
The Changes Occurring in Proteins during Processing and Storage of Fermented Meat Products and Their Regulation by Lactic Acid Bacteria

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Protein Changes in Fermented Meat Products
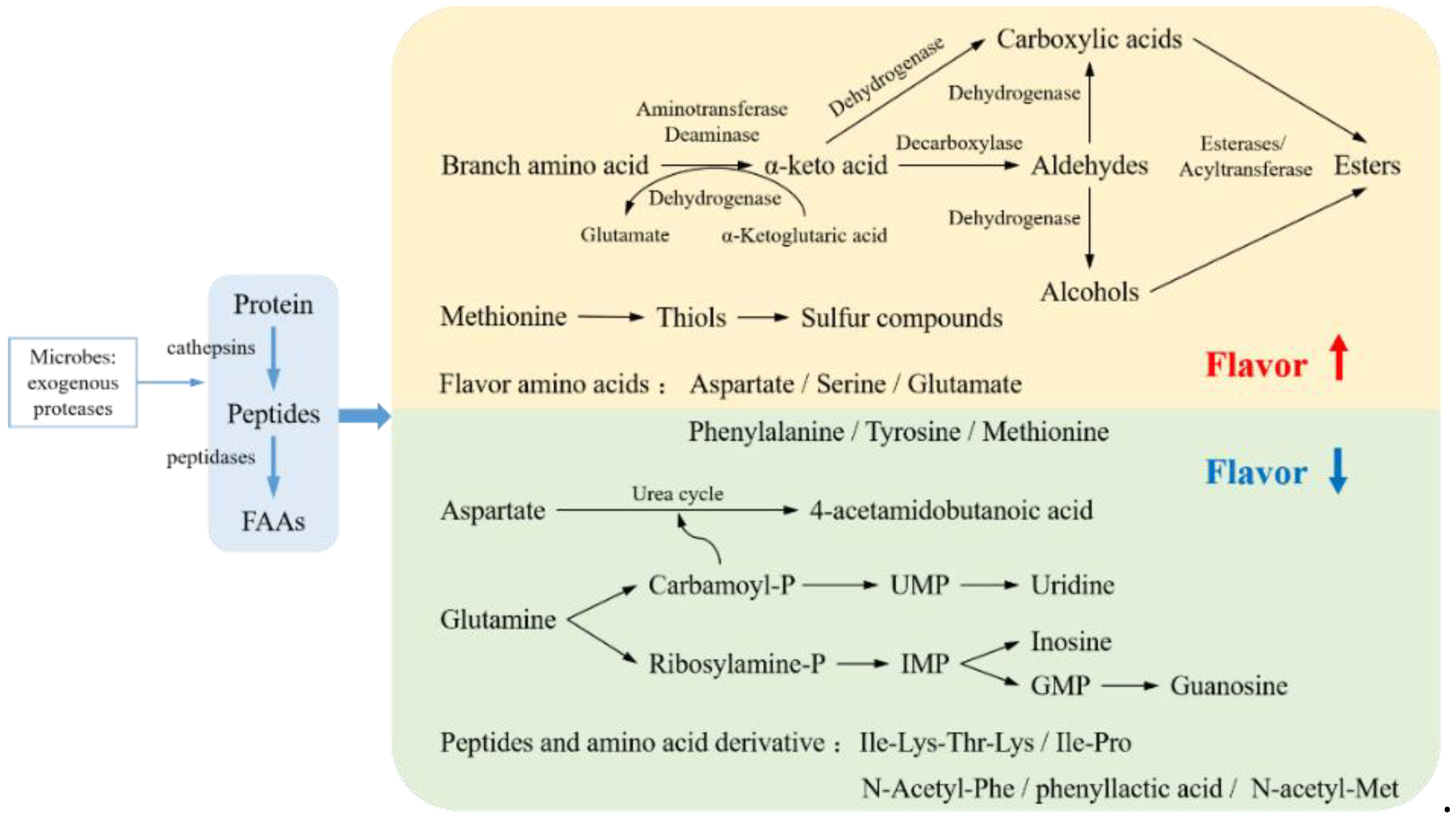
2.1. Protein Degradation
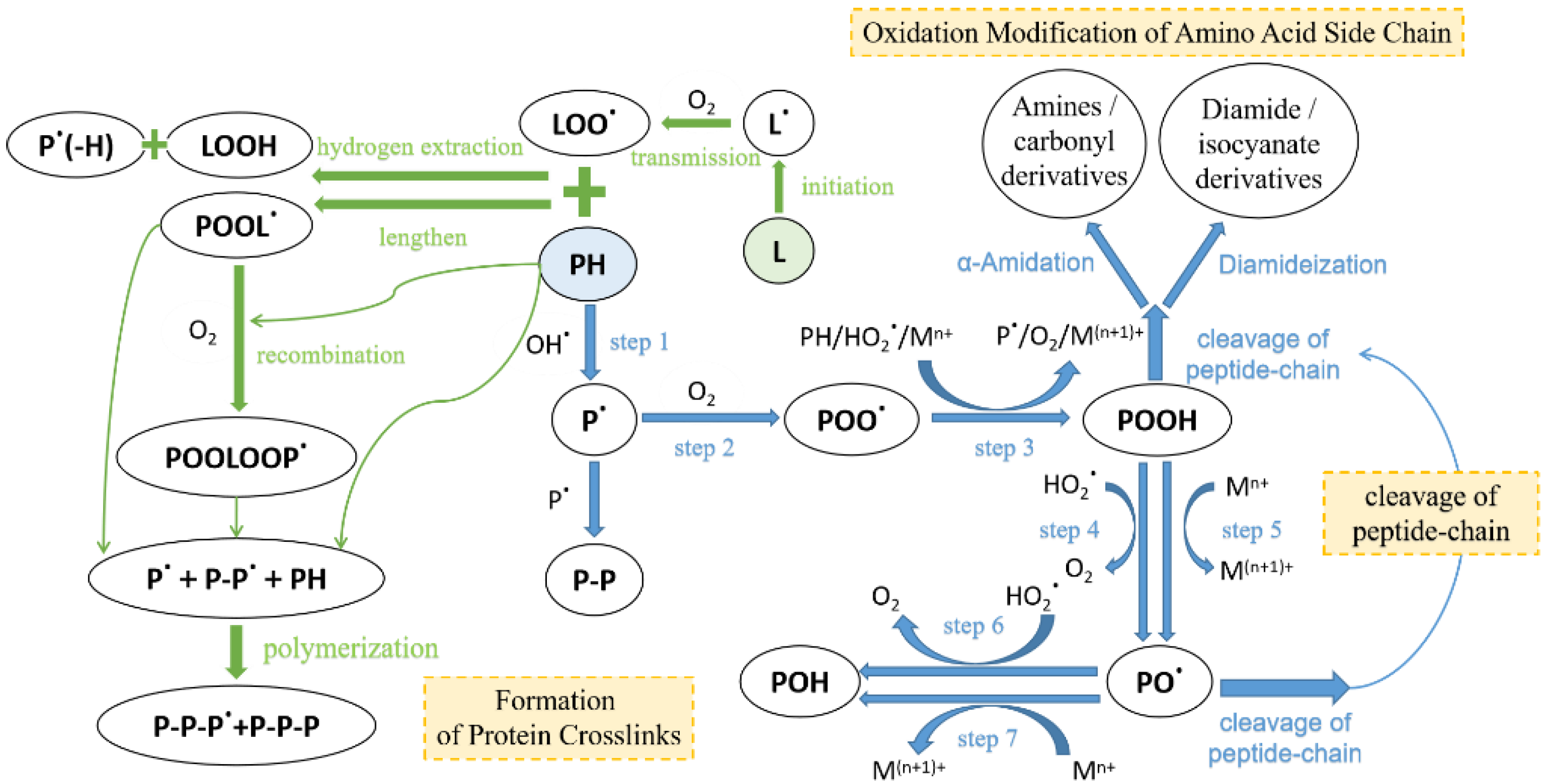
2.2. Protein Oxidation
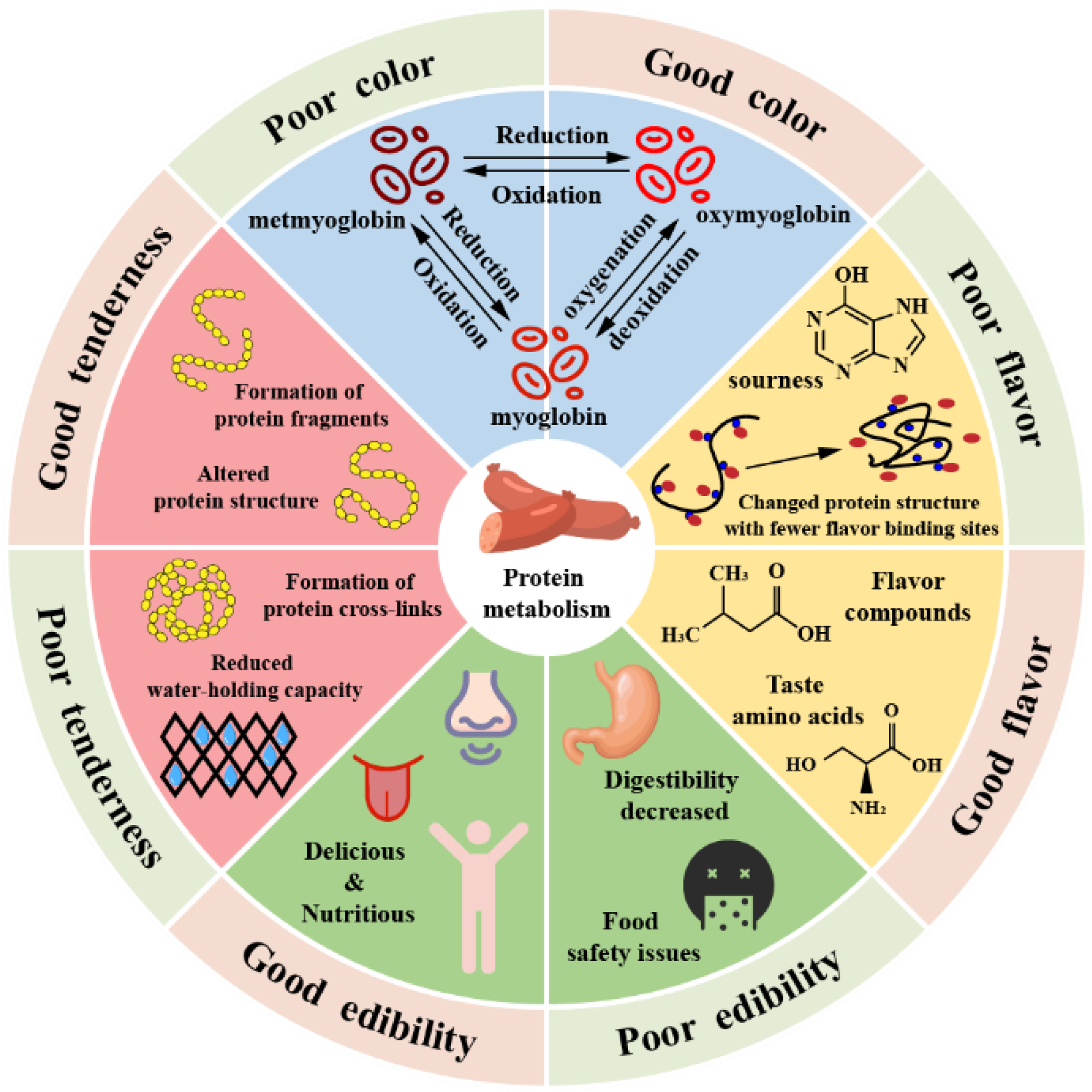
2.3. Effects of Protein Changes
2.3.1. Effects on Protein Structure
2.3.2. Impact on Product Quality
3. Regulatory Role of LAB
3.1. Acid Production
3.2. Protease Production
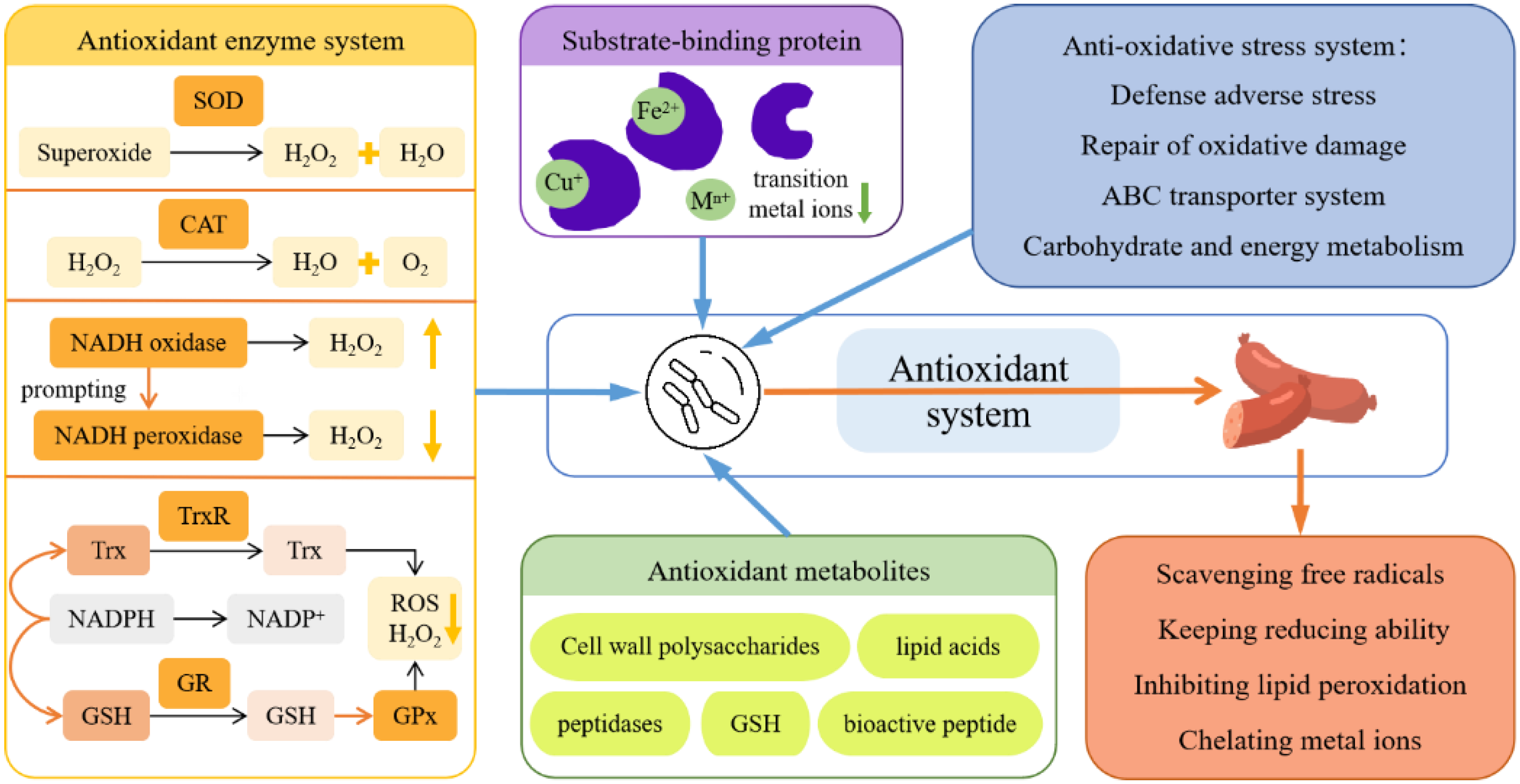
3.3. Antioxidant Properties
3.4. Effect on Quality
4. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Z.L.; Wang, Z.X.; Ji, L.L.; Zhang, J.M.; Zhao, Z.P.; Zhang, R.; Bai, T.; Hou, B.; Zhang, Y.; Liu, D.Y.; et al. A review: Microbial diversity and function of fermented meat products in China. Front. Microbiol. 2021, 12, 645435. [Google Scholar] [CrossRef] [PubMed]
- Talon, R.; Leroy, S.; Lebert, I. Microbial ecosystems of traditional fermented meat products: The importance of indigenous starters. Meat Sci. 2007, 77, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Halagarda, M.; Wójciak, K.M. Health and safety aspects of traditional European meat products. A review. Meat Sci. 2022, 184, 108623. [Google Scholar] [CrossRef] [PubMed]
- Aarti, C.; Khusro, A.; Varghese, R.; Arasu, M.V.; Agastian, P.; Al-Dhabi, N.A.; Ilavenil, S.; Choi, K.C. In vitro studies on probiotic and antioxidant properties of Lactobacillus brevis strain LAP2 isolated from Hentak, a fermented fish product of North-East India. LWT-Food Sci. Technol. 2017, 86, 438–446. [Google Scholar] [CrossRef]
- Pringsulaka, O.; Thongngam, N.; Suwannasai, N.; Atthakor, W.; Pothivejkul, K.; Rangsiruji, A. Partial characterisation of bacteriocins produced by lactic acid bacteria isolated from Thai fermented meat and fish products. Food Control 2012, 23, 547–551. [Google Scholar] [CrossRef]
- Soladoye, O.P.; Juárez, M.L.; Aalhus, J.L.; Shand, P.; Estévez, M. Protein oxidation in processed meat: Mechanisms and potential implications on human health. Compr. Rev. Food Sci. Food Saf. 2015, 14, 106–122. [Google Scholar] [CrossRef]
- Berardo, A.; Claeys, E.; Vossen, E.; Leroy, F.; De Smet, S. Protein oxidation affects proteolysis in a meat model system. Meat Sci. 2015, 106, 78–84. [Google Scholar] [CrossRef]
- Ying, H.Y.; Lang, Z.; Qian, L.; Yan, W.; Qian, C.; Hua, K.B. The potential correlation between bacterial diversity and the characteristic volatile flavour of traditional dry sausages from Northeast China. Food Microbiol. 2020, 91, 103505. [Google Scholar] [CrossRef]
- Zhang, Y.; Qin, Y.X.; Wang, Y.; Huang, Y.X.; Li, P.F.; Li, P.L. Lactobacillus plantarum LPL-1, a bacteriocin producing strain, changed the bacterial community composition and improved the safety of low-salt fermented sausages. LWT-Food Sci. Technol. 2020, 128, 109385. [Google Scholar] [CrossRef]
- Cao, C.C.; Feng, M.Q.; Sun, J.; Xu, X.L.; Zhou, G.H. Screening of lactic acid bacteria with high protease activity from fermented sausages and antioxidant activity assessment of its fermented sausages. CyTA-J. Food 2019, 17, 347–354. [Google Scholar] [CrossRef]
- Mei, L.; Pan, D.M.; Guo, T.T.; Ren, H.J.; Wang, L. Role of Lactobacillus plantarum with antioxidation properties on Chinese sausages. LWT-Food Sci. Technol. 2022, 162, 113427. [Google Scholar] [CrossRef]
- Feng, T.; Wang, J. Oxidative stress tolerance and antioxidant capacity of lactic acid bacteria as probiotic: A systematic review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.Y.; Li, Y.J.; Li, X.-a.; Zhang, H.W.; Chen, Q.; Kong, B.H. Application of lactic acid bacteria for improving the quality of reduced-salt dry fermented sausage: Texture, color, and flavor profiles. LWT-Food Sci. Technol. 2022, 154, 112723. [Google Scholar] [CrossRef]
- Chen, Q.; Kong, B.H.; Han, Q.; Xia, X.F.; Xu, L. The role of bacterial fermentation in lipolysis and lipid oxidation in Harbin dry sausages and its flavour development. LWT-Food Sci. Technol. 2017, 77, 389–396. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Pan, D.D.; Cao, J.X.; Zhou, G.H. A comprehensive review on molecular mechanism of defective dry-cured ham with excessive pastiness, adhesiveness, and bitterness by proteomics insights. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3838–3857. [Google Scholar] [CrossRef]
- Dalmis, U.; Soyer, A. Effect of processing methods and starter culture (Staphylococcus xylosus and Pediococcus pentosaceus) on proteolytic changes in Turkish sausages (sucuk) during ripening and storage. Meat Sci. 2008, 80, 345–354. [Google Scholar] [CrossRef]
- Ruiz-Moyano, S.; Martín, A.; Benito, M.J.; Hernández, A.; Casquete, R.; de Guia Córdoba, M. Application of Lactobacillus fermentum HL57 and Pediococcus acidilactici SP979 as potential probiotics in the manufacture of traditional Iberian dry-fermented sausages. Food Microbiol. 2011, 28, 839–847. [Google Scholar] [CrossRef]
- Ikonić, P.; Tasić, T.; Petrović, L.; Škaljac, S.; Jokanović, M.; Mandić, A.; Ikonić, B. Proteolysis and biogenic amines formation during the ripening of Petrovská klobása, traditional dry-fermented sausage from Northern Serbia. Food Control 2013, 30, 69–75. [Google Scholar] [CrossRef]
- Virgili, R.; Saccani, G.; Gabba, L.; Tanzi, E.; Bordini, C.S. Changes of free amino acids and biogenic amines during extended ageing of Italian dry-cured ham. LWT-Food Sci. Technol. 2007, 40, 871–878. [Google Scholar] [CrossRef]
- Wang, D.B.; Hu, G.H.; Wang, H.T.; Wang, L.M.; Zhang, Y.H.; Zou, Y.F.; Zhao, L.H.; Liu, F.; Jin, Y. Effect of mixed starters on proteolysis and formation of biogenic amines in dry fermented mutton sausages. Foods 2021, 10, 2939. [Google Scholar] [CrossRef]
- Yu, D.; Feng, M.Q.; Sun, J. Influence of mixed starters on the degradation of proteins and the formation of peptides with antioxidant activities in dry fermented sausages. Food Control 2021, 123, 107743. [Google Scholar] [CrossRef]
- Luan, X.X.; Feng, M.Q.; Sun, J. Effect of Lactobacillus plantarum on antioxidant activity in fermented sausage. Food Res. Int. 2021, 144, 110351. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Guo, M.T.; Wang, Q.L.; Dong, J.; Lu, S.L.; Lyu, B.; Ma, X.L. Antioxidant activities of peptides derived from mutton ham, Xuanwei ham and Jinhua ham. Food Res. Int. 2021, 142, 110195. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Wu, J.P.; Wan, J.; Zhou, Y.; Zhu, Q.J. Extraction and identification of bioactive peptides from Panxian dry-cured ham with multifunctional activities. LWT-Food Sci. Technol. 2022, 160, 113326. [Google Scholar] [CrossRef]
- Estévez, M. Protein carbonyls in meat systems: A review. Meat Sci. 2011, 89, 259–279. [Google Scholar] [CrossRef]
- Davies, M.J. The oxidative environment and protein damage. BBA-Proteins Proteom. 2005, 1703, 93–109. [Google Scholar] [CrossRef]
- Zhang, W.G.; Xiao, S.; Ahn, D.U. Protein oxidation: Basic principles and implications for meat quality. Crit. Rev. Food Sci. Nutr. 2013, 53, 1191–1201. [Google Scholar] [CrossRef]
- Wang, Z.M.; Zhou, H.; Zhou, K.; Tu, J.; Xu, B.C. An underlying softening mechanism in pale, soft and exudative—Like rabbit meat: The role of reactive oxygen species—Generating systems. Food Res. Int. 2022, 151, 110853. [Google Scholar] [CrossRef]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef]
- Stadtman, E.R.; Levine, R.L. Free radical-mediated oxidation of free amino acids and amino acid residues in proteins. Amino Acids 2003, 25, 207–218. [Google Scholar] [CrossRef]
- Park, D.; Xiong, Y.L. Oxidative modification of amino acids in porcine myofibrillar protein isolates exposed to three oxidizing systems. Food Chem. 2007, 103, 607–616. [Google Scholar] [CrossRef]
- Lv, C.Y.; Xu, C.; Gan, J.; Jiang, Z.H.; Wang, Y.M.; Cao, X.L. Roles of proteins/enzymes from animal sources in food quality and function. Foods 2021, 10, 1988. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Y.; Luo, H.L.; Cao, J.X.; Pan, D.D. Structural characteristics of Sheldrake meat and secondary structure of myofibrillar protein: Effects of oxidation. Int. J. Food Prop. 2017, 20, 1553–1566. [Google Scholar] [CrossRef]
- Zhang, D.; Li, H.J.; Emara, A.M.; Hu, Y.; Wang, Z.F.; Wang, M.Q.; He, Z.F. Effect of in vitro oxidation on the water retention mechanism of myofibrillar proteins gel from pork muscles. Food Chem. 2020, 315, 126226. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, D.H.; Tu, J.C.; Zhong, Y.J.; Zhang, D.; Wang, Z.M.; Tao, X.Q. Mechanisms of change in gel water-holding capacity of myofibrillar proteins affected by lipid oxidation: The role of protein unfolding and cross-linking. Food Chem. 2021, 344, 128587. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.P.; Feng, D.Y.; Xiao, J.; Wu, F.; He, X.J.; Xia, M.H.; Dong, T.; Liu, Y.H.; Tan, H.Z.; Zou, S.G.; et al. Studies on meat color, myoglobin content, enzyme activities, and genes associated with oxidative potential of pigs slaughtered at different growth stages. Asian-Australas. J. Anim. Sci. 2017, 30, 1739–1750. [Google Scholar] [CrossRef]
- Mitacek, R.M.; Ke, Y.L.; Prenni, J.E.; Jadeja, R.; VanOverbeke, D.L.; Mafi, G.G.; Ramanathan, R. Mitochondrial degeneration, depletion of NADH, and oxidative stress decrease color stability of wet-aged beef longissimus steaks. J. Food Sci. 2019, 84, 38–50. [Google Scholar] [CrossRef]
- Purslow, P.P.; Gagaoua, M.; Warner, R.D. Insights on meat quality from combining traditional studies and proteomics. Meat Sci. 2021, 174, 108423. [Google Scholar] [CrossRef]
- Guo, X.; Wang, Y.Q.; Lu, S.L.; Wang, J.Y.; Fu, H.H.; Gu, B.Y.; Lyu, B.; Wang, Q.L. Changes in proteolysis, protein oxidation, flavor, color and texture of dry-cured mutton ham during storage. LWT-Food Sci. Technol. 2021, 149, 111860. [Google Scholar] [CrossRef]
- Harkouss, R.; Astruc, T.; Lebert, A.; Gatellier, P.; Loison, O.; Safa, H.; Portanguen, S.; Parafita, E.; Mirade, P.-S. Quantitative study of the relationships among proteolysis, lipid oxidation, structure and texture throughout the dry-cured ham process. Food Chem. 2015, 166, 522–530. [Google Scholar] [CrossRef]
- Lonergan, E.H.; Zhang, W.; Lonergan, S.M. Biochemistry of postmortem muscle—Lessons on mechanisms of meat tenderization. Meat Sci. 2010, 86, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, Y.; He, J.; Pan, D.D.; Wang, H.F.; Cao, J.X. Evaluating the profile of myofibrillar proteins and its relationship with tenderness among five styles of dry-cured hams. Int. J. Food Sci. Technol. 2020, 56, 259–268. [Google Scholar] [CrossRef]
- Malheiros, J.M.; Braga, C.P.; Grove, R.A.; Ribeiro, F.A.; Calkins, C.R.; Adamec, J.; Chardulo, L.A.L. Influence of oxidative damage to proteins on meat tenderness using a proteomics approach. Meat Sci. 2019, 148, 64–71. [Google Scholar] [CrossRef]
- Zhang, M.; He, L.C.; Li, C.L.; Yang, F.; Zhao, S.L.; Liang, Y.H.; Jin, G.F. Effects of gamma ray irradiation-induced protein hydrolysis and oxidation on tenderness change of fresh pork during storage. Meat Sci. 2020, 163, 108058. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.Z.; Zhou, F.B.; Zhao, M.M.; Yang, B.; Cui, C. Physicochemical changes of myofibrillar proteins during processing of Cantonese sausage in relation to their aggregation behaviour and in vitro digestibility. Food Chem. 2011, 129, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Lametsch, R.; Lonergan, S.; Huff-Lonergan, E. Disulfide bond within µ-calpain active site inhibits activity and autolysis. BBA-Proteins Proteom. 2008, 1784, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Gagaoua, M.; Mullen, A.M.; Kelly, A.L.; Sweeney, T.; Cafferky, J.; Viala, D.; Hamill, R.M. A proteomic study for the discovery of beef tenderness biomarkers and prediction of warner-bratzler shear force measured on longissimus thoracis muscles of young limousin-sired bulls. Foods 2021, 10, 952. [Google Scholar] [CrossRef]
- Liao, R.Y.; Xia, Q.; Zhou, C.Y.; Geng, F.; Wang, Y.; Sun, Y.Y.; He, J.; Pan, D.D.; Cao, J.X. LC-MS/MS-based metabolomics and sensory evaluation characterize metabolites and texture of normal and spoiled dry-cured hams. Food Chem. 2022, 371, 131156. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Liu, F.; Jin, Z.; Xia, X. Ecological succession and functional characteristics of lactic acid bacteria in traditional fermented foods. Crit. Rev. Food Sci. Nutr. 2022, 1–15. [Google Scholar] [CrossRef]
- Tian, Z.Q.; Zhu, Q.J.; Chen, Y.S.; Zhou, Y.; Hu, K.; Li, H.Y.; Lu, K.; Zhou, J.; Liu, Y.; Chen, X. Studies on flavor compounds and free amino acid dynamic characteristics of fermented pork loin ham with a complex starter. Foods 2022, 11, 1501. [Google Scholar] [CrossRef]
- Chen, Q.; Hu, Y.Y.; Wen, R.X.; Wang, Y.; Qin, L.G.; Kong, B.H. Characterisation of the flavour profile of dry fermented sausages with different NaCl substitutes using HS-SPME-GC-MS combined with electronic nose and electronic tongue. Meat Sci. 2021, 172, 108338. [Google Scholar] [CrossRef] [PubMed]
- Estévez, M.; Ventanas, S.; Heinonen, M. Formation of Strecker aldehydes between protein carbonyls—α-Aminoadipic and γ-glutamic semialdehydes-and leucine and isoleucine. Food Chem. 2011, 128, 1051–1057. [Google Scholar] [CrossRef]
- Guichard, E. Flavour retention and release from protein solutions. Biotechnol. Adv. 2006, 24, 226–229. [Google Scholar] [CrossRef]
- Wang, H.T.; Guan, H.N.; Zhang, H.W.; Liu, H.T.; Chen, Q.; Kong, B.H. Elucidation of interaction mechanisms between myofibrillar proteins and ethyl octanoate by SPME-GC-MS, molecular docking and dynamics simulation. LWT-Food Sci. Technol. 2022, 154, 112787. [Google Scholar] [CrossRef]
- Zhou, F.B.; Zhao, M.M.; Su, G.W.; Sun, W.Z. Binding of aroma compounds with myofibrillar proteins modified by a hydroxyl-radical-induced oxidative system. J. Agric. Food Chem. 2014, 62, 9544–9552. [Google Scholar] [CrossRef]
- Zhang, J.; Kang, D.-C.; Zhang, W.-G.; Lorenzo, J.-M. Recent advantage of interactions of protein-flavor in foods: Perspective of theoretical models, protein properties and extrinsic factors. Trends Food Sci. Technol. 2021, 111, 405–425. [Google Scholar] [CrossRef]
- Flores, M.; Mora, L.; Reig, M.; Toldrá, F. Risk assessment of chemical substances of safety concern generated in processed meats. Food Sci. Hum. Wellness 2019, 8, 244–251. [Google Scholar] [CrossRef]
- Ma, J.B.; Wang, X.Y.; Li, Q.; Zhang, L.; Wang, Z.; Han, L.; Yu, Q.L. Oxidation of myofibrillar protein and crosslinking behavior during processing of traditional air-dried yak (Bos grunniens) meat in relation to digestibility. LWT-Food Sci. Technol. 2021, 142, 110984. [Google Scholar] [CrossRef]
- Guyon, C.; Meynier, A.; De Lamballerie, M. Protein and lipid oxidation in meat: A review with emphasis on high-pressure treatments. Trends Food Sci. Technol. 2016, 50, 131–143. [Google Scholar] [CrossRef]
- Liu, L.; Li, S.S.; Zheng, J.X.; Bu, T.T.; He, G.Q.; Wu, J.P. Safety considerations on food protein-derived bioactive peptides. Trends Food Sci. Technol. 2020, 96, 199–207. [Google Scholar] [CrossRef]
- Han, Q.; Kong, B.H.; Chen, Q.; Sun, F.D.; Zhang, H. In vitro comparison of probiotic properties of lactic acid bacteria isolated from Harbin dry sausages and selected probiotics. J. Funct. Foods 2017, 32, 391–400. [Google Scholar] [CrossRef]
- Wang, J.; Hou, J.N.; Zhang, X.; Hu, J.R.; Yu, Z.H.; Zhu, Y.C. Improving the flavor of fermented sausage by increasing its bacterial quality via inoculation with lactobacillus plantarum MSZ2 and staphylococcus xylosus YCC3. Foods 2022, 11, 736. [Google Scholar] [CrossRef] [PubMed]
- Arnau, J.; Guerrero, L.; Sárraga, C. The effect of green ham pH and NaCl concentration on cathepsin activities and the sensory characteristics of dry-cured hams. J. Sci. Food Agric. 1998, 77, 387–392. [Google Scholar] [CrossRef]
- Zhao, G.M.; Zhou, G.H.; Wang, Y.L.; Xu, X.L.; Huan, Y.J.; Wu, J.Q. Time-related changes in cathepsin B and L activities during processing of Jinhua ham as a function of pH, salt and temperature. Meat Sci. 2005, 70, 381–388. [Google Scholar] [CrossRef]
- Berardo, A.; Devreese, B.; Maere, H.D.; Stavropoulou, D.A.; Royen, G.V.; Leroy, F.; Smet, S.D. Actin proteolysis during ripening of dry fermented sausages at different pH values. Food Chem. 2017, 221, 1322–1332. [Google Scholar] [CrossRef]
- Hu, Y.Y.; Li, Y.J.; Zhu, J.M.; Kong, B.H.; Liu, Q.; Chen, Q. Improving the taste profile of reduced-salt dry sausage by inoculating different lactic acid bacteria. Food Res. Int. 2021, 145, 110391. [Google Scholar] [CrossRef] [PubMed]
- Fadda, S.; Vildoza, M.J.; Vignolo, G. The acidogenic metabolism of lactobacillus plantarum crl 681 improves sarcoplasmic protein hydrolysis during meat fermentation. J. Muscle Foods 2010, 21, 545–556. [Google Scholar] [CrossRef]
- Casaburi, A.; Monaco, R.D.; Cavella, S.; Toldrá, F.; Ercolini, D.; Villani, F. Proteolytic and lipolytic starter cultures and their effect on traditional fermented sausages ripening and sensory traits. Food Microbiol. 2008, 25, 335–347. [Google Scholar] [CrossRef]
- Siedler, S.; Balti, R.; Neves, A.R. Bioprotective mechanisms of lactic acid bacteria against fungal spoilage of food. Curr. Opin. Biotechnol. 2019, 56, 138–146. [Google Scholar] [CrossRef]
- Yang, F.; Xia, W.-S.; Zhang, X.-W.; Xu, Y.-S.; Jiang, Q.-X. A comparison of endogenous and microbial proteolytic activities during fast fermentation of silver carp inoculated with Lactobacillus plantarum. Food Chem. 2016, 207, 86–92. [Google Scholar] [CrossRef]
- Sun, F.D.; Hu, Y.Y.; Chen, Q.; Kong, B.H.; Liu, Q. Purification and biochemical characteristics of the extracellular protease from Pediococcus pentosaceus isolated from Harbin dry sausages. Meat Sci. 2019, 156, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.D.; Li, Q.X.; Liu, H.T.; Kong, B.H.; Liu, Q. Purification and biochemical characteristics of the protease from Lactobacillus brevis R4 isolated from Harbin dry sausages. LWT-Food Sci. Technol. 2019, 113, 108287. [Google Scholar] [CrossRef]
- Sun, F.D.; Sun, Q.X.; Zhang, H.; Kong, B.H.; Xia, X.F. Purification and biochemical characteristics of the microbial extracellular protease from Lactobacillus curvatus isolated from Harbin dry sausages. Int. J. Biol. Macromol. 2019, 133, 987–997. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.D.; Tao, R.; Liu, Q.; Wang, H.; Kong, B.H. Effects of temperature and pH on the structure of a metalloprotease from Lactobacillus fermentum R6 isolated from Harbin dry sausages and molecular docking between protease and meat protein. J. Sci. Food Agric. 2021, 101, 5016–5027. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.D.; Hu, Y.Y.; Yin, X.Y.; Kong, B.H.; Qin, L.G. Production, purification and biochemical characterization of the microbial protease produced by Lactobacillus fermentum R6 isolated from Harbin dry sausages. Process Biochem. 2020, 89, 37–45. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.P.; Wang, Y.Y.; Xu, H.; Mei, X.Q.; Yu, D.Y.; Wang, Y.B.; Li, W.F. Antioxidant properties of probiotic bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef]
- Lee, J.; Hwang, K.T.; Chung, M.Y.; Cho, D.H.; Park, C.S. Resistance of lactobacillus casei KCTC 3260 to Reactive Oxygen Species (ROS): Role for a metal ion chelating effect. J. Food Sci. 2005, 70, 388–391. [Google Scholar] [CrossRef]
- Liu, R.; Li, S.Y.; Yang, B.; Chen, L.; Ge, Q.F.; Xiong, G.Y.; Yu, H.; Wu, M.A.; Zhang, W.A. Investigation of the antioxidant capacity of cell-free extracts from Lactobacillus plantarum NJAU-01 obtained by different cell disruption methods. LWT-Food Sci. Technol. 2021, 152, 112393. [Google Scholar] [CrossRef]
- Amaretti, A.; Di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef]
- Talwalkar, A.; Kailasapathy, K. Metabolic and biochemical responses of probiotic bacteria to oxygen. J. Dairy Sci. 2003, 86, 2537–2546. [Google Scholar] [CrossRef]
- Landis, G.N.; Tower, J. Superoxide dismutase evolution and life span regulation. Mech. Ageing Dev. 2005, 126, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Guzmán, M.; Toral, M.; Romero, M.; Jiménez, R.; Galindo, P.; Sánchez, M.; Zarzuelo, M.J.; Olivares, M.; Gálvez, J.; Duarte, J. Antihypertensive effects of probiotics Lactobacillus strains in spontaneously hypertensive rats. Mol. Nutr. Food Res. 2015, 59, 2326–2336. [Google Scholar] [CrossRef] [PubMed]
- Carmel-Harel, O.; Storz, G. Roles of the glutathione- and thioredoxin-dependent reduction systems in the Escherichia coli and Saccharomyces cerevisiae responses to oxidative stress. Annu. Rev. Microbiol. 2000, 54, 439–461. [Google Scholar] [CrossRef]
- Arner, E.; Holmgren, A. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Toalá, J.E.; Estrada-Montoya, M.C.; Liceaga, A.M.; Garcia, H.S.; González-Aguilar, G.A.; Vallejo-Cordoba, B.; González-Córdova, A.F.; Hernández-Mendoza, A. An insight on antioxidant properties of the intracellular content of Lactobacillus casei CRL-431. LWT-Food Sci. Technol. 2019, 102, 58–63. [Google Scholar] [CrossRef]
- Li, S.Y.; Zhao, Y.J.; Zhang, L.; Zhang, X.; Huang, L.; Li, D.; Niu, C.H.; Yang, Z.N.; Wang, Q. Antioxidant activity of Lactobacillus plantarum strains isolated from traditional Chinese fermented foods. Food Chem. 2012, 135, 1914–1919. [Google Scholar] [CrossRef]
- Bleau, C.; Monges, A.; Rashidan, K.; Laverdure, J.-P.; Lacroix, M.; Van Calsteren, M.-R.; Millette, M.; Savard, R.; Lamontagne, L. Intermediate chains of exopolysaccharides from Lactobacillus rhamnosus RW-9595M increase IL-10 production by macrophages. J. Appl. Microbiol. 2010, 108, 666–675. [Google Scholar] [CrossRef]
- Aarti, C.; Khusro, A. Functional and technological properties of exopolysaccharide producing autochthonous Lactobacillus plantarum strain AAS3 from dry fish based fermented food. LWT-Food Sci. Technol. 2019, 114, 108387. [Google Scholar] [CrossRef]
- Lee, Y.G.; Cho, J.Y.; Moon, J.H. Isolation and antioxidative activity of amino acid derivatives produced by Leuconostoc mesenteroides. Food Sci. Biotechnol. 2016, 25, 329–334. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, C.; Liu, H.T.; Chen, Q.; Kong, B.H. Proteomic response strategies of Pediococcus pentosaceus R1 isolated from Harbin dry sausages to oxidative stress. Food Biosci. 2021, 44, 101364. [Google Scholar] [CrossRef]
- Huang, L.; Zeng, X.Q.; Sun, Z.; Wu, A.J.; He, J.; Dang, Y.L.; Pan, D.D. Production of a safe cured meat with low residual nitrite using nitrite substitutes. Meat Sci. 2020, 162, 108027. [Google Scholar] [CrossRef] [PubMed]
- Lian, Z.Y.; Li, Y.Q. Isolation of antibacterial, nitrosylmyoglobin forming lactic acid bacteria and their potential use in meat processing. Front. Microbiol. 2020, 11, 1315. [Google Scholar] [CrossRef]
- Cao, K.H.; Zhang, K.P.; Ma, M.R.; Ma, J.J.; Tian, J.J.; Jin, Y. Lactobacillus mediates the expression of NPC1L1, CYP7A1, and ABCG5 genes to regulate cholesterol. Food Sci. Nutr. 2021, 9, 6882–6891. [Google Scholar] [CrossRef]
- Li, J.Z.; Li, Q.K.; Gao, N.; Wang, Z.H.; Li, F.; Li, J.P.; Shan, A.S. Exopolysaccharides produced by Lactobacillus rhamnosus GG alleviate hydrogen peroxide-induced intestinal oxidative damage and apoptosis through the Keap1/Nrf2 and Bax/Bcl-2 pathways in vitro. Food Funct. 2021, 12, 9632–9641. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.Y.; Chan, B.D.; Sham, T.T.; Lee, M.M.L.; Chan, C.O.; Chau, C.T.; Mok, D.K.W.; Kwan, Y.W.; Tai, W.C.S. Lactobacillus casei strain shirota ameliorates dextran sulfate sodium-induced colitis in mice by increasing taurine-conjugated bile acids and inhibiting NF-κB signaling via stabilization of Iκ B α. Front. Nutr. 2022, 9, 816836. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Liu, Y.L.; Ai, Z.Y.; Wang, Y.S.; Liu, J.M.; Piao, C.H.; Wang, Y.H. Lactobacillus fermentum KP-3-fermented ginseng ameliorates alcohol-induced liver disease in C57BL/6N mice through the AMPK and MAPK pathways. Food Funct. 2020, 11, 9801–9809. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Peptide Sequence | Length | Mass (Da) | Protein of Origin | References |
|---|---|---|---|---|---|
| Fermented sausage | LPVKY | 5 | 618.3741 | Phosphatidylinositol 3-kinase catalytic subunit type 3 | [22] |
| PFGDTH | 6 | 672.2867 | CDP-6-deoxy-delta-3,4-glucoseen reductase | ||
| QPSLVH | 6 | 679.3653 | Aminoglycoside transferase | ||
| LPVTVR | 6 | 683.433 | Rho guanine nucleotide exchange factor 2 | ||
| LPSLKF | 6 | 703.4268 | Phosphoglycerate kinase | ||
| HKLPVK | 6 | 720.4646 | Myoglobin | ||
| YGLDEK | 6 | 723.3439 | Serine/threonine-protein kinase A | ||
| LDRKDL | 6 | 758.4286 | Lipid A biosynthesis lauroyltransferase | ||
| LDLRDK | 6 | 758.4286 | Tetratricopeptide repeat protein 27 | ||
| EGGLSKLTLDKLDVKGK | 17 | 1800.036 | Phosphoglycerate kinas | ||
| EARSKLTLDKLDVKGK | 16 | 1800.047 | Phosphoglycerate kinase | ||
| ESDLLAAFR | 9 | 1020.524 | Adenosine kinase | ||
| EVSHGSDQVKAHGQK | 15 | 1605.786 | Hemoglobin subunit alpha | ||
| ELSNKLTLDK | 10 | 1159.645 | Phosphoglycerate kinase | ||
| VDLNGGSHAGNK | 12 | 1167.563 | Beta-enolase isoform X1 | ||
| EGADSEMALFGEAAPYLRKSEKEVGKN | 27 | 2925.418 | Myosin-1 isoform X1 | ||
| NPPKF | 5 | 601.3224 | Myosin | ||
| KGGSLELTLDKLDVKGK | 17 | 1800.036 | Phosphoglycerate kinase | ||
| Mutton Ham | MWTD | 4 | 551.61 | Uncharacterized protein | [23] |
| SAGNPN | 6 | 558.20 | Uncharacterized protein | ||
| APYMM | 5 | 611.76 | Uncharacterized protein | ||
| VFDPEG | 6 | 662.69 | Myosin-2 | ||
| FWIIE | 5 | 706.84 | Spectrin alpha chain-like protein | ||
| DKEEFV | 6 | 744.81 | Myosin-7 | ||
| GVDNPGHP | 8 | 791.81 | Creatine Kinase M-type | ||
| MVHMASK | 7 | 803.00 | Glyceraldehyde-3-phosphate dehydrogenase | ||
| PAPAPPKE | 8 | 805.91 | Titin | ||
| FGNTHNK | 7 | 816.87 | Creatine Kinase M-type | ||
| MDAIKKK | 7 | 833.05 | Tropomyosin | ||
| IEEALGDK | 8 | 873.94 | Beta-enolase | ||
| IPPKIPEGE | 9 | 887.01 | Troponin T | ||
| GLRKHER | 7 | 895.02 | Myosin-4 | ||
| ERFSKDE | 7 | 909.93 | Uncharacterized protein | ||
| ERSFKDE | 7 | 909.93 | Troponin T | ||
| PFGNTHNK | 8 | 913.99 | Creatine Kinase M-type | ||
| NVINGGSHAG | 10 | 924.97 | Glyceraldehyde-3-phosphate dehydrogenase | ||
| HIITHGEE | 8 | 935.02 | Myosin regulatory light chain 2 | ||
| GRKFRNPK | 8 | 1002.18 | Beta-enolase | ||
| DVAGHGQEVL | 10 | 1024.10 | Myosin regulatory light chain 2 | ||
| IDDMIPAQK | 9 | 1030.20 | Creatine Kinase M-type | ||
| Jinhua Ham | GKKFNV | 6 | 565.28 | Transcription activator BRGl | [23] |
| LVVDGVK | 7 | 728.88 | Uncharacterized protein | ||
| DKEEFV | 6 | 744.81 | Myosin-7 | ||
| VDIINAK | 7 | 772.45 | Uncharacterized protein | ||
| VHMASKE | 7 | 801.39 | Glyceraldehyde-3-phosphate dehydrogenase | ||
| MDAIKKK | 7 | 833.05 | Tropomyosin | ||
| LVVDGVKL | 8 | 841.53 | Creatine Kinase M-type | ||
| IPPKIPEGE | 9 | 887.01 | Troponin T | ||
| KAGTTPKGK | 9 | 887.04 | Maturase K | ||
| YGEKLKR | 7 | 893.04 | Binding protein | ||
| ALPHAIMR | 8 | 908.11 | Actin | ||
| PFGNTHNK | 8 | 913.99 | Creatine Kinase M-type | ||
| VKQKGPDF | 8 | 918.50 | Pyruvate kinase | ||
| AGQAFRKF | 8 | 924.05 | Uncharacterized protein | ||
| MVHMASKE | 8 | 932.43 | Glyceraldehyde-3-phosphate dehydrogenase | ||
| IEEALGDKA | 9 | 945.02 | Beta-enolase | ||
| VITHGDAKD | 9 | 955.48 | Myosin regulatory light chain 2 | ||
| LQNHPEHS | 8 | 960.99 | Glyceraldehyde-3-phosphate dehydrogenase | ||
| EAGPSIVHR | 9 | 965.07 | Actin | ||
| LRDKAKEL | 8 | 972.12 | Binding protein | ||
| AGFAGDDAPR | 10 | 975.99 | Actin | ||
| FPMNPPKF | 8 | 977.49 | Myosin | ||
| DVGDWRKN | 8 | 989.05 | Troponin T | ||
| VAPEEHPTL | 9 | 992.08 | Actin | ||
| KKAGTTPKGK | 10 | 1015.21 | Troponin T |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Cheng, F.; Wang, Y.; Han, J.; Gao, F.; Tian, J.; Zhang, K.; Jin, Y. The Changes Occurring in Proteins during Processing and Storage of Fermented Meat Products and Their Regulation by Lactic Acid Bacteria. Foods 2022, 11, 2427. https://doi.org/10.3390/foods11162427
Wang D, Cheng F, Wang Y, Han J, Gao F, Tian J, Zhang K, Jin Y. The Changes Occurring in Proteins during Processing and Storage of Fermented Meat Products and Their Regulation by Lactic Acid Bacteria. Foods. 2022; 11(16):2427. https://doi.org/10.3390/foods11162427
Chicago/Turabian StyleWang, Daixun, Feng Cheng, Yi Wang, Jun Han, Fang Gao, Jianjun Tian, Kaiping Zhang, and Ye Jin. 2022. "The Changes Occurring in Proteins during Processing and Storage of Fermented Meat Products and Their Regulation by Lactic Acid Bacteria" Foods 11, no. 16: 2427. https://doi.org/10.3390/foods11162427