
Optimization of Sonication Parameters to Produce a Cashew Apple Bagasse Puree Rich in Superoxide Dismutase
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cashew Apple Bagasse
2.2. Sonication
2.3. Enzyme Extraction
2.4. Protein Determination
2.5. Enzyme Assays
2.6. H2O2 Content
2.7. Expression of Results
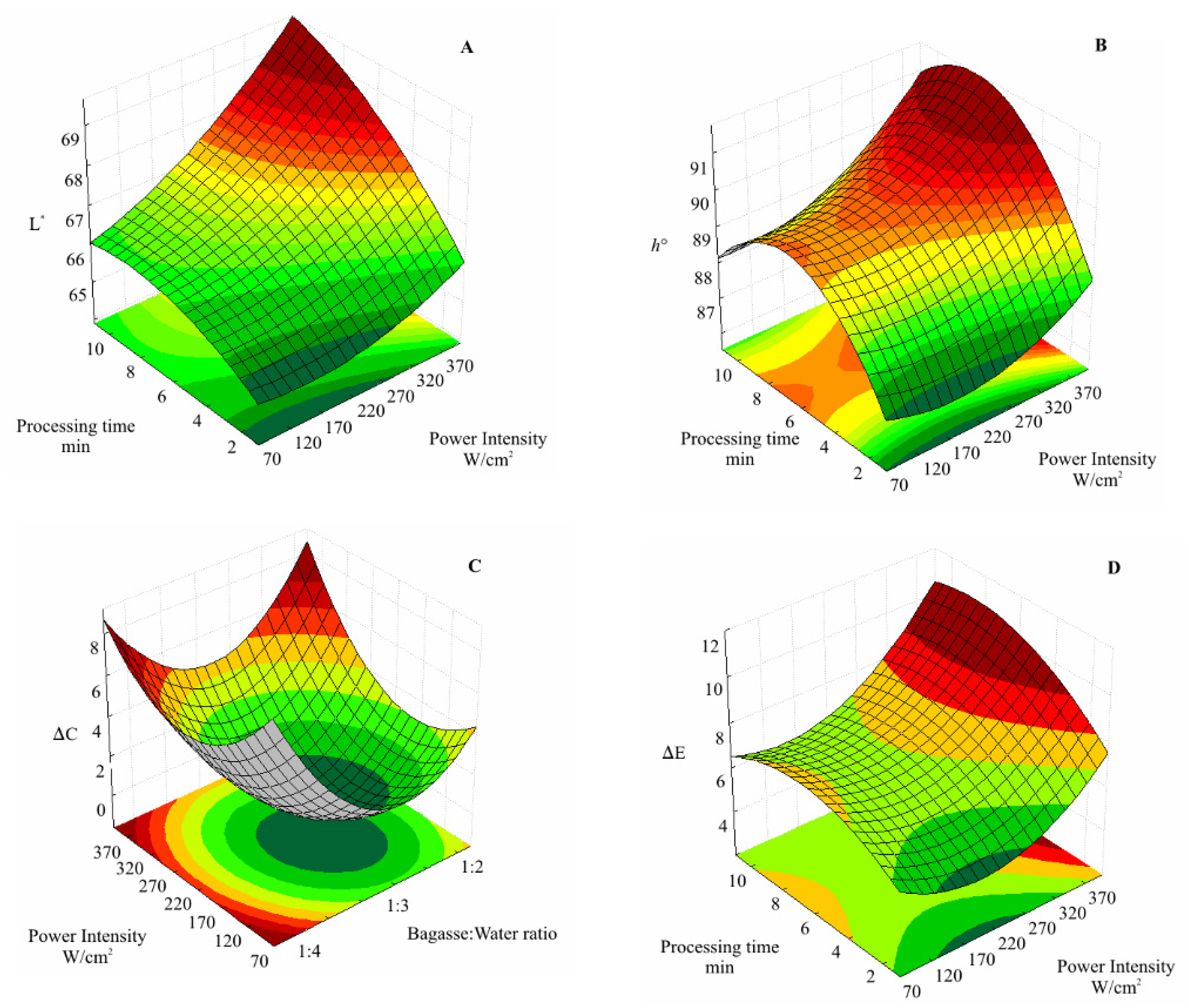
2.8. Color
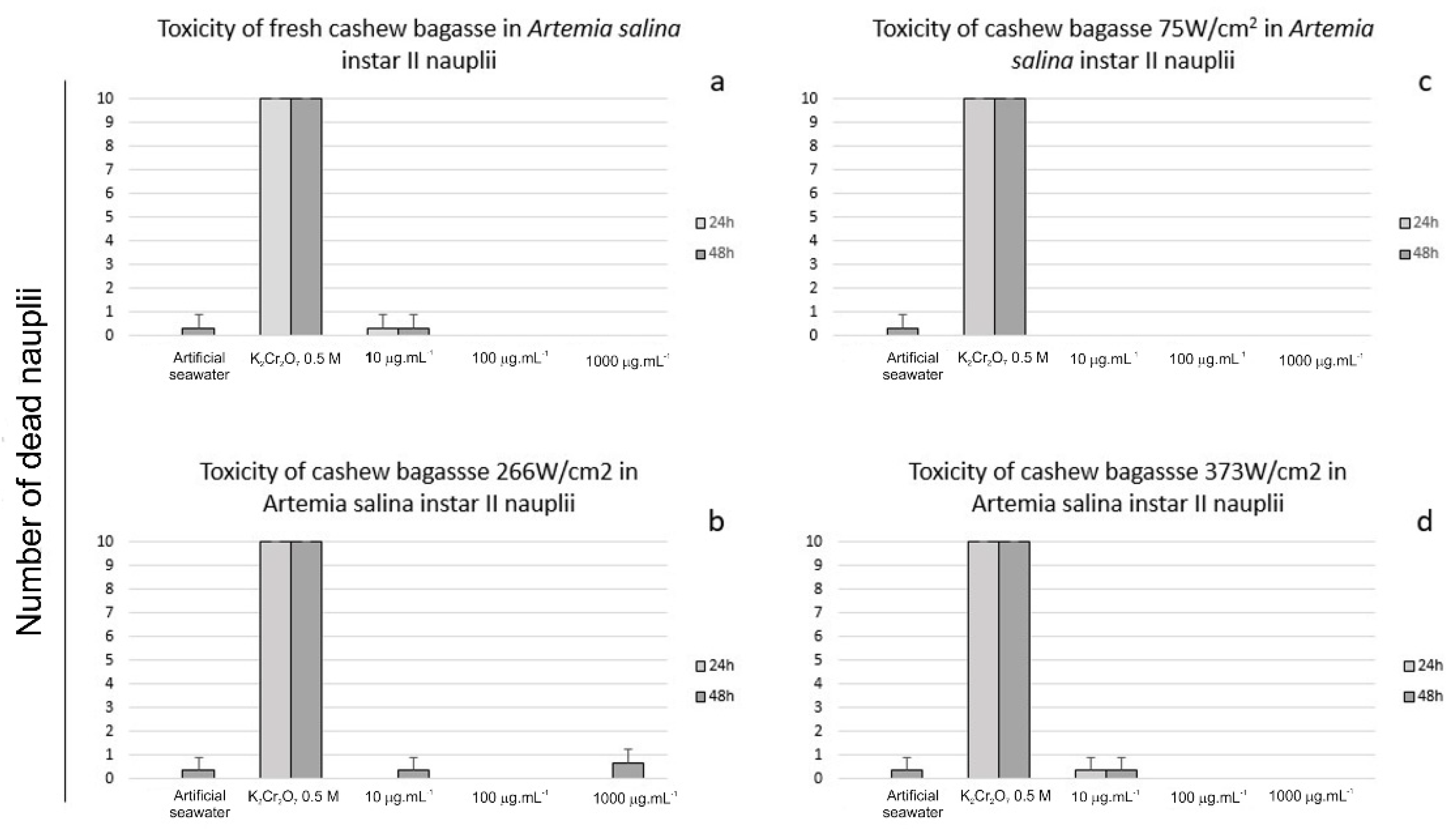
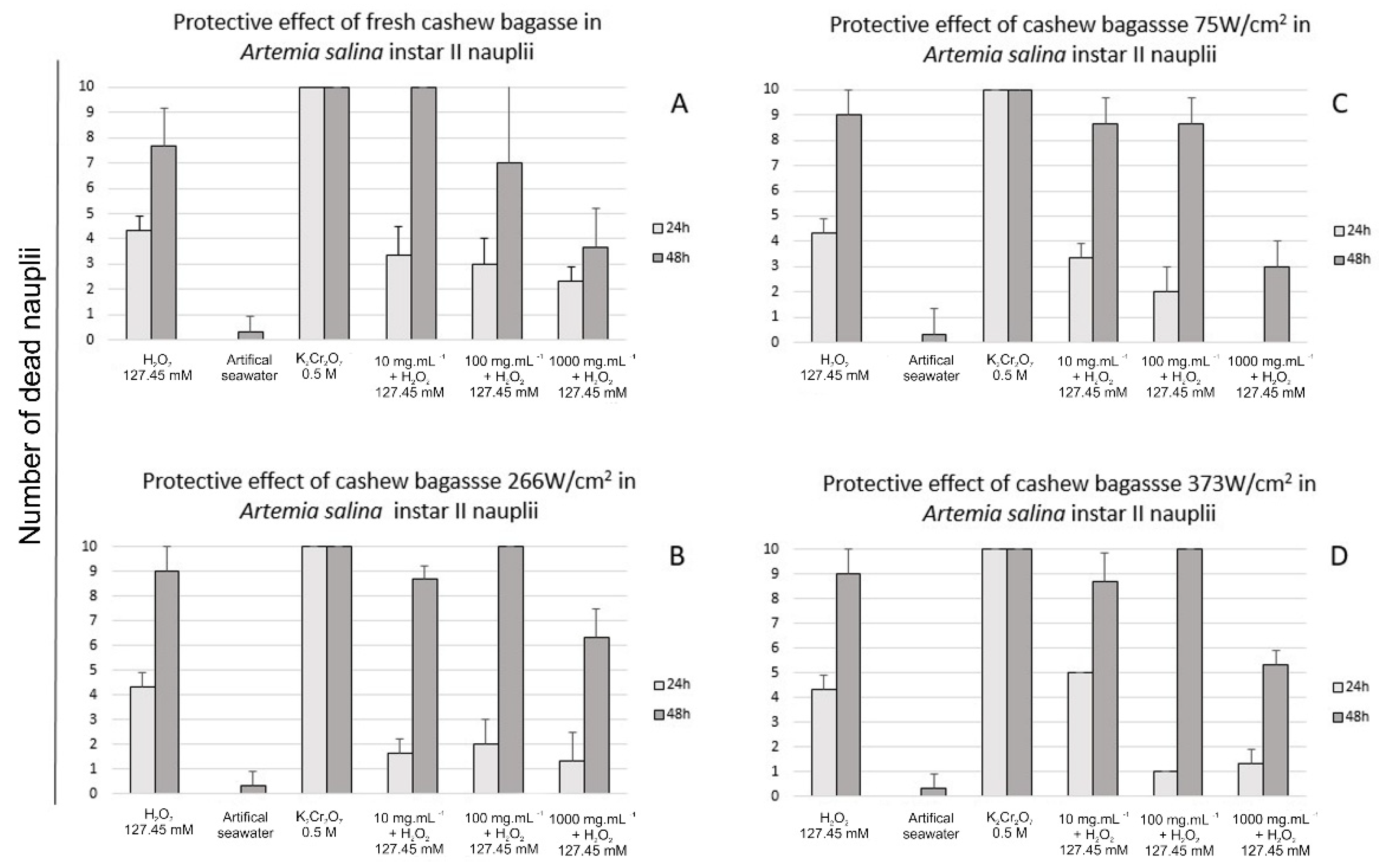
2.9. Toxicity against Artemia salina
2.9.1. Toxicity Assays
2.9.2. Protective Effect of Sonicated Cashew Apple Bagasse Puree against H2O2
2.10. Morphological Assays
2.10.1. Light Microscopy
2.10.2. Scanning Electron Microscopy (S.E.M.)
2.11. Statistical Analysis
3. Results
3.1. Activity of Antioxidative Enzymes
3.2. Toxicity and Protective Effect against Artemia salina
3.3. Morphological Evaluation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Čakar, U.; Obajić, S.; Vidović, B.; Djordjević, B. Nutritional and Lifestyle Habits of European Pharmacy Undergraduate Students. Prog. Nutr. 2018, 20, 38–45. [Google Scholar] [CrossRef]
- Bafana, A.; Dutt, S.; Kumar, S.; Ahuja, P.S. Superoxide Dismutase: An Industrial Perspective. Crit. Rev. Biotechnol. 2011, 31, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Cloarec, M.; Caillard, P.; Provost, J.C.; Dever, J.M.; Elbeze, Y.; Zamaria, N. Glisodin, a Vegetal SOD with Gliadin, as Preventative Agent vs. Atherosclerosis, as Confirmed with Carotid Ultrasound-B Imaging. Eur. Ann. Allergy Clin. Immunol. 2007, 39, 45–50. [Google Scholar] [PubMed]
- Fontas, E.; Montaudié, H.; Passeron, T. Oral Gliadin-Protected Superoxide Dismutase in Addition to Phototherapy for Treating Non-Segmental Vitiligo: A 24-Week Prospective Randomized Placebo-Controlled Study. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 1725–1729. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide Dismutases: Role in Redox Signaling, Vascular Function, and Diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef]
- Rosa, A.C.; Corsi, D.; Cavi, N.; Bruni, N.; Dosio, F. Superoxide Dismutase Administration: A Review of Proposed Human Uses. Molecules 2021, 26, 1844. [Google Scholar] [CrossRef]
- Xu, J.; Duan, X.; Yang, J.; Beeching, J.R.; Zhang, P. Enhanced Reactive Oxygen Species Scavenging by Overproduction of Superoxide Dismutase and Catalase Delays Postharvest Physiological Deterioration of Cassava Storage Roots. Plant Physiol. 2013, 161, 1517–1528. [Google Scholar] [CrossRef]
- Vouldoukis, I.; Lacan, D.; Kamate, C.; Coste, P.; Calenda, A.; Mazier, D.; Conti, M.; Dugas, B. Antioxidant and Anti-Inflammatory Properties of a Cucumis Melo LC. Extract Rich in Superoxide Dismutase Activity. J. Ethnopharmacol. 2004, 94, 67–75. [Google Scholar] [CrossRef]
- Sulastri, A.; Soemardji, A.A.; Sukrasno, S.; Amaliya, A. The Influence of Maturity Stage on Superoxide Dismutase and Catalase Activity in Red Guava Fruit; Atlantis Press: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Zafra, A.; Castro, A.J.; de Alché, J.D. Identification of Novel Superoxide Dismutase Isoenzymes in the Olive (Olea Europaea L.) Pollen. BMC Plant Biol. 2018, 18, 114. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, J.; Asch, F. Extraction, Storage Duration, and Storage Temperature Affect the Activity of Ascorbate Peroxidase, Glutathione Reductase, and Superoxide Dismutase in Rice Tissue. Biology 2019, 8, 70. [Google Scholar] [CrossRef]
- Sharma, S.; Bahuguna, S.; Kaur, N.; Chaudhary, N. Biochemical Aspects of Superoxide Dismutase Isolated from Amaranthus Spinosus: A Therapeutically Important Plant. Int. J. Genet. Eng. Biotechnol. 2014, 3073, 35–42. [Google Scholar]
- Gawlik-Dziki, U. Modification of Enzymatic and Non-Enzymatic in Vitro Oxidative Defence System by Bioaccessible Phytonutrients of Selected Spices. LWT Food Sci. Technol. 2014, 57, 434–441. [Google Scholar] [CrossRef]
- Hua, L.Y.; Ning, Z.; Yuebao, N. Determination of SOD in Black Ginger Extract and Its Effect on the Liver of Rats with Type 2 Diabetes. Food Sci. Technol. 2022, 42, 1–6. [Google Scholar] [CrossRef]
- Moreno-González, M.; Ottens, M. A Structured Approach to Recover Valuable Compounds from Agri-Food Side Streams. Food Bioprocess Technol. 2021, 14, 1387–1406. [Google Scholar] [CrossRef]
- Li, Y.; Fabiano-Tixier, A.S.; Tomao, V.; Cravotto, G.; Chemat, F. Green Ultrasound-Assisted Extraction of Carotenoids Based on the Bio-Refinery Concept Using Sunflower Oil as an Alternative Solvent. Ultrason. Sonochem. 2013, 20, 12–18. [Google Scholar] [CrossRef]
- Comarella, C.G.; Sautter, C.K.; Ebert, L.C.; Penna, N.G. Phenolic Compounds and Sensory Evaluation of Juice from Isabel Grapes Treated with Ultrasound. Braz. J. Food Technol. 2012, 4, 69–73. [Google Scholar] [CrossRef]
- Safari, M.; Ghanati, F.; Behmanesh, M.; Hajnorouzi, A.; Nahidian, B.; Mina, G. Enhancement of Antioxidant Enzymes Activity and Expression of CAT and PAL Genes in Hazel (Corylus Avellana L.) Cells in Response to Low-Intensity Ultrasound. Acta Physiol. Plant. 2013, 35, 2847–2855. [Google Scholar] [CrossRef]
- Bermúdez-Aguirre, D.; Mobbs, T.; Barbosa-Cánovas, G.V. Ultrasound Technologies for Food and Bioprocessing. In Ultrasound applications in Food Processing; Feng, H., Barbosa-Canovas, G., Weiss, J., Eds.; Springer: New York, NY, USA, 2011; pp. 65–105. [Google Scholar]
- Miguel, T.B.A.R.; Porto, E.C.M.; de Paiva Pinheiro, S.K.; de Castro Miguel, E.; Fernandes, F.A.N.; Rodrigues, S. Protective Effect of Natural and Processed Coconut Water by Non-Thermal Technologies Against Oxidative Stress in Brine Shrimp (Artemia salina). Food Bioprocess Technol. 2021, 14, 702–716. [Google Scholar] [CrossRef]
- Mason, T.J. Practical Sonochemistry User’s Guide to Application in Chemistry and Chemical Engineering; Ellis Horwood: New York, NY, USA, 1991. [Google Scholar]
- Wang, L.; Li, Z.; Huang, J.; Liu, D.; Lefebvre, C.; Fan, J. Effect of Ultrasound-Assisted Extraction of Polyphenols from Apple Peels in Water CO2 Systems. Food Bioprocess Technol. 2022, 15, 1157–1167. [Google Scholar] [CrossRef]
- Wissemann, K.W.; Lee, C.Y. Polyphenoloxidase Activity during Grape Maturation and Wine Production. Am. J. Enol. Vitic. 1980, 31, 206–211. [Google Scholar]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases. Plant Physiol. 1977, 309, 309–314. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A Spectrophotometric Method for Measuring the Breakdown of Hydrogen Peroxide by Catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Matsuno, H.; Uritani, I. Physiological Behavior of Peroxidase Isozymes in Sweet Potato Root Tissue Injured by Cutting or with Black Rot. Plant Cell Physiol. 1972, 13, 1091–1101. [Google Scholar]
- Mapelli, S.; Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The Effect of Drought and Ultraviolet Radiation on Growth and Stress Markers in Pea and Wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar]
- Meyer, B.N.; Ferrigni, N.R.; Putnam, J.E. Brine Shrimp: A Convenient General Bioassay for Active Plant Constituents. Planta Med. 1982, 45, 31–34. [Google Scholar] [CrossRef]
- Johari, S.A.; Rasmussen, K.; Gulumian, M.; Ghazi-Khansari, M.; Tetarazako, N.; Kashiwada, S.; Asghari, S.; Park, J.W.; Yu, I.J. Introducing a New Standardized Nanomaterial Environmental Toxicity Screening Testing Procedure, ISO/TS 20787: Aquatic Toxicity Assessment of Manufactured Nanomaterials in Saltwater Lakes Using Artemia Sp. Nauplii. Toxicol. Mech. Methods 2019, 29, 95–109. [Google Scholar] [CrossRef]
- Cao, X.; Cai, C.; Wang, Y.; Zheng, X. Effects of Ultrasound Processing on Physicochemical Parameters, Antioxidants, and Color Quality of Bayberry Juice. J. Food Qual. 2019. [Google Scholar] [CrossRef]
- Rojas-Contreras, J.A.; de la Rosa, A.P.B.; de León-Rodríguez, A. Expression and Characterization of a Recombinant Psychrophilic Cu/Zn Superoxide Dismutase from Deschampsia Antarctica E. Desv. [Poaceae]. Appl. Biochem. Biotechnol. 2015, 175, 3287–3296. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.G.; Fernandes, F.A.N.; de Siqueira Oliveira, L.; de Miranda, M.R.A. Influence of Ultrasound on Fresh-Cut Mango Quality Through Evaluation of Enzymatic and Oxidative Metabolism. Food Bioprocess Technol. 2015, 8, 1532–1542. [Google Scholar] [CrossRef]
- Chen, B.; Huang, J.; Wang, J.; Huang, L. Ultrasound Effects on the Antioxidative Defense Systems of Porphyridium Cruentum. Colloids Surf B Biointerfaces 2008, 61, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Fonteles, T.V.; Leite, A.K.F.; da Silva, A.R.A.; Fernandes, F.A.N.; Rodrigues, S. Sonication Effect on Bioactive Compounds of Cashew Apple Bagasse. Food Bioprocess Technol. 2017, 10, 1854–1864. [Google Scholar] [CrossRef]
- Chemat, F.; Zill-e-Huma; Khan, M.K. Applications of Ultrasound in Food Technology: Processing, Preservation and Extraction. Ultrason. Sonochem. 2011, 18, 813–835. [Google Scholar] [CrossRef]
- Kentish, S.; Ashokkumar, M. The Physical and Chemical Effects of Ultrasound. In Ultrasound Technologies for Food and Bioprocessing; Feng, H., Barbosa-Canóvas, G.V., Weiss, J., Eds.; Springer: London, UK, 2011; p. 678. ISBN 9781441974716. [Google Scholar]
- Wan, J.; Mawson, R.; Ashokkumar, M.; Ronacher, K.; Coventry, J.; Roginski, H.; Versteeg, C. Emerging Processing Technologies for Functional Foods. Aust. J. Dairy Technol. 2005, 60, 167–169. [Google Scholar]
- Martinez, M.V.; Whitaker, J.R. The Biochemistry and Control of Enzymatic Browning. Trends Food Sci. Technol. 1995, 6, 195–200. [Google Scholar] [CrossRef]
- Fadimu, G.J.; Gill, H.; Farahnaky, A.; Truong, T. Investigating the Impact of Ultrasound Pretreatment on the Physicochemical, Structural, and Antioxidant Properties of Lupin Protein Hydrolysates. Food Bioprocess Technol. 2021, 14, 2004–2019. [Google Scholar] [CrossRef]
- Choi, M.H.; Kim, G.H.; Lee, H. Effect of Ascorbic Acid Retention on Juice Color and Pigment Stability in Blood Orange (Citrus Sinensis) Juice during Refrigerated Storage. Food Res. Int. 2002, 35, 753–759. [Google Scholar] [CrossRef]
- Piechowiak, T.; Skóra, B.; Balawejder, M. Ozone Treatment Induces Changes in Antioxidative Defense System in Blueberry Fruit During Storage. Food Bioprocess Technol. 2020, 13, 1240–1245. [Google Scholar] [CrossRef]
- Čakar, U.; Čolović, M.; Milenković, D.; Medić, B.; Krstić, D.; Petrović, A.; Ðoređvić, B. Protective Effects of Fruit Wines against Hydrogen Peroxide—Induced Oxidative Stress in Rat Synaptosomes. Agronomy 2021, 11, 1414. [Google Scholar] [CrossRef]
- Yu Dong, X.; Liang Yang, Y. A Novel Approach to Enhance Blueberry Quality During Storage Using Cold Plasma at Atmospheric Air Pressure. Food Bioprocess Technol. 2019, 12, 1409–1421. [Google Scholar] [CrossRef]
- Gerbino, E.; Ghibaudo, F.; Tymczyszyn, E.E.; Gomez-Zavaglia, A.; Hugo, A.A. Probiotics, Galacto-Oligosaccharides, and Zinc Antagonize Biological Effects of Enterohaemorrhagic Escherichia Coli on Cultured Cells and Brine Shrimp Model. LWT 2020, 128, 109435. [Google Scholar] [CrossRef]
- Victoria Barahona, M.; Sánchez-Fortún, S. Protective Effect Induced by Atropine, Carbamates, and 2-Pyridine Aldoxime Methoiodide Artemia Salina Larvae Exposed to Fonofos and Phosphamidon. Ecotoxicol. Environ. Saf. 2007, 66, 65–73. [Google Scholar] [CrossRef]
- Lindsay, J.; Metcalf, J.S.; Codd, G.A. Protection against the Toxicity of Microcystin-LR and Cylindrospermopsin in Artemia Salina and Daphnia Spp. by Pre-Treatment with Cyanobacterial Lipopolysaccharide (LPS). Toxicon 2006, 48, 995–1001. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Factors | Coded Levels | ||
|---|---|---|---|
| −1 | 0 | +1 | |
| Power Intensity (W/cm2) | 75 | 226 | 373 |
| Bagasse: water ratio (g/mL) | 1:4 | 1:3 | 1:2 |
| Processing time (min) | 2 | 6 | 10 |
| Assay | Power Intensity (W/cm2) | Time (min) | Bagasse: Water Ratio | Residual (%) | |||||
|---|---|---|---|---|---|---|---|---|---|
| SOD | CAT | APX | H2O2 | PPO | POD | ||||
| 1 | 75 | 2 | 1:2 | 171.31 ± 0.08 | 100.61 ± 0.01 | 100.00 ± 0.10 | 84.21 ± 0.03 | 153.20 ± 2.45 | 77.32 ± 0.00 |
| 2 | 75 | 10 | 1:2 | 83.96 ± 1.67 | 112.45 ± 0.01 | 59.46 ± 0.09 | 175.00 ± 0.03 | 173.96 ± 2.89 | 34.97 ± 0.26 |
| 3 | 373 | 2 | 1:2 | 72.99 ± 0.99 | 95.91 ± 0.06 | 74.16 ± 0.23 | 140.78 ± 0.03 | 217.13 ± 1.86 | 98.46 ± 0.88 |
| 4 | 373 | 10 | 1:2 | 126.16 ± 0.24 | 85.67 ± 0.04 | 93.21 ± 0.45 | 486.84 ± 0.03 | 179.02 ± 2.21 | 48.18 ± 0.29 |
| 5 | 75 | 2 | 1:4 | 249.00 ± 0.76 | 108.76 ± 0.00 | 97.55 ± 0.05 | 248.68 ± 0.03 | 185.52 ± 5.72 | 37.05 ± 0.00 |
| 6 | 75 | 10 | 1:4 | 376.28 ± 0.56 | 110.86 ± 0.08 | 91.87 ± 0.09 | 164.47 ± 0.03 | 179.30 ± 9.31 | 56.47 ± 0.00 |
| 7 | 373 | 2 | 1:4 | 162.71 ± 0.97 | 119.22 ± 0.08 | 52.11 ± 0.08 | 109.21 ± 0.03 | 189.97 ± 2.31 | 49.41 ± 0.49 |
| 8 | 373 | 10 | 1:4 | 300.09 ± 1.67 | 108.76 ± 0.00 | 100.00 ± 0.01 | 161.84 ± 0.03 | 200 ± 3.20 | 39.11 ± 0.29 |
| 9 | 226 | 6 | 1:2 | 172.15 ± 0.54 | 104.56 ± 0.02 | 100.00 ± 0.32 | 109.21 ± 0.03 | 78.19 ± 0.78 | 27.61 ± 0.26 |
| 10 | 226 | 6 | 1:4 | 345.77 ± 0.57 | 112.95 ± 0.64 | 58.33 ± 0.09 | 98.68 ± 0.03 | 125.63 ± 4.29 | 49.41 ± 0.19 |
| 11 | 75 | 6 | 1:2 | 124.37 ± 0.76 | 87.71 ± 0.00 | 64.14 ± 0.00 | 119.73 ± 0.03 | 74.75 ± 3. 10 | 22.92 ± 0.39 |
| 12 | 373 | 6 | 1:3 | 44.11± 0.21 | 98.68 ± 0.00 | 72.16 ± 0.29 | 307.89 ± 0.03 | 97.50 ± 3.13 | 28.14 ± 0.36 |
| 13 | 226 | 2 | 1:3 | 92.01 ± 0.12 | 104.16 ± 0.09 | 53.45 ± 0.00 | 115.78 ± 0.03 | 127.05 ± 3.28 | 45.31 ± 0.54 |
| 14 | 226 | 10 | 1:3 | 85.29 ± 0.53 | 97.06 ± 0.08 | 50.00 ± 0.07 | 156.57 ± 0.03 | 129.59 ± 4.23 | 31.07 ± 0.00 |
| 15 | 226 | 6 | 1:3 | 103.36 ± 0.36 | 85.78 ± 0.01 | 51.00 ± 0.15 | 102.63 ± 0.03 | 83 ± 1.20 | 23.30 ± 0.19 |
| 16 | 226 | 6 | 1:3 | 105.00 ± 1.04 | 87.42 ± 0.03 | 49.77 ± 0.29 | 105.00 ± 0.03 | 84.57 ± 0.88 | 22.92 ± 0.67 |
| 17 | 226 | 6 | 1:3 | 102.07 ± 0.45 | 85.99 ± 0.05 | 47.00 ± 0.04 | 102.00 ± 0.03 | 85.54 ± 0.99 | 22.92 ± 0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonteles, T.; Leite, A.K.; Miguel, T.; Fernandes, F.; Pinheiro, S.; Miguel, E.; Rodrigues, S. Optimization of Sonication Parameters to Produce a Cashew Apple Bagasse Puree Rich in Superoxide Dismutase. Foods 2022, 11, 2694. https://doi.org/10.3390/foods11172694
Fonteles T, Leite AK, Miguel T, Fernandes F, Pinheiro S, Miguel E, Rodrigues S. Optimization of Sonication Parameters to Produce a Cashew Apple Bagasse Puree Rich in Superoxide Dismutase. Foods. 2022; 11(17):2694. https://doi.org/10.3390/foods11172694
Chicago/Turabian StyleFonteles, Thatyane, Ana Karoline Leite, Thaiz Miguel, Fabiano Fernandes, Sergimar Pinheiro, Emílio Miguel, and Sueli Rodrigues. 2022. "Optimization of Sonication Parameters to Produce a Cashew Apple Bagasse Puree Rich in Superoxide Dismutase" Foods 11, no. 17: 2694. https://doi.org/10.3390/foods11172694