
Molecular Link in Flavonoid and Amino Acid Biosynthesis Contributes to the Flavor of Changqing Tea in Different Seasons

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Detection of Flavor Components of Changqing Tea
2.3. RNA Extraction and Illumina Sequencing
2.4. Genome Alignment and Gene Prediction
2.5. Differential Expression Analysis
2.6. Quantitative Real-Time PCR Analysis (QRT-PCR)
2.7. Metabolites Extraction and Detection
2.8. Statistical Analysis
3. Results
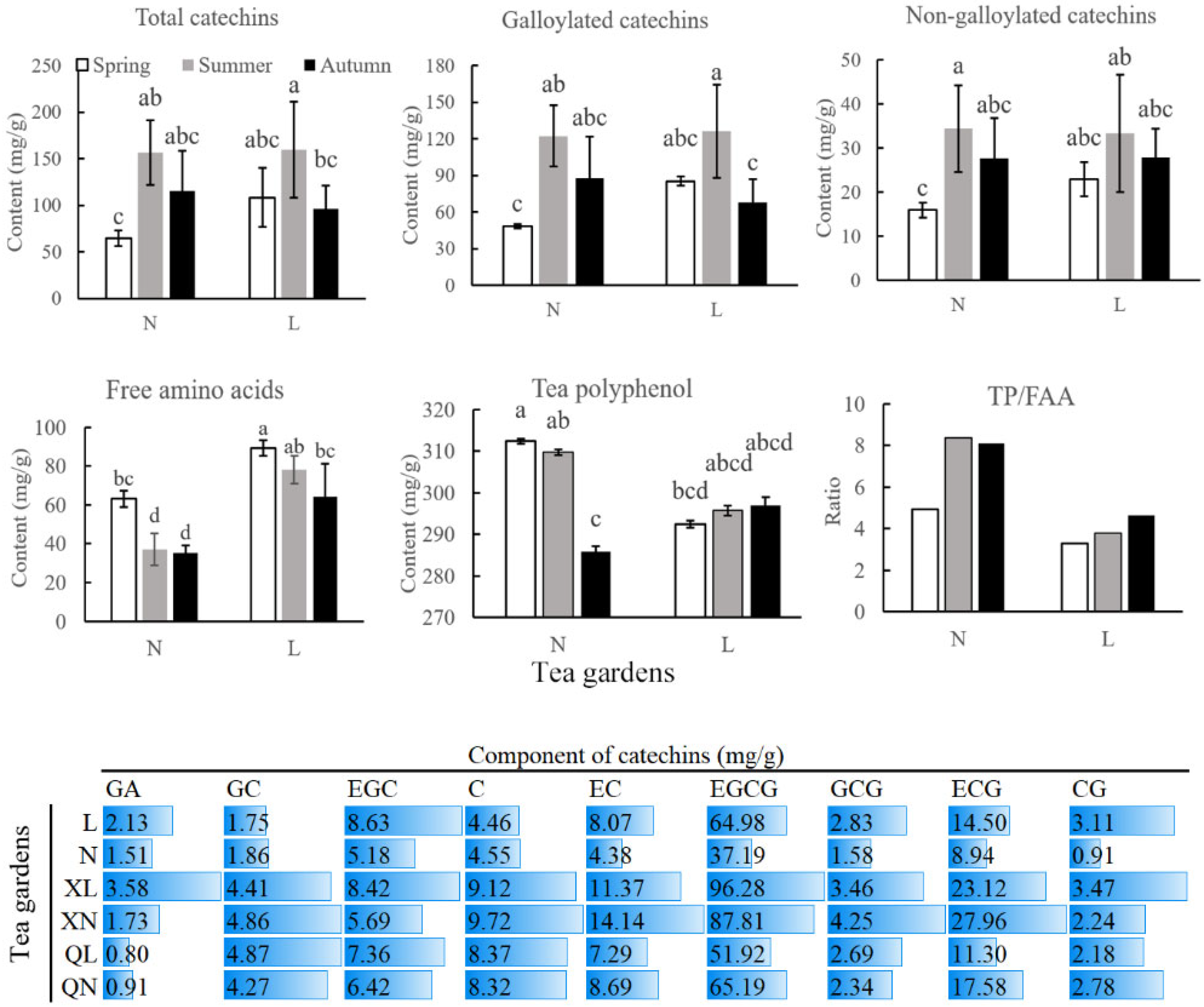
3.1. Flavor Components of Changqing Tea
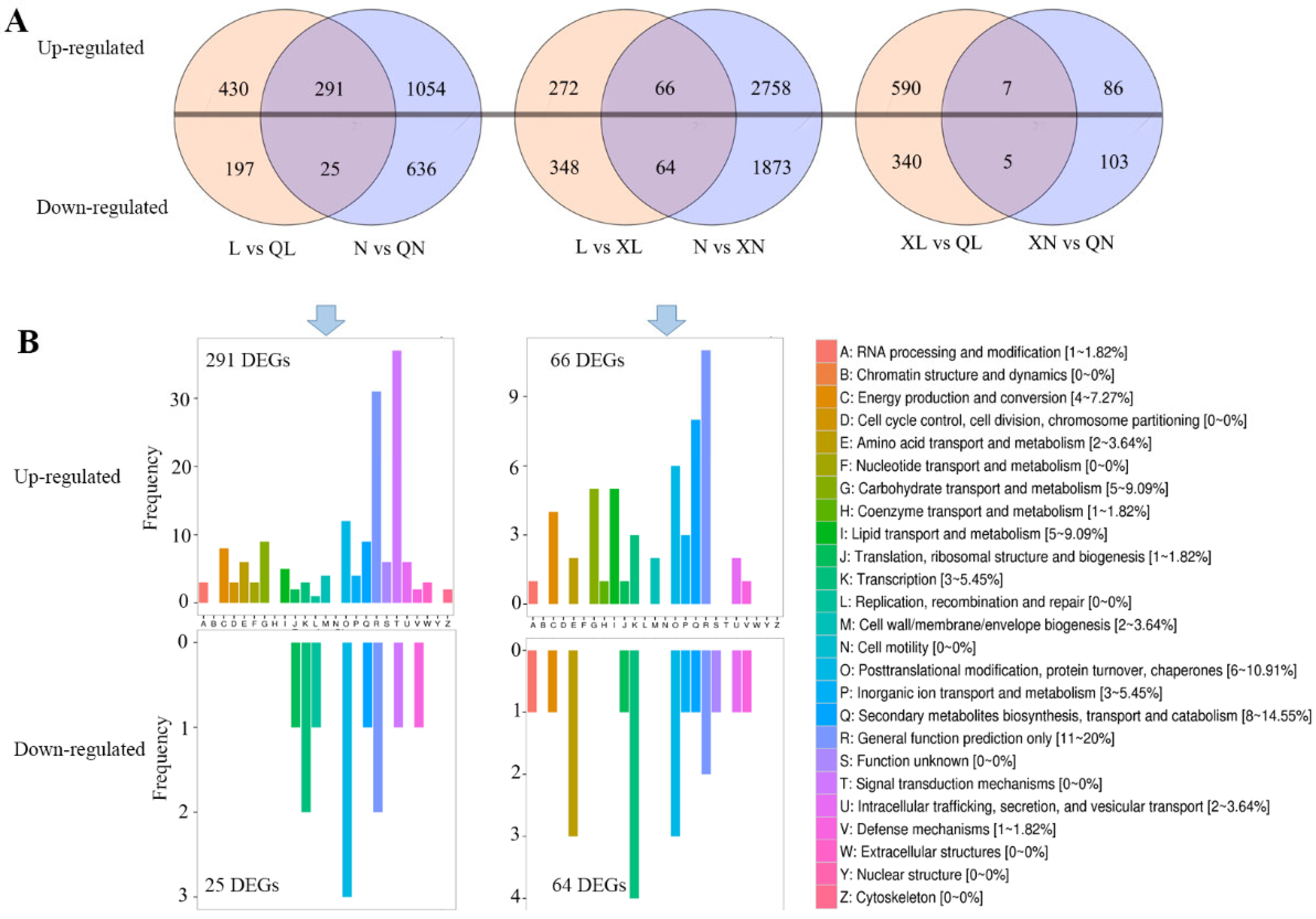
3.2. Identification of Differential Expressed Genes
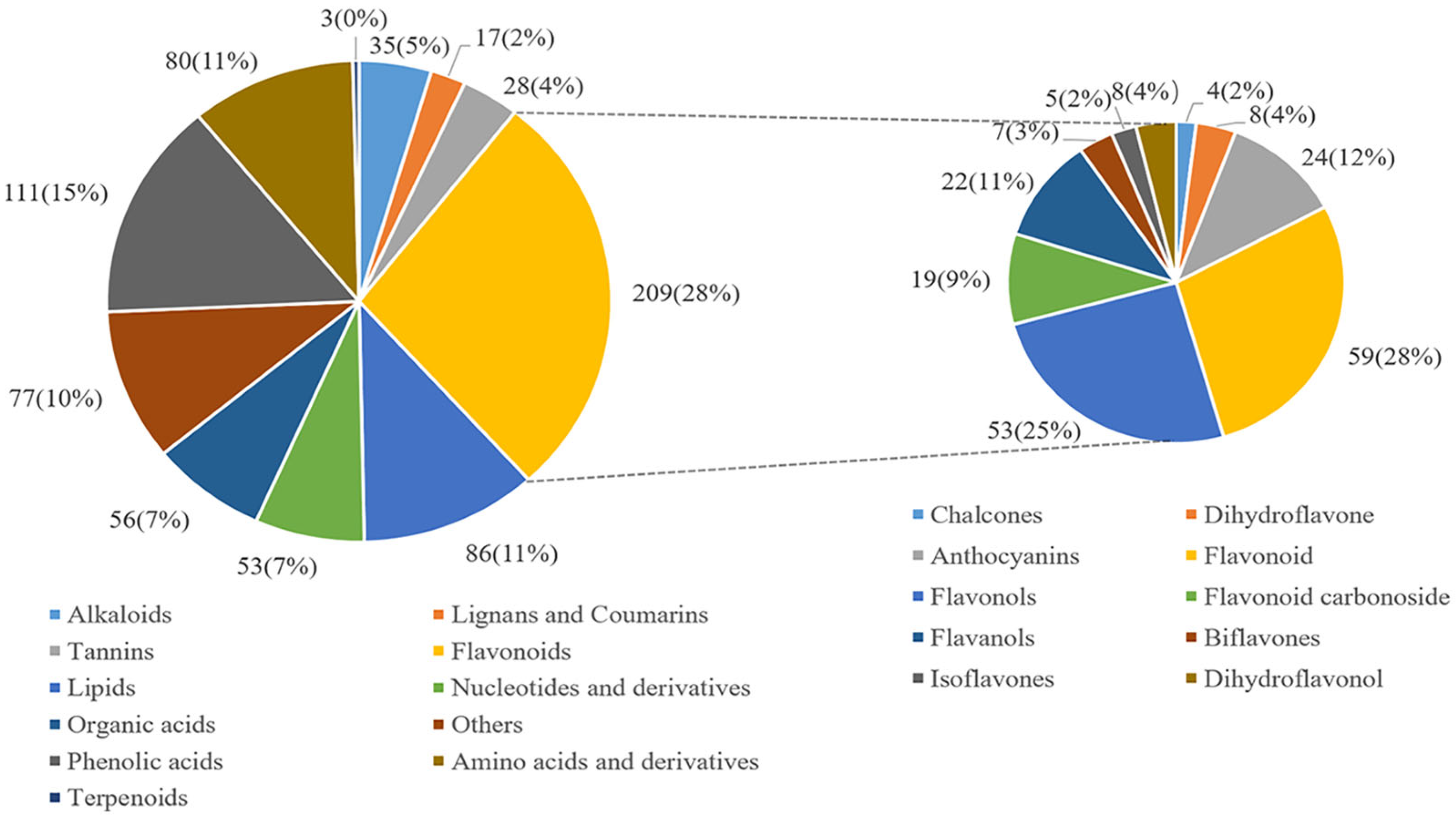
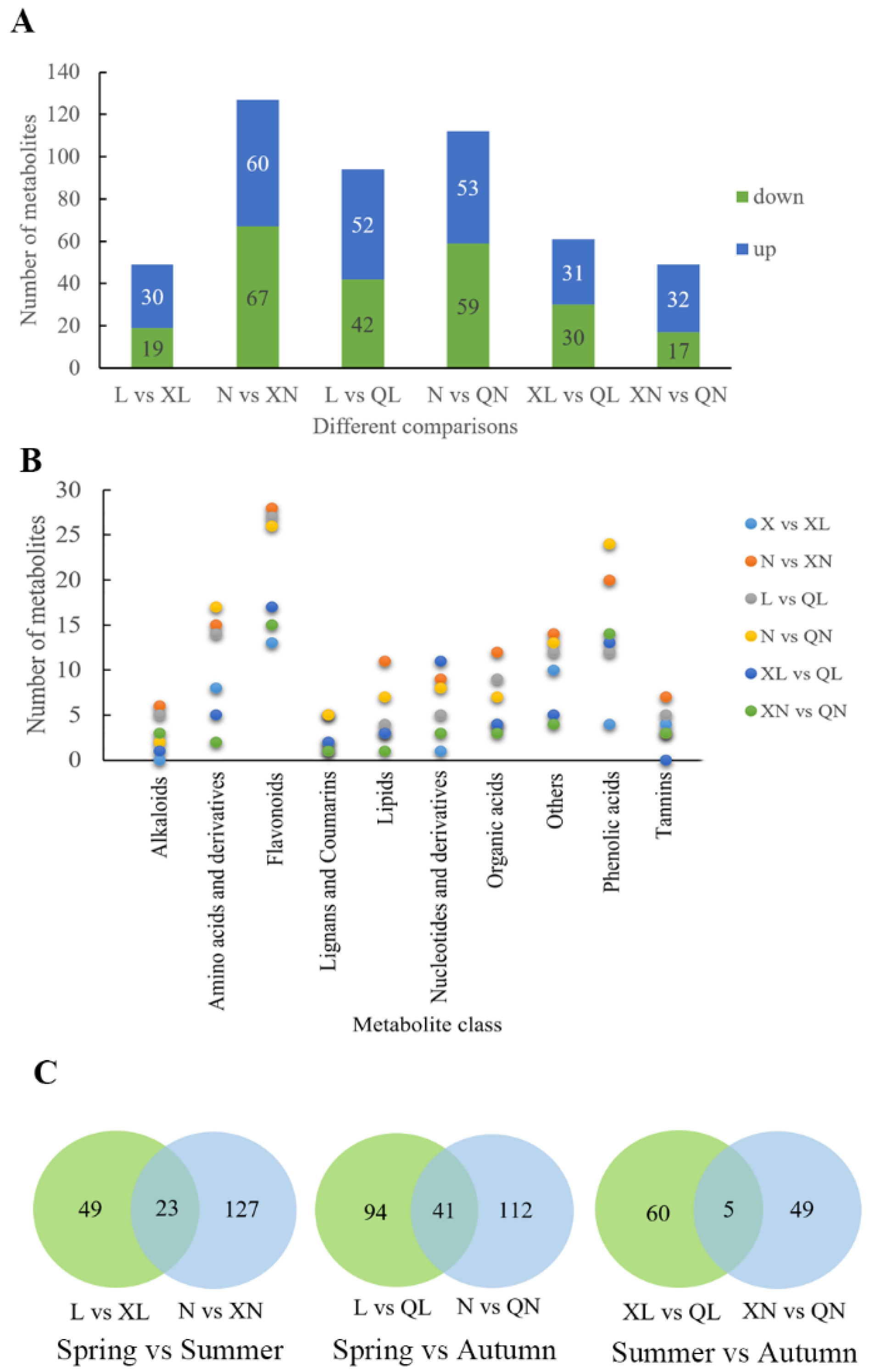
3.3. Identification of Differential Metabolites
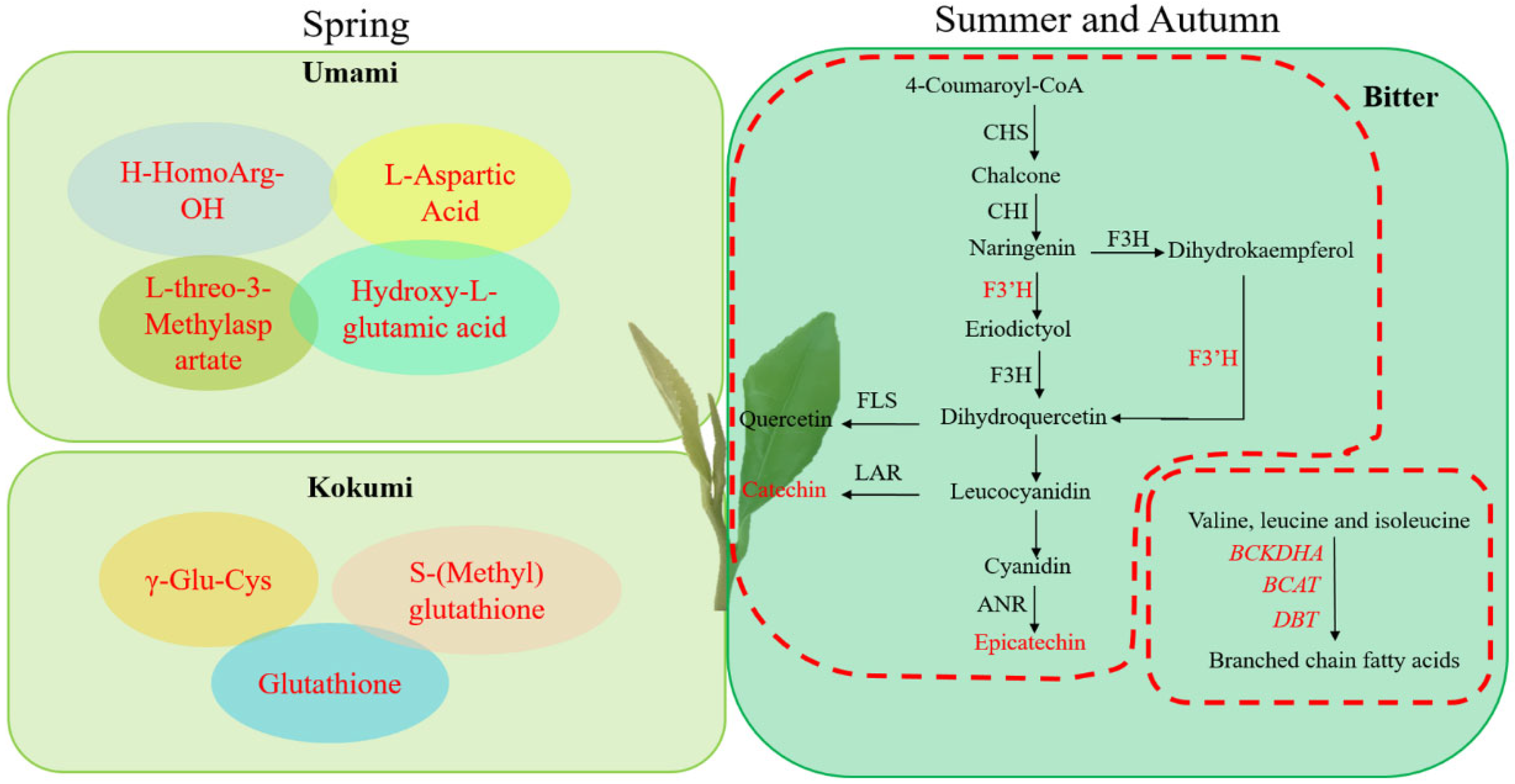
3.4. Flavonoid Biosynthesis of Changqing Tea in Different Seasons
3.5. Amino Acid Metabolism of Changqing Tea in Different Seasons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kfoury, N.; Scott, E.R.; Orians, C.M.; Ahmed, S.; Cash, S.B.; Griffin, T.; Matyas, C.; Stepp, J.R.; Han, W.; Xue, D.; et al. Plant-climate interaction effects: Changes in the relative distribution and concentration of the volatile tea leaf metabolome in 2014–2016. Front. Plant Sci. 2019, 10, 1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Zhao, Y.; Sun, Y.; Yang, S.; Yang, X. Characterisation of polysaccharides from green tea of Huangshan Maofeng with antioxidant and hepatoprotective effects. Food Chem. 2013, 141, 3415–3423. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Guo, H.; Zong, B.; He, P.; Fan, F.; Gong, S. Rapid and non-destructive discrimination of special-grade flat green tea using Near-infrared spectroscopy. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2019, 206, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.L. Tea in China. In Caffeine; Spiller, G.A., Ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 1–6. [Google Scholar]
- Zhuang, X.G.; Shi, X.S.; Wang, H.F.; Wang, L.L.; Fang, J.X. Rapid determination of green tea origins by near-infrared spectroscopy and multi-wavelength statistical discriminant analysis. J. Appl. Spectrosc. 2019, 86, 76–82. [Google Scholar] [CrossRef]
- Zhuang, X.-G.; Wang, L.-L.; Wu, X.-Y.; Fang, J.-X. Origin identification of Shandong green tea by moving window back propagation artificial neural network based on near infrared spectroscopy. J. Infrared Millim. Waves 2016, 35, 200–205. [Google Scholar]
- Ma, Q.; Song, L.; Niu, Z.; Li, J.; Wang, Y.; Sun, H.; Ren, Z.; Zhao, H.; Guo, S.; Ding, Z. Red light regulates the metabolite biosynthesis in the leaves of “Huangjinya” through amino acid and phenylpropanoid metabolisms. Front. Plant Sci. 2022, 12, 810888. [Google Scholar] [CrossRef]
- Xia, E.; Tong, W.; Hou, Y.; An, Y.; Chen, L.; Wu, Q.; Liu, Y.; Yu, J.; Li, F.; Li, R.; et al. The reference genome of tea plant and resequencing of 81 diverse accessions provide insights into its genome evolution and adaptation. Mol. Plant 2020, 13, 1013–1026. [Google Scholar] [CrossRef]
- Deng, Y.; Jianqi, L.I.; Songfeng, W.U.; Zhu, Y.; Chen, Y.; Fuchu, H.E. Integrated nr database in protein annotation system and its localization. Comput. Eng. 2006, 32, 71–74. [Google Scholar]
- Apweiler, R.; Bairoch, A.; Wu, C.H.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; Magrane, M.; et al. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2004, 32, D115–D119. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Krylov, D.M.; Makarova, K.S.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; Rao, B.S.; et al. A comprehensive evolutionary classification of proteins encoded in complete eukaryotic genomes. Genome Biol. 2004, 5, R7. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ma, Q.; Li, H.; Zou, Z.; Arkorful, E.; Lv, Q.; Zhou, Q.; Chen, X.; Sun, K.; Li, X. Transcriptomic analyses identify albino-associated genes of a novel albino tea germplasm ‘Huabai 1’. Hortic. Res. 2018, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.-K.; Ni, D.-J.; Yao, M.-Z.; Ma, J.-Q.; Chen, L. Metabolic and transcriptome analysis reveals metabolite variation and flavonoid regulatory networks in fresh shoots of tea (Camellia sinensis) over three seasons. Front. Agric. Sci. Eng. 2021, 8, 215–230. [Google Scholar]
- Djuardi, A.U.P.; Yuliana, N.D.; Ogawa, M.; Akazawa, T.; Suhartono, M.T. Emulsifying properties and antioxidant activity of soy protein isolate conjugated with tea polyphenol extracts. J. Food Sci. Technol. 2020, 57, 3591–3600. [Google Scholar] [CrossRef]
- Kim, H.-S.; Quon, M.J.; Kim, J.-A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Zhou, Y.; Chen, F.; Yan, X.; Lai, Y.; Wei, C.; Chen, X.; Xu, J.; Wang, X. Isolation, diversity, and antimicrobial and immunomodulatory activities of endophytic actinobacteria from tea cultivars Zijuan and Yunkang-10 (Camellia sinensis var. assamica). Front. Microbiol. 2018, 9, 1304. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhang, T. Antimicrobial activities of tea polyphenol on phytopathogens: A review. Molecules 2019, 24, 816. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Xu, S.; Fang, J.; Jiang, H. The protective effect of polyphenols for colorectal cancer. Front. Immunol. 2020, 11, 1407. [Google Scholar] [CrossRef]
- Lecumberri, E.; Dupertuis, Y.M.; Miralbell, R.; Pichard, C. Green tea polyphenol epigallocatechin-3-gallate (EGCG) as adjuvant in cancer therapy. Clin. Nutr. 2013, 32, 894–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.Y.; Liang, Y.R.; Wang, H.; Wang, K.R.; Lu, J.L.; Zhang, G.H.; Lin, W.P.; Li, M.; Fang, Q.Y. A study on the chemical composition of albino tea cultivars. J. Hortic. Sci. Biotechnol. 2006, 81, 809–812. [Google Scholar] [CrossRef]
- Feng, L.; Gao, M.-J.; Hou, R.-Y.; Hu, X.-Y.; Zhang, L.; Wan, X.-C.; Wei, S. Determination of quality constituents in the young leaves of albino tea cultivars. Food Chem. 2014, 155, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Li, J.; Li, Y.; Yuan, L.; Liu, S.; Huang, J.A.; Liu, Z. Dynamic changes in catechin levels and catechin biosynthesis-related gene expression in albino tea plants (Camellia sinensis L.). Plant Physiol. Biochem. 2013, 71, 132–143. [Google Scholar] [CrossRef]
- Kerio, L.C.; Wachira, F.N.; Wanyoko, J.K.; Rotich, M.K. Total polyphenols, catechin profiles and antioxidant activity of tea products from purple leaf coloured tea cultivars. Food Chem. 2013, 136, 1405–1413. [Google Scholar] [CrossRef]
- Liu, J.; Liu, M.; Fang, H.; Zhang, Q.; Ruan, J. Accumulation of amino acids and flavonoids in young tea shoots is highly correlated with carbon and nitrogen metabolism in roots and mature leaves. Front. Plant Sci. 2021, 12, 756433. [Google Scholar] [CrossRef]
- Shen, Z.; He, Y.; Li, Y.; Yang, T.; Xu, X.; Jiang, C. Insights into the profiling changes of amino acid content in an albino mutant (Camellia sinensis cv. Huangshanbaicha) during the albinostic stage. Sci. Hortic. 2020, 260, 108732. [Google Scholar] [CrossRef]
- Zhao, C.J.; Schieber, A.; Gänzle, M.G. Formation of taste-active amino acids, amino acid derivatives and peptides in food fermentations—A review. Food Res. Int. 2016, 89, 39–47. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Total Reads | GC Content | Q30 | Mapped Reads | Uniq Mapped Reads |
|---|---|---|---|---|---|
| L1 | 44,484,588 | 45.35% | 94.84% | 38,627,832 (86.83%) | 32,337,944 (72.69%) |
| L2 | 46,058,340 | 45.37% | 94.89% | 40,035,790 (86.92%) | 33,536,472 (72.81%) |
| L3 | 38,975,414 | 45.72% | 94.49% | 33,881,495 (86.93%) | 28,279,355 (72.56%) |
| N1 | 44,248,808 | 45.38% | 94.45% | 38,350,376 (86.67%) | 32,204,533 (72.78%) |
| N2 | 42,729,226 | 45.52% | 94.51% | 36,935,621 (86.44%) | 30,977,168 (72.50%) |
| N3 | 39,548,916 | 45.51% | 94.20% | 34,339,197 (86.83%) | 28,631,767 (72.40%) |
| XL1 | 43,648,802 | 45.51% | 94.78% | 37,879,990 (86.78%) | 31,889,023 (73.06%) |
| XL2 | 43,559,986 | 45.67% | 94.84% | 37,926,766 (87.07%) | 32,015,677 (73.50%) |
| XL3 | 43,038,378 | 45.49% | 94.82% | 37,442,835 (87.00%) | 31,632,998 (73.50%) |
| XN1 | 43,957,652 | 45.58% | 94.51% | 38,161,588 (86.81%) | 32,217,373 (73.29%) |
| XN2 | 41,767,546 | 45.29% | 93.91% | 36,196,630 (86.66%) | 30,599,340 (73.26%) |
| XN3 | 43,149,206 | 45.43% | 94.94% | 37,390,956 (86.66%) | 31,694,135 (73.45%) |
| QL1 | 48,320,578 | 45.43% | 95.29% | 41,721,506 (86.34%) | 35,084,628 (72.61%) |
| QL2 | 53,669,856 | 45.58% | 94.97% | 46,314,442 (86.30%) | 38,931,575 (72.54%) |
| QL3 | 52,343,966 | 45.51% | 94.32% | 44,353,491 (84.73%) | 37,160,579 (70.99%) |
| QN1 | 41,956,338 | 45.65% | 94.44% | 36,024,372 (85.86%) | 30,405,207 (72.47%) |
| QN2 | 44,766,142 | 45.29% | 94.29% | 38,612,001 (86.25%) | 32,659,135 (72.95%) |
| QN3 | 42,372,422 | 45.49% | 94.12% | 36,598,453 (86.37%) | 30,941,615 (73.02%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Qin, M.; Song, L.; Sun, H.; Zhang, H.; Wu, H.; Ren, Z.; Liu, H.; Duan, G.; Wang, Y.; et al. Molecular Link in Flavonoid and Amino Acid Biosynthesis Contributes to the Flavor of Changqing Tea in Different Seasons. Foods 2022, 11, 2289. https://doi.org/10.3390/foods11152289
Ma Q, Qin M, Song L, Sun H, Zhang H, Wu H, Ren Z, Liu H, Duan G, Wang Y, et al. Molecular Link in Flavonoid and Amino Acid Biosynthesis Contributes to the Flavor of Changqing Tea in Different Seasons. Foods. 2022; 11(15):2289. https://doi.org/10.3390/foods11152289
Chicago/Turabian StyleMa, Qingping, Mengyao Qin, Laichao Song, Haiwei Sun, Hong Zhang, Huanhuan Wu, Zhihong Ren, Hui Liu, Gang Duan, Yu Wang, and et al. 2022. "Molecular Link in Flavonoid and Amino Acid Biosynthesis Contributes to the Flavor of Changqing Tea in Different Seasons" Foods 11, no. 15: 2289. https://doi.org/10.3390/foods11152289