
Metabolomics Combined with Transcriptomics Analysis Revealed the Amino Acids, Phenolic Acids, and Flavonol Derivatives Biosynthesis Network in Developing Rosa roxburghii Fruit

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Determination of Bioactive Substance Content and Antioxidant Capacity
2.3. Metabolite Extraction and ESI-Q TRAP-MS/MS Analysis
2.4. Qualitative and Quantitative Analysis of Metabolites
2.5. Transcriptome Information and Prediction of Transcription Factors
2.6. Quantitative RT-PCR Analysis
2.7. Statistical Analysis
3. Results
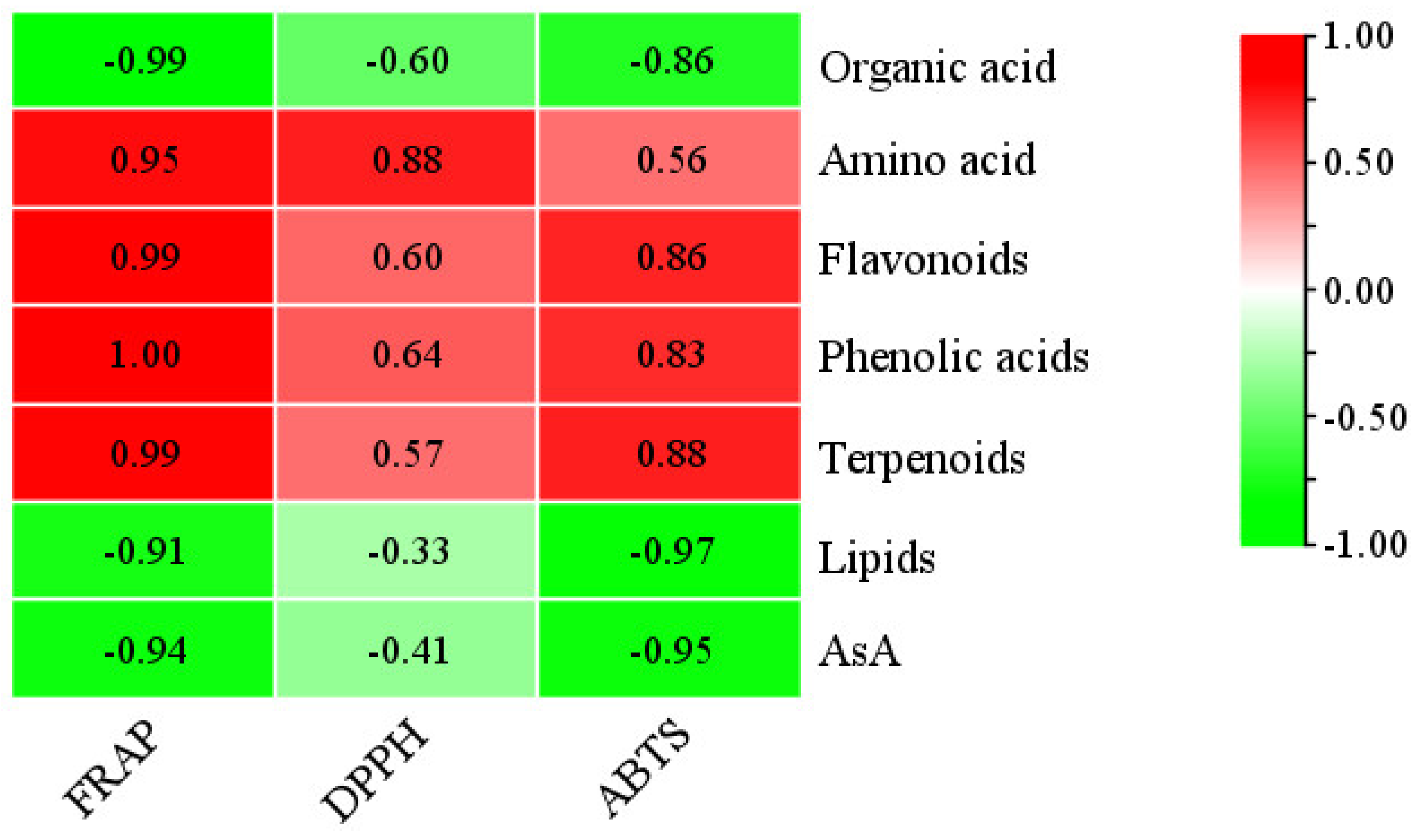
3.1. Bioactive Substance Content and Antioxidant Capacity in R. roxburghii during Fruit Development
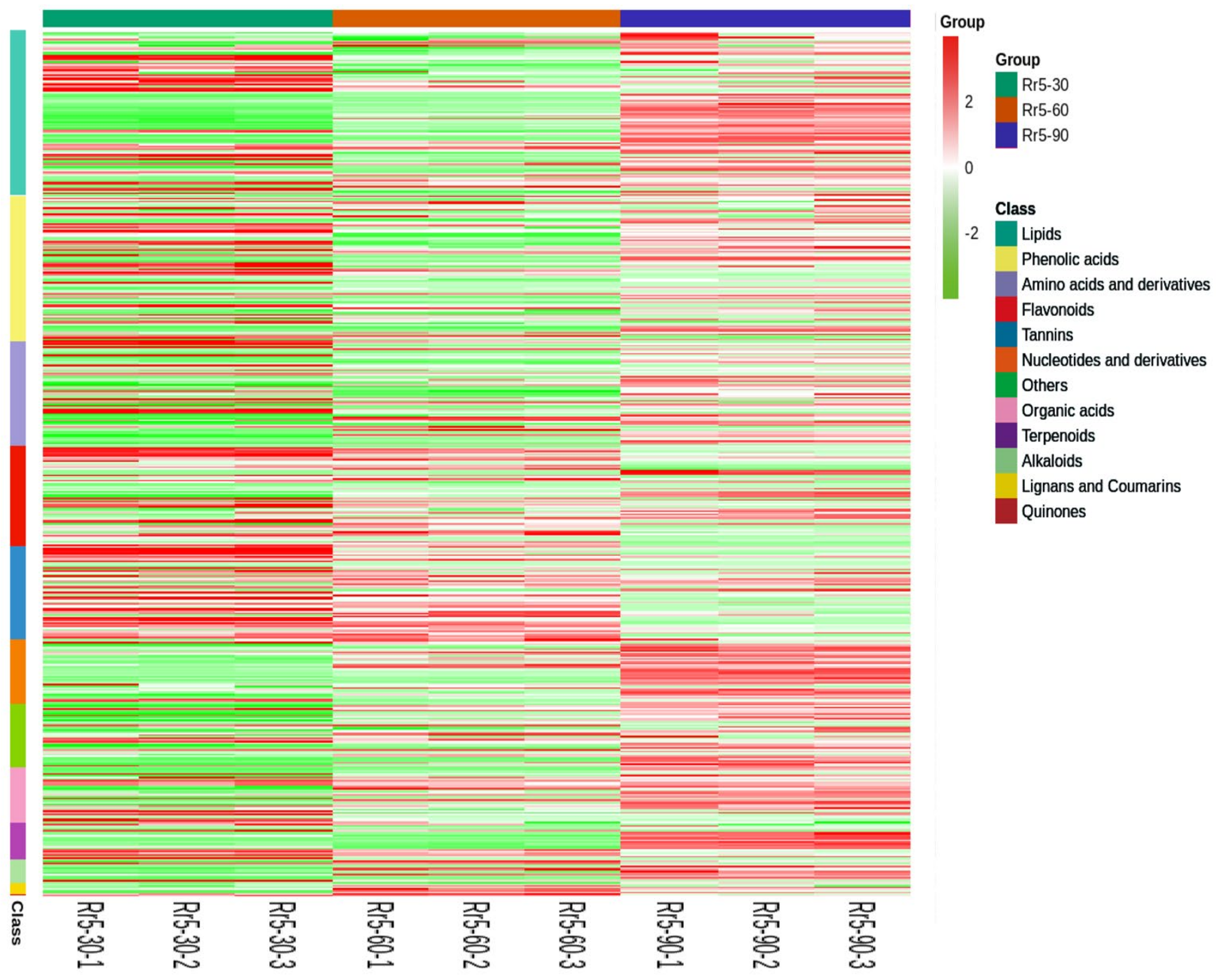
3.2. Metabolome Profiling of R. roxburghii Fruits at Different Ripening Stages
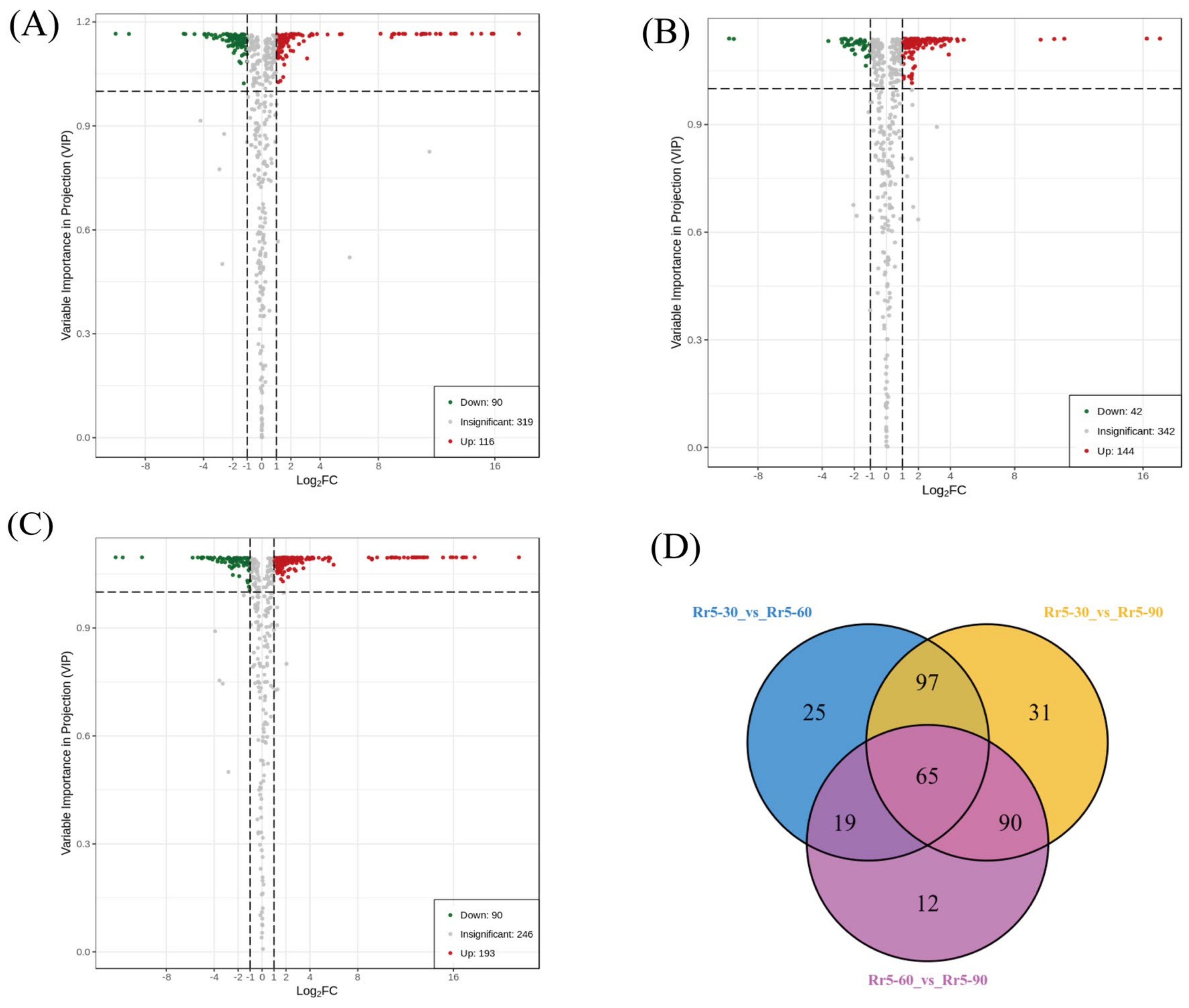
3.3. Differentially Accumulated Metabolites between Young and Mature Fruits of R. roxburghii
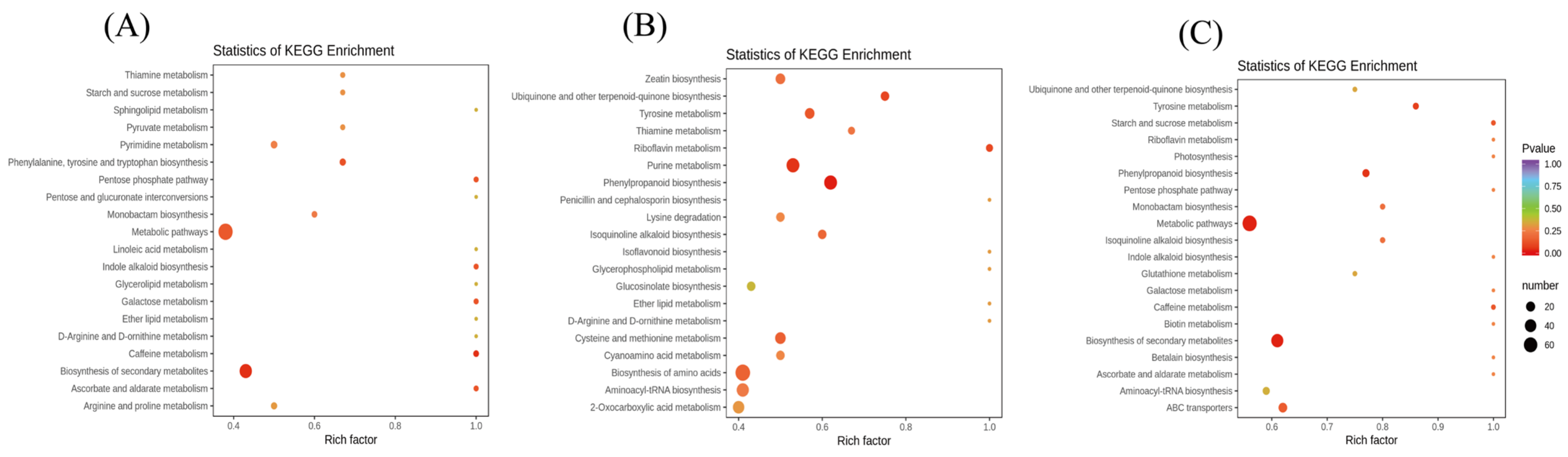
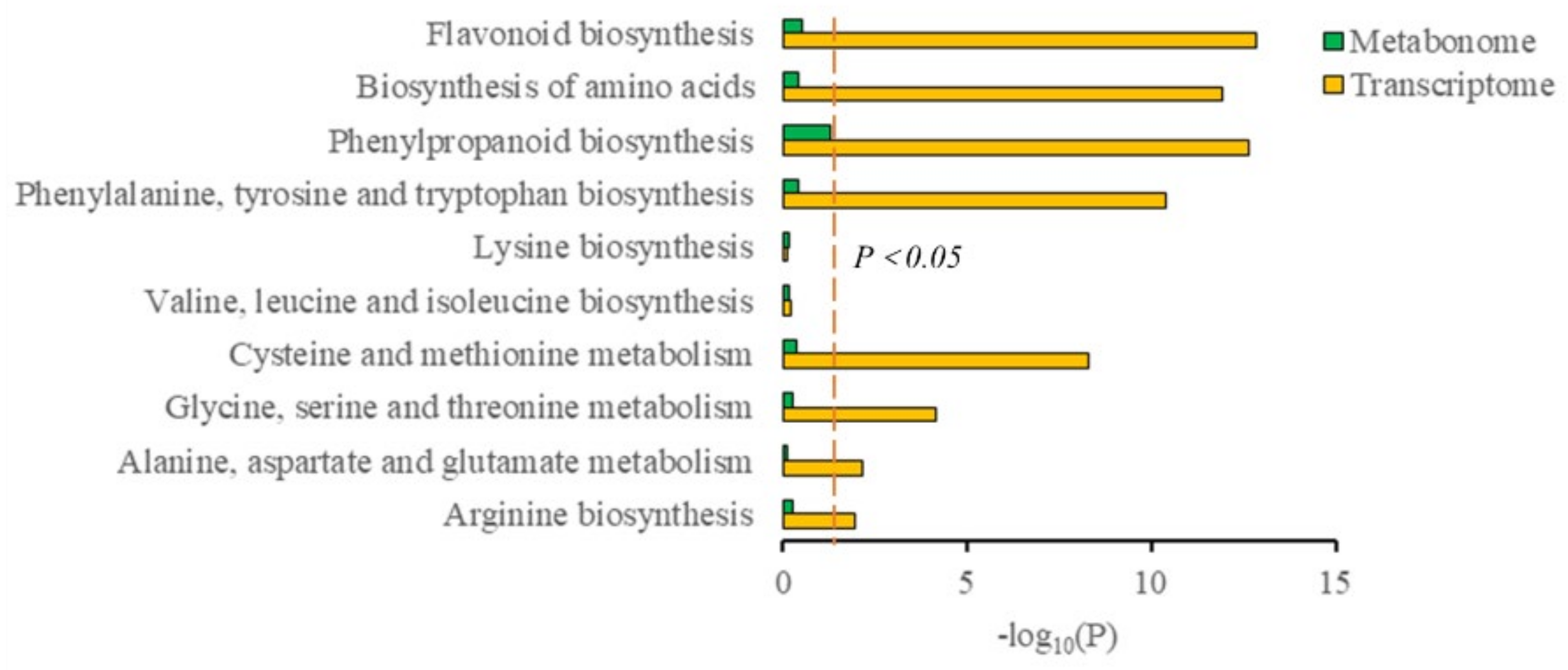
3.4. Conjoint Analysis of Transcriptome and Metabolome
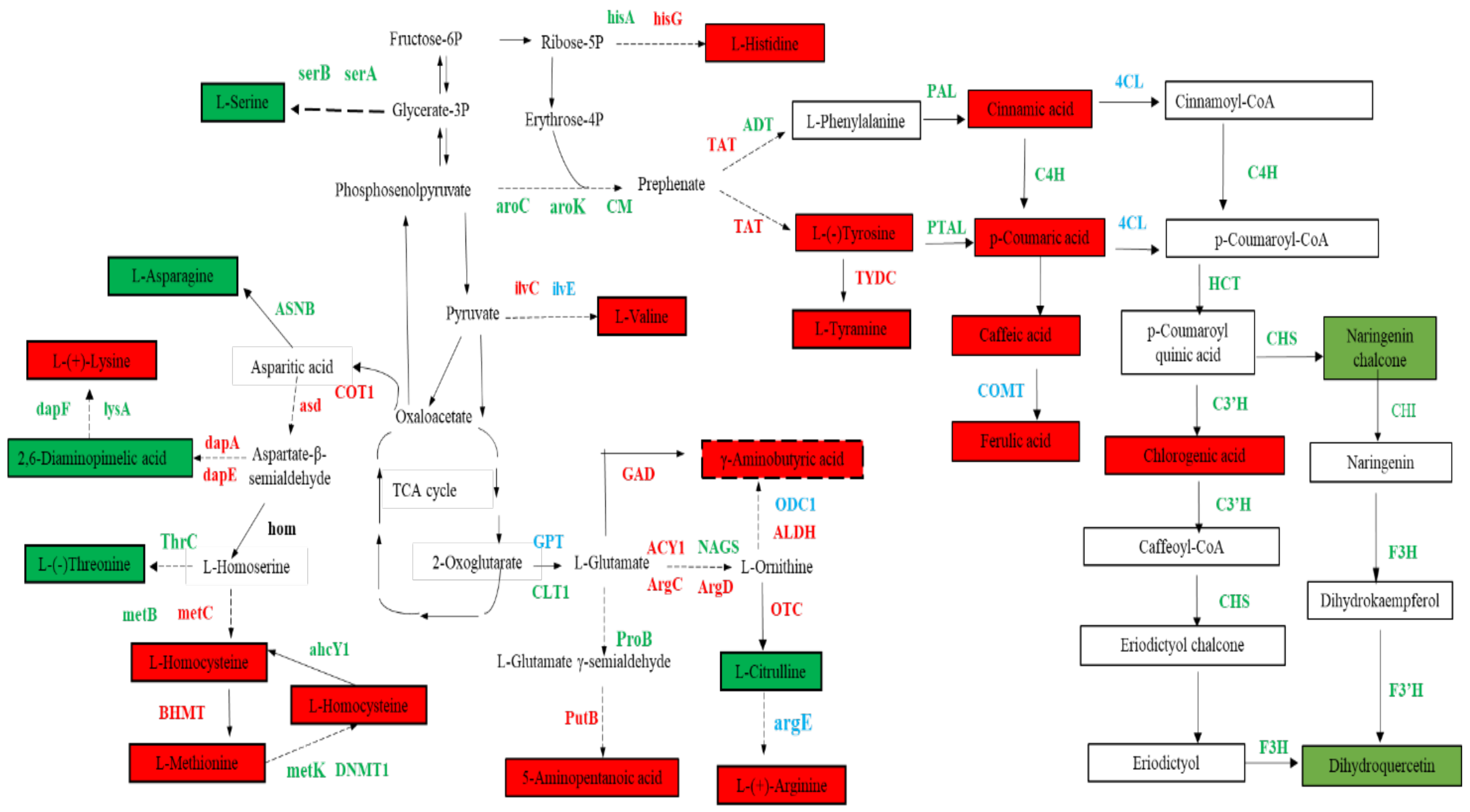
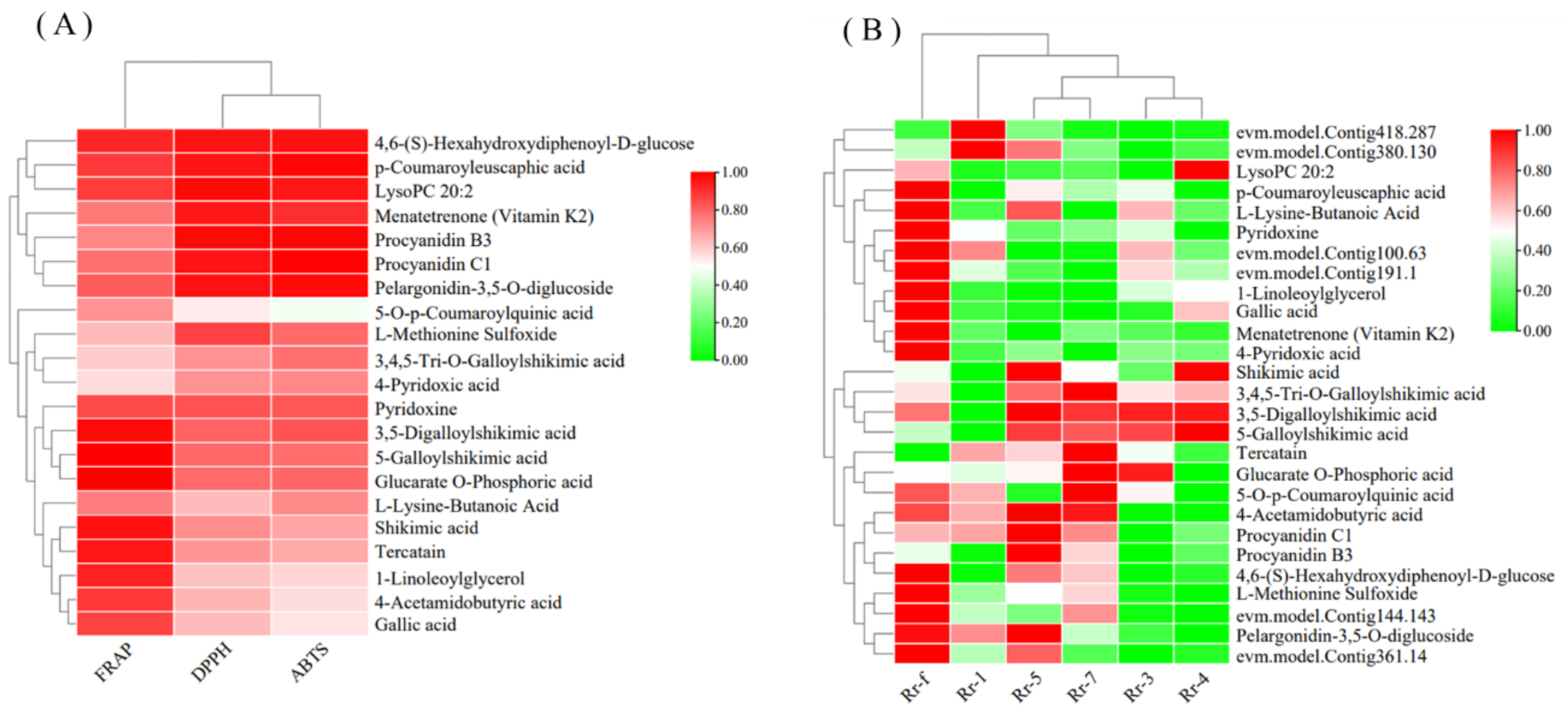
3.5. Biosynthesis of Amino Acids, Phenolic Acids, and Flavonol Derivatives
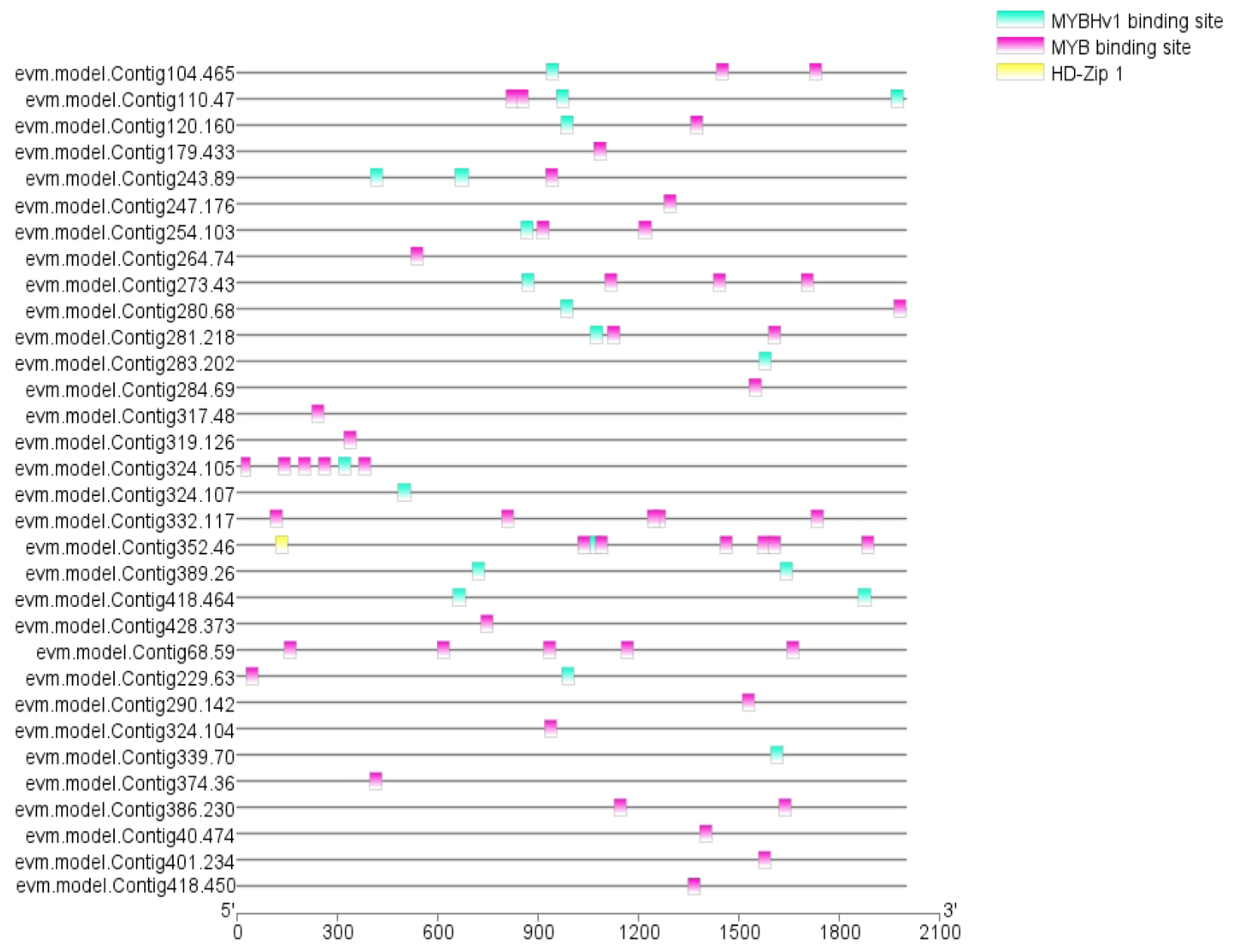
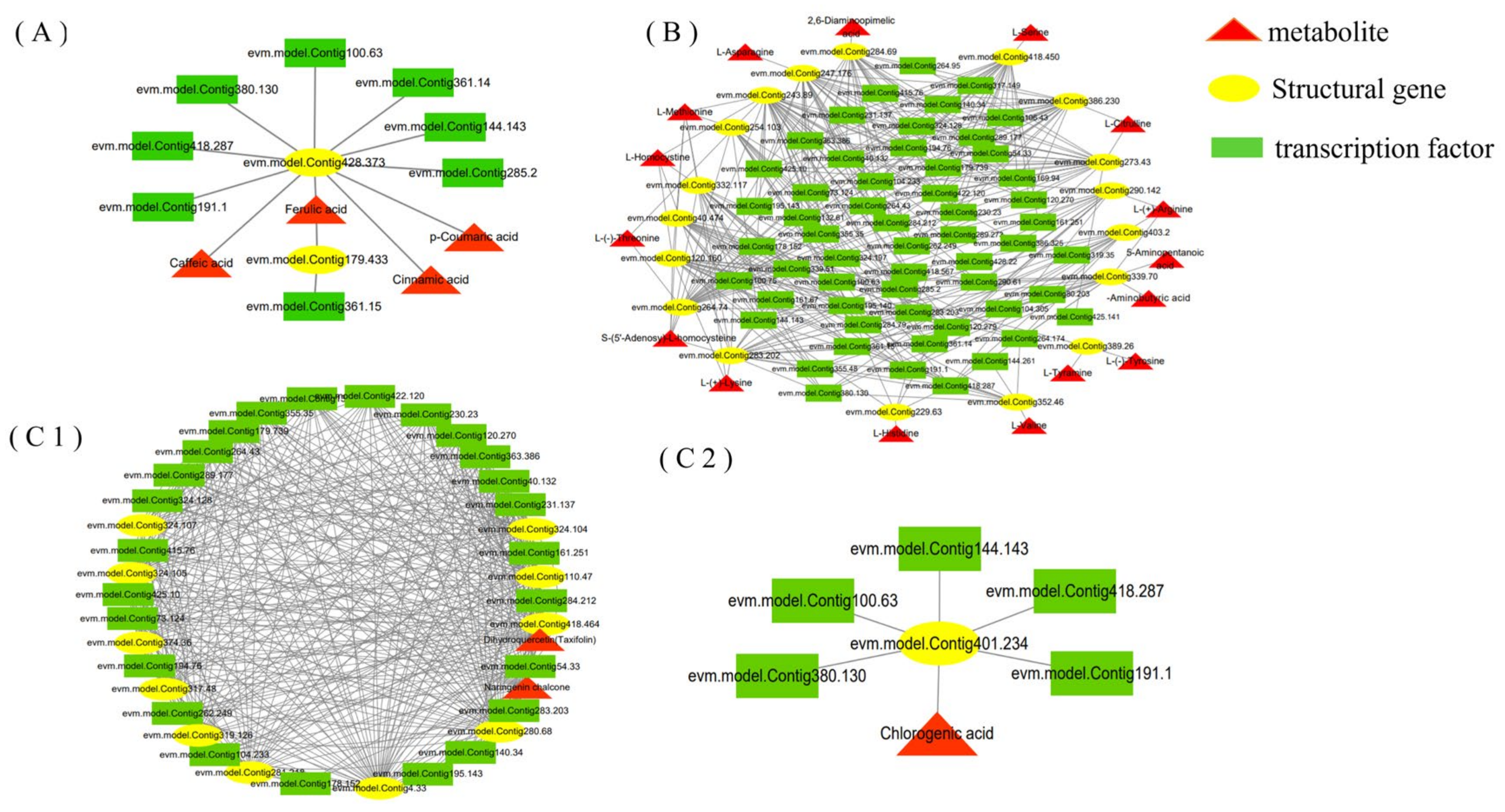
3.6. RrMYBs in the Regulation of Amino Acid, Phenolic Acid, and Flavonol Derivative Biosynthesis
3.7. Key RrMYBs Expression and Metabolites in Different R. roxburghii Genotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- An, H.M.; Liu, M.; Yang, M.; Fang, W.G. Analysis of organic acid components and ascorbic acid content of Rosa roxburghii. China Agric. Sci. 2011, 44, 2094–2100. [Google Scholar]
- Lu, M.; An, H.M.; Wang, D.P. Characterization of amino acid composition in fruits of three Rosa roxburghii genotypes. Hortic. Plant J. 2017, 3, 232–236. [Google Scholar] [CrossRef]
- Zeng, F.; Ge, Z.; Limwachiranon, J.; Li, L.; Feng, S.; Wang, Y.; Luo, Z. Antioxidant and tyrosinase inhibitory activity of Rosa roxburghii fruit and identification of main bioactive phytochemicals by UPLC-Triple-TOF/MS. Int. J. Food Sci. Technol. 2017, 52, 897–905. [Google Scholar] [CrossRef]
- Qin, J.J.; Li, Q.J.; Xue, Y.; Lin, M.A.; Yang, X.S. Extraction method and application of total triterpenes from Rosa roxburghii Tratt α-Study on glucosidase inhibitory activity. Food Ind. Sci. Technol. 2014, 35, 186–189. [Google Scholar]
- Zhou, G.Z.; Lu, M.; An, H.M. Analysis on the content of main active substances and their antioxidant activity during fruit development of Rosa roxburghii. Food Sci. 2018, 39, 20–25. [Google Scholar]
- Jiang, L.L.; Lu, M.; Rao, T.Z.; Liu, Z.Y.; Wu, X.M.; An, H.M. Comparative analysis of fruit metabolome using widely targeted metabolomics reveals nutritional characteristics of fifferent Rosa roxburghii genotypes. Foods 2022, 11, 850. [Google Scholar] [CrossRef]
- Xu, S.J.; Zhang, F.; Wang, L.J.; Hao, M.H.; Yang, X.J.; Li, N.N.; Ji, H.L.; Xu, P. Flavonoids of Rosa roxburghii Tratt offers protection against radiation induced apoptosis and inflammation in mouse thymus. Apoptosis 2018, 23, 470–483. [Google Scholar] [CrossRef]
- Yuan, H.; Wang, Y.; Chen, H.; Cai, X. Protective effect of flavonoids from Rosa roxburghii Tratt on myocardial cells via autophagy. 3 Biotech 2020, 10, 58. [Google Scholar] [CrossRef]
- Chen, C.; Tan, S.; Ren, T.; Wang, H.; Dai, X.; Wang, H. Polyphenol from Rosa roxburghii Tratt Fruit Ameliorates the Symptoms of Diabetes by Activating the P13K/AKT Insulin Pathway in db/db Mice. Foods 2022, 11, 636. [Google Scholar] [CrossRef]
- Yan, X.; Zhang, X.; Lu, M.; He, Y.; An, H. De novo sequencing analysis of the Rosa roxburghii fruit transcriptome reveals putative ascorbate biosynthetic genes and EST-SSR markers. Gene 2015, 561, 54–62. [Google Scholar] [CrossRef]
- Lu, M.; Ma, W.T.; Liu, Y.Q.; An, H.M.; Ludlow, R.A. Transcriptome analysis reveals candidate lignin-related genes and transcription factors in Rosa roxburghii during fruit ripening. Plant Mol. Biol. Report. 2020, 38, 331–342. [Google Scholar] [CrossRef]
- Huang, X.L.; Yan, H.Q.; Zhai, L.S.; Yang, Z.T.; Yi, Y. Characterization of the Rosa roxburghii Tratt transcriptome and analysis of MYB genes. PLoS ONE 2019, 14, e0203014. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; An, H.; Li, L. Genome Survey Sequencing for the Characterization of the Genetic Background of Rosa roxburghii Tratt and Leaf Ascorbate Metabolism Genes. PLoS ONE 2016, 11, e0147530. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.Q.; Lu, M.; An, H.M. SSR information analysis and molecular marker development of Rosa roxburghii transcriptome. J. Hortic. 2015, 42, 341–349. [Google Scholar]
- Meng, J.; Wang, B.; He, G.; Wang, Y.; Tang, X.; Wang, S.; Ma, Y.; Fu, C.; Chai, G.; Zhou, G. Metabolomics integrated with transcriptomics reveals redirection of the phenylpropanoids metabolic flux in Ginkgo biloba. Agric. Food Chem. 2019, 67, 3284–3291. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, G.; Ma, B.; Zhong, C.; He, N. Metabolic profiling and transcriptome analysis of mulberry leaves provide insights into flavonoid biosynthesis. Agric. Food Chem. 2020, 68, 1494–1504. [Google Scholar] [CrossRef] [PubMed]
- Umer, M.J.; Safdar, L.B.; Gebremeskel, H.; Zhao, S.; Yuan, P.; Zhu, H.; Kaseb, M.O.; Anees, M.; Lu, X.; He, N. Identification of key gene networks controlling organic acid and sugar metabolism during watermelon fruit development by integrating metabolic phenotypes and gene expression profiles. Hortic. Res. 2020, 7, 193. [Google Scholar] [CrossRef]
- Zhang, Z.; Tian, C.; Zhang, Y.; Li, C.; Feng, S. Transcriptomic and metabolomic analysis provides insights into anthocyanin and procyanidin accumulation in pear. BMC Plant Biol. 2020, 20, 129. [Google Scholar] [CrossRef] [Green Version]
- Fortes, A.M.; Agudelo, R.P.; Silva, M.S.; Ali, K.; Sousa, L.; Maltese, F.; Choi, Y.H.; Grimplet, J.; Martinez, Z.J.M.; Verpoorte, R.; et al. Transcript and metabolite analysis in Trincadeira cultivar reveals novel information regarding the dynamics of grape ripening. BMC Plant Biol. 2011, 11, 149. [Google Scholar] [CrossRef] [Green Version]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Huang, R.S.; Yu, Y.X.; Hu, Y.; Sheng, X.B. Determination of total flavonoids in abrus cantoniensis and its dynamic changes. China J. Chin. Mater. Med. 2006, 31, 1428–1431. [Google Scholar]
- Zhou, Q.X.; Zhang, J.S.; Gu, M.; Gu, Z.L. Determination of total triterpenoid acids in Hawthorn and its extract. Chin. Wild Plant Resour. 2004, 5, 43–44. [Google Scholar]
- Wang, X.K.; Huang, J.L. Principles and Techniques of Plant Physiological Biochemical Experiment; Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Andrés, M.; Franco, D.; Sineiro, J.; Domnguez, H.; Lema, J.M. Antioxidant activity of extracts from Gevuina avellana and Rosa rubiginosa defatted seeds. Food Res. Int. 2001, 34, 103–109. [Google Scholar]
- Martinez, V.C.; Penas, E.; Ciska, E.; Piskula, M.K.; Kozlowska, H.; Vidal, V.C.; Frias, J. Time dependence of bioactive compounds and antioxidant capacity during germination of different cultivars of broccoli and radish seeds. Food Chem. 2010, 120, 710–716. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, D.F.; Chen, J.H.; Guo, J.; Chen, H.B.; Li, P.F. Comparative study on the content of polyphenols and flavonoids and their antioxidant activity of seven citrus species. Food Res. Dev. 2019, 40, 69–74. [Google Scholar]
- Zhang, M.M.; Zhou, P.Y.; Zhang, S.T.; Li, L.X.; Cui, Y.; Sun, B.S. Analysis of polyphenol content and antioxidant activity of wine grape. J. Shenyang Pharm. Univ. 2019, 36, 539–548. [Google Scholar]
- Zhao, Q.F.; Tan, W.; Dong, Z.G.; Li, X.M.; Tan, M.; Tang, X.P. Comparative analysis of fruit quality of different nutritional lines of three wine grape varieties, ‘pinlizhu’, ‘Shiraz’ and ‘Pinot Noir’. Chin. Agron. Bull. 2018, 34, 56–60. [Google Scholar]
- Zhang, X.; Bi, J.F.; Chen, Q.Q.; Lv, Y.; Wu, X.Y.; Li, X.; Guo, J. Component analysis and antioxidant activity of four berries. Food Sci. Technol. 2020, 45, 52–58. [Google Scholar]
- Guo, X.; Shakeel, M.; Wang, D.; Qu, S.Y.; Yang, S.Y.; Ahmad, S.; Son, Z.J. Metabolome and transcriptome profiling unveil the mechanisms of light-induced anthocyanin synthesis in rabbiteye blueberry (vaccinium ashei: Reade). BMC Plant Biol. 2022, 22, 223. [Google Scholar] [CrossRef]
- Chen, X.; Cai, W.; Xia, J.; Yu, H.; Wang, Q.; Pang, F.; Zhao, M. Metabolomic and Transcriptomic Analyses Reveal that Blue Light Promotes Chlorogenic Acid Synthesis in Strawberry. J. Agric. Food Chem. 2020, 68, 12485–12492. [Google Scholar] [CrossRef]
- Wang, F.; Chen, L.; Chen, S.; Chen, H.; Liu, Y. Characterization of two closely related citrus cultivars using UPLC-ESI-MS/MS-based widely targeted metabolomics. PLoS ONE 2021, 16, e0254759. [Google Scholar] [CrossRef]
- Ma, W.; Xu, L.; Gao, S.; Lyu, X.; Cao, X.; Yao, Y. Melatonin alters the secondary metabolite profile of grape berry skin by promoting VvMYB14-mediated ethylene biosynthesis. Hortic. Res. 2021, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, G.; Nishioka, I.; Nishizawa, M.; Yamagishi, T.; Kashiwada, Y.; Dutschman, G.E.; Bodner, A.J.; Kilkuskie, R.E.; Cheng, Y.C.; Lee, K.H. Anti-AIDS agents, 2: Inhibitory effects of tannins on HIV reverse transcriptase and HIV replication in H9 lymphocyte cells. J. Nat. Prod. 1990, 53, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.F. Exploration of Natural Substances Against Influenza Virus (H1N1); Taipei Medical University: Taibei, Taiwan, 2009. [Google Scholar]
- Cerulli, A.; Masullo, M.; Mari, A.; Balato, A.; Filosa, R.; Lembo, S.; Napolitano, A.; Piacente, S. Phenolics from Castanea sativa leaves and their effects on UVB-induced damage. Nat. Prod. Res. 2018, 32, 1170–1175. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, Y.; Zheng, Y.; Zhao, J.; Yu, H.; Zhu, J. Relationship between Neuroprotective Effects and Structure of Procyanidins. Molecules 2020, 27, 2308. [Google Scholar] [CrossRef]
- Khalifa, I.; Zhu, W.; Nafie, M.S.; Dutta, K.; Li, C. Anti-COVID-19 effects of ten structurally different hydrolysable tannins through binding 1 with the catalytic-closed sites of COVID-19 main protease: An in-silico approach 2. Reverse Vaccinol. 2020, 3, 0277. [Google Scholar]
- Li, W.; Gu, M.; Gong, P.; Wang, J.; Yang, H. Glycosides changed the stability and antioxidant activity of pelargonidin. LWT—Food Sci. Technol. 2021, 147, 111581. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, Y.; Wu, H.; Wu, C.; Ji, E.; Xu, J.; Zhang, Y.; Wei, J.; Zhao, Y.; Yang, H. Systematic survey of the alteration of the faecal microbiota in rats with gastrointestinal disorder and modulation by multicomponent drugs. Front. Pharmacol. 2021, 12, 670335. [Google Scholar] [CrossRef]
- Kim, J.A.; Lau, E.; Tay, D.; Blanco, D.E.J. Antioxidant and NF-κB inhibitory constituents isolated from Morchella esculenta. Nat. Prod. Res. 2011, 25, 1412–1417. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Song, J.; Zhao, S.; Jiang, H.; Chi, A. Therapeutic effect and metabolic mechanism of a selenium-polysaccharide from Ziyang green tea on chronic fatigue syndrome. Polymers 2018, 10, 1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, B.; Zhu, Q.; Zhao, Y.; Ge, W.; Zhao, Y.; Song, Q.; Zhou, Y.; Shi, H.; Zhang, Y. Phthalate exposure and childhood overweight and obesity: Urinary metabolomic evidence. Environ. Int. 2018, 121, 159–168. [Google Scholar] [CrossRef]
- Batory, M.; Rotsztejn, H. Shikimic acid in the light of current knowledge. J. Cosmet. Dermatol. 2022, 21, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; He, N.; Men, P.; Song, C.; Zhai, S. The efficacy and safety of menatetrenone in the management of osteoporosis: A systematic review and meta-analysis of randomized controlled trials. Osteoporos. Int. 2019, 30, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Obeid, R.; Geisel, J.; Nix, W. 4-pyridoxic acid/pyridoxine ratio in patients with type 2 diabetes is related to global cardiovascular risk scores. Diagnostics 2019, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Ding, Y.; Nie, Y.; Wang, X.J.; An, Y.Q.; Roessner, U.; Walker, R.; Du, B.; Bai, J.G. Plant metabolomics integrated with transcriptomics and rhizospheric bacterial community indicates the mitigation effects of Klebsiella oxytoca P620 on p-hydroxybenzoic acid stress in cucumber. J. Hazard. Mater. 2021, 415, 125756. [Google Scholar] [CrossRef]
- Fodor, M.; Mikola, E.E.; Geösel, A.; Stefanovits-Bányai, É.; Mednyánszky, Z. Application of Near-Infrared Spectroscopy to Investigate Some Endogenic Properties of Pleurotus ostreatus Cultivars. Sensors 2020, 20, 6632. [Google Scholar] [CrossRef]
- Akashi, K.; Miyake, C.; Yokota, A. Citrulline, a novel compatible solute in drought-tolerant wild watermelon leaves, is an efficient hydroxyl radical scavenger. FEBS Lett. 2001, 508, 438–442. [Google Scholar] [CrossRef] [Green Version]
- Akinyede, A.I.; Fagbemi, T.N.; Osundahunsi, O.F.; Aluko, R.E. Amino acid composition and antioxidant properties of the enzymatic hydrolysate of calabash nutmeg (Monodora myristica) and its membrane ultrafiltration peptide fractions. J. Food Biochem. 2021, 45, 13437. [Google Scholar] [CrossRef]
- Hwang, H.S.; Winkler, M.J.K.; Liu, S.X. Study on antioxidant activity of amino acids at frying temperatures and their interaction with rosemary extract, green tea extract, and ascorbic acid. J. Food Sci. 2019, 84, 3614–3623. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Fang, B.; Jiang, L.; Guo, H.Y.; Cui, J.Y.; Ren, F.Z. Structure-activity relationship of a series of antioxidant tripeptides derived from β-Lactoglobulin using QSAR modeling. Dairy Sci. Technol. 2015, 95, 451–463. [Google Scholar] [CrossRef]
- Catanesi, M.; Brandolini, L.; Angelo, M.; Benedetti, E.; Tupone, M.G.; Alfonsetti, M.; Cabri, E.; Iaconis, D.; Fratelli, M.; Cimini, A.; et al. L-methionine protects against oxidative stress and mitochondrial dysfunction in an in vitro model of Parkinson’s disease. Antioxidants 2021, 10, 1467. [Google Scholar] [CrossRef]
- Guo, Y.W. Amino Acids in Nutrition and Health; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Clark, T.C.; Tinsley, J.; Sigholt, T.; Macqueen, D.J.; Martin, S.A.M. Arginine, ornithine and citrulline supplementation in rainbow trout: Free amino acid dynamics and gene expression responses to bacterial infection. Fish Shellfish. Immunol. 2020, 98, 374–390. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, R.E.; Nazarian, B.; Saboori, S.; Falahi, E.; Hekmatdoost, A. Effects of Arginine supplementation on glycemic profile: Evidence from a systematic review and meta-analysis of clinical trials. J. Integr. Med. 2020, 18, 284–291. [Google Scholar] [CrossRef]
- Wang, J. Study on the Effect of Amino Acids on Blood Glucose and Its Related Mechanism; Chinese People’s Liberation Army Medical College: Beijing, China, 2012. [Google Scholar]
- Fremont, N.; Riefler, M.; Stolz, A.; Schmülling, T. The Arabidopsis TUMOR PRONE5 gene encodes an acetylornithine aminotransferase required for arginine biosynthesis and root meristem maintenance in blue light. Plant Physiol. 2013, 161, 1127–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.; Joshi, M.; Dipiazza, J.; Joshi, V. Functional Relevance of Citrulline in the Vegetative Tissues of Watermelon During Abiotic Stresses. Front. Plant Sci. 2015, 151, 352–364. [Google Scholar] [CrossRef]
- Ata, N.; Yusuf, M.; Tan, B.C.; Husaini, A.; Mohd Yusuf, Y.; Majid, N.A.; Khalid, N. Expression profiles of flavonoid-related gene, 4 coumarate: Coenzyme A ligase, and optimization of culturing conditions for the selected flavonoid production in Boesenbergia rotunda. Plant Cell Tissue Organ Cult. 2015, 123, 47–55. [Google Scholar]
- Fan, C.; Hu, H.; Wang, L.; Zhou, Q.; Huang, X. Enzymological mechanism for the regulation of lanthanum chloride on flavonoid synthesis of soybean seedlings under enhanced ultraviolet-B radiation. Environ. Sci. Pollut. Res. 2014, 21, 8792–8800. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, S.; Li, H.; Wang, L.; Liu, Y.; Niu, L.; Yang, Q.; Meng, D.; Fu, Y. Genome-wide analysis and characterization of R2R3-MYB family in pigeon pea (Cajanus cajan) and their functional identification in phenylpropanoids biosynthesis. Planta 2021, 254, 64. [Google Scholar] [CrossRef]
- Premathilake, A.T.; Ni, J.; Bai, S.; Tao, R.; Ahmad, M.; Teng, Y. R2R3-MYB transcription factor PpMYB17 positively regulates flavonoid biosynthesis in pear fruit. Planta 2020, 252, 59. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, B.L.; Wei, L.; Ye, G.J. Frame-shift Mutation Causes the Function Loss of TaMYB-A1 Regulating Anthocyanin Biosynthesis in Triticum aestivum. Cereal Res. Commun. 2017, 45, 35–46. [Google Scholar]
- Ding, Z.Q.; Chen, T.Z.; Liu, T.L.; Liu, X.S.; Zhang, B.L.; Zhou, X.G. Functional analysis of MYB transcription factor GHRAX3 induced by drought in cotton. China Agric. Sci. 2015, 48, 3569–3579. [Google Scholar]
- Li, X.Y. Screening and Functional Analysis of REVEILLEs Transcription Factors Regulating the Circadian Accumulation of Anthocyanin in Pear Peel; Northwest University of Agriculture and Forestry Science and Technology: Xian, China, 2021; pp. 40–50. [Google Scholar]
- Kidokoro, S.; Hayashi, K.; Haraguchi, H.; Ishikawa, T.; Soma, F.; Konoura, I.; Toda, S.; Mizoi, J.; Suzuki, T.; Shinozaki, K.; et al. Posttranslational regulation of multiple clock-related transcription factors triggers cold-inducible gene expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2021048118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, P.; Zhao, J.; Chen, G. Synteny-based mapping of causal point mutations relevant to sand rice (Agriophyllum squarrosum) trichomeless1 mutant by RNA-sequencing. J. Plant Physiol. 2018, 231, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.P.; Yang, S.Q.; Xu, K.; Luo, L.J. Expression Characteristics and Transcriptional Self-Excited Activity Analysis of Drought Tolerance Gene OsMYB105 in Rice. 2022. Available online: https://kns.cnki.net/kcms/detail/46.1068.S.20220411.0008.002.html (accessed on 20 March 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Acid (% FW) | Amino Acid (μmol/g FW) | Flavonoids (mg/100 g FW) | Total Phenols (mg/100 g FW) | Terpenoids (mg/100 g FW) | AsA Content (mg/100 g FW) | Lipids (% FW) | DPPH (μmol TE/g FW) | FRAP (μmol TE/g FW) | ABTS (μmol TE/g FW) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Rr5-30 | 0.58 ± 0.04 c | 22.23 ± 1.96 a | 2250.12 ± 139.77 a | 4540.25 ± 451.04 a | 7543.93 ± 39.71 a | 70.78 ± 0.23 c | 0.90 ± 0.07 a | 78.57 ± 0.41 a | 1902.78 ± 78.84 a | 154.29 ± 3.89 a |
| Rr5-60 | 0.83 ± 0.07 b | 9.05 ± 1.57 b | 1416.51 ± 58.23 b | 3430.60 ± 50.15 b | 5024.89 ± 286.07 b | 285.08 ± 10.12 b | 0.94 ± 0.04 a | 58.76 ± 3.42 c | 891.67 ± 43.83 b | 58.96 ± 3.46 c |
| Rr5-90 | 1.12 ± 0.04 a | 6.95 + 0.94 b | 456.32 + 30.86 c | 2376.71 ±202.62 c | 1701.63 ± 23.51 c | 861.13 ± 15.56 a | 1.11 ± 0.16 a | 66.04 ± 3.23 b | 140.97 ± 8.72 c | 66.03 ± 3.21 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, N.; Jiang, L.; Liu, Y.; Zou, S.; Lu, M.; An, H. Metabolomics Combined with Transcriptomics Analysis Revealed the Amino Acids, Phenolic Acids, and Flavonol Derivatives Biosynthesis Network in Developing Rosa roxburghii Fruit. Foods 2022, 11, 1639. https://doi.org/10.3390/foods11111639
Li N, Jiang L, Liu Y, Zou S, Lu M, An H. Metabolomics Combined with Transcriptomics Analysis Revealed the Amino Acids, Phenolic Acids, and Flavonol Derivatives Biosynthesis Network in Developing Rosa roxburghii Fruit. Foods. 2022; 11(11):1639. https://doi.org/10.3390/foods11111639
Chicago/Turabian StyleLi, Nanyu, Lanlan Jiang, Yiyi Liu, Shimei Zou, Min Lu, and Huaming An. 2022. "Metabolomics Combined with Transcriptomics Analysis Revealed the Amino Acids, Phenolic Acids, and Flavonol Derivatives Biosynthesis Network in Developing Rosa roxburghii Fruit" Foods 11, no. 11: 1639. https://doi.org/10.3390/foods11111639