | 1.300 | 0.009 | up | 1.032 | 0.402 | / |
| 34 | Q8GTK0 | Starch synthase | GBSSII | 1.297 | 0.021 | up | / | / | / |
| 35 | A0A0P0VKS3 | Aldose 1-epimerase | Os02g0575800 | 1.200 | 0.003 | up | 0.972 | 0.589 | / |
| 36 | Q0E4I5 | UTP-glucose-1-phosphate uridylyltransferase | Os02g0117700 | / | / | / | 1.378 | 0.020 | up |
| 37 | Q6ESW3/ A3A4Z8 | Glycosyltransferase | Os02g0242900 | / | / | / | 1.412 | 0.000 | up |
| 38 | Q10CK4 | α-1,4 glucan phosphorylase | LOC_Os03g55090 | 0.470 | 0.001 | down | 0.812 | 0.458 | / |
| Gluetin | |||||||||
| 39 | Q84X94 | Glutelin | GluB-5 | 1.473 | 0.011 | up | 0.971 | 0.863 | / |
| 40 | A0A0N7KF02/ Q6ESW6 | Glutelin | GluB-5-like, Os02g0242600 | 1.076 | 0.378 | / | 1.432 | 0.028 | up |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Zhang, D.; Liu, J.; Shang, B.; Duan, X.; Sun, H. Storage Drives Alterations of Proteomic and Protein Structural Properties in Rice (Oryza sativa L.). Foods 2022, 11, 3541. https://doi.org/10.3390/foods11213541
Wang Q, Zhang D, Liu J, Shang B, Duan X, Sun H. Storage Drives Alterations of Proteomic and Protein Structural Properties in Rice (Oryza sativa L.). Foods. 2022; 11(21):3541. https://doi.org/10.3390/foods11213541
Chicago/Turabian StyleWang, Qian, Dong Zhang, Jianlei Liu, Bo Shang, Xiaoliang Duan, and Hui Sun. 2022. "Storage Drives Alterations of Proteomic and Protein Structural Properties in Rice (Oryza sativa L.)" Foods 11, no. 21: 3541. https://doi.org/10.3390/foods11213541