
Effect of Addition of Tryptophan on Aggregation of Apo-α-Lactalbumin Induced by UV-Light


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Sample Preparation and Illumination
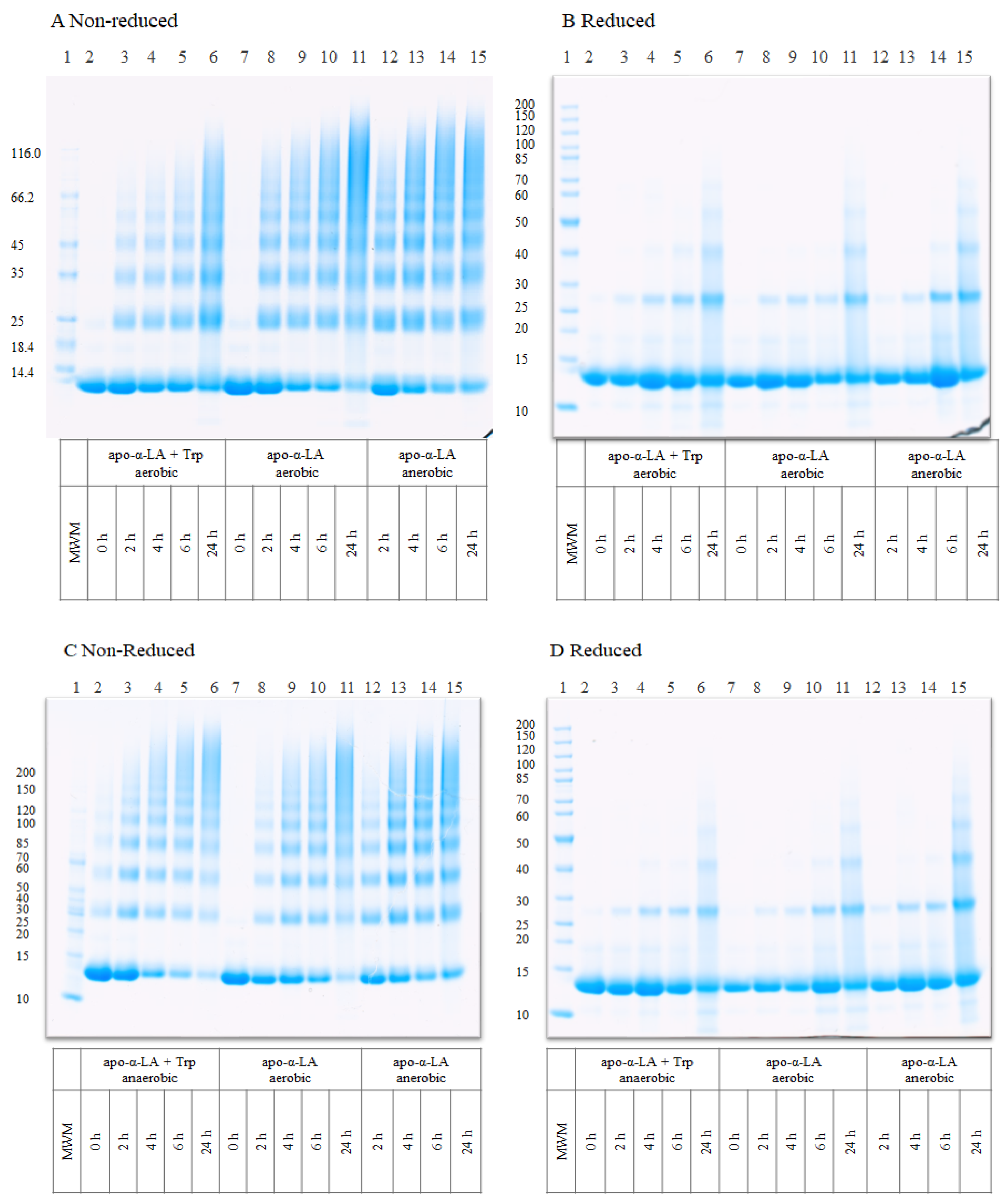
2.3. Characterization of Protein Aggregates by Gel Electrophoresis
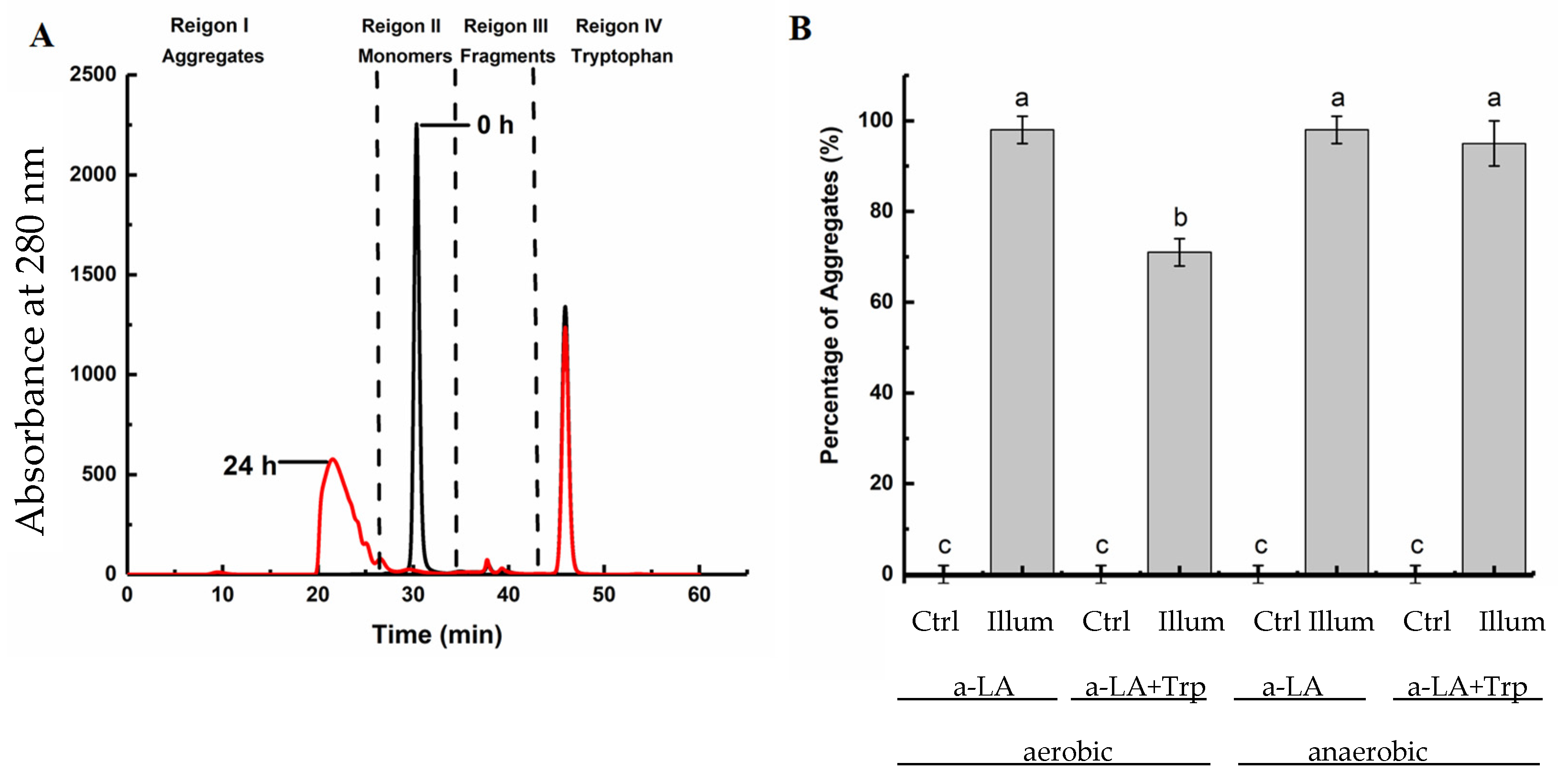
2.4. Determination of the Molecular Mass Distribution of Protein Aggregates by Size Exclusion Chromatography (SEC)
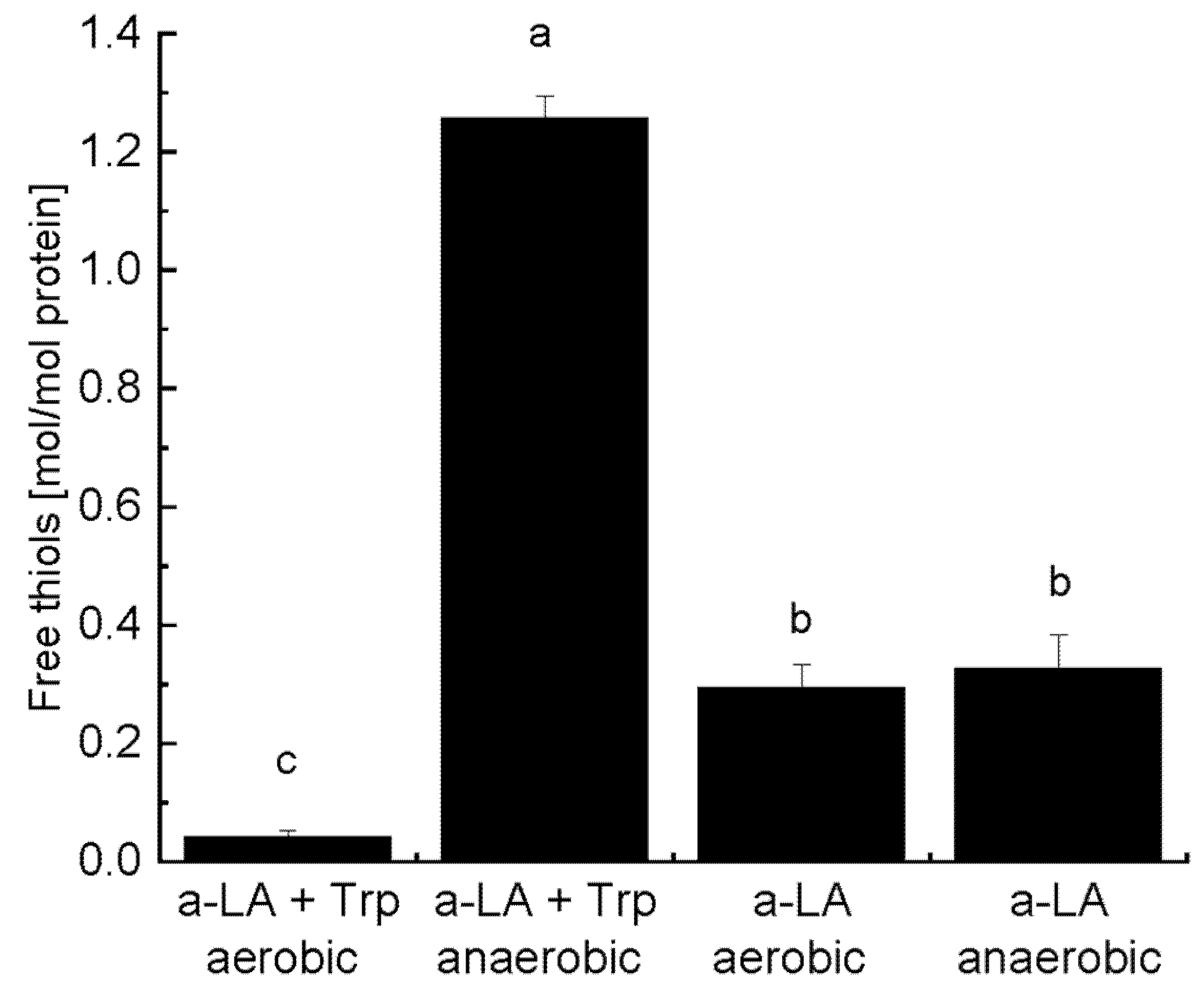
2.5. Quantification of Free Thiols by 4,4′-Dithiodipyridine (4-DPS) Derivatization
2.6. Quantification of Oxidation Products from Trp, Tyr, and Phe
2.7. Statistical Analysis
3. Results and Discussion
3.1. UV-B-Induced Aggregation of Apo-α-LA in the Absence and Presence of Free Trp and Oxygen
3.2. Formation of Reducible Covalent Cross-Links and Release of Free Thiols during UV-B Illumination
3.3. Formation of Oxidation Products from Aromatic Amino Acids after UV-B Light Treatment
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Díaz, O.; Pereira, C.D.; Cobos, A. Functional properties of ovine whey protein concentrates produced by membrane technology after clarification of cheese manufacture by-products. Food Hydrocoll. 2004, 18, 601–610. [Google Scholar] [CrossRef]
- Ryan, K.N.; Foegeding, E.A. Formation of soluble whey protein aggregates and their stability in beverages. Food Hydrocoll. 2015, 43, 265–274. [Google Scholar] [CrossRef]
- Bryant, C.M.; McClements, D.J. Molecular basis of protein functionality with special consideration of cold-set gels derived from heat- denatured whey. Trends Food Sci. Technol. 1998, 9, 143–151. [Google Scholar] [CrossRef]
- De Wit, J.N. Nutritional and Functional Characteristics of Whey Proteins in Food Products. J. Dairy Sci. 1998, 81, 597–608. [Google Scholar] [CrossRef]
- White, S.S.; Fox, K.M.; Jervis, S.M.; Drake, M.A. Influence of heating and acidification on the flavor of whey protein isolate. J. Dairy Sci. 2013, 96, 1366–1379. [Google Scholar] [CrossRef]
- Jansson, T.; Nielsen, S.B.; Petersen, M.A.; Lund, M.N. Temperature-dependency of unwanted aroma formation in reconstituted whey protein isolate solutions. Int. Dairy J. 2020, 104, 104653. [Google Scholar] [CrossRef]
- Zhao, Z.; Engholm-Keller, K.; Poojary, M.M.; Boelt, S.G.; Rogowska-Wrzesinska, A.; Skibsted, L.H.; Davies, M.J.; Lund, M.N. Generation of Aggregates of α-Lactalbumin by UV-B Light Exposure. J. Agric. Food Chem. 2020, 68, 6701–6714. [Google Scholar] [CrossRef] [PubMed]
- Vanaman, T.C.; Brew, K.H.R. The disulfide bonds of bovine alpha-lactalbumin. J. Biol. Chem. 1970, 245, 4583–4590. [Google Scholar] [CrossRef]
- Gammelgaard, S.K.; Petersen, S.B.; Haselmann, K.F.; Nielsen, P.K. Direct Ultraviolet Laser-Induced Reduction of Disulfide Bonds in Insulin and Vasopressin. ACS Omega 2020, 5, 7962–7968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asquith, R.S.; Hirst, L. The photochemical degradation of cystine in aqueous solution in the presence of air. BBA Gen. Subj. 1969, 184, 345–357. [Google Scholar] [CrossRef]
- Vanhooren, A.; Devreese, B.; Vanhee, K.; Van Beeumen, J.; Hanssens, I. Photoexcitation of tryptophan groups induces reduction of two disulfide bonds in goat α-lactalbumin. Biochemistry 2002, 41, 11035–11043. [Google Scholar] [CrossRef]
- Pattison, D.I.; Rahmanto, A.S.; Davies, M.J. Photo-oxidation of proteins. Photochem. Photobiol. Sci. 2012, 11, 38–53. [Google Scholar] [CrossRef]
- Correia, M.; Neves-Petersen, M.T.; Parracino, A.; Di Gennaro, A.K.; Petersen, S.B. Photophysics, photochemistry and energetics of UV light induced disulphide bridge disruption in apo-α-lactalbumin. J. Fluoresc. 2012, 22, 323–337. [Google Scholar] [CrossRef]
- Bent, D.V.; Hayon, E. Excited state chemistry of aromatic amino acids and related peptides. III. Tryptophan. J. Am. Chem. Soc. 1975, 97, 2612–2619. [Google Scholar] [CrossRef] [PubMed]
- Bent, D.V.; Hayon, E. Excited state chemistry of aromatic amino acids and related peptides. I. Tyrosine. J. Am. Chem. Soc. 1975, 97, 2599–2606. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Poojary, M.M.; Skibsted, L.H.; Lund, M.N.; Lund, M.N. Cleavage of Disulfide Bonds in Cystine by UV-B Illumination Mediated by Tryptophan or Tyrosine as Photosensitizers. J. Agric. Food Chem. 2020, 68, 6900–6909. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.R.; Nielsen, S.B.; Zhao, Z.; Olsen, K.; Nielsen, J.H.; Lund, M.N. Control of α-Lactalbumin Aggregation by Modulation of Temperature and Concentration of Calcium and Cysteine. J. Agric. Food Chem. 2018, 66, 7110–7120. [Google Scholar] [CrossRef]
- Zainudin, M.A.M.; Poojary, M.M.; Jongberg, S.; Lund, M.N. Light exposure accelerates oxidative protein polymerization in beef stored in high oxygen atmosphere. Food Chem. 2019, 299, 125132. [Google Scholar] [CrossRef]
- Gomyo, T.; Fujimaki, M. Studies on Changes of Protein by Dye Sensitized Photooxidation. Agric. Biol. Chem. 1970, 34, 302–309. [Google Scholar] [CrossRef]
- Haywood, J.; Mozziconacci, O.; Allegre, K.M.; Kerwin, B.A.; Schöneich, C. Light-induced conversion of trp to gly and gly hydroperoxide in IgG1. Mol. Pharm. 2013, 10, 1146–1150. [Google Scholar] [CrossRef]
- Neves-Petersen, M.T.; Klitgaard, S.; Pascher, T.; Skovsen, E.; Polivka, T.; Yartsev, A.; Sundström, V.; Petersen, S.B. Flash photolysis of cutinase: Identification and decay kinetics of transient intermediates formed upon UV excitation of aromatic residues. Biophys. J. 2009, 97, 211–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dose, K. The photolysis of cystine in the presence of aromatic amino acids. Photochem. Photobiol. 1968, 7, 671–673. [Google Scholar] [CrossRef]
- Creed, D. The photophysics and photochemistry of the near-uv absorbing amino acids-III. Cystine and its simple derivatives. Photochem. Photobiol. 1984, 39, 577–583. [Google Scholar] [CrossRef]
- Kristo, E.; Hazizaj, A.; Corredig, M. Structural changes imposed on whey proteins by UV irradiation in a continuous UV light reactor. J. Agric. Food Chem. 2012, 60, 6204–6209. [Google Scholar] [CrossRef]
- Catalfo, A.; Bracchitta, G.; De Guidi, G. Role of aromatic amino acid tryptophan UVA-photoproducts in the determination of drug photosensitization mechanism: A comparison between methylene blue and naproxen. Photochem. Photobiol. Sci. 2009, 8, 1467–1475. [Google Scholar] [CrossRef]
- Hellwig, M. The Chemistry of Protein Oxidation in Food. Angew. Chem. Int. Ed. 2019, 58, 16742–16763. [Google Scholar] [CrossRef]
- Dalsgaard, T.K.; Otzen, D.; Nielsen, J.H.; Larsen, L.B. Changes in structures of milk proteins upon photo-oxidation. J. Agric. Food Chem. 2007, 55, 10968–10976. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oxidation Product | Apo-α-LA Separated from Trp | Apo-α-LA Separated from Trp | Free Trp Separated from Apo-α-LA | Free Trp Separated from Apo-α-LA | LOD (µM) |
|---|---|---|---|---|---|
| Aerobic | Anaerobic | Aerobic | Anaerobic | ||
| 3-OH-Kyn | 56 ± 7 | 56 ± 4 | nd | nd | 2.2 |
| NFK | nd | nd | nd | nd | 1.2 |
| Kyn | nd | nd | 58 ± 2 | nd | 1.0 |
| 5-OH-Trp | nd | nd | 11 ± 6 | nd | 5.2 |
| Tryptamine | nd | nd | 2 ± 0 | 3 ± 1 | 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Li, R.; Poojary, M.M.; Nielsen, S.B.; Lund, M.N. Effect of Addition of Tryptophan on Aggregation of Apo-α-Lactalbumin Induced by UV-Light. Foods 2021, 10, 1577. https://doi.org/10.3390/foods10071577
Zhao Z, Li R, Poojary MM, Nielsen SB, Lund MN. Effect of Addition of Tryptophan on Aggregation of Apo-α-Lactalbumin Induced by UV-Light. Foods. 2021; 10(7):1577. https://doi.org/10.3390/foods10071577
Chicago/Turabian StyleZhao, Zichen, Renjie Li, Mahesha M. Poojary, Søren B. Nielsen, and Marianne N. Lund. 2021. "Effect of Addition of Tryptophan on Aggregation of Apo-α-Lactalbumin Induced by UV-Light" Foods 10, no. 7: 1577. https://doi.org/10.3390/foods10071577