
The Role of ptsH in Stress Adaptation and Virulence in Cronobacter sakazakii BAA-894

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
2.2. Construction of the ptsH Mutant
2.3. Growth Analysis of C. sakazakii WT Strain and ΔptsH Mutant
2.4. Environmental Stress Tolerance of C. sakazakii WT Strain and ΔptsH Mutant
2.5. Superoxide Dismutase (SOD) Activity Assessment
2.6. Specific Biofilm Formation (SBF) Assay
2.7. Adhesion to HT-29 Cells
2.8. RNA-Seq Analysis
2.9. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.10. Statistical Analysis
3. Results
3.1. Growth Curve
3.2. The ΔptsH Mutant Decreased Tolerance to Heat and SGF
3.3. The ΔptsH Mutant Enhanced Tolerance to Osmotic Stress
3.4. The ΔptsH Mutant Enhanced Tolerance to Oxidative Stress
3.5. The Deletion of ptsH Increased SOD Activity and Upregulated Genes Related to Oxidative and Osmotic Stress
3.6. The ΔptsH Mutant Attenuated the SBF Index and Adhesion to HT-29 Cells of C. sakazakii
3.7. Overall Transcriptome Comparison of WT Strain and ΔptsH Mutant
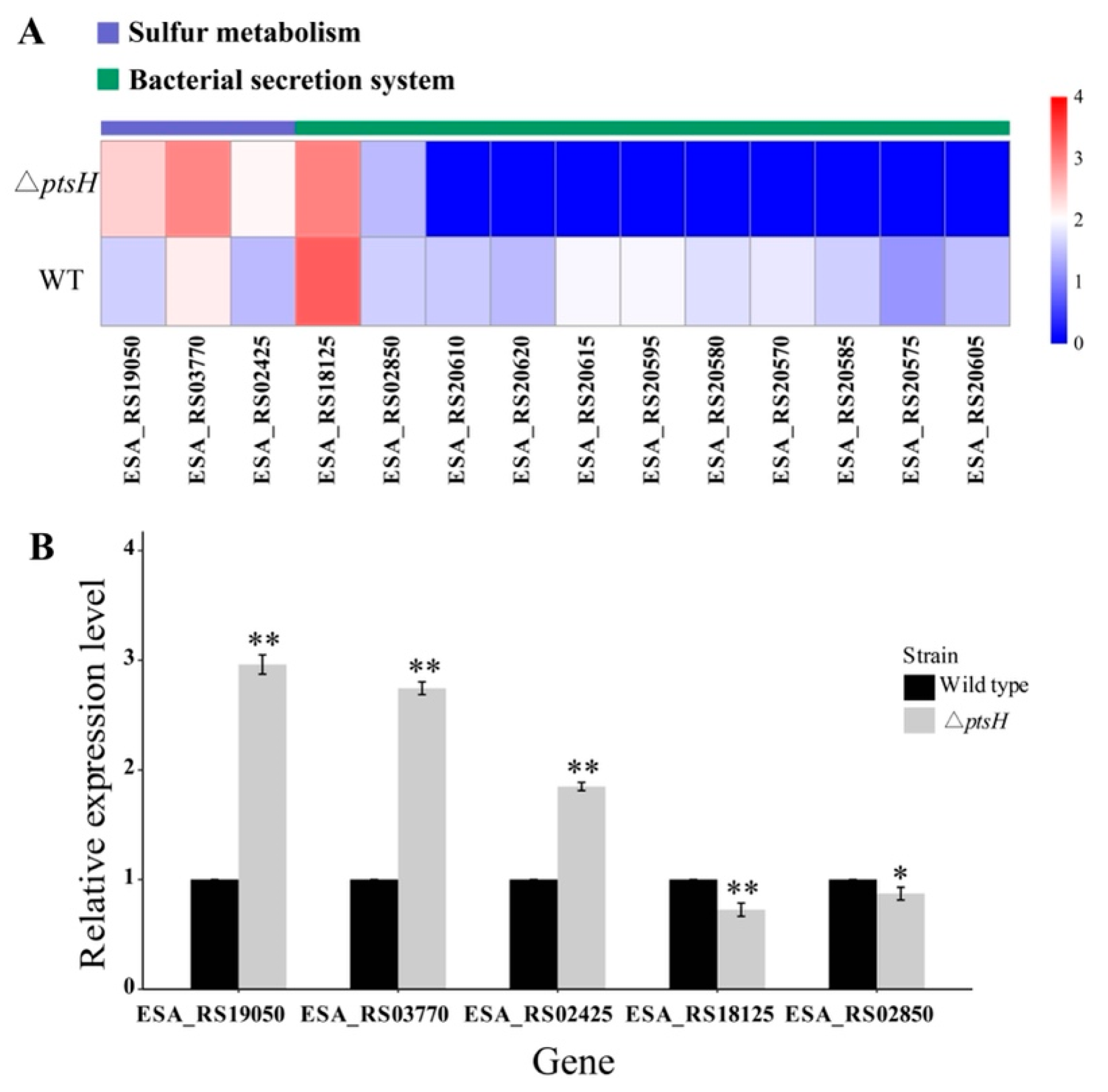
3.8. Analysis of Significant Differential Pathways
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Healy, B.; Cooney, S.; O’Brien, S.; Iversen, C.; Whyte, P.; Nally, J.; Callanan, J.J.; Fanning, S. Cronobacter (Enterobacter sakazakii): An Opportunistic Foodborne Pathogen. Foodborne Pathog. Dis. 2010, 7, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Beuchat, L.R.; Kim, H.; Gurtler, J.B.; Lin, L.-C.; Ryu, J.-H.; Richards, G.M. Cronobacter sakazakii in Foods and Factors Affecting Its Survival, Growth, and Inactivation. Int. J. Food Microbiol. 2009, 136, 204–213. [Google Scholar] [CrossRef]
- Fang, T.; Gurtler, J.B.; Huang, L. Growth Kinetics and Model Comparison of Cronobacter sakazakii in Reconstituted Powdered Infant Formula. J. Food Sci. 2012, 77, E247–E255. [Google Scholar] [CrossRef]
- Kim, K.; Jang, S.S.; Kim, S.K.; Park, J.-H.; Heu, S.; Ryu, S. Prevalence and Genetic Diversity of Enterobacter sakazakii in Ingredients of Infant Foods. Int. J. Food Microbiol. 2008, 122, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Fakruddin, M.; Rahaman, M.; Ahmed, M.M.; Hoque, M.M. Stress Tolerant Virulent Strains of Cronobacter sakazakii from Food. Biol. Res. 2014, 47, 63. [Google Scholar] [CrossRef]
- Liu, Q.; Mittal, R.; Emami, C.N.; Iversen, C.; Ford, H.R.; Prasadarao, N.V. Human Isolates of Cronobacter sakazakii Bind Efficiently to Intestinal Epithelial Cells In Vitro to Induce Monolayer Permeability and Apoptosis. J. Surg. Res. 2012, 176, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Amalaradjou, M.A.R.; Venkitanarayanan, K. Effect of Trans-Cinnamaldehyde on Reducing Resistance to Environmental Stresses in Cronobacter sakazakii. Foodborne Pathog. Dis. 2011, 8, 403–409. [Google Scholar] [CrossRef]
- Townsend, S.M.; Hurrell, E.; Gonzalez-Gomez, I.; Lowe, J.; Frye, J.G.; Forsythe, S.; Badger, J.L. Enterobacter sakazakii Invades Brain Capillary Endothelial Cells, Persists in Human Macrophages Influencing Cytokine Secretion and Induces Severe Brain Pathology in the Neonatal Rat. Microbiology 2007, 153, 3538–3547. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial Biofilms: From the Natural Environment to Infectious Diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Zhao, L.; Poh, C.N.; Wu, J.; Zhao, X.; He, Y.; Yang, H. Effects of Electrolysed Water Combined with Ultrasound on Inactivation Kinetics and Metabolite Profiles of Escherichia coli Biofilms on Food Contact Surface. Innov. Food Sci. Emerg. Technol. 2022, 76, 102917. [Google Scholar] [CrossRef]
- Grimm, M.; Stephan, R.; Iversen, C.; Manzardo, G.G.; Rattei, T.; Riedel, K.; Ruepp, A.; Frishman, D.; Lehner, A. Cellulose as an Extracellular Matrix Component Present in Enterobacter sakazakii Biofilms. J. Food Prot. 2008, 71, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.-H.; Hauk, P.; Cho, K.; Eo, Y.; Ma, X.; Stephens, K.; Cha, S.; Jeong, M.; Suh, J.-Y.; Sintim, H.O.; et al. Evidence of Link between Quorum Sensing and Sugar Metabolism in Escherichia coli Revealed via Cocrystal Structures of LsrK and HPr. Sci. Adv. 2018, 4, eaar7063. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, J.; Francke, C.; Postma, P.W. How Phosphotransferase System-Related Protein Phosphorylation Regulates Carbohydrate Metabolism in Bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 939–1031. [Google Scholar] [CrossRef]
- Gao, T.; Ding, M.; Yang, C.-H.; Fan, H.; Chai, Y.; Li, Y. The Phosphotransferase System Gene PtsH Plays an Important Role in MnSOD Production, Biofilm Formation, Swarming Motility, and Root Colonization in Bacillus cereus 905. Res. Microbiol. 2019, 170, 86–96. [Google Scholar] [CrossRef]
- Antunes, A.; Derkaoui, M.; Terrade, A.; Denizon, M.; Deghmane, A.-E.; Deutscher, J.; Delany, I.; Taha, M.-K. The Phosphocarrier Protein HPr Contributes to Meningococcal Survival during Infection. PLoS ONE. 2016, 11, e0162434. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; He, G.; Zhang, J. Physiological and Proteomic Analysis of Lactobacillus casei in Response to Acid Adaptation. J. Ind. Microbiol. Biotechnol. 2014, 41, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Chen, T.; Zhou, L.; Yang, H. Effect of Chlorine Sanitizer on Metabolic Responses of Escherichia coli Biofilms “Big Six” during Cross-Contamination from Abiotic Surface to Sponge Cake. Food Res. Int. 2022, 157, 111361. [Google Scholar] [CrossRef]
- Mao, Q.; Jiang, J.; Wu, X.; Xu, R.; Ma, Y.; Zhang, Y.; Shao, S.; Wang, Q. Coordinate Regulation of Carbohydrate Metabolism and Virulence by PtsH in Pathogen Edwardsiella piscicida. Appl. Microbiol. Biotechnol. 2022, 106, 2063–2077. [Google Scholar] [CrossRef]
- Cao, Y.; Li, L.; Zhang, Y.; Liu, F.; Xiao, X.; Li, X.; Yu, Y. SdiA Plays a Crucial Role in Stress Tolerance of C. sakazakii CICC 21544. LWT 2021, 143, 111189. [Google Scholar] [CrossRef]
- Fernández-Gómez, P.; López, M.; Prieto, M.; González-Raurich, M.; Alvarez-Ordóñez, A. The Role of the General Stress Response Regulator RpoS in Cronobacter sakazakii Biofilm Formation. Food Res. Int. 2020, 136, 109508. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, Y.; Chen, K.; Zhang, L.; Zhang, Y.; Wang, X.; Xia, X. The Role of PhoP/PhoQ Two Component System in Regulating Stress Adaptation in Cronobacter sakazakii. Food Microbiol. 2021, 100, 103851. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Enterobacter sakazakii Infections Associated with the Use of Powdered Infant Formula—Tennessee, 2001. MMWR. Morb. Mortal. Wkly. Rep. 2002, 51, 297–300. [Google Scholar]
- Xu, T.; Cao, H.; Zhu, W.; Wang, M.; Du, Y.; Yin, Z.; Chen, M.; Liu, Y.; Yang, B.; Liu, B. RNA-seq-based Monitoring of Gene Expression Changes of Viable but Non-culturable State of Vibrio cholerae Induced by Cold Seawater. Environ. Microbiol. Rep. 2018, 10, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Xu, L.; Xiao, L.; Zhu, K.; Song, Y.; Li, C.; Zhu, L.; Shen, X.; Wang, Y. RovM and CsrA Negatively Regulate Urease Expression in Yersinia pseudotuberculosis. Front. Microbiol. 2018, 9, 348. [Google Scholar] [CrossRef]
- Dancer, G.; Mah, J.; Rhee, M.; Hwang, I.; Kang, D. Resistance of Enterobacter sakazakii (Cronobacter Spp.) to Environmental Stresses. J. Appl. Microbiol. 2009, 107, 1606–1614. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-Y.; Kim, S.-K.; Choi, S.-Y.; You, D.-H.; Lee, S.-C.; Bang, W.-S.; Yuk, H.-G. Effect of Acid, Desiccation and Heat Stresses on the Viability of Cronobacter sakazakii during Rehydration of Powdered Infant Formula and in Simulated Gastric Fluid. Food Control. 2015, 50, 336–341. [Google Scholar] [CrossRef]
- Hu, Y.; Hu, Q.; Wei, R.; Li, R.; Zhao, D.; Ge, M.; Yao, Q.; Yu, X. The XRE Family Transcriptional Regulator SrtR in Streptococcus suis Is Involved in Oxidant Tolerance and Virulence. Front. Cell. Infect. Microbiol. 2019, 8, 452. [Google Scholar] [CrossRef]
- Naves, P.; del Prado, G.; Huelves, L.; Gracia, M.; Ruiz, V.; Blanco, J.; Dahbi, G.; Blanco, M.; del Carmen Ponte, M.; Soriano, F. Correlation between Virulence Factors and In Vitro Biofilm Formation by Escherichia coli Strains. Microb. Pathog. 2008, 45, 86–91. [Google Scholar] [CrossRef]
- Shi, C.; Yan, C.; Sui, Y.; Sun, Y.; Guo, D.; Chen, Y.; Jin, T.; Peng, X.; Ma, L.; Xia, X. Thymoquinone Inhibits Virulence Related Traits of Cronobacter sakazakii ATCC 29544 and Has Anti-Biofilm Formation Potential. Front. Microbiol. 2017, 8, 2220. [Google Scholar] [CrossRef]
- Li, G.; Yan, C.; Xu, Y.; Feng, Y.; Wu, Q.; Lv, X.; Yang, B.; Wang, X.; Xia, X. Punicalagin Inhibits Salmonella Virulence Factors and Has Anti-Quorum-Sensing Potential. Appl. Environ. Microbiol. 2014, 80, 6204–6211. [Google Scholar] [CrossRef]
- Cheng, C.; Yan, X.; Liu, B.; Jiang, T.; Zhou, Z.; Guo, F.; Zhang, Q.; Li, C.; Fang, T. SdiA Enhanced the Drug Resistance of Cronobacter sakazakii and Suppressed Its Motility, Adhesion and Biofilm Formation. Front. Microbiol. 2022, 13, 901912. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for Integration and Interpretation of Large-Scale Molecular Data Sets. Nucleic Acids Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V. Oxidative Stress and Mechanisms of Protection against It in Bacteria. Biochemistry 2001, 66, 476–489. [Google Scholar]
- Holý, O.; Forsythe, S. Cronobacter Spp. as Emerging Causes of Healthcare-Associated Infection. J. Hosp. Infect. 2014, 86, 169–177. [Google Scholar] [CrossRef]
- Stephens, K.; Pozo, M.; Tsao, C.-Y.; Hauk, P.; Bentley, W.E. Bacterial Co-Culture with Cell Signaling Translator and Growth Controller Modules for Autonomously Regulated Culture Composition. Nat. Commun. 2019, 10, 4129. [Google Scholar] [CrossRef]
- Pätzold, L.; Brausch, A.-C.; Bielefeld, E.-L.; Zimmer, L.; Somerville, G.A.; Bischoff, M.; Gaupp, R. Impact of the Histidine-Containing Phosphocarrier Protein HPr on Carbon Metabolism and Virulence in Staphylococcus aureus. Microorganisms 2021, 9, 466. [Google Scholar] [CrossRef]
- Liang, Q.; Zhang, F.; Li, Y.; Zhang, X.; Li, J.; Yang, P.; Qi, Q. Comparison of Individual Component Deletions in a Glucose-Specific Phosphotransferase System Revealed Their Different Applications. Sci. Rep. 2015, 5, 13200. [Google Scholar] [CrossRef]
- Yan, Q.; Power, K.A.; Cooney, S.; Fox, E.; Gopinath, G.R.; Grim, C.J.; Tall, B.D.; McCusker, M.P.; Fanning, S. Complete Genome Sequence and Phenotype Microarray Analysis of Cronobacter sakazakii SP291: A Persistent Isolate Cultured from a Powdered Infant Formula Production Facility. Front. Microbiol. 2013, 4, 256. [Google Scholar] [CrossRef]
- Edelson-Mammel, S.; Porteous, M.; Buchanan, R. Acid Resistance of Twelve Strains of Enterobacter sakazakii, and the Impact of Habituating the Cells to an Acidic Environment. J. Food Sci. 2006, 71, M201–M207. [Google Scholar] [CrossRef]
- Orieskova, M.; Kajsik, M.; Szemes, T.; Holy, O.; Forsythe, S.; Turna, J.; Drahovska, H. Contribution of the Thermotolerance Genomic Island to Increased Thermal Tolerance in Cronobacter Strains. Antonie Van Leeuwenhoek 2016, 109, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Pan, M.; Wan, C.; Shah, N.P.; Tao, X.; Wei, H. Physiological and Transcriptional Responses and Cross Protection of Lactobacillus plantarum ZDY2013 under Acid Stress. J. Dairy Sci. 2016, 99, 1002–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, B.; Hasic, M.; Chen, C.; Wai, S.N.; Milton, D.L. Type VI Secretion Modulates Quorum Sensing and Stress Response in Vibrio anguillarum. Environ. Microbiol. 2009, 11, 3018–3028. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Pan, J.; Wang, Y.; Shen, X. Type VI Secretion Systems Present New Insights on Pathogenic Yersinia. Front. Cell. Infect. Microbiol. 2018, 8, 260. [Google Scholar] [CrossRef] [PubMed]
- Lertpiriyapong, K.; Gamazon, E.R.; Feng, Y.; Park, D.S.; Pang, J.; Botka, G.; Graffam, M.E.; Ge, Z.; Fox, J.G. Campylobacter jejuni Type VI Secretion System: Roles in Adaptation to Deoxycholic Acid, Host Cell Adherence, Invasion, and In Vivo Colonization. PLoS ONE 2012, 7, e42842. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, X.; Shou, J.; Zong, B.; Zhang, Y.; Tan, J.; Chen, J.; Hu, L.; Zhu, Y.; Chen, H.; et al. Roles of Hcp Family Proteins in the Pathogenesis of the Porcine Extraintestinal Pathogenic Escherichia coli Type VI Secretion System. Sci. Rep. 2016, 6, 26816. [Google Scholar] [CrossRef]
- Azcarate-Peril, M.A.; Altermann, E.; Hoover-Fitzula, R.L.; Cano, R.J.; Klaenhammer, T.R. Identification and Inactivation of Genetic Loci Involved with Lactobacillus acidophilus Acid Tolerance. Appl. Environ. Microbiol. 2004, 70, 5315–5322. [Google Scholar] [CrossRef]
- Panyakampol, J.; Cheevadhanarak, S.; Sutheeworapong, S.; Chaijaruwanich, J.; Senachak, J.; Siangdung, W.; Jeamton, W.; Tanticharoen, M.; Paithoonrangsarid, K. Physiological and Transcriptional Responses to High Temperature in Arthrospira (Spirulina) platensis C1. Plant Cell Physiol. 2015, 56, 481–496. [Google Scholar] [CrossRef]
- Duval, V.; Lister, I.M. MarA, SoxS and Rob of Escherichia coli—Global Regulators of Multidrug Resistance, Virulence and Stress Response. Int. J. Biotechnol. Wellness Ind. 2013, 2, 101. [Google Scholar] [CrossRef]
- Osaili, T.; Forsythe, S. Desiccation Resistance and Persistence of Cronobacter Species in Infant Formula. Int. J. Food Microbiol. 2009, 136, 214–220. [Google Scholar] [CrossRef]
- Weber, H.; Polen, T.; Heuveling, J.; Wendisch, V.F.; Hengge, R. Genome-Wide Analysis of the General Stress Response Network in Escherichia coli: σS-Dependent Genes, Promoters, and Sigma Factor Selectivity. J. Bacteriol. 2005, 187, 1591–1603. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.M.; Norris, S.J.; Weinstock, G.M.; White, O.; Sutton, G.G.; Dodson, R.; Gwinn, M.; Hickey, E.K.; Clayton, R.; Ketchum, K.A. Complete Genome Sequence of Treponema pallidum, the Syphilis Spirochete. Science 1998, 281, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Qiao, J.; Wang, X. Role of Sigma Factor RpoS in Cronobacter sakazakii Environmental Stress Tolerance. Bioengineered 2021, 12, 2791–2809. [Google Scholar] [CrossRef] [PubMed]
- Price, N.L.; Raivio, T.L. Characterization of the Cpx Regulon in Escherichia coli Strain MC4100. J. Bacteriol. 2009, 191, 1798–1815. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Ordóñez, A.; Begley, M.; Clifford, T.; Deasy, T.; Collins, B.; Hill, C. Transposon Mutagenesis Reveals Genes Involved in Osmotic Stress and Drying in Cronobacter sakazakii. Food Res. Int. 2014, 55, 45–54. [Google Scholar] [CrossRef]
- Hengge-Aronis, R.; Lange, R.; Henneberg, N.; Fischer, D. Osmotic Regulation of RpoS-Dependent Genes in Escherichia coli. J. Bacteriol. 1993, 175, 259–265. [Google Scholar] [CrossRef]
- Serrano, L.M.; Molenaar, D.; Wels, M.; Teusink, B.; Bron, P.A.; De Vos, W.M.; Smid, E.J. Thioredoxin Reductase Is a Key Factor in the Oxidative Stress Response of Lactobacillus plantarum WCFS1. Microb. Cell Factories 2007, 6, 29. [Google Scholar] [CrossRef]
- Bin, P.; Huang, R.; Zhou, X. Oxidation Resistance of the Sulfur Amino Acids: Methionine and Cysteine. BioMed Res. Int. 2017, 2017, 9584932. [Google Scholar] [CrossRef]
- Wang, H.; Li, H.-X.; Fang, F.; Guo, J.; Chen, Y.-P.; Yan, P.; Yang, J.-X. Underlying Mechanisms of ANAMMOX Bacteria Adaptation to Salinity Stress. J. Ind. Microbiol. Biotechnol. 2019, 46, 573–585. [Google Scholar] [CrossRef]
- Feeney, A.; Kropp, K.A.; O’Connor, R.; Sleator, R.D. Cronobacter sakazakii: Stress Survival and Virulence Potential in an Opportunistic Foodborne Pathogen. Gut Microbes 2014, 5, 711–718. [Google Scholar] [CrossRef]
- Wang, H.; Wilksch, J.J.; Chen, L.; Tan, J.W.; Strugnell, R.A.; Gee, M.L. Influence of Fimbriae on Bacterial Adhesion and Viscoelasticity and Correlations of the Two Properties with Biofilm Formation. Langmuir 2017, 33, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Bowden, S.D.; Hopper-Chidlaw, A.C.; Rice, C.J.; Ramachandran, V.K.; Kelly, D.J.; Thompson, A. Nutritional and Metabolic Requirements for the Infection of HeLa Cells by Salmonella enterica serovar Typhimurium. PLoS ONE 2014, 9, e96266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, A.; Hardison, R.L.; Wallace, R.M.; Fitch, J.; Heimlich, D.R.; Bryan, M.O.; Dubois, L.; John-Williams, L.S.; Sebra, R.P.; White, P.; et al. Reprioritization of Biofilm Metabolism Is Associated with Nutrient Adaptation and Long-Term Survival of Haemophilus influenzae. NPJ Biofilms Microbiomes 2019, 5, 33. [Google Scholar] [CrossRef]
- Smith, T.J.; Font, M.E.; Kelly, C.M.; Sondermann, H.; O’Toole, G.A. An N-Terminal Retention Module Anchors the Giant Adhesin LapA of Pseudomonas fluorescens at the Cell Surface: A Novel Subfamily of Type I Secretion Systems. J. Bacteriol. 2018, 200, e00734-17. [Google Scholar] [CrossRef]
- Nishikawa, S.; Shinzawa, N.; Nakamura, K.; Ishigaki, K.; Abe, H.; Horiguchi, Y. The Bvg-repressed Gene BrtA, Encoding Biofilm-associated Surface Adhesin, Is Expressed during Host Infection by Bordetella bronchiseptica. Microbiol. Immunol. 2016, 60, 93–105. [Google Scholar] [CrossRef] [PubMed]
- De Pace, F.; de Paiva, J.B.; Nakazato, G.; Lancellotti, M.; Sircili, M.P.; Stehling, E.G.; da Silveira, W.D.; Sperandio, V. Characterization of IcmF of the Type VI Secretion System in an Avian Pathogenic Escherichia coli (APEC) Strain. Microbiology 2011, 157, 2954–2962. [Google Scholar] [CrossRef]
- Ma, J.; Bao, Y.; Sun, M.; Dong, W.; Pan, Z.; Zhang, W.; Lu, C.; Yao, H. Two Functional Type VI Secretion Systems in Avian Pathogenic Escherichia coli Are Involved in Different Pathogenic Pathways. Infect. Immun. 2014, 82, 3867–3879. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Ling, N.; Gao, J.; Zhang, X.; Zhang, M.; Tong, L.; Zeng, H.; Zhang, J.; Wu, Q. Roles of Outer Membrane Protein W (OmpW) on Survival, Morphology, and Biofilm Formation under NaCl Stresses in Cronobacter sakazakii. J. Dairy Sci. 2018, 101, 3844–3850. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Relevant Characteristics | Sources |
|---|---|---|
| E. coli S17 λpir | Tpr Smr recA thi pro rK- mK- RP4:2-Tc:MuKm Tn7 λ pir (thi pro hsdRhsdM + recA RP4-2-Tc:: Mu-Km-Tn7) | [23] |
| Cronobacter sakazakii | ||
| BAA-894 | WT, NA-induced mutant of C. sakazakii ATCC BAA-894, NAr | Laboratory collection |
| ΔptsH | ptsH deletion mutant of BAA-894, NAr | This study |
| Plasmid | ||
| pDM4 | Suicide vector, mobRK2, oriR6K, pir, sacB, Cmr | [24] |
| pDM4::UP+DOWN | Construct used for in-frame deletion of ptsH, Cmr | This study |
| Primer | Sequence (5′-3′) |
|---|---|
| UP-F | ACATGCATGCGAAAGCGGAAGAGATT |
| UP-R | GCTGGAACATTGTATTTCCCC |
| DOWN-F | GGGGAAATACAATGTTCCAGCGAACTCGAGTAAGTTCC |
| DOWN-R | ACGCGTCGACCTCTTCTTCGGTT |
| Primer | Sequence (5′-3′) |
|---|---|
| ESA_04030 | F, CCAGGGCTACACACGTGCTA |
| R, TCTCGCGAGGTCGCTTCT | |
| osmY | F, CGCGAAGGAACGATGTCAT |
| R, CCACCACCAGCGAAATCAA | |
| rpoS | F, CTGGTGGATTCGTCAGACCAT |
| R, GCGAATCGTACGGGTTTGG | |
| cpxR | F, TTGAGCTGGGCGCGGATGAT |
| R, TGCCGAGCACTTCCTGGCTT | |
| cpxA | F, ACCGCCCGCATCTTCGCCATTT |
| R, AACAACCGCCGCCACCACATCA | |
| sodA | F, CGAATCTGCCGGTTGAAGA |
| R, CTTGTCCGCCGGAACCT | |
| ESA_RS19050 | F, TATCCTCGCCGAGCCGACTGTT |
| R, CAGCCGCCGAACACGTCATCAA | |
| ESA_RS03770 | F, TGGAGTCACGTAACCCAAGC |
| R, CGACGGTAATCAGCTCGGTT | |
| ESA_RS02425 | F, CTTTCGGGCTCCGGAAAATC |
| R, TCAGGATTTGATGATATCTTCGCG | |
| ESA_RS18125 | F, GGCTGGACCGACATCACCTCTT |
| R, CGCTGGAGCAGTGCTTCAGGAT | |
| ESA_RS02850 | F, GACACGCTTCTGGATGAGGT |
| R, TCCCTTAACGCCTCTTTCACC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Li, J.; Yang, Y.; Yang, G.; Shi, Y.; Wang, S.; Wang, M.; Xia, X. The Role of ptsH in Stress Adaptation and Virulence in Cronobacter sakazakii BAA-894. Foods 2022, 11, 2680. https://doi.org/10.3390/foods11172680
Sun Y, Li J, Yang Y, Yang G, Shi Y, Wang S, Wang M, Xia X. The Role of ptsH in Stress Adaptation and Virulence in Cronobacter sakazakii BAA-894. Foods. 2022; 11(17):2680. https://doi.org/10.3390/foods11172680
Chicago/Turabian StyleSun, Yi, Jiahui Li, Yanpeng Yang, Gaoji Yang, Yiqi Shi, Shuo Wang, Muxue Wang, and Xiaodong Xia. 2022. "The Role of ptsH in Stress Adaptation and Virulence in Cronobacter sakazakii BAA-894" Foods 11, no. 17: 2680. https://doi.org/10.3390/foods11172680