
Supplementation of Labneh with Passion Fruit Peel Enhanced Survival of E. coli Nissle 1917 during Simulated Gastrointestinal Digestion and Adhesion to Caco-2 Cells


 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Chemicals and Culture Media
2.1.2. Bacterial Strain and Cell Line
2.2. Preparation of PFPP
2.3. Production of Probiotic Labneh
2.4. Kinetics Parameters of Probiotic Labneh
2.5. Postacidification of Probiotic Labneh
2.6. Microbiological Analyses of Probiotic Labneh
2.7. Sensory Analyses of Probiotic Labneh
2.8. In Vitro Evaluation of EcN Survival under Conditions Simulating the GI Tract
2.9. Adhesion of EcN to Caco-2 Cells
2.10. Statistical Analysis
3. Results
3.1. The Chemical Composition of PFPP
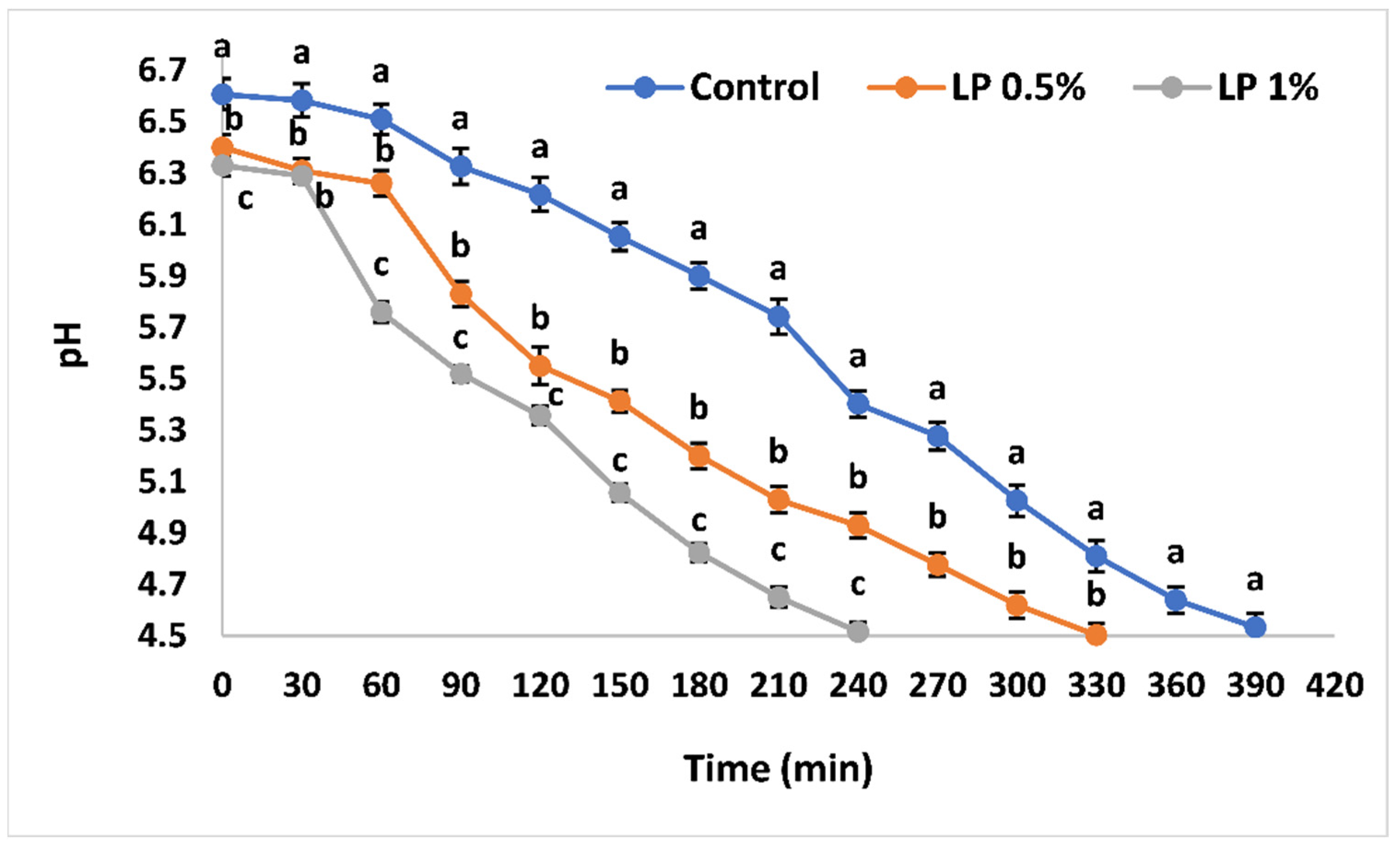
3.2. The Influence of PFPP Addition on the Fermentation Kinetics of Labneh
3.3. Postacidification and Titratable Acidity
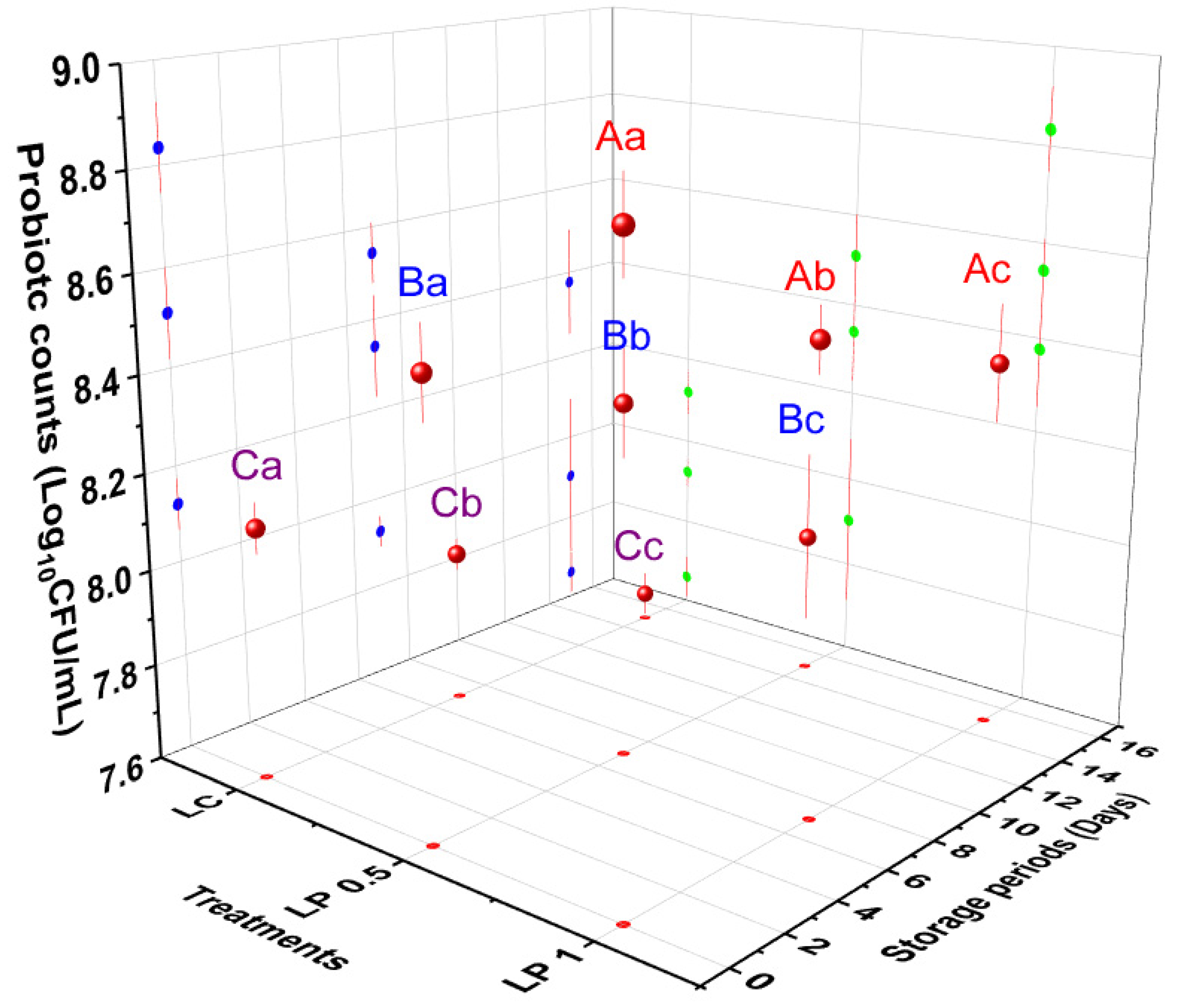
3.4. Viability of EcN during Storage of Labneh
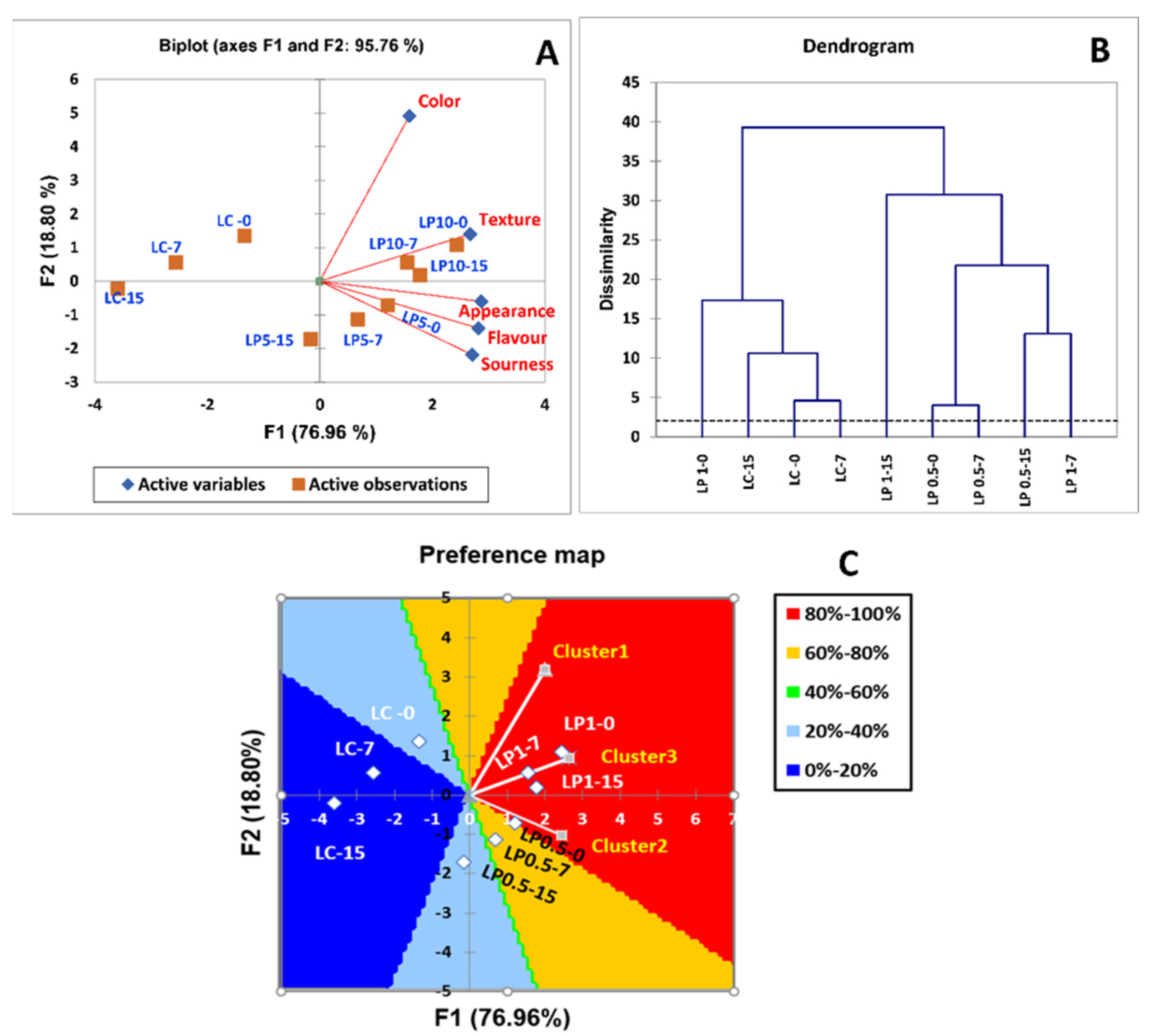
3.5. Cluster Analysis and Preference Map of Probiotic Labneh Enriched with PFPP
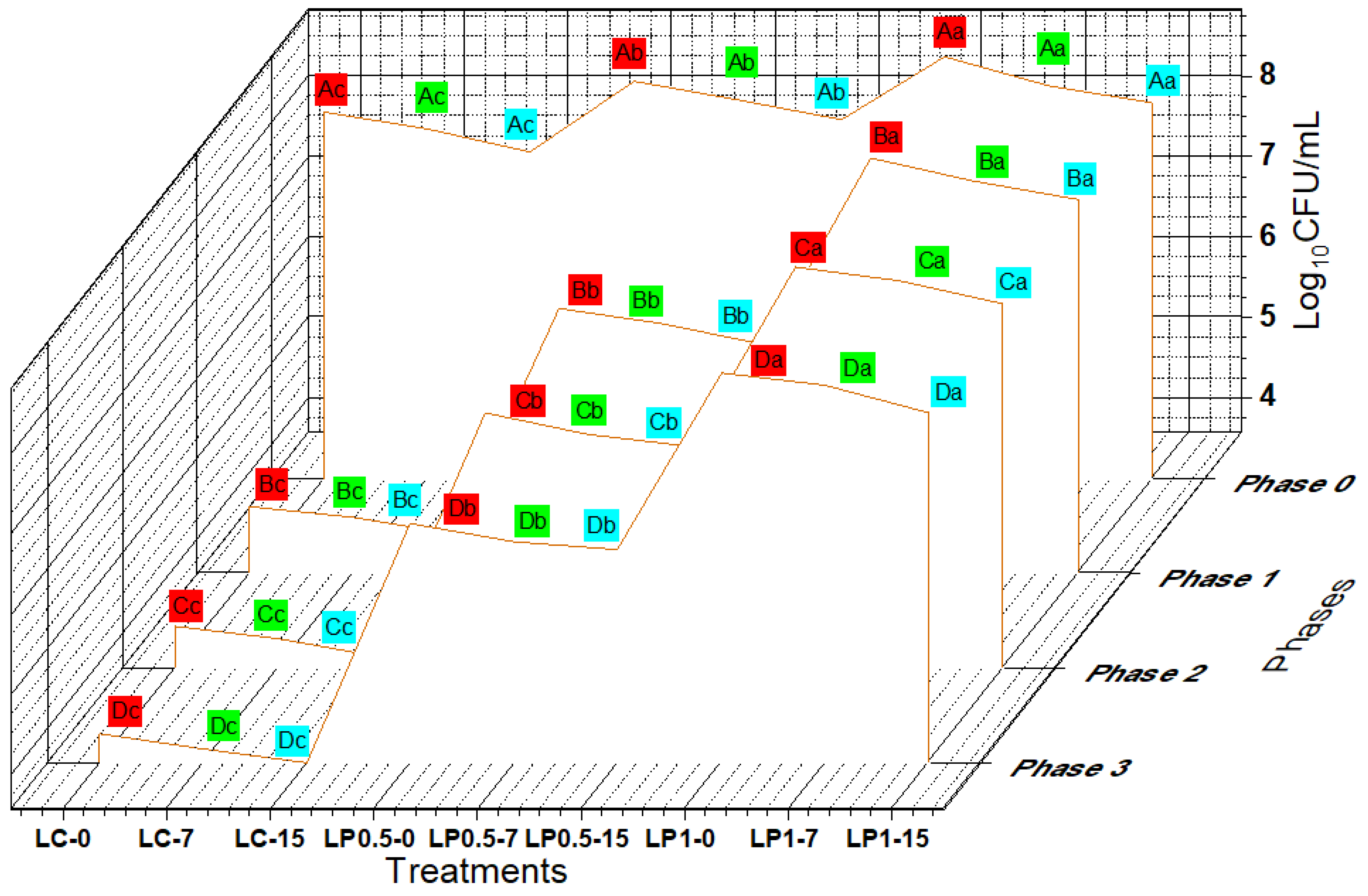
3.6. Survival of EcN against the Simulated GI Stress
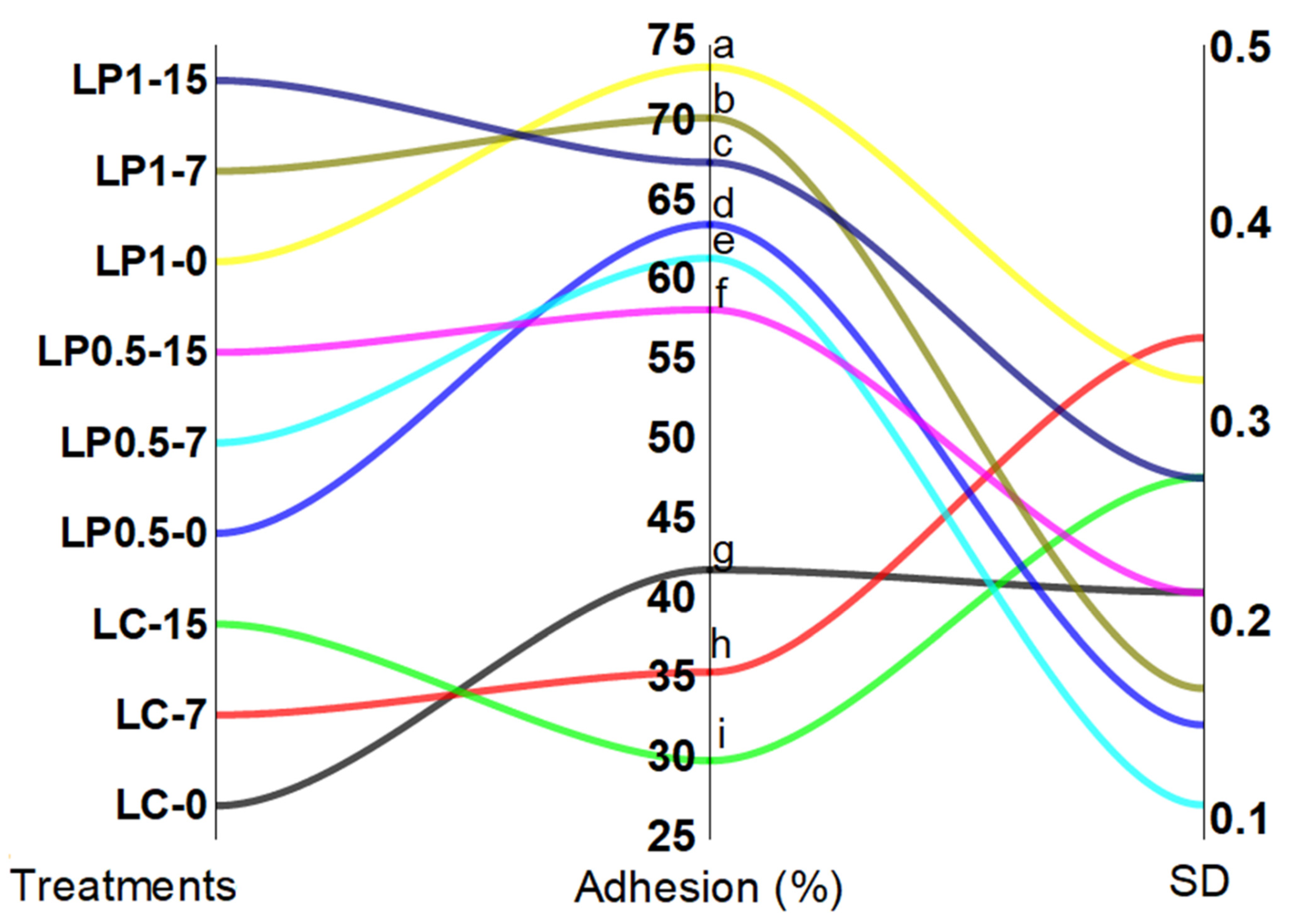
3.7. EcN Adhesion to Intestinal Cell In Vitro
3.8. Effect of Digestion on the Total Phenolic Content of PFPP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Essa, M.M.; Bishir, M.; Bhat, A.; Chidambaram, S.B.; Al-Balushi, B.; Hamdan, H.; Govindarajan, N.; Freidland, R.P.; Qoronfleh, M.W. Functional foods and their impact on health. J. Food Sci. Technol. 2021, 59, 1–15. [Google Scholar] [CrossRef]
- Damián, M.R.; Cortes-Perez, N.G.; Quintana, E.T.; Ortiz-Moreno, A.; Garfias Noguez, C.; Cruceño-Casarrubias, C.E.; Sánchez Pardo, M.E.; Bermúdez-Humarán, L.G. Functional Foods, Nutraceuticals and Probiotics: A Focus on Human Health. Microorganisms 2022, 10, 1065. [Google Scholar] [CrossRef]
- Hu, R.; Lin, H.; Li, J.; Zhao, Y.; Wang, M.; Sun, X.; Min, Y.; Gao, Y.; Yang, M. Probiotic Escherichia coli Nissle 1917-derived outer membrane vesicles enhance immunomodulation and antimicrobial activity in RAW264.7 macrophages. BMC Microbiol. 2020, 20, 268. [Google Scholar] [CrossRef]
- Grozdanov, L.; Raasch, C.; Schulze, J.; Sonnenborn, U.; Gottschalk, G.; Hacker, J.; Dobrindt, U. Analysis of the Genome Structure of the Nonpathogenic Probiotic Escherichia coli Strain Nissle 1917. J. Bacteriol. 2004, 186, 5432–5441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graziani, C.; Petito, V.; Del Chierico, F.; Mangiola, F.; Pecere, S.; Schiavoni, E.; Pizzoferrato, M.; Lopetuso, L.R.; Putignani, L.; Gasbarrini, A.; et al. P115 Escherichia coli Nissle 1917 modulate gut microbiota composition in ulcerative colitis patients. J. Crohn’s Colitis 2017, 11, S133–S134. [Google Scholar] [CrossRef]
- Toloza, L.; Giménez, R.; Fábrega, M.J.; Alvarez, C.S.; Aguilera, L.; Cañas, M.A.; Martín-Venegas, R.; Badia, J.; Baldomà, L. The secreted autotransporter toxin (Sat) does not act as a virulence factor in the probiotic Escherichia coli strain Nissle 1917. BMC Microbiol. 2015, 15, 250. [Google Scholar] [CrossRef] [Green Version]
- Jacobi, C.A.; Malfertheiner, P. Escherichia coli Nissle 1917 (Mutaflor): New Insights into an Old Probiotic Bacterium. Dig. Dis. 2011, 29, 600–607. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Chen, Z.; Shah, N.P.; El-Nezami, H. Effects of Lactobacillus rhamnosus GG and Escherichia coli Nissle 1917 Cell-Free Supernatants on Modulation of Mucin and Cytokine Secretion on Human Intestinal Epithelial HT29-MTX Cells. J. Food Sci. 2018, 83, 1999–2007. [Google Scholar] [CrossRef]
- Darwish, M.S.; Qiu, L.; Taher, M.A.; Zaki, A.A.; Abou-Zeid, N.A.; Dawood, D.H.; Shalabi, O.M.A.K.; Khojah, E.; Elawady, A.A. Health Benefits of Postbiotics Produced by E. coli Nissle 1917 in Functional Yogurt Enriched with Cape Gooseberry (Physalis peruviana L.). Fermentation 2022, 8, 128. [Google Scholar] [CrossRef]
- Khojah, E.; Gomaa, M.S.; Elsherbiny, E.G.; Elawady, A.; Darwish, M.S. The In Vitro Analysis of Postbiotics in Functional Labneh to Be Used as Powerful Tool to Improve Cell Surfaces Properties and Adherence Potential of Probiotic Strains. Fermentation 2022, 8, 122. [Google Scholar] [CrossRef]
- Madhu, A.N.; Amrutha, N.; Prapulla, S.G. Characterization and Antioxidant Property of Probiotic and Synbiotic Yogurts. Probiotics Antimicrob. Proteins 2012, 4, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Elshaghabee, F.M.F.; El-Hussein, A.; Mohamed, M.S.M. Enhancement of Labneh Quality by Laser-Induced Modulation of Lactocaseibacillus casei NRRL B-1922. Fermentation 2022, 8, 132. [Google Scholar] [CrossRef]
- Casarotti, S.N.; Carneiro, B.M.; Penna, A.L.B. Evaluation of the effect of supplementing fermented milk with quinoa flour on probiotic activity. J. Dairy Sci. 2014, 97, 6027–6035. [Google Scholar] [CrossRef] [Green Version]
- Matouskova, P.; Hoova, J.; Rysavka, P.; Marova, I. Stress Effect of Food Matrices on Viability of Probiotic Cells during Model Digestion. Microorganisms 2021, 9, 1625. [Google Scholar] [CrossRef]
- Santos Monteiro, S.; Beserra, A.S.Y.; Oliveira, M.L.H.; Pasquali, M.A. Production of Probiotic Passion Fruit (Passiflora edulis Sims f. flavicarpa Deg.) Drink Using Lactobacillus reuteri and Microencapsulation via Spray Drying. Foods 2020, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- do Espírito Santo, A.P.; Perego, P.; Converti, A.; Oliveira, M.N. Influence of milk type and addition of passion fruit peel powder on fermentation kinetics, texture profile and bacterial viability in probiotic yoghurts. LWT 2012, 47, 393–399. [Google Scholar] [CrossRef]
- Cao, Q.; Teng, J.; Wei, B.; Huang, L.; Xia, N. Phenolic compounds, bioactivity, and bioaccessibility of ethanol extracts from passion fruit peel based on simulated gastrointestinal digestion. Food Chem. 2021, 356, 129682. [Google Scholar] [CrossRef]
- Elbermawi, A.; Darwish, M.S.; El-Awady, A.A.; Zaki, A.A.; Qiu, L.; Samra, R.M. Isolation and biological activities of compounds from Rumex vesicarius L. and their use as a component of a synbiotic preparation. Food Chem. X 2022, 14, 100306. [Google Scholar] [CrossRef] [PubMed]
- Balabanova, T.; Petkova, N.; Ivanova, M.; Panayotov, N. Design of Labneh cheese fortified with alginate-encapsulated pepper (Capsicum annuum) extracts. Emir. J. Food Agric. 2020, 32, 559–566. [Google Scholar] [CrossRef]
- Ning, X.; Luo, Z.; Chen, Z.; Zhou, C.; Xie, C.; Du, W.; Wang, L. Fortification of set yogurt with passion fruit juice: Effects on fermentation kinetics, physicochemical properties, and functionality. J. Dairy Sci. 2021, 104, 4084–4093. [Google Scholar] [CrossRef]
- Buriti, F.C.A.; Castro, I.A.; Saad, S.M.I. Viability of Lactobacillus acidophilus in synbiotic guava mousses and its survival under in vitro simulated gastrointestinal conditions. Int. J. Food Microbiol. 2010, 137, 121–129. [Google Scholar] [CrossRef]
- Fasoli, S.; Marzotto, M.; Rizzotti, L.; Rossi, F.; Dellaglio, F.; Torriani, S. Bacterial composition of commercial probiotic products as evaluated by PCR-DGGE analysis. Int. J. Food Microbiol. 2003, 82, 59–70. [Google Scholar] [CrossRef]
- Masco, L.; Huys, G.; De Brandt, E.; Temmerman, R.; Swings, J. Culture-dependent and culture-independent qualitative analysis of probiotic products claimed to contain bifidobacteria. Int. J. Food Microbiol. 2005, 102, 221–230. [Google Scholar] [CrossRef]
- dos Reis, L.C.R.; Facco, E.M.P.; Salvador, M.; Flôres, S.H.; de Oliveira Rios, A. Antioxidant potential and physicochemical characterization of yellow, purple and orange passion fruit. J. Food Sci. Technol. 2018, 55, 2679–2691. [Google Scholar] [CrossRef] [PubMed]
- Dias, C.O.; dos Santos Opuski de Almeida, J.; Pinto, S.S.; de Oliveira Santana, F.C.; Verruck, S.; Müller, C.M.O.; Prudêncio, E.S.; de Mello Castanho Amboni, R.D. Development and physico-chemical characterization of microencapsulated bifidobacteria in passion fruit juice: A functional non-dairy product for probiotic delivery. Food Biosci. 2018, 24, 26–36. [Google Scholar] [CrossRef]
- Varghese, K.S.; Mishra, H.N. Modelling of acidification kinetics and textural properties in dahi (Indian yogurt) made from buffalo milk using response surface methodology. Int. J. Dairy Technol. 2008, 61, 284–289. [Google Scholar] [CrossRef]
- Almeida, K.E.; Tamime, A.Y.; Oliveira, M.N. Influence of total solids contents of milk whey on the acidifying profile and viability of various lactic acid bacteria. LWT Food Sci. Technol. 2009, 42, 672–678. [Google Scholar] [CrossRef]
- McCann, T.H.; Fabre, F.; Day, L. Microstructure, rheology and storage stability of low-fat yoghurt structured by carrot cell wall particles. Food Res. Int. 2011, 44, 884–892. [Google Scholar] [CrossRef]
- De Souza Oliveira, R.P.; Perego, P.; Converti, A.; De Oliveira, M.N. Growth and acidification performance of probiotics in pure culture and co-culture with Streptococcus thermophilus: The effect of inulin. LWT Food Sci. Technol. 2009, 42, 1015–1021. [Google Scholar] [CrossRef]
- Zare, F.; Boye, J.I.; Champagne, C.P.; Orsat, V.; Simpson, B.K. Probiotic Milk Supplementation with Pea Flour: Microbial and Physical Properties. Food Bioprocess Technol. 2013, 6, 1321–1331. [Google Scholar] [CrossRef]
- Hashim, I.B.; Khalil, A.H.; Afifi, H.S. Quality characteristics and consumer acceptance of yogurt fortified with date fiber. J. Dairy Sci. 2009, 92, 5403–5407. [Google Scholar] [CrossRef]
- Sendra, E.; Fayos, P.; Lario, Y.; Fernández-López, J.; Sayas-Barberá, E.; Pérez-Alvarez, J.A. Incorporation of citrus fibers in fermented milk containing probiotic bacteria. Food Microbiol. 2008, 25, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Staffolo, M.D.; Bertola, N.; Martino, M.; Bevilacqua, Y.A. Influence of dietary fiber addition on sensory and rheological properties of yogurt. Int. Dairy J. 2004, 14, 263–268. [Google Scholar] [CrossRef]
- Vasiljevic, T.; Shah, N.P. Probiotics—From Metchnikoff to bioactives. Int. Dairy J. 2008, 18, 714–728. [Google Scholar] [CrossRef]
- Kailasapathy, K.; Harmstorf, I.; Phillips, M. Survival of Lactobacillus acidophilus and Bifidobacterium animalis ssp. lactis in stirred fruit yogurts. LWT Food Sci. Technol. 2008, 41, 1317–1322. [Google Scholar] [CrossRef]
- Auty, M.A.E.; Gardiner, G.E.; McBrearty, S.J.; O’Sullivan, E.O.; Mulvihill, D.M.; Collins, J.K.; Fitzgerald, G.F.; Stanton, C.; Ross, R.P. Direct In Situ Viability Assessment of Bacteria in Probiotic Dairy Products Using Viability Staining in Conjunction with Confocal Scanning Laser Microscopy. Appl. Environ. Microbiol. 2001, 67, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Moreno, Y.; Collado, M.C.; Ferrús, M.A.; Cobo, J.M.; Hernández, E.; Hernández, M. Viability assessment of lactic acid bacteria in commercial dairy products stored at 4 C using LIVE/DEAD® BacLightTM staining and conventional plate counts. Int. J. Food Sci. Technol. 2006, 41, 275–280. [Google Scholar] [CrossRef]
- Possemiers, S.; Marzorati, M.; Verstraete, W.; Van de Wiele, T. Bacteria and chocolate: A successful combination for probiotic delivery. Int. J. Food Microbiol. 2010, 141, 97–103. [Google Scholar] [CrossRef]
- Lavermicocca, P. Highlights on new food research. Dig. Liver Dis. 2006, 38 (Suppl. 2), S295–S299. [Google Scholar] [CrossRef]
- Valerio, F.; De Bellis, P.; Lonigro, S.L.; Morelli, L.; Visconti, A.; Lavermicocca, P. In vitro and in vivo survival and transit tolerance of potentially probiotic strains carried by artichokes in the gastrointestinal tract. Appl. Environ. Microbiol. 2006, 72, 3042–3045. [Google Scholar] [CrossRef] [Green Version]
- Nagy, G.; Dobrindt, U.; Grozdanov, L.; Hacker, J.; Emődy, L. Transcriptional regulation through RfaH contributes to intestinal colonization by Escherichia coli. FEMS Microbiol. Lett. 2005, 244, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, C.; Rani, R.S.; Usha, A. Evaluation of safety, antimicrobial activity and probiotic properties of Escherichia coli Nissle 1917 isolated from Idli batter. Res. J. Biotechnol. 2016, 11, 7. [Google Scholar]
- Campanella, D.; Rizzello, C.G.; Fasciano, C.; Gambacorta, G.; Pinto, D.; Marzani, B.; Scarano, N.; De Angelis, M.; Gobbetti, M. Exploitation of grape marc as functional substrate for lactic acid bacteria and bifidobacteria growth and enhanced antioxidant activity. Food Microbiol. 2017, 65, 25–35. [Google Scholar] [CrossRef]
- de Souza, E.L.; de Albuquerque, T.M.R.; Dos Santos, A.S.; Massa, N.M.L.; de Brito Alves, J.L. Potential interactions among phenolic compounds and probiotics for mutual boosting of their health-promoting properties and food functionalities–A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1645–1659. [Google Scholar] [CrossRef]
- Kemsawasd, V.; Chaikham, P.; Rattanasena, P. Survival of immobilized probiotics in chocolate during storage and with an in vitro gastrointestinal model. Food Biosci. 2016, 16, 37–43. [Google Scholar] [CrossRef]
- Succi, M.; Tremonte, P.; Pannella, G.; Tipaldi, L.; Cozzolino, A.; Coppola, R.; Sorrentino, E. Survival of commercial probiotic strains in dark chocolate with high cocoa and phenols content during the storage and in a static in vitro digestion model. J. Funct. Foods 2017, 35, 60–67. [Google Scholar] [CrossRef]
- Sánchez, B.; Bressollier, P.; Urdaci, M.C. Exported proteins in probiotic bacteria: Adhesion to intestinal surfaces, host immunomodulation and molecular cross-talking with the host. FEMS Immunol. Med. Microbiol. 2008, 54, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Saxelin, M.; Lassig, A.; Karjalainen, H.; Tynkkynen, S.; Surakka, A.; Vapaatalo, H.; Järvenpää, S.; Korpela, R.; Mutanen, M.; Hatakka, K. Persistence of probiotic strains in the gastrointestinal tract when administered as capsules, yoghurt, or cheese. Int. J. Food Microbiol. 2010, 144, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Marcináková, M.; Klingberg, T.D.; Lauková, A.; Budde, B.B. The effect of pH, bile and calcium on the adhesion ability of probiotic enterococci of animal origin to the porcine jejunal epithelial cell line IPEC-J2. Anaerobe 2010, 16, 120–124. [Google Scholar] [CrossRef]
- Deepika, G.; Rastall, R.A.; Charalampopoulos, D. Effect of food models and low-temperature storage on the adhesion of Lactobacillus rhamnosus GG to Caco-2 cells. J. Agric. Food Chem. 2011, 59, 8661–8666. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.; Tsakalidou, E.; Nychas, G.J.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Ryu, E.H.; Chang, H.C. In vitro study of potentially probiotic lactic acid bacteria strains isolated from kimchi. Ann. Microbiol. 2013, 63, 1387–1395. [Google Scholar] [CrossRef]
- Ranadheera, C.S.; Evans, C.A.; Adams, M.C.; Baines, S.K. In vitro analysis of gastrointestinal tolerance and intestinal cell adhesion of probiotics in goat’s milk ice cream and yogurt. Food Res. Int. 2012, 49, 619–625. [Google Scholar] [CrossRef]
- Barba, F.J.; Mariutti, L.R.B.; Bragagnolo, N.; Mercadante, A.Z.; Barbosa-Cánovas, G.V.; Orlien, V. Bioaccessibility of bioactive compounds from fruits and vegetables after thermal and nonthermal processing. Trends Food Sci. Technol. 2017, 67, 195–206. [Google Scholar] [CrossRef]
- Quan, W.; Tao, Y.; Lu, M.; Yuan, B.; Chen, J.; Zeng, M.; Qin, F.; Guo, F.; He, Z. Stability of the phenolic compounds and antioxidant capacity of five fruit (apple, orange, grape, pomelo and kiwi) juices during in vitro-simulated gastrointestinal digestion. Int. J. Food Sci. Technol. 2018, 53, 1131–1139. [Google Scholar] [CrossRef]
- AOAC, H.W. International A: Official Methods of Analysis of the AOAC International; The Association: Arlington County, VA, USA, 2000. [Google Scholar]
- Seixas, F.L.; Fukuda, D.L.; Turbiani, F.R.B.; Garcia, P.S.; Petkowicz, C.L.d.O.; Jagadevan, S.; Gimenes, M.L. Extraction of pectin from passion fruit peel (Passiflora edulis f. flavicarpa) by microwave-induced heating. Food Hydrocoll. 2014, 38, 186–192. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters/Treatments | Control | LP 0.5% | LP 1% |
|---|---|---|---|
| Initial pH | 6.61 ± 0.03 a | 6.4 ± 0.02 b | 6.33 ± 0.02 c |
| tpH5.5 (min) | 228.05 ± 0.23 a | 149.3 ± 0.68 b | 103.4 ± 1.3 c |
| tpH5.0 (min) | 314.8 ± 0.69 a | 230.9 ± 1.32 b | 165.4 ± 1.7 c |
| tpH4.5 (min) | 401.5 ± 1.56 a | 312.4 ± 2.1 b | 227.4 ± 2.1 c |
| Vmax (10–3pH/min) | 11.33 ± 0.67 c | 14.2 ± 1.3 b | 17.67 ± 0.88 a |
| Tmax (min) | 240 a | 90 b | 60 c |
| Treatment | pH | Titratable Acidity (% Lactic Acid) | ||||
|---|---|---|---|---|---|---|
| Storage Period (Days) | ||||||
| 0 | 7 | 15 | 0 | 7 | 15 | |
| Control | 4.48 ± 0.01 Aa | 4.39 ± 0.03 Ba | 4.32 ± 0.02 Ca | 0.86 ± 0.03 Cc | 0.99 ± 0.03 Bc | 1.08 ± 0.02 Ac |
| 0.5% PFPP | 4.42 ± 0.02 Ab | 4.35 ± 0.02 Bb | 4.25 ± 0.03 Cb | 0.95 ± 0.02 Cb | 1.08 ± 0.02 Bb | 1.16 ± 0.02 Ab |
| 1% PFPP | 4.37 ± 0.03 Ac | 4.32 ± 0.02 Bc | 4.20 ± 0.02 Cc | 1.05 ± 0.03 Ca | 1.14 ± 0.03 Ba | 1.22 ± 0.01 Aa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darwish, M.S.; Abou-Zeid, N.A.; Khojah, E.; AL Jumayi, H.A.; Alshehry, G.A.; Algarni, E.H.; Elawady, A.A. Supplementation of Labneh with Passion Fruit Peel Enhanced Survival of E. coli Nissle 1917 during Simulated Gastrointestinal Digestion and Adhesion to Caco-2 Cells. Foods 2022, 11, 1663. https://doi.org/10.3390/foods11111663
Darwish MS, Abou-Zeid NA, Khojah E, AL Jumayi HA, Alshehry GA, Algarni EH, Elawady AA. Supplementation of Labneh with Passion Fruit Peel Enhanced Survival of E. coli Nissle 1917 during Simulated Gastrointestinal Digestion and Adhesion to Caco-2 Cells. Foods. 2022; 11(11):1663. https://doi.org/10.3390/foods11111663
Chicago/Turabian StyleDarwish, Mohamed Samir, Noha A. Abou-Zeid, Ebtihal Khojah, Huda A. AL Jumayi, Garsa A. Alshehry, Eman H. Algarni, and Asmaa A. Elawady. 2022. "Supplementation of Labneh with Passion Fruit Peel Enhanced Survival of E. coli Nissle 1917 during Simulated Gastrointestinal Digestion and Adhesion to Caco-2 Cells" Foods 11, no. 11: 1663. https://doi.org/10.3390/foods11111663