
Survival of Escherichia coli O157:H7 during Moderate Temperature Dehydration of Plant-Based Foods

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Food Samples
2.2. Inoculum Selection and Preparation
2.3. Sample Inoculation
2.4. Isothermal Studies
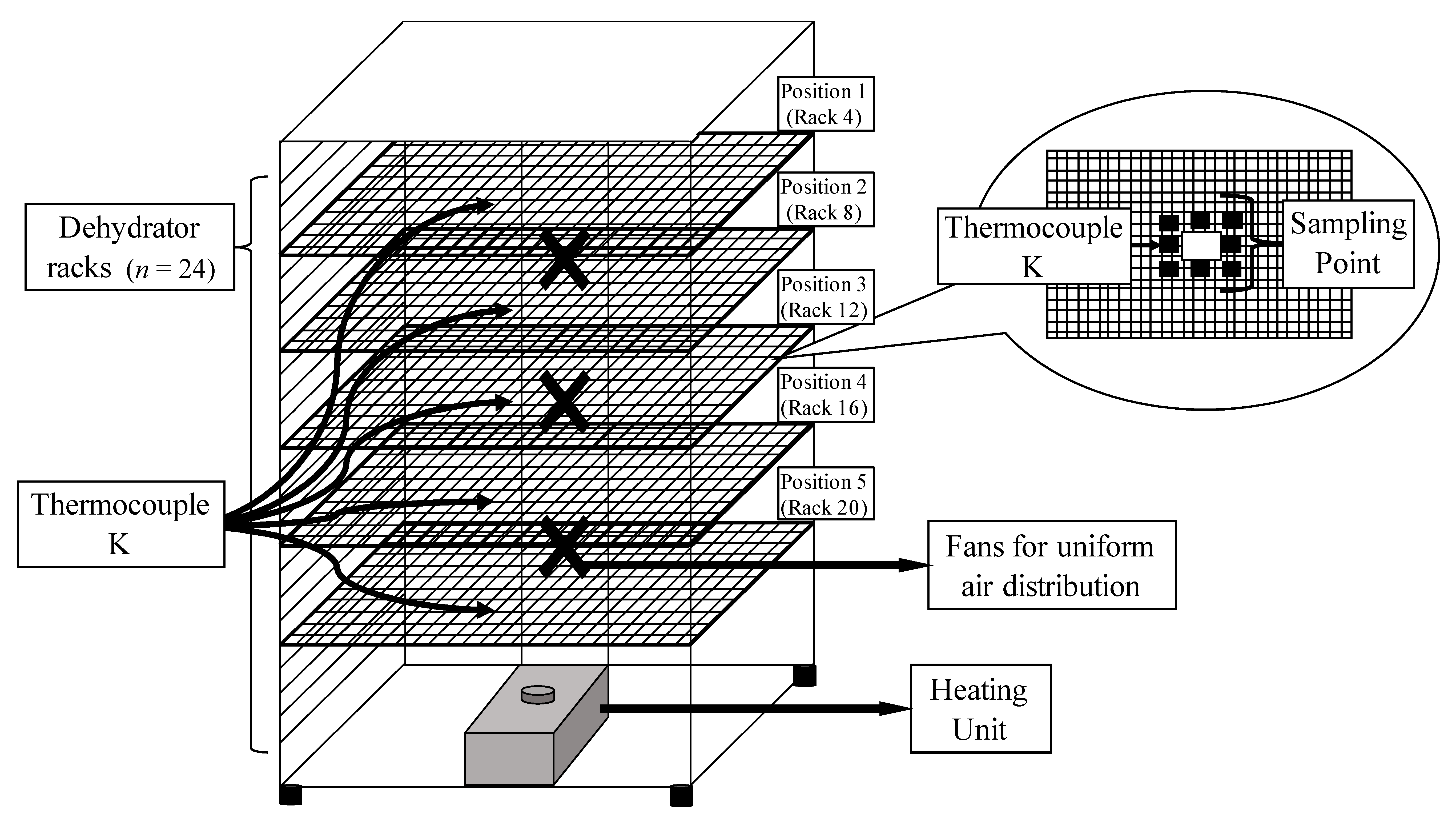
2.5. Inoculated Pack Dehydration Studies
2.6. Isothermal Data Modeling
2.7. Statistical Modeling
3. Results and Discussion
3.1. Inactivation of E. coli O157:H7 during Isothermal Treatments
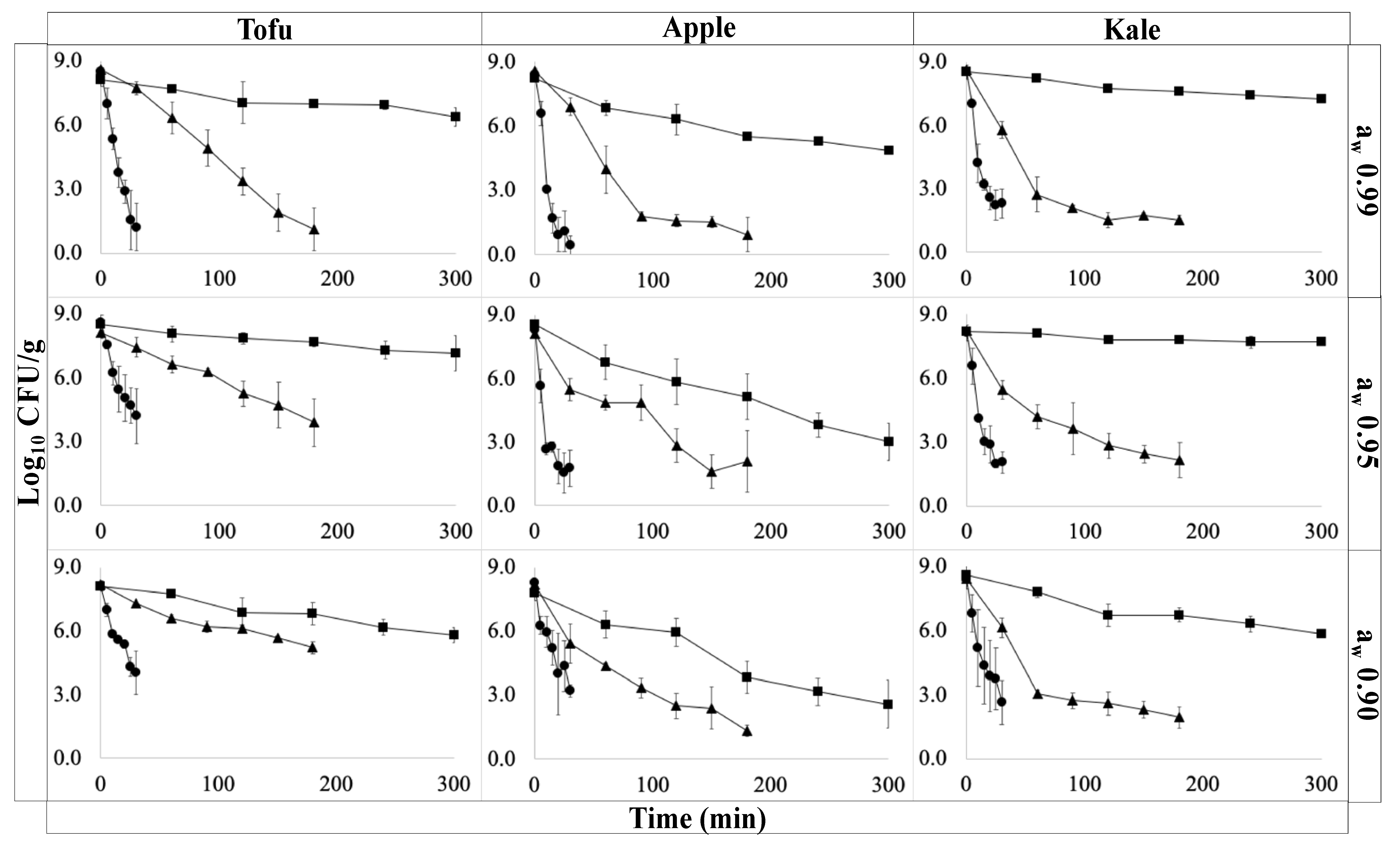
3.2. The Impact of Sample Location on Temperature, aw, and Microbial Inactivation in Inoculated Pack Studies
3.3. Changes in E. coli O157:H7 during Inoculated Pack Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beuchat, L.R.; Komitopoulou, E.; Beckers, H.; Betts, R.P.; Bourdichon, F.; Fanning, S.; Joosten, H.M.; Ter-Kuile, B.H. Low-water activity foods: Increased concern as vehicles of foodborne pathogens. J. Food Prot. 2013, 76, 150–172. [Google Scholar] [CrossRef] [PubMed]
- Bourdoux, S.; Li, D.; Rajkovic, A.; Devlieghere, F.; Uyttendaele, M. Performance of drying technologies to ensure microbial safety of dried fruits and vegetables. Comp. Rev. Food Sci. Food Saf. 2016, 15, 1056–1066. [Google Scholar] [CrossRef]
- Jayaraman, K.S.; Das-Gupta, D.K. Drying of fruits and vegetables. In Handbook of Industrial Drying, 4th ed.; Mujumdar, A.S., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 611–635. [Google Scholar]
- Saravacos, G.D.; Kostaropoulos, A.E. Food dehydration equipment. In Handbook of Food Processing Equipment, 1st ed.; Saravacos, G.D., Kostaropoulos, A.E., Eds.; Springer: New York, NY, USA, 2002; pp. 331–382. [Google Scholar]
- Burnham, J.A.; Kendall, P.A.; Sofos, J.N. Ascorbic acid enhances destruction of Escherichia coli O157:H7 during home-type drying of apple slices. J. Food Prot. 2001, 64, 1244–1248. [Google Scholar] [CrossRef]
- Derrickson-Tharrington, E.; Kendall, P.A.; Sofos, J.N. Inactivation of Escherichia coli O157:H7 during storage or drying of apple slices pretreated with acidic solutions. Int. J. Food Microbiol. 2005, 99, 79–89. [Google Scholar] [CrossRef]
- Dipersio, P.A.; Kendall, P.A.; Calicioglu, M.; Sofos, J.N. Inactivation of Salmonella during drying and storage of apple slices treated with acidic or sodium metabisulfite solutions. J. Food Prot. 2003, 12, 2245–2251. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). List of Selected Multistate Foodborne Outbreak Investigations. 2020. Available online: https://www.cdc.gov/foodsafety/outbreaks/multistate-outbreaks/outbreaks-list.html (accessed on 28 June 2020).
- European Food Safety Authority (EFSA); Panel on Biological Hazards (BIOHAZ). Scientific Opinion on the Risk Posed by Pathogens in Food of Non-Animal Origin. Part 1 (Outbreak Data Analysis and Risk Ranking of Food/Pathogen Combinations). EFSA J. 2013, 11, 3025. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention (CDC). Multistate Outbreak of Salmonella Typhimurium Infections Linked to Dried Coconut (Final Update). 2018. Available online: https://www.cdc.gov/salmonella/typhimurium-03-18/index.html (accessed on 28 June 2020).
- Centers for Disease Control and Prevention (CDC). Outbreak of E. coli Infections Linked to Flour (Final Update). 2019. Available online: https://www.cdc.gov/ecoli/2019/flour-05-19/index.html (accessed on 28 June 2020).
- Mujumdar, A.S. Principles, classification, selection of dryers. In Handbook of Industrial Drying, 4th ed.; Mujumdar, A.S., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 3–30. [Google Scholar]
- Kiang, C.S.; Jon, C.K. Heat Pump Drying Systems. In Handbook of Industrial Drying, 4th ed.; Mujumdar, A.S., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 1101–1126. [Google Scholar]
- Cai, S.; Worobo, R.W.; Snyder, A.B. Combined effect of storage condition, surface integrity, and length of shelf life on the growth of Listeria monocytogenes and spoilage microbiota on refrigerated ready-to-eat products. J. Food Prot. 2019, 82, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration (FDA). Bad Bug Book, Foodborne Pathogenic Microorganisms and Natural Toxins, 2nd ed.; Lampel, K., Al-Khaldi, S., Cahill, S., Eds.; Center for Food Safety and Applied Nutrition: Washington, DC, USA, 2012. Available online: https://www.fda.gov/files/food/published/Bad-Bug-Book-2nd-Edition-%28PDF%29.pdf (accessed on 20 December 2019).
- Linton, R. Food Safety Hazards in Foodservice and Food Retail Establishments. 2003. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.222.2542&rep=rep1&type=pdf (accessed on 20 December 2019).
- Food and Drug Administration (FDA). H.R. 2751 FDA Food Safety Modernization Act. 2011. Available online: https://www.fda.gov/food/guidance-regulation-food-and-dietary-supplements/food-safety-modernization-act-fsma (accessed on 20 February 2020).
- Santillana-Farakos, S.M.; Frank, J.F.; Schaffner, D.W. Modeling the influence of temperature, water activity and water mobility on the persistence of Salmonella in low-moisture foods. Int. J. Food Microbiol. 2013, 166, 280–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdramidis, V.P.; Geeraerd, A.H.; Gaze, J.E.; Kondjoyan, A.; Boyd, A.R.; Shaw, H.L.; Van Impe, J.F. Quantitative description of Listeria monocytogenes inactivation kinetics with temperature and water activity as the influencing factors; model prediction and methodological validation on dynamic data. J. Food Eng. 2006, 76, 79–88. [Google Scholar] [CrossRef]
- Villa-Rojas, R.; Tang, J.M.; Wang, S.J.; Gao, M.X.; Kang, D.H.; Mah, J.H.; Gray, P.; Sosa-Morales, M.E.; Lopez-Malo, A. Thermal inactivation of Salmonella Enteritidis PT 30 in almond kernels as influenced by water activity. J. Food Prot. 2013, 76, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Snyder, A.B.; Perry, J.J.; Yousef, A.E. Developing and optimizing bacteriophage treatment to control enterohemorrhagic Escherichia coli on fresh produce. Int. J. Food Microbiol. 2016, 236, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.F.; Hilderbrandt, I.M.; Casulli, K.E.; Dolan, K.D.; Marks, B.P. Modeling the effect of temperature and water activity on the thermal resistance of Salmonella Enteritidis PT 30 in wheat flour. J. Food Prot. 2016, 79, 2058–2065. [Google Scholar] [CrossRef] [PubMed]
- Lenth, R. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.4.8. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 1 September 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 September 2021).
- Mattick, K.L.; Jorgensen, F.; Wang, P.; Pound, J.; Vandeven, M.H.; Ward, L.R.; Legan, J.D.; Lappin-Scott, H.M.; Humphrey, T.J. Effect of challenge temperature and solute type on heat tolerance of Salmonella serovars at low water activity. Appl. Environ. Microbiol. 2001, 67, 4128–4136. [Google Scholar] [CrossRef] [Green Version]
- Usaga, J.; Worobo, R.W.; Padilla-Zakour, O.I. Effect of acid adaptation and acid shock on thermal tolerance and survival of Escherichia coli O157:H7 and O111 in apple juice. J. Food Prot. 2014, 77, 1656–1663. [Google Scholar] [CrossRef] [PubMed]
- Amaha, M.; Sakaguchi, K.I. Effects of carbohydrates, proteins, and bacterial cells in the heating media on the heat resistance of Clostridium sporogenes. J. Bacteriol. 1954, 68, 338–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buerman, E.C.; Worobo, R.W.; Padilla-Zakour, O.I. Thermal resistance of xerophilic fungi in Low-Water-Activity (0.70 to 0.80) confectionery model foods. J. Food Protect. 2019, 82, 390–394. [Google Scholar] [CrossRef]
- He, Y.S.; Li, Y.; Salazar, J.K.; Yang, J.Y.; Tortorello, M.L.; Zhang, W. Increased water activity reduces the thermal resistance of Salmonella enterica in peanut butter. Appl. Environ. Microbiol. 2013, 79, 4763–4767. [Google Scholar] [CrossRef] [Green Version]
- Gautam, B.; Govindan, B.N.; Gänzle, M.; Roopesh, M.S. Influence of water activity on the heat resistance of Salmonella enterica in selected low-moisture foods. Int. J. Food Microbiol. 2020, 334, 108813. [Google Scholar] [CrossRef]
- Chen, L.; Wei, X.; Chaves, B.D.; Jones, D.; Ponder, M.A.; Subbiah, J. Inactivation of Salmonella enterica and Enterococcus faecium NRRL B2354 on cumin seeds using gaseous ethylene oxide. Food Microbiol. 2021, 94, 103656. [Google Scholar] [CrossRef]
- Bianchini, A.; Stratton, J.; Weier, S.; Hartter, T.; Plattner, B.; Rokey, G.; Hertzel, G.; Gompa, L.; Martinez, B.; Eskridge, K.M. Use of Enterococcus faecium as a surrogate for Salmonella enterica during extrusion of a balanced carbohydrate-protein meal. J. Food Protect. 2014, 77, 75–82. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, L.; Cheng, T.; Guan, X.; Wang, S. Effects of water activity, temperature and particle size on thermal inactivation of Escherichia coli ATCC 25922 in red pepper powder. Food Control 2020, 107, 106817. [Google Scholar] [CrossRef]
- Brar, P.K.; Proano, L.G.; Friedrich, L.M.; Harris, L.J.; Danyluk, M.D. Survival of Salmonella, Escherichia coli O157:H7, and Listeria monocytogenes on raw peanut and pecan kernels stored at −24, 4, and 22 °C. J. Food Protect. 2015, 78, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Pickens, S.R.; Hildebrandt, I.M.; Burbick, S.J.; Grasso-Kelley, E.M.; Keller, S.E.; Anderson, N.M. Thermal inactivation of Salmonella Agona in Low–Water Activity foods: Predictive models for the combined effect of temperature, water activity, and food component. J. Food Protect. 2018, 81, 1411–1417. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Tang, J.; Zhu, M. Water activity influence on the thermal resistance of Salmonella in soy protein powder at elevated temperatures. Food Control 2020, 113, 107160. [Google Scholar] [CrossRef]
- Liu, S.; Rojas, R.V.; Gray, P.; Zhu, M.; Tang, J. Enterococcus faecium as a Salmonella surrogate in the thermal processing of wheat flour: Influence of water activity at high temperatures. Food Microbiol. 2018, 74, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Lau, S.K.; Chaves, B.D.; Danao, M.C.; Agarwal, S.; Subbiah, J. Effect of water activity on the thermal inactivation kinetics of Salmonella in milk powders. J. Dairy Sci. 2020, 103, 6904–6917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, L.; Lan, R.; Zhou, X.; Kou, X.; Wang, S. Thermal inactivation of Aspergillus flavus in peanut kernels as influenced by temperature, water activity and heating rate. Food Microbiol. 2018, 76, 237–244. [Google Scholar] [CrossRef]
- Xu, J.; Tang, J.; Jin, Y.; Song, J.; Yang, R.; Sablani, S.S.; Zhu, M. High temperature water activity as a key factor influencing survival of Salmonella Enteritidis PT30 in thermal processing. Food Control 2018, 98, 520–528. [Google Scholar] [CrossRef]
- Giannakourou, M.C.; Stoforos, N.G. A theoretical analysis for assessing the variability of secondary model thermal inactivation kinetic parameters. Foods 2017, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Lang, E.; Chemlal, L.; Molin, P.; Guyot, S.; Alvarez-Martin, P.; Perrier-Cornet, J.M.; Dantigny, P.; Gervais, P. Modeling the heat inactivation of foodborne pathogens in milk powder: High relevance of the substrate water activity. Food Res. Int. 2017, 99, 577–585. [Google Scholar] [CrossRef]
- Brackett, R.E.; Ocasio, W.; Waters, K.; Barach, J.; Wan, J. Validation and Verification: A practical, industry-driven framework developed to support the requirements of the Food Safety Modernization Act (FSMA) of 2011. Food Protect. Trends 2014, 34, 410–425. [Google Scholar]
- Brar, P.K.; Danyluk, M.D. Nuts and grains: Microbiology and preharvest contamination risks. Preharvest Food Saf. 2018, 105–121. [Google Scholar] [CrossRef]
- Cai, S.; Phinney, D.; Heldman, D.; Snyder, A.B. All treatment parameters affect environmental surface sanitation efficacy, but their relative importance depends on the microbial target. Appl. Environ. Microbiol. 2020, 87, e01748-20. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Product | Estimations from Equation (2) (Temperature Only) | Estimations from Equation (3) (Temperature & aw) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| aw | Temp (°C) | D-Value (min ± SE) | z-Value (°C ± SE) | RMSE (log CFU/g) | D49°C (min ± SE) | z-Value (°C ± SE) | Raw (Unitless ± SE) | RMSE (log CFU/g) | |
| Apple | 0.99 | 49 | 78.2 ± 8.2 | 7.4 ± 0.6 | 1.26 | 71.9 ± 10.1 | 8.7 ± 0.5 | 1.1 ± 0.7 | 1.54 |
| 54.5 | 19.3 ± 2.3 | ||||||||
| 60 | 3.0 ± 0.5 | ||||||||
| 0.95 | 49 | 45.9 ± 3.9 | 8.9 ± 1.1 | 1.86 | |||||
| 54.5 | 25.2 ± 2.9 | ||||||||
| 60 | 3.5 ± 0.5 | ||||||||
| 0.9 | 49 | 54.5 ± 6.8 | 10.9 ± 1.2 | 1.12 | |||||
| 54.5 | 23.1 ± 2.5 | ||||||||
| 60 | 5.5 ± 0.8 | ||||||||
| Tofu | 0.99 | 49 | 173.3 ± 35.1 | 6.9 ± 0.3 | 1.31 | 107.3 ± 14.2 | 7.7 ± 0.4 | 0.6 ± 0.5 | 1.24 |
| 54.5 | 24.1 ± 1.6 | ||||||||
| 60 | 3.8 ± 0.3 | ||||||||
| 0.95 | 49 | 173.5 ± 16.6 | 6.9 ± 0.4 | 1.11 | |||||
| 54.5 | 43.3 ± 4.5 | ||||||||
| 60 | 6.1 ± 0.8 | ||||||||
| 0.9 | 49 | 107.6 ± 12.7 | 7.6 ± 0.8 | 1.18 | |||||
| 54.5 | 55.4 ± 4.6 | ||||||||
| 60 | 6.7 ± 0.6 | ||||||||
| Kale | 0.99 | 49 | 174.1 ± 27.2 | 7.5 ± 0.6 | 0.62 | 143.9 ± 17.0 | 7.0 ± 0.3 | 3.3 ± 0.5 | 0.75 |
| 54.5 | 19.6 ± 2.8 | ||||||||
| 60 | 3.8 ± 0.4 | ||||||||
| 0.95 | 49 | 257.4 ± 71.0 | 7.1 ± 0.5 | 0.64 | |||||
| 54.5 | 22.8 ± 3.0 | ||||||||
| 60 | 4.0 ± 0.4 | ||||||||
| 0.9 | 49 | 86.3 ± 10.8 | 8.7 ± 0.8 | 0.82 | |||||
| 54.5 | 22.2 ± 3.1 | ||||||||
| 60 | 4.8 ± 0.8 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rana, Y.S.; Eberly, P.M.; Suehr, Q.J.; Hildebrandt, I.M.; Marks, B.P.; Snyder, A.B. Survival of Escherichia coli O157:H7 during Moderate Temperature Dehydration of Plant-Based Foods. Foods 2021, 10, 2162. https://doi.org/10.3390/foods10092162
Rana YS, Eberly PM, Suehr QJ, Hildebrandt IM, Marks BP, Snyder AB. Survival of Escherichia coli O157:H7 during Moderate Temperature Dehydration of Plant-Based Foods. Foods. 2021; 10(9):2162. https://doi.org/10.3390/foods10092162
Chicago/Turabian StyleRana, Yadwinder Singh, Philip M. Eberly, Quincy J. Suehr, Ian M. Hildebrandt, Bradley P. Marks, and Abigail B. Snyder. 2021. "Survival of Escherichia coli O157:H7 during Moderate Temperature Dehydration of Plant-Based Foods" Foods 10, no. 9: 2162. https://doi.org/10.3390/foods10092162