
Changes in Caprine Milk Fat Globule Membrane Proteins after Heat Treatment Using a Label-Free Proteomics Technique

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Treatment and Reagents
2.2. MFGM Protein Extraction
2.3. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.4. Protein Digestion
2.5. Liquid Chromatography-Tandem Mass Spectrometry Analysis (LC-MS/MS)
2.6. Data Analysis
3. Results
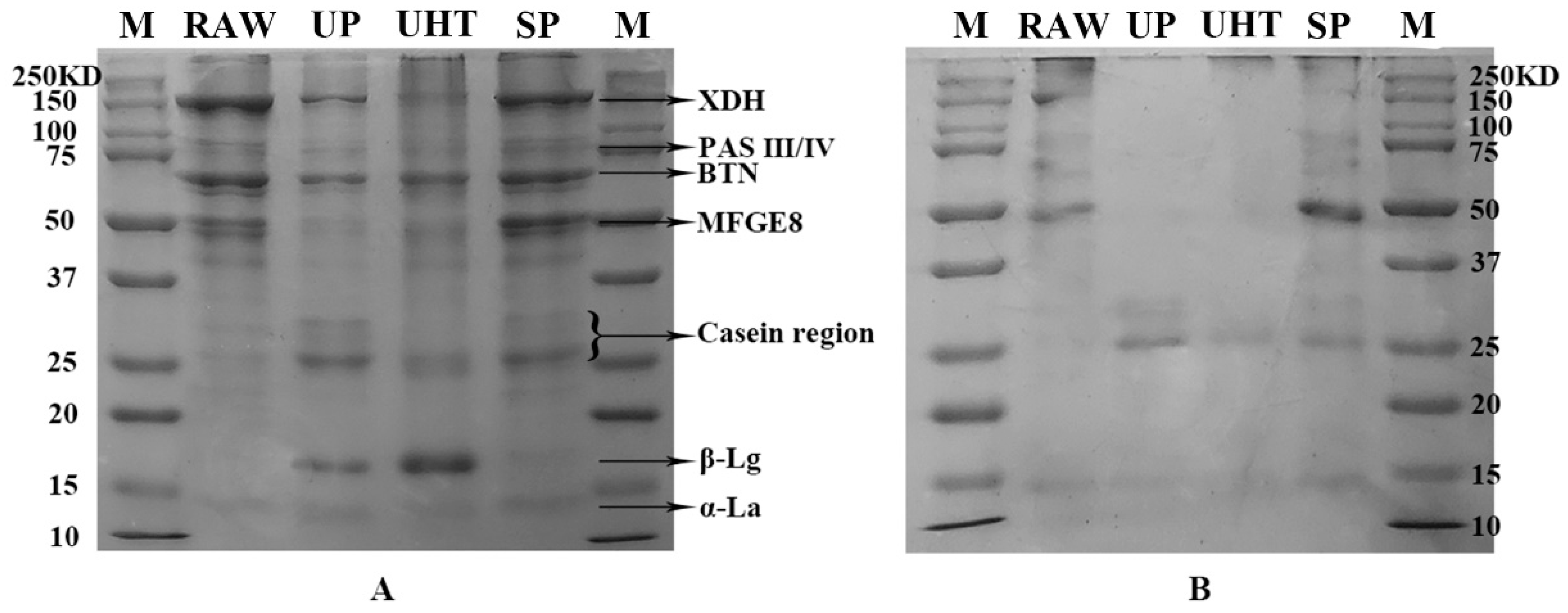
3.1. SDS-PAGE of MFGM Proteins
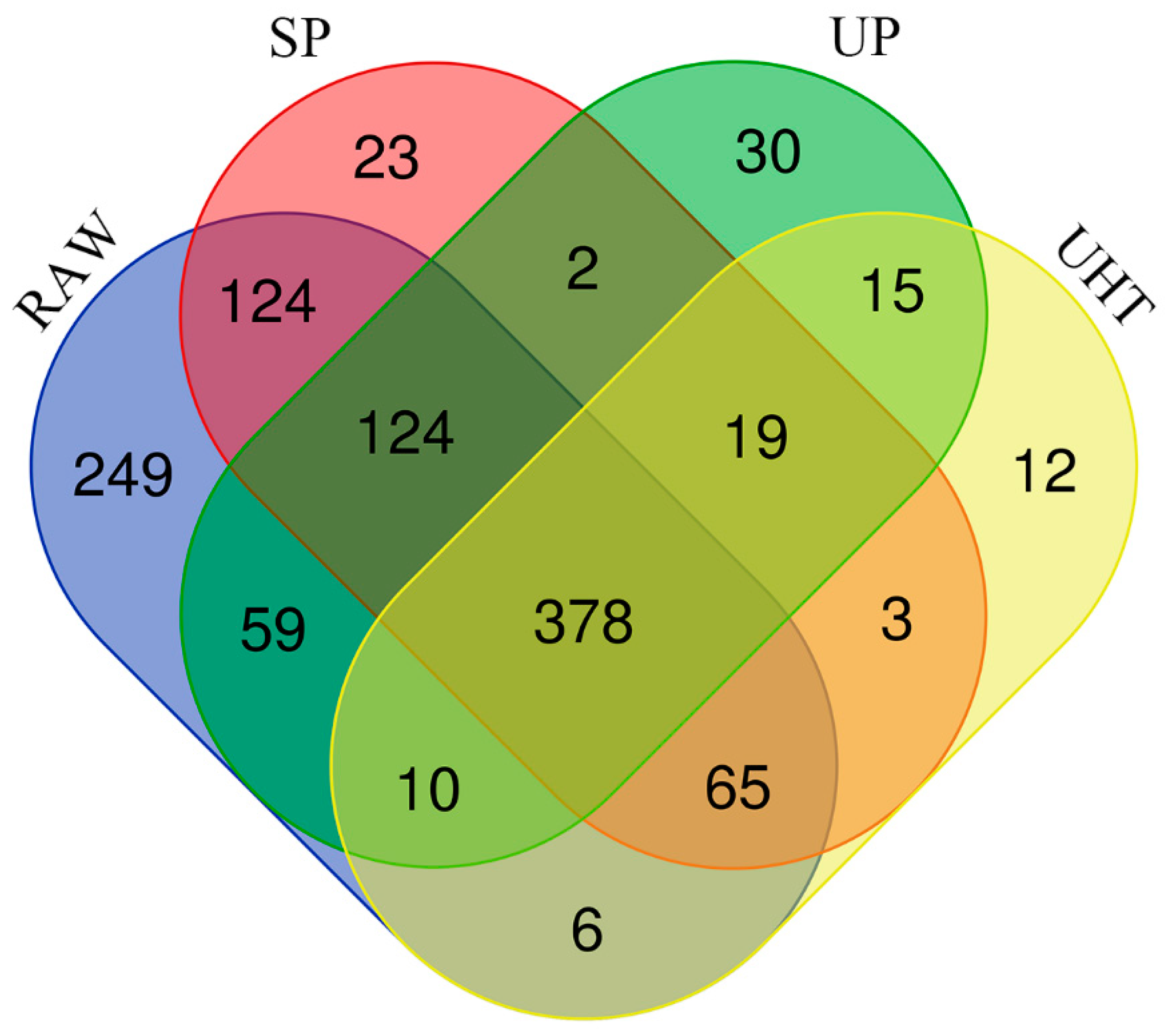
3.2. Identification and Quantification of MFGM Proteins
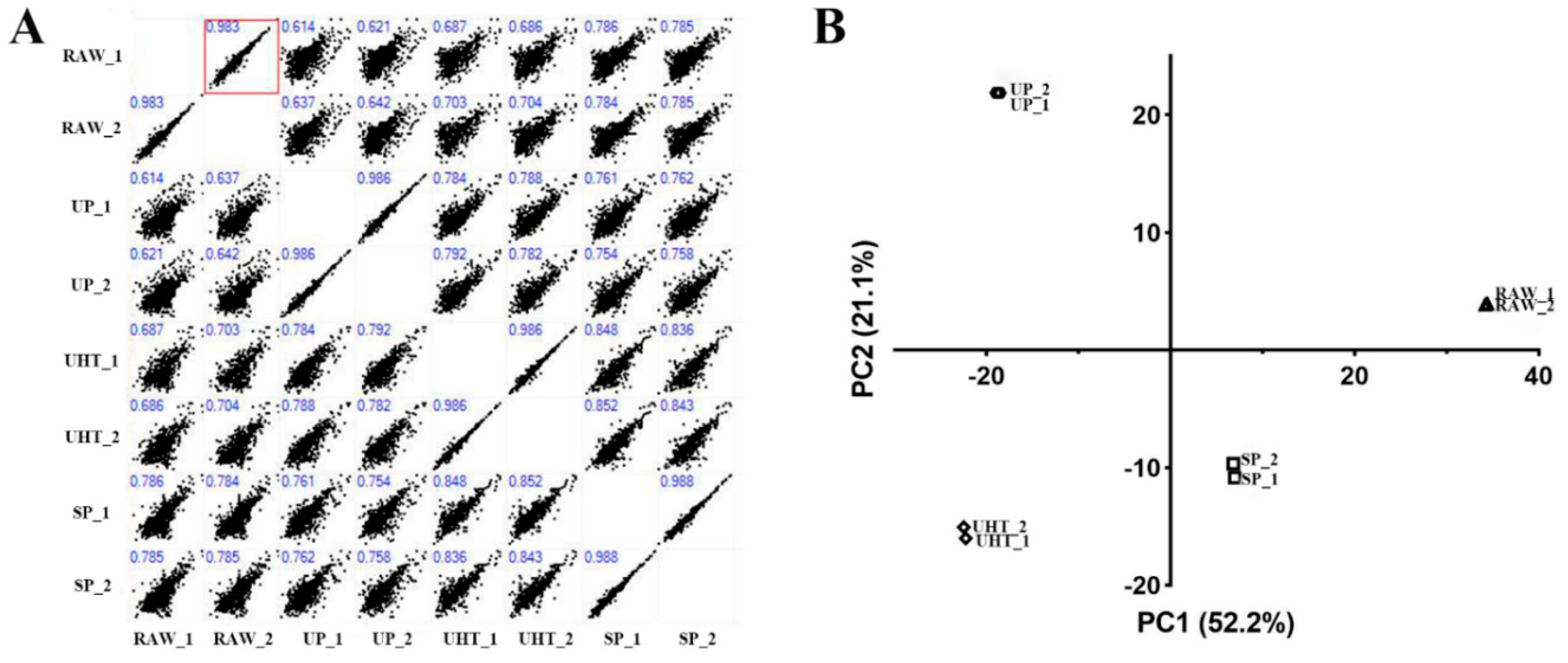
3.3. Correlation and PCA Analysis of MFGM Proteins from Different Heat Treatment Groups
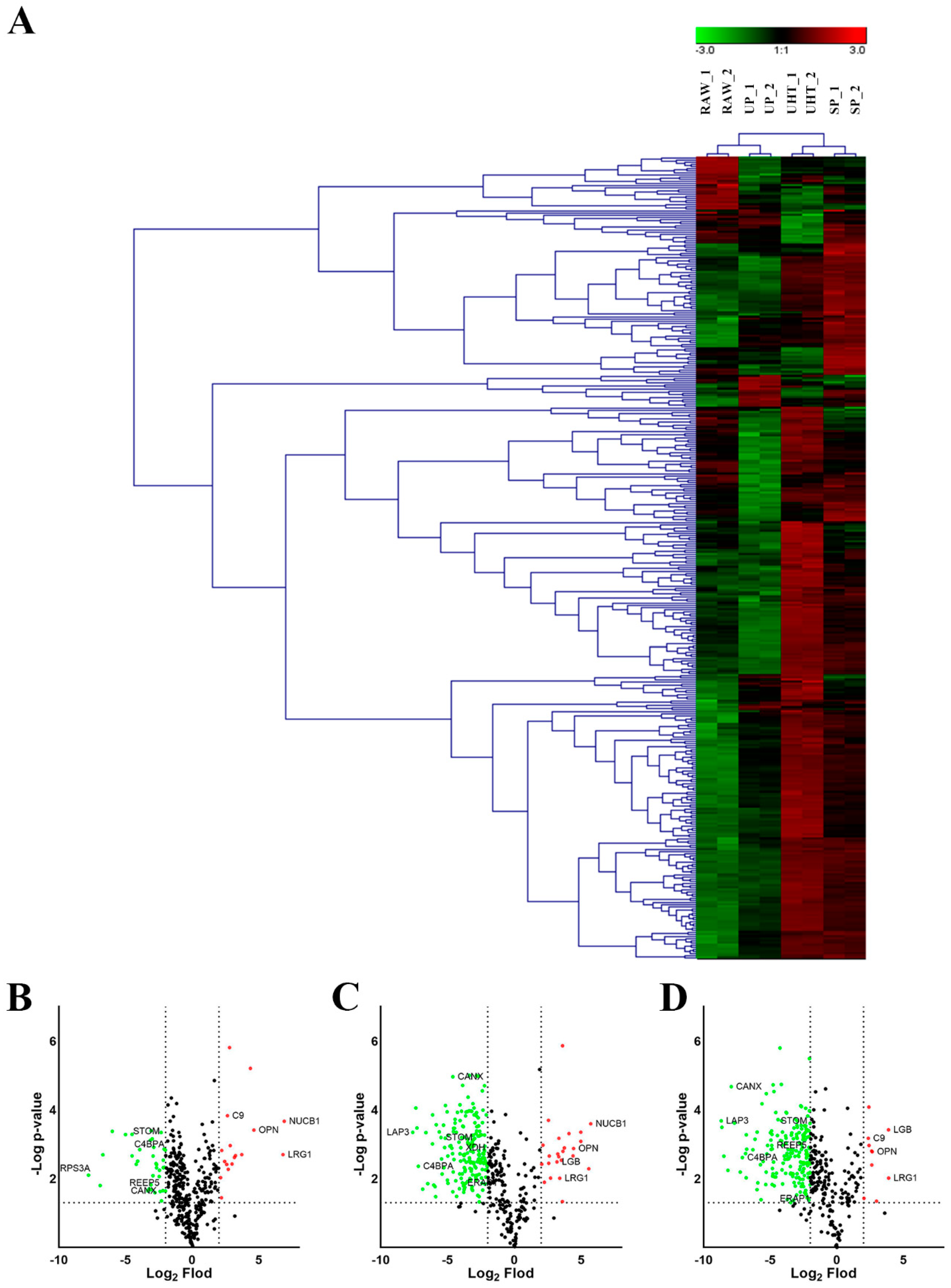
3.4. Significant Differences in the Changes MFGM Proteins among Different Heat Treatment
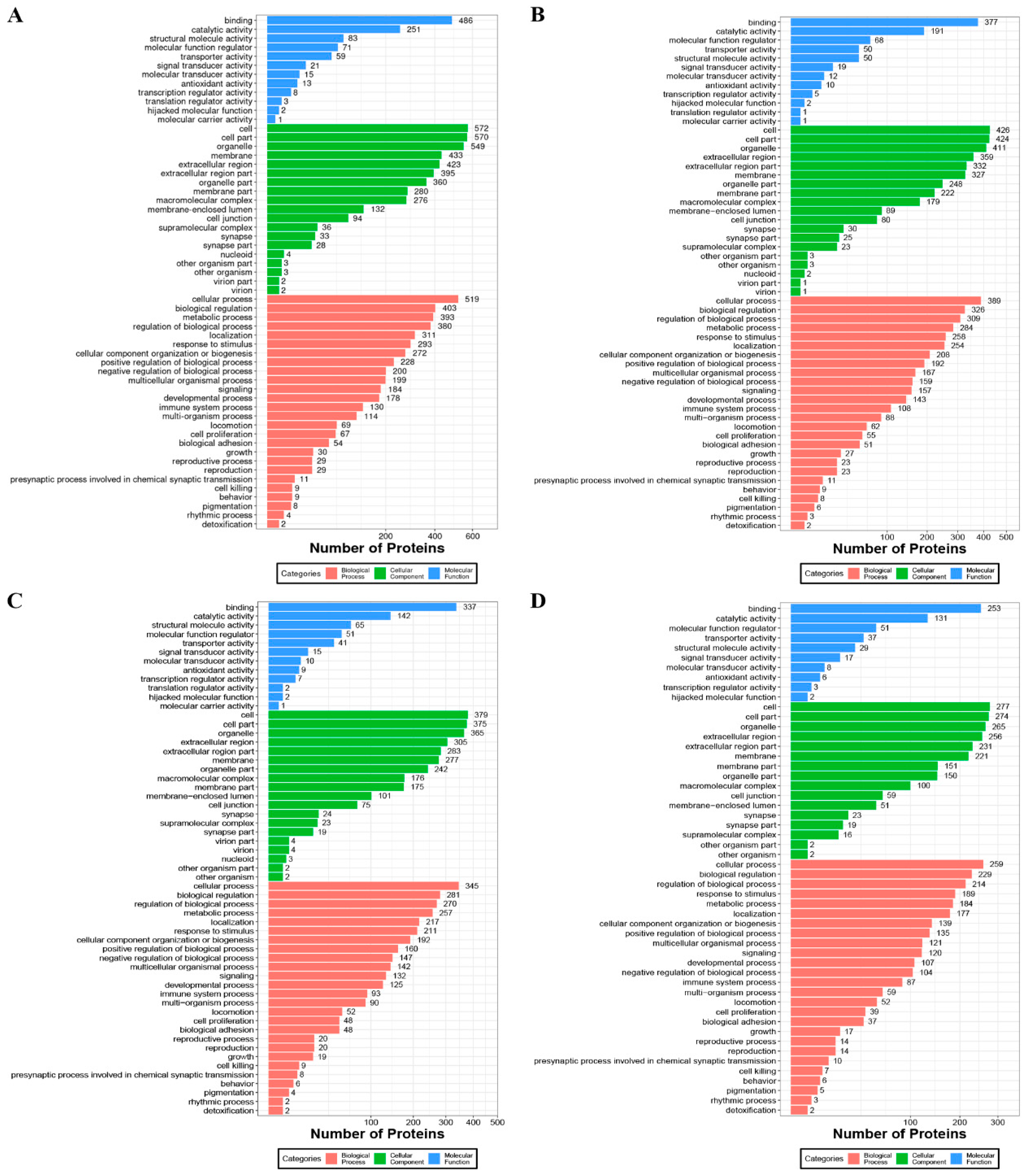
3.5. GO Analysis of Caprine MFGM Proteins in Relation to Different Heat Treatments
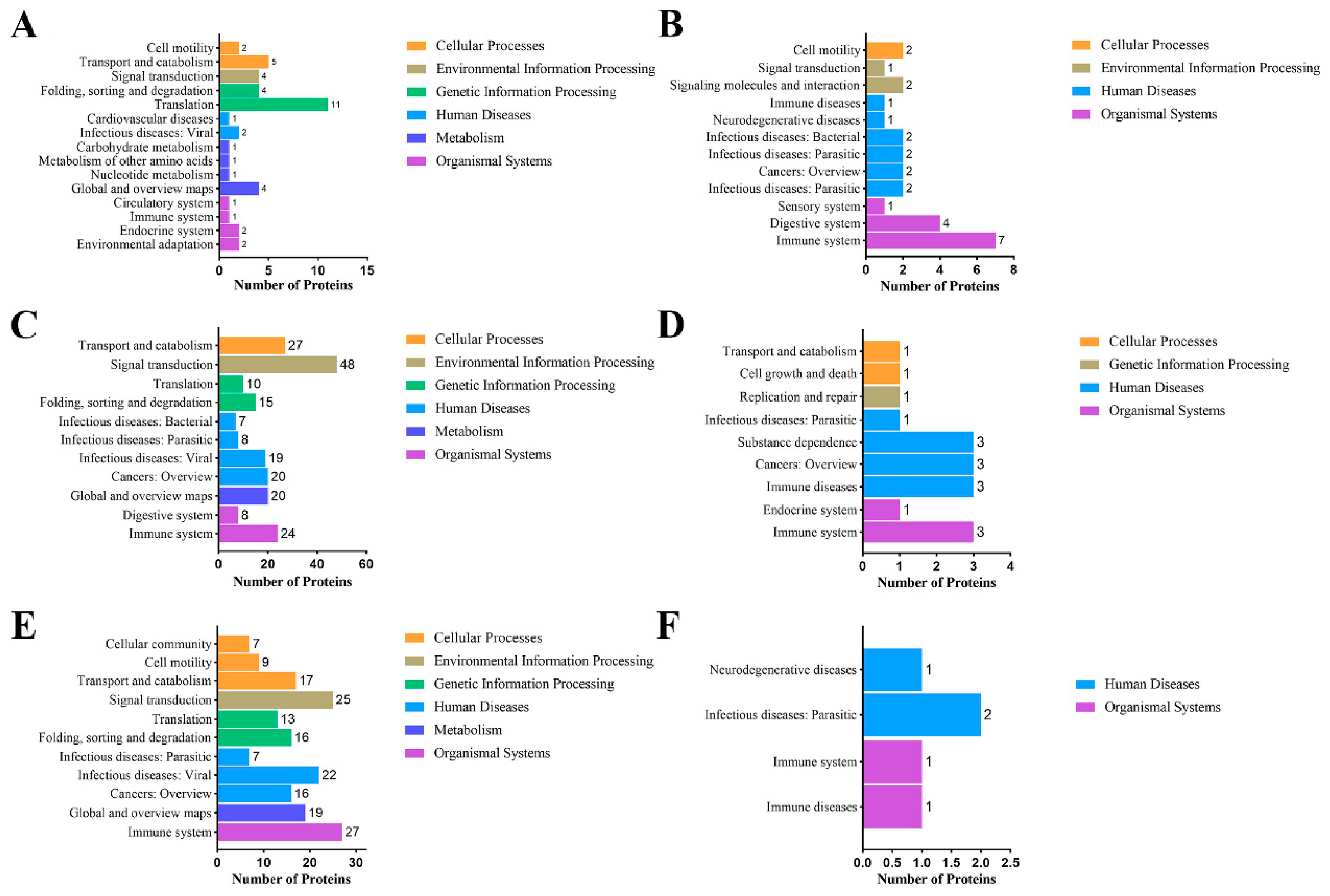
3.6. Pathway Analysis of Unique MFGM Proteins and Significantly Different MFGM Proteins
4. Discussion
4.1. Reliability of the Identified MFGM Proteins by Using Label-Free Proteomics Technique
4.2. Changes in the Caprine MFGM Proteins after Different Heat Treatment
4.3. Potential Physiological Functions Changes in MFGM Proteins after Different Heat Treatments
4.4. The Interaction between Skimmed Milk Proteins and MFGM Proteins under Heat Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, X.; Spencer, G.W.K.; Ong, L.; Gras, S.L. Beta casein proteins—A comparison between caprine and bovine milk. Trends Food Sci. Technol. 2022, 121, 30–43. [Google Scholar] [CrossRef]
- Clark, S.; Mora García, M.B. A 100-Year Review: Advances in goat milk research. J. Dairy Sci. 2017, 100, 10026–10044. [Google Scholar] [CrossRef]
- Selvaggi, M.; Laudadio, V.; Dario, C.; Tufarelli, V. Major proteins in goat milk: An updated overview on genetic variability. Mol. Biol. Rep. 2014, 41, 1035–1048. [Google Scholar] [CrossRef]
- Affolter, M.; Grass, L.; Vanrobaeys, F.; Casado, B.; Kussmann, M. Qualitative and quantitative profiling of the bovine milk fat globule membrane proteome. J. Proteom. 2010, 73, 1079–1088. [Google Scholar] [CrossRef]
- Wooding, F.B.P. The structure of the milk fat globule membrane. J. Ultrastruct. Res. 1971, 37, 388–400. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, N.; Zhao, X.; Yang, J.; Zhang, Y.; Han, R.; Qi, Y.; Zhao, S.; Li, S.; Wen, F.; et al. Changes in bovine milk fat globule membrane proteins caused by heat procedures using a label-free proteomic approach. Food Res. Int. 2018, 113, 1–8. [Google Scholar] [CrossRef]
- Dewettinck, K.; Rombaut, R.; Thienpont, N.; Le, T.T.; Messens, K.; Van Camp, J. Nutritional and technological aspects of milk fat globule membrane material. Int. Dairy J. 2008, 18, 436–457. [Google Scholar] [CrossRef]
- Yao, Y.; Zhao, G.; Yan, Y.; Mu, H.; Jin, Q.; Zou, X.; Wang, X. Milk fat globules by confocal Raman microscopy: Differences in human, bovine and caprine milk. Food Res. Int. 2016, 80, 61–69. [Google Scholar] [CrossRef]
- Raza, G.S.; Herzig, K.-H.; Leppäluoto, J. Invited review: Milk fat globule membrane—A possible panacea for neurodevelopment, infections, cardiometabolic diseases, and frailty. J. Dairy Sci. 2021, 104, 7345–7363. [Google Scholar] [CrossRef]
- Cebo, C.; Caillat, H.; Bouvier, F.; Martin, P. Major proteins of the goat milk fat globule membrane. J. Dairy Sci. 2010, 93, 868–876. [Google Scholar] [CrossRef] [Green Version]
- Zamora, A.; Guamis, B.; Trujillo, A.J. Protein composition of caprine milk fat globule membrane. Small Rumin. Res. 2009, 82, 122–129. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, L.; Wu, Y.; Zhou, P. Changes in milk fat globule membrane proteome after pasteurization in human, bovine and caprine species. Food Chem. 2019, 279, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, C.; Sun, X.; Jiang, S.; Guo, M. Characterization of the milk fat globule membrane proteome in colostrum and mature milk of Xinong Saanen goats. J. Dairy Sci. 2020, 103, 3017–3024. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wang, X.; Zhang, W.; Liu, L.; Pang, X.; Zhang, S.; Lv, J. Comparative proteomics of milk fat globule membrane in different species reveals variations in lactation and nutrition. Food Chem. 2016, 196, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Anema, S.G. Storage stability and age gelation of reconstituted ultra-high temperature skim milk. Int. Dairy J. 2017, 75, 56–67. [Google Scholar] [CrossRef]
- Shelke, P.A.; Sabikhi, L.; Khetra, Y.; Ganguly, S.; Baig, D. Effect of skim milk addition and heat treatment on characteristics of cow milk Ricotta cheese manufactured from Cheddar cheese whey. LWT 2022, 162, 113405. [Google Scholar] [CrossRef]
- Mahomud, M.S.; Katsuno, N.; Zhang, L.; Nishizu, T. Physical, rheological, and microstructural properties of whey protein enriched yogurt influenced by heating the milk at different pH values. J. Food Processing Preserv. 2017, 41, e13236. [Google Scholar] [CrossRef]
- Lu, J.; Pickova, J.; Daniel, G.; Langton, M. The role of key process steps on microstructural organisation of fat globules and lipid profiles in UHT milk processed in a pilot plant unit. Int. Dairy J. 2020, 109, 104741. [Google Scholar] [CrossRef]
- Sakkas, L.; Moutafi, A.; Moschopoulou, E.; Moatsou, G. Assessment of heat treatment of various types of milk. Food Chem. 2014, 159, 293–301. [Google Scholar] [CrossRef]
- Ye, A.; Singh, H.; James Oldfield, D.; Anema, S. Kinetics of heat-induced association of β-lactoglobulin and α-lactalbumin with milk fat globule membrane in whole milk. Int. Dairy J. 2004, 14, 389–398. [Google Scholar] [CrossRef]
- Sharma, P.; Oey, I.; Everett, D.W. Thermal properties of milk fat, xanthine oxidase, caseins and whey proteins in pulsed electric field-treated bovine whole milk. Food Chem. 2016, 207, 34–42. [Google Scholar] [CrossRef]
- Zanabria, R.; Tellez, A.M.; Griffiths, M.W.; Corredig, M. The antiproliferative properties of the milk fat globule membrane are affected by extensive heating. Dairy Sci. Technol. 2014, 94, 439–453. [Google Scholar] [CrossRef]
- Arena, S.; Renzone, G.; Novi, G.; Scaloni, A. Redox proteomics of fat globules unveils broad protein lactosylation and compositional changes in milk samples subjected to various technological procedures. J. Proteom. 2011, 74, 2453–2475. [Google Scholar] [CrossRef] [PubMed]
- Chandra Roy, M.; Zhang, L.; Liu, X.; Zhou, P. Investigation of caprine milk serum proteome and glycated proteome changes during heat treatment using robust ion mobility time-of-flight proteomic techniques. Int. Dairy J. 2020, 110, 104798. [Google Scholar] [CrossRef]
- Zhang, L.; Boeren, S.; Smits, M.; van Hooijdonk, T.; Vervoort, J.; Hettinga, K. Proteomic study on the stability of proteins in bovine, camel, and caprine milk sera after processing. Food Res. Int. 2016, 82, 104–111. [Google Scholar]
- Yuan, J.; Liu, D.; Peng, X.; Jiang, Y.; Gao, Y.; Liu, X.; Regenstein, J.M.; Zhou, P. The heat stability of caprine and bovine micellar casein dispersions. Int. Dairy J. 2022, 131, 105373. [Google Scholar] [CrossRef]
- Lee, H.; Zavaleta, N.; Chen, S.-Y.; Lönnerdal, B.; Slupsky, C. Effect of bovine milk fat globule membranes as a complementary food on the serum metabolome and immune markers of 6-11-month-old Peruvian infants. NPJ Sci. Food 2018, 2, 6. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.; Sun, X.; Guo, M. Comparative Proteomics of Whey and Milk Fat Globule Membrane Proteins of Guanzhong Goat and Holstein Cow Mature Milk. J. Food Sci. 2019, 84, 244–253. [Google Scholar] [CrossRef]
- Lu, J.; Liu, L.; Pang, X.; Zhang, S.; Jia, Z.; Ma, C.; Zhao, L.; Lv, J. Comparative proteomics of milk fat globule membrane in goat colostrum and mature milk. Food Chem. 2016, 209, 10–16. [Google Scholar] [CrossRef]
- Marques, I.T.O.; Vasconcelos, F.R.; Alves, J.P.M.; Montenegro, A.R.; Fernandes, C.C.L.; Oliveira, F.B.B.; Silva, C.P.; Nagano, C.S.; Figueiredo, F.C.; Beserra, F.J.; et al. Proteome of milk fat globule membrane and mammary gland tissue in goat fed different lipid supplementation. Small Rumin. Res. 2021, 199, 106378. [Google Scholar] [CrossRef]
- Sha, W.; Hou, C.; Yuan, E.; Li, Y.; Ren, J. Different processed milk with residual xanthine oxidase activity and risk of increasing serum uric acid level. Food Biosci. 2021, 40, 100892. [Google Scholar] [CrossRef]
- Lee, S.J.; Sherbon, J.W. Chemical changes in bovine milk fat globule membrane caused by heat treatment and homogenization of whole milk. J. Dairy Res. 2002, 69, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Gallier, S.; Cui, J.; Olson, T.D.; Rutherfurd, S.M.; Ye, A.; Moughan, P.J.; Singh, H. In vivo digestion of bovine milk fat globules: Effect of processing and interfacial structural changes. I. Gastric digestion. Food Chem. 2013, 141, 3273–3281. [Google Scholar]
- Ye, A.; Anema, S.G.; Singh, H. Behaviour of homogenized fat globules during the spray drying of whole milk. Int. Dairy J. 2007, 17, 374–382. [Google Scholar]
- Yvon, S.; Schwebel, L.; Belahcen, L.; Tormo, H.; Peter, M.; Haimoud-Lekhal, D.A.; Eutamene, H.; Jard, G. Effects of thermized donkey milk with lysozyme activity on altered gut barrier in mice exposed to water-avoidance stress. J. Dairy Sci. 2019, 102, 7697–7706. [Google Scholar] [CrossRef]
- Callewaert, L.; Van Herreweghe, J.M.; Vanderkelen, L.; Leysen, S.; Voet, A.; Michiels, C.W. Guards of the great wall: Bacterial lysozyme inhibitors. Trends Microbiol. 2012, 20, 501–510. [Google Scholar] [CrossRef]
- Smith, I.A.; Knezevic, B.R.; Ammann, J.U.; Rhodes, D.A.; Aw, D.; Palmer, D.B.; Mather, I.H.; Trowsdale, J. BTN1A1, the mammary gland butyrophilin, and BTN2A2 are both inhibitors of T cell activation. J. Immunol. 2010, 184, 3514–3525. [Google Scholar] [CrossRef]
- Trégoat, V.; Montagne, P.; Cuillière, M.L.; Béné, M.C.; Faure, G. C3/C4 concentration ratio reverses between colostrum and mature milk in human lactation. J. Clin. Immunol. 1999, 19, 300–304. [Google Scholar] [CrossRef]
- Trégoat, V.; Montagne, P.; Cuillière, M.L.; Béné, M.C.; Faure, G.C. Sequential C3 and C4 levels in human milk in relation to prematurity and parity. Clin. Chem. Lab. Med. 2000, 38, 609–613. [Google Scholar] [CrossRef]
- Ye, A.; Singh, H.; Taylor, M.W.; Anema, S. Interactions of whey proteins with milk fat globule membrane proteins during heat treatment of whole milk. Dairy Sci. Technol. 2004, 84, 411–417. [Google Scholar] [CrossRef]
- Corredig, M.; Dalgleish, D.G. Effect of different heat treatments on the strong binding interactions between whey proteins and milk fat globules in whole milk. J. Dairy Res. 1996, 63, 441–449. [Google Scholar] [CrossRef]
- Dalsgaard, T.K.; Nielsen, J.H.; Larsen, L.B. Proteolysis of milk proteins lactosylated in model systems. Mol. Nutr. Food Res. 2007, 51, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Du, M.; Mao, X. Change in interfacial properties of milk fat globules by homogenization and thermal processing plays a key role in their in vitro gastrointestinal digestion. Food Hydrocoll. 2019, 96, 331–342. [Google Scholar] [CrossRef]
- Houlihan, A.V.; Goddard, P.A.; Nottingham, S.M.; Kitchen, B.J.; Masters, C.J. Interactions between the bovine milk fat globule membrane and skim milk components on heating whole milk. J. Dairy Res. 1992, 59, 187–195. [Google Scholar] [CrossRef]
- Hansen, S.F.; Nielsen, S.D.; Rasmusen, J.T.; Larsen, L.B.; Wiking, L. Disulfide bond formation is not crucial for the heat-induced interaction between β-lactoglobulin and milk fat globule membrane proteins. J. Dairy Sci. 2020, 103, 5874–5881. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, D.; Zhang, L.; Zhu, Y.; Han, M.; Wang, Y.; Tang, J.; Zhou, P. Changes in Caprine Milk Fat Globule Membrane Proteins after Heat Treatment Using a Label-Free Proteomics Technique. Foods 2022, 11, 2705. https://doi.org/10.3390/foods11172705
Yan D, Zhang L, Zhu Y, Han M, Wang Y, Tang J, Zhou P. Changes in Caprine Milk Fat Globule Membrane Proteins after Heat Treatment Using a Label-Free Proteomics Technique. Foods. 2022; 11(17):2705. https://doi.org/10.3390/foods11172705
Chicago/Turabian StyleYan, Daomin, Lina Zhang, Yixuan Zhu, Mengyu Han, Yancong Wang, Jun Tang, and Peng Zhou. 2022. "Changes in Caprine Milk Fat Globule Membrane Proteins after Heat Treatment Using a Label-Free Proteomics Technique" Foods 11, no. 17: 2705. https://doi.org/10.3390/foods11172705