
Effects of Linseed Meal and Carotenoids from Different Sources on Egg Characteristics, Yolk Fatty Acid and Carotenoid Profile and Lipid Peroxidation

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Design
2.3. Dietary Treatments
2.4. Performance Parameters
2.5. Sampling Collection and Procedures
2.6. Color Measurement
2.7. Egg Yolk Carotenoid Analysis
2.8. Egg Yolk Fatty Acid Analysis and Lipid Quality Indices
2.9. Lipid Oxidative Status of the Yolk
2.10. Yolk Cholesterol Content
2.11. Statistical Analyses
3. Results
3.1. Proximate Composition, Fatty Acid and Carotenoid Profile of Dried Kapia Pepper, Sea Buckthorn Pomace and Carrot
3.2. Performance Parameters
3.3. External and Internal Egg Quality Parameters
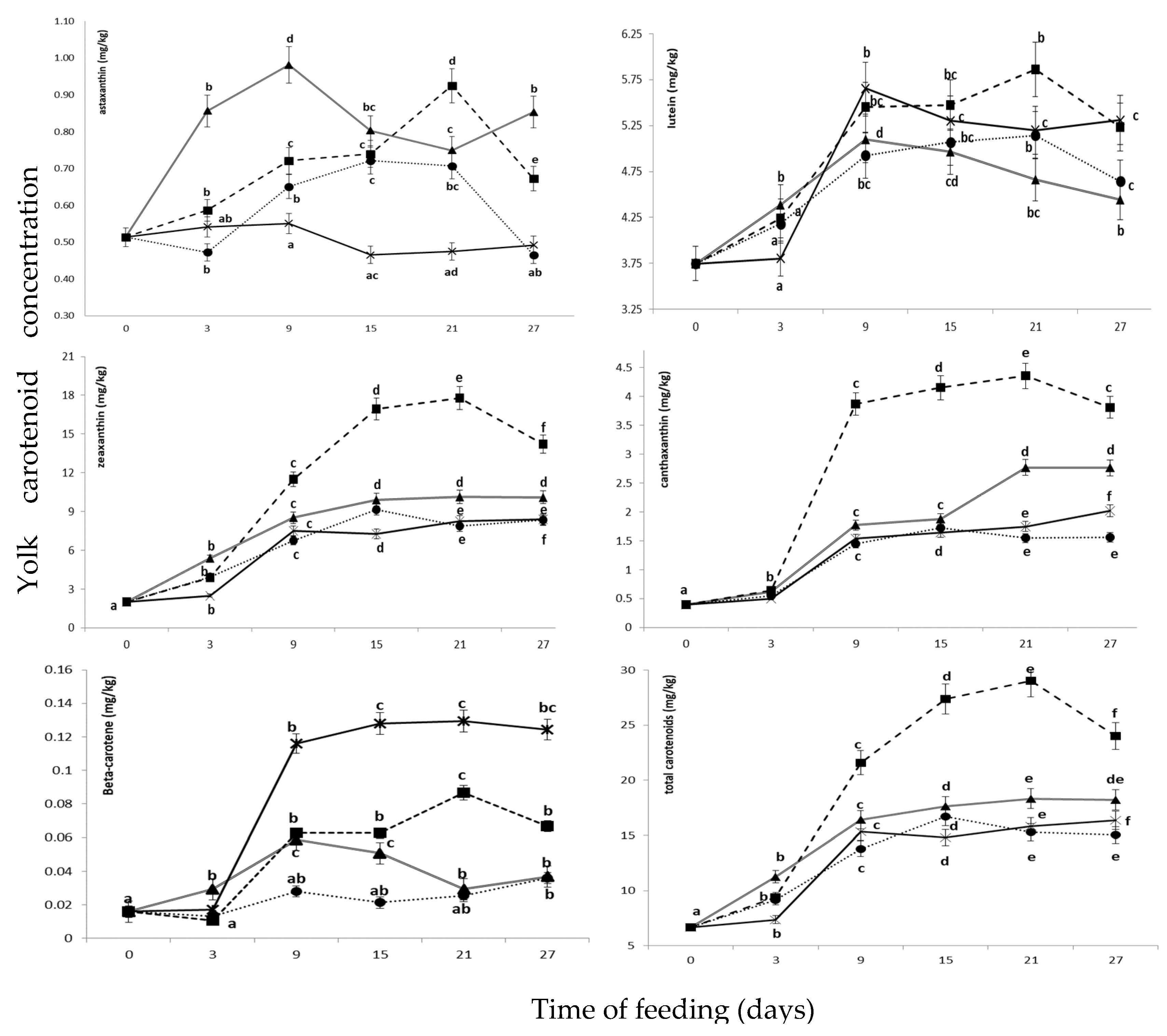
3.4. Yolk Color and Carotenoid Concentration in Eggs Yolk
3.5. Egg Yolk Fatty Acids Profile
3.6. Egg Yolk Cholesterol Content and Lipid Oxidation
4. Discussion
4.1. Chemical Composition
4.2. Performance Parameters
4.3. External and Internal Egg Quality Parameters
4.4. Yolk Color and Carotenoid Content
4.5. Egg Yolk Fatty Acids Profile
4.6. Cholesterol Content and Lipid Oxidative Status of the Yolk
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Omri, B.; Chalghoumi, R.; Izzo, L.; Ritieni, A.; Lucarini, M.; Durazzo, A.; Abdouli, H.; Santini, A. Effect of dietary incorporation of linseed alone or together with tomato-red pepper mix on laying hens’ egg yolk fatty acids profile and health lipid indexes. Nutrients 2019, 11, 813. [Google Scholar] [CrossRef] [Green Version]
- Mapiye, C.; Chimonyo, M.; Dzama, K.; Hugo, A.; Strydom, P.E.; Muchenje, V. Fatty acid composition of beef from Nguni Steers supplemented with Acacia karroo leaf-meal. J. Food Compos. Anal. 2011, 24, 523–528. [Google Scholar] [CrossRef]
- Attia, Y.A.; Al-Harthi, M.A.; Korish, M.A.; Shiboob, M.M. Fatty acid and cholesterol profiles, hypocholesterolemic, atherogenic, and thrombogenic indices of broiler meat in the retail market. Lipids Health Dis. 2017, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Campos, J.; Severino, P.; Ferreira, C.; Zielinska, A.; Santini, A.; Souto, S.B.; Souto, E.B. Linseed essential oil—Source of lipids as active ingredients for pharmaceuticals and nutraceuticals. Curr. Med. Chem. 2018, 26, 4537–4558. [Google Scholar] [CrossRef] [PubMed]
- Kljak, K.; Carović-Stanko, K.; Kos, I.; Janječić, Z.; Kiš, G.; Duvnjak, M.; Safner, T.; Bedeković, D. Plant carotenoids as pigment sources in laying hen diets: Effect on yolk color, carotenoid content, oxidative stability and sensory properties of eggs. Foods 2021, 10, 721. [Google Scholar] [CrossRef] [PubMed]
- Altuntaş, A.; Aydin, R. Fatty acid composition of egg yolk from chickens fed a diet including marigold (Tagetes erecta L.). J. Lipids 2014, 2014, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Kang, S.K.; Heo, Y.J.; Shin, D.W.; Park, T.E.; Han, G.G.; Jin, G.-D.; Lee, H.-B.; Jung, E.; Kim, H.S.; et al. Influence of flaxseed oil on fecal microbiota, egg quality and fatty acid composition of egg yolks in laying hens. Curr. Microbiol. 2016, 72, 259–266. [Google Scholar] [CrossRef]
- Simopoulos, A. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Surai, P.F.; Simons, P.C.M.; Dvorska, J.E.; Aradas, F.; Sparks, N.H.C. Antioxidant-enriched eggs: Opportunities and limitations. In The Amazing Egg: Nature’s Perfect Functional Food for Health Promotion; Sim, J.S., Sunwoo, H.H., Eds.; University of Alberta: Edmonton, AB, Canada, 2006; pp. 68–93. [Google Scholar]
- Panaite, T.D.; Nour, V.; Vlaicu, P.A.; Ropota, M.; Corbu, A.R.; Saracila, M. Flaxseed and dried tomato waste used together in laying hens diet. Arch. Anim. Nutr. 2019, 73, 222–238. [Google Scholar] [CrossRef]
- Loetscher, Y.; Kreuzer, M.; Messikommer, R.E. Late laying hens deposit dietary antioxidants preferentially in the egg and not in the body. J. Appl. Poultry Res. 2014, 23, 647–660. [Google Scholar] [CrossRef]
- Marounek, M.; Pebriansyah, A. Use of carotenoids in feed mixtures for poultry: A review. Agric. Trop. Subtrop. 2018, 51, 107–111. [Google Scholar] [CrossRef]
- Ciccone, M.M.; Cortese, F.; Gesualdo, M.; Carbonara, S.; Zito, A.; Ricci, G.; De Pascalis, F.; Scicchitano, P.; Riccioni, G. Dietary intake of carotenoids and their antioxidant and anti-inflammatory effects in cardiovascular care. Mediat. Inflamm. 2013, 782137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria, A.G.; Graziano, R.; Nicolantonio, D.O. Carotenoids: Potential allies of cardiovascular health? Food Nutr. Res. 2015, 59, 26762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spasevski, N.J.; Dragojlović, D.M.; Čolović, D.S.; Vidosavljević, S.Ž.; Peulić, T.A.; Rakita, S.M.; Kokić, B.M. Influence of dietary carrot and paprika on egg physical characteristics and yolk color. Food Feed Res. 2018, 45, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Tkacz, K.; Wojdyło, A.; Turkiewicz, I.P.; Bobak, Ł.; Nowicka, P. Anti-oxidant and anti-enzymatic activities of sea buckthorn (Hippophaë rhamnoides L.) fruits modulated by chemical components. Antioxidants 2019, 8, 618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özer, A.; Erdost, H.; Zõk, B. Histological investigations on the effects of feeding a diet containing red hot pepper on the reproductive organs of the chicken. Phytother. Res. 2005, 19, 501–505. [Google Scholar] [CrossRef]
- Al-Kassie, G.A.M.; Mamdooh, A.M.A.; Saba, J.A. The effects of using hot red pepper as a diet supplement on some performance traits in broiler. Pak. J. Nutr. 2011, 9, 842–845. [Google Scholar] [CrossRef]
- Cervantes-Paz, B.; Yahia, E.M.; de Ornelas-Paz, J.J.; VictoriaCampos, C.I.; Ibarra-Junquera, V.; Pérez-Martínez, J.D.; Escalante-Minakata, P. Antioxidant activity and content of chlorophylls and carotenoids in raw and heat-processed Jalapeno peppers at intermediate stages of ripening. Food Chem. 2014, 146, 188–196. [Google Scholar] [CrossRef]
- Arimboor, R.; Natarajan, R.B.; Menon, K.R.; Chandrasekhar, L.P.; Moorkoth, V. Red pepper (Capsicum annuum) carotenoids as a source of natural food colors: Analysis and stability-a review. J. Food Sci. Technol. Mys. 2015, 52, 1258–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palevitch, D.; Craker, L.E. Nutritional and medicinal importance of red pepper (Capsicum spp.). J. Herbs Spices Med. Plants 1995, 3, 55–83. [Google Scholar] [CrossRef]
- Daood, H.G.; Vinkler, M.; Markus, F.; Hebshi, E.A.; Biacs, P.A. Antioxidant vitamin content of spice red pepper (paprika) as affected by technological and varietal factors. Food Chem. 1996, 55, 365–372. [Google Scholar] [CrossRef]
- Özer, A.; Erdost, H.; Zık, B.; Özfiliz, N. Histological investigations on the effects of feeding with a diet containing red hot pepper on the reproductive system organs of the cock. Turk. J. Vet. Anim. Sci. 2006, 30, 7–15. [Google Scholar]
- Lokaewmanee, K.; Yamauchi, K.; Okuda, N. Effects of dietary red pepper on egg yolk colour and histological intestinal morphology in laying hens. J. Anim. Physiol. Anim. Nutr. 2013, 97, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M.; Nakajima, S.; Miyagawa, S.; Nakawaga, J.; Okumura, J. Feeding behavior, abdominal fat and laying performance in laying hens given diets containing red pepper. Jpn. Poult. Sci. 1994, 31, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.D.; Karki, S.; Thakur, N.S.; Attri, S. Chemical composition, functional properties and processing of carrot—A review. J. Food Sci. Technol. Mys. 2012, 49, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B. Sugars, acids, ethyl β-D-glucopyranose and a methyl inositol in sea buckthorn (Hippophae rhamnoides) berries. Food Chem. 2009, 112, 89–97. [Google Scholar] [CrossRef]
- Bal, L.M.; Meda, V.; Naik, S.N.; Satya, S. Sea buckthorn berries: A potential source of valuable nutrients for nutraceuticals and cosmoceuticals. Food Res. Int. 2011, 44, 1718–1727. [Google Scholar] [CrossRef]
- Michel, T.; Destandau, E.; Le Floch, G.; Lucchesi, M.E.; Elfakir, C. Antimicrobial, antioxidant and phytochemical investigations of sea buckthorn (Hippophae rhamnoides L.) leaf, stem, root and seed. Food Chem. 2012, 131, 754–760. [Google Scholar] [CrossRef]
- Nour, V.; Panaite, T.D.; Corbu, A.R.; Ropota, M.; Turcu, R.P. Nutritional and bioactive compounds in dried sea-buckthorn pomace. Erwerbs-Obstbau 2021, 63, 91–98. [Google Scholar] [CrossRef]
- Dumbrava, D.; Matiuti, M.; Druga, M.; Lupea, A.; Ianculov, I.; Clep, C. Effect of seabuckthorn berry flour from hens food on egg yolk carotenoidic pigments content. Ann. Facul. Eng. Hunedoara. 2006, 4, 156–160. [Google Scholar]
- Shaker, M.M.; Al-Beitawi, N.A.; Bláha, J.; Mahmoud, Z. The effect of sea buckthorn (Hippophae rhamnoides L.) fruit residues on performance and egg quality of laying hens. J. Appl. Anim. Res. 2018, 46, 422–426. [Google Scholar] [CrossRef] [Green Version]
- Association of Official Analytical Chemists (AOAC). Official Method of Analysis, 13th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2000. [Google Scholar]
- European Parliament, Council of the European Union. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes; Council of Europe: Strasbourg, France, 2010. [Google Scholar]
- Lohmann. Lohmann Brown Management Guide; March 2005; Lohmann Tierzucht GmbH: Cuxhaven, Germany, 2005. [Google Scholar]
- European Council Directive. Certain Marketing Standards for Eggs; Chapter II: Grades of eggs. Article 7: Grading of Grade A Eggs of Regulation (EC) No 2295/2003; Comission European Communities: Brussels, Belgium, 2006. [Google Scholar]
- Safaa, H.M.; Jiménez-Moreno, E.; Valencia, D.G.; Frikha, M.; Serrano, M.P.; Mateos, G.G. Effect of main cereal of the diet and particle size of the cereal on productive performance and egg quality of brown egg laying hens in early phase of production. Poult. Sci. 2009, 88, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Laudadio, V.; Ceci, E.; Lastella, N.M.B.; Introna, M.; Tufarelli, V. Low-fiber alfalfa (Medicago sativa L.) meal in the laying hen diet: Effects on productive traits and egg quality. Poult. Sci. 2014, 93, 1868–1874. [Google Scholar] [CrossRef]
- Untea, A.E.; Varzaru, I.; Panaite, T.D.; Gavris, T.; Lupu, A.; Ropota, M. The effects of dietary inclusion of bilberry and walnut leaves in laying hens’ diets on the antioxidant properties of eggs. Animals 2020, 10, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Association of Official Analytical Chemists (AOAC). Official Method 994.10: Cholesterol in Foods-Direct Saponification-Gas Chromatographic Methods; Association of Official Analytical Chemists: Arlington, VA, USA, 1996. [Google Scholar]
- Suresh, P.V.; Raj, K.R.; Nidheesh, T.; Pal, G.K.; Sakhare, P.Z. Application of chitosan for improvement of quality and shelf life of table eggs under tropical room conditions. J. Food Sci. Technol. Mys. 2015, 52, 6345–6354. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, P.; Suchý, P.; Straková, E.; Doležalová, J. The effect of a diet supplemented with sea-buckthorn pomace on the colour and viscosity of the egg yolk. Acta Vet. Brno 2017, 86, 303–308. [Google Scholar] [CrossRef] [Green Version]
- Hammershøj, M.; Kidmose, U. Coloured carrot varieties as forage material for egg layers. World. Poult. Sci. J. 2006, 62, S168–S170. [Google Scholar]
- Hammershøj, M.; Kidmose, U.; Steenfeldt, S. Deposition of carotenoids in egg yolk by short-term supplement of coloured carrot (Daucus carota) varieties as forage material for egg-laying hens. J. Sci. Food Agric. 2010, 90, 1163–1171. [Google Scholar] [CrossRef] [Green Version]
- Bozalan, K.N.; Karadeniz, F. Carotenoid profile, total phenolic content, and antioxidant activity of carrots. Int. J. Food Prop. 2011, 14, 1060–1068. [Google Scholar] [CrossRef]
- Søltoft, M.; Bysted, A.; Madsen, K.H.; Mark, A.B.; Bügel, S.G.; Nielsen, J.; Knuthsen, P. Effects of organic and conventional growth systems on the content of carotenoids in carrot roots, and on intake and plasma status of carotenoids in humans. J. Sci. Food Agric. 2011, 91, 767–775. [Google Scholar] [CrossRef]
- Koka, N.; Karadeniz, F. Changes of bioactive compounds and anti-oxidant activity during cold storage of carrots. Int. J. Food Sci. Tech. 2008, 43, 2019–2025. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Martínez-Guirado, C.; del Mar Rebolloso-Fuentes, M.; Carrique-Pérez, A. Nutrient composition and antioxidant activity of 10 pepper (Capsicum annuun) varieties. Eur. Food Res. Technol. 2006, 224, 1–9. [Google Scholar] [CrossRef]
- Teleszko, M.; Wojdylo, A.; Rudzinska, M.; Oszmianski, J.; Golis, T. Analysis of lipophilic and hydrophilic bioactive compounds content in sea buckthorn (Hippophae rhamnoides L.) berries. J. Agric. Food Chem. 2015, 63, 4120–4129. [Google Scholar] [CrossRef] [PubMed]
- Aaby, K.; Martinsen, B.K.; Borge, G.I.; Røen, D. Bioactive compounds and color of sea buckthorn (Hippophae rhamnoides L.) purees as affected by heat treatment and high-pressure homogenization. Int. J. Food Prop. 2020, 23, 651–664. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Jin, L.; Wu, F.; Thacker, P.; Li, X.; You, J.; Wang, X.; Liu, S.; Li, S.; Xu, Y. Effect of red pepper (Capsicum frutescens) powder or red pepper pigment on the performance and egg yolk color of laying hens. Asian Aust. J. Anim. Sci. 2012, 25, 1605–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou-Elkhair, R.; Selim, S.; Hussein, E. Effect of supplementing layer hen diet with phytogenic feed additives on laying performance, egg quality, egg lipid peroxidation and blood biochemical constituents. Anim. Nutr. 2018, 4, 399–400. [Google Scholar] [CrossRef]
- Chand, N.; Naz, S.; Irfan, M.; Khan, R.U.; Rehman, Z.U. Effect of sea buckthorn (Hippophae rhamnoides L.) seed supplementation on egg quality and cholesterol of Rhode Island Red × Fayoumi laying hens. Korean J. Food. Sci. Anim. Res. 2018, 38, 468–475. [Google Scholar] [CrossRef]
- Gurbuz, Y.; Yasar, S.; Karaman, M. Effects of addition of the red pepper from 4th harvest to corn or wheat based diets on egg-yolk colour and egg production in laying hens. Int. J. Poult. Sci. 2003, 2, 107–111. [Google Scholar] [CrossRef]
- Moeini, M.M.; Ghazi, S.H.; Sadeghi, S.; Malekizadeh, M. The effect of red pepper (Capsicum annuum) and marigold flower (Tageteserectus) powder on egg production, egg yolk color and some blood metabolites of laying hens. Iran. J. Appl. Anim. Sci. 2013, 3, 301–305. [Google Scholar]
- Hamilton, P.B.; Tirado, J.F.; Garcia-Hernandez, F. Deposition in egg yolks of the carotenoids from saponified and unsaponified oleoresin of red pepper (Capsicum annuum). Poult. Sci. 1990, 69, 462–470. [Google Scholar] [CrossRef]
- Bohn, T. Carotenoids and markers of oxidative stress in human observational studies and intervention trials: Implications for chronic diseases. Antioxidants 2019, 8, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böhm, V.; Lietz, G.; Olmedilla-Alonso, B.; Phelan, D.; Reboul, E.; Bánati, D.; Borel, P.; Corte-Real, J.; de Lera, A.R.; Desmarchelier, C.; et al. From carotenoid intake to carotenoid blood and tissue concentrations—Implications for dietary intake recommendations. Nutr. Rev. 2021, 79, 544–573. [Google Scholar] [CrossRef] [PubMed]
- Na, J.C.; Song, J.Y.; Lee, B.D.; Lee, S.J.; Lee, C.Y.; An, G.H. Effect of polarity on absorption and accumulation of carotenoids by laying hens. Anim. Feed Sci. Technol. 2004, 117, 305–315. [Google Scholar] [CrossRef]
- Van Elswyk, M. Comparison of n–3 fatty acid sources in laying hen rations for improvement of whole egg nutritional quality: A review. Br. J. Nutr. 1997, 78, S61–S69. [Google Scholar] [CrossRef] [Green Version]
- Roth-Maier, D.A.; Eder, K.; Kirchgessner, M. Live performance and fatty acid composition of meat in broiler chickens fed diets with various amounts of ground or whole flaxseed. J. Anim. Physiol. Anim. Nutr. 1998, 79, 260–268. [Google Scholar] [CrossRef]
- Sytařová, I.; Orsavová, J.; Snopek, L.; Mlček, J.; Byczńnski, L.; Mišurcová, L. Impact of phenolic compounds and vitamins C and E on antioxidant activity of sea buckthorn (Hippophae rhamnoides L.) berries and leaves of diverse ripening times. Food Chem. 2020, 310, 125784. [Google Scholar] [CrossRef]
- Nguyen, Q.V.; Malau-Aduli, B.S.; Cavalieri, J.; Nichols, P.D.; Malau-Aduli, A.E.O. Enhancing omega-3 long-chain polyunsaturated fatty acid content of dairy-derived foods for human consumption. Nutrients 2019, 11, 743. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Joint FAO/WHO Expert Consultation on Fats and Fatty Acids in Human Nutrition; WHO: Geneva, Switzerland, 2008; Available online: http://www.who.int/nutrition/topics/FFA_interim_recommendations/en/ (accessed on 10 May 2021).
- Anderson, B.M.; Ma, D.W. Are all n-3 polyunsaturated fatty acids created equal? Lipids Health Dis. 2009, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Kones, R.; Howell, S.; Rumana, U. n-3 Polyunsaturated Fatty Acids and Cardiovascular Disease: Principles, Practices, Pitfalls, and Promises—A Contemporary Review. Med. Princ. Pract. 2017, 26, 497–508. [Google Scholar] [CrossRef]
- Duru, M.; Duru Arslan, A.; Karadas, K.; Eyduran, E.; Cinli, H.; Tariq, M.M. Effect of carrot (Daucus carota) leaf powder on external and internal egg characteristics of Hy-Line White laying hens. Pak. J. Zool. 2017, 49, 125–132. [Google Scholar] [CrossRef]
- Sahin, N.; Akdemir, F.; Orhan, C.; Kucuk, O.; Hayirli, A.; Sahin, K. Lycopene-enriched quail egg as functional food for humans. Food Res. Int. 2008, 41, 295–300. [Google Scholar] [CrossRef]
- Skřivan, M.; Marounek, M.; Englmaierová, M.; Skřivanová, E. Effect of increasing doses of marigold (Tagetes erecta) flower extract on eggs carotenoids content, colour and oxidative stability. J. Anim. Feed Sci. 2016, 25, 58–64. [Google Scholar] [CrossRef]
- Young, A.J.; Lowe, G.L. Carotenoids–Antioxidant Properties. Antioxidants 2018, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Akdemir, F.; Orhan, C.; Sahin, N.; Sahin, K.; Hayirli, A. Tomato powder in laying hen diets: Effects on concentrations of yolk carotenoids and lipid peroxidation. Br. Poult. Sci. 2012, 53, 675–680. [Google Scholar] [CrossRef]
- An, B.K.; Choo, W.D.; Kang, C.W.; Lee, J.; Lee, K.W. Effects of dietary lycopene or tomato paste on laying performance and serum lipids in laying hens and on malondialdehyde content in egg yolk upon storage. J. Poult. Sci. 2019, 56, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamed, M.; Kalita, D.; Bartolo, M.E.; Jayanty, S.S. Capsaicinoids, polyphenols and antioxidant activities of Capsicum annuum: Comparative study of the effect of ripening stage and cooking methods. Antioxidants 2019, 8, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chávez-Mendoza, C.; Sanchez, E.; Muñoz-Marquez, E.; Sida-Arreola, J.P.; Flores-Cordova, M.A. Bioactive compounds and antioxidant activity in different grafted varieties of bell pepper. Antioxidants 2015, 4, 427–446. [Google Scholar] [CrossRef] [Green Version]
- Yeum, K.J.; Beretta, G.; Krinsky, N.I.; Russell, R.M.; Aldini, G. Synergistic interactions of antioxidant nutrients in a biological model system. Nutrition 2009, 25, 839–846. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Items | Experimentl Diets 1 | |||
|---|---|---|---|---|
| Control (C) | Kapia Pepper (E1) | Sea Buckthorn Pomace (E2) | Carrot (E3) | |
| Ingredients, % | ||||
| Corn | 50.00 | 50.00 | 50.00 | 50.00 |
| Soybean meal | 29.52 | 20.57 | 19.91 | 21.30 |
| Sunflower meal | 4.22 | 7.21 | 8.00 | 6.49 |
| Vegetable oil | 3.50 | 1.31 | 1.16 | 1.3 |
| Linseed meal | - | 6.00 | 6.00 | 6.00 |
| Kapia pepper (dehydrated) | - | 2.00 | - | - |
| Sea buckthorn pomace (dehydrated) | - | - | 2.00 | - |
| Carrot (dehydrated) | - | - | - | 2.00 |
| DL-methionine | 0.08 | 0.17 | 0.17 | 0.17 |
| L-lysine | 0.02 | 0.02 | 0.04 | 0.02 |
| Calcium carbonate | 9.81 | 9.85 | 9.85 | 9.85 |
| Monocalcium phosphate | 1.40 | 1.42 | 1.42 | 1.42 |
| Salt | 0.40 | 0.40 | 0.40 | 0.40 |
| Choline premix | 0.05 | 0.05 | 0.05 | 0.05 |
| Vitamin-mineral premix 2 | 1.00 | 1.00 | 1.00 | 1.00 |
| Calculated analysis, % | ||||
| Metabolizable energy (MJ/kg) | 2780.00 | 2780.00 | 2780.00 | 2780.00 |
| Crude protein | 17.50 | 17.10 | 17.20 | 17.10 |
| Crude fat | 4.20 | 4.70 | 4.83 | 4.69 |
| Chemical analysis, % | ||||
| Fatty acids composition (% of total fat) | ||||
| ΣSFA | 28.81 | 22.30 | 25.02 | 25.82 |
| ΣMUFA | 34.82 | 29.79 | 32.93 | 34.04 |
| ∑ PUFA, from which: | 35.91 | 47.60 | 41.89 | 39.83 |
| Σ n-3 PUFA | 2.67 | 10.63 | 9.29 | 7.84 |
| ∑ n-6 PUFA | 33.24 | 36.98 | 32.60 | 31.99 |
| ∑ n-6/∑ n-3 | 12.45 | 3.48 | 3.51 | 4.08 |
| Carotenoid profile (mg/kg) | ||||
| Astaxanthin | 0.122 | 0.129 | 0.121 | 0.124 |
| Lutein | 0.309 | 0.429 | 0.329 | 0.314 |
| Zeaxanthin | 0.333 | 0.385 | 0.465 | 0.323 |
| Canthaxanthin | 0.125 | 0.154 | 0.127 | 0.128 |
| β-carotene | 0.056 | 0.218 | 0.231 | 0.719 |
| Total carotenoid, mg/kg | 0.945 | 1.315 | 1.273 | 1.608 |
| Nutrients | Kapia Pepper | Buckthorn Pomace | Carrot |
|---|---|---|---|
| Proximate composition (%) | |||
| Dry matter | 91.31 ± 3.78 | 94.60 ± 4.42 | 88.29 ± 3.19 |
| Crude protein | 9.34 ± 0.35 | 15.42 ± 0.68 | 6.11 ± 0.28 |
| Ether extract | 1.34 ± 0.08 | 15.08 ± 0.56 | 0.92 ± 0.04 |
| Crude fibre | 9.15 ± 0.38 | 30.84 ± 1.23 | 7.03 ± 0.44 |
| Ash | 5.38 ± 0.28 | 1.94 ± 0.12 | 6.27 ± 0.26 |
| Fatty acids composition (% of total fat) | |||
| ΣSFA | 27.72 ± 1.21 | 34.28 ± 1.57 | 30.09 ± 1.39 |
| ΣMUFA | 14.13 ± 0.68 | 57.10 ± 2.66 | 23.42 ± 1.07 |
| ∑PUFA, from which: | 58.04 ± 2.78 | 8.31 ± 0.46 | 46.28 ± 1.89 |
| Σ n-3 PUFA | 12.48 ± 0.45 | 2.74 ± 0.22 | 2.99 ± 0.18 |
| ∑ n-6 PUFA | 45.56 ± 2.18 | 5.57 ± 0.25 | 43.29 ± 1.82 |
| ∑ n-6/∑n-3 | 3.65 ± 0.23 | 2.04 ± 0.14 | 14.50 ± 0.65 |
| Carotenoid profile (mg/kg) | |||
| Astaxanthin | 0.501 ± 0.04 | 0.478 ± 0.03 | 0.147 ± 0.01 |
| Lutein | 5.631 ± 0.29 | 0.696 ± 0.05 | 0.237 ± 0.02 |
| Zeaxanthin | 1.528 ± 0.07 | 6.960 ± 0.36 | 0.031 ± 0.01 |
| Canthaxanthin | 0.971 ± 0.04 | 0.136 ± 0.01 | n.d. |
| Trans-apo-carotenal | 0.161 ± 0.01 | 0.236 ± 0.02 | 0.002 ± 0.001 |
| Lycopene | 0.351 ± 0.02 | 0.091 ± 0.01 | 0.259 ± 0.01 |
| β-carotene | 7.576 ± 0.34 | 8.076 ± 0.38 | 32.586 ± 1.46 |
| Total carotenoid content | 16.719 ± 0.75 | 16.673 ± 0.82 | 33.262 ± 1.51 |
| Items | Dietary Treatments | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| C | E1 | E2 | E3 | |||
| Initial body weight (g/hen) | 1951.00 | 1950.50 | 1958.57 | 1952.50 | 19.118 | 0.9988 |
| Final body weight (g/hen) | 1940.00 | 1968.42 | 1961.58 | 1930.53 | 19.943 | 0.8941 |
| Feed intake (g/day/hen) | 115.82 b | 115.96 b | 119.53 a | 120.96 a | 0.656 | 0.0075 |
| Feed conversion ratio (g feed/g egg) | 2.02 b | 2.07 ab | 2.13 a | 2.10 ab | 0.015 | 0.0670 * |
| Total carotenoid intake (mg/day/hen) | 0.11 c | 0.15 b | 0.15 b | 0.19 a | 0.006 | <0.0001 |
| Laying percentage (%) | 91.51 | 91.75 | 90.89 | 92.70 | 0.480 | 0.6139 |
| Total eggs/period (eggs) | 1150 | 1156 | 1134 | 1169 | ||
| Egg size classification ** | ||||||
| Extra-large egg (XL), >73 g (%) | 4.29 b | 3.41 b | 4.35 b | 6.25 a | 0.261 | 0.0003 |
| Large egg (L), 63–73 g (%) | 50.57 b | 38.44 a | 57.25 c | 49.43 b | 0.862 | <0.0001 |
| Medium egg (M), 53–63 g (%) | 45.65 b | 56.62 a | 39.25 c | 43.93 b | 0.831 | <0.0001 |
| Small egg (S), <53 g (%) | 2.65 b | 5.13 a | 2.71 b | 3.28 b | 0.273 | 0.0005 |
| Egg Weight and Components (g) | Fresh Egg (Value) | Shell Quality (mm; kgF) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Whole Egg | Albumen | Yolk | Eggshell | Albumen pH | Yolk pH | Haugh Unit | Eggshell Thickness | Eggshell Breaking Strength | |
| 2 weeks | |||||||||
| C | 62.67 | 38.26 | 16.02 | 8.39 | 8.29 | 5.79 c | 74.60 a | 0.34 bc | 3.96 |
| E1 | 60.82 | 36.69 | 15.7 | 8.43 | 8.29 | 5.97 a | 75.92 a | 0.34 abc | 4.27 |
| E2 | 61.02 | 36.84 | 15.88 | 8.3 | 8.38 | 5.92 b | 75.82 a | 0.36 ab | 4.30 |
| E3 | 60.93 | 36.36 | 15.94 | 8.62 | 8.29 | 6.00 a | 76.13 a | 0.34 bc | 3.90 |
| 4 weeks | |||||||||
| C | 61.09 | 37.00 | 15.84 | 8.25 | 8.41 | 5.81 c | 62.22 b | 0.33 c | 3.90 |
| E1 | 61.56 | 37.17 | 15.72 | 8.59 | 8.27 | 5.90 b | 65.59 b | 0.36 ab | 3.98 |
| E2 | 62.55 | 37.36 | 16.50 | 8.66 | 8.30 | 5.90 b | 63.13 b | 0.36 ab | 3.95 |
| E3 | 60.80 | 36.47 | 15.56 | 8.51 | 8.30 | 5.90 b | 61.87 b | 0.37 a | 3.63 |
| SEM | 0.565 | 0.545 | 0.294 | 0.164 | 16.139 | 0.006 | 1.981 | 0.006 | 0.167 |
| Main effects | |||||||||
| Diet | |||||||||
| C | 61.88 | 37.63 | 15.93 | 8.32 | 8.35 | 5.80 c | 68.41 | 0.33 b | 3.93 |
| E1 | 61.19 | 36.93 | 15.71 | 8.51 | 8.28 | 5.94 a | 70.75 | 0.35 a | 4.13 |
| E2 | 61.78 | 37.10 | 16.19 | 8.48 | 8.34 | 5.91 b | 69.47 | 0.35 a | 4.12 |
| E3 | 60.87 | 36.42 | 15.75 | 8.56 | 8.30 | 5.95 a | 69.00 | 0.36 a | 3.77 |
| Time | |||||||||
| 2 weeks | 61.36 | 37.04 | 15.89 | 8.43 | 8.31 | 5.92 a | 75.62 a | 0.345 b | 4.109 a |
| 4 weeks | 61.50 | 37.00 | 15.91 | 8.50 | 19.75 | 5.88 b | 63.20 b | 0.354 a | 3.867 b |
| p-Value | |||||||||
| Diet | 0.2255 | 0.1732 | 0.3477 | 0.488 | 0.3959 | <0.0001 | 0.678 | 0.0002 | 0.0998 |
| Time | 0.7232 | 0.9232 | 0.9234 | 0.5552 | 0.3182 | <0.0001 | <0.0001 | 0.0306 | 0.0432 |
| Diet × Time | 0.0436 | 0.3269 | 0.3663 | 0.3631 | 0.3947 | <0.0001 | 0.8003 | 0.0083 | 0.8224 |
| Parameter | Yolk Roche Color Fan Score | L* | a* | b* | C | h |
|---|---|---|---|---|---|---|
| 2 weeks | ||||||
| C | 5.94 c | 71.87 ab | −1.96 d | 40.36 b | 39.16 c | 93.14 a |
| E1 | 14.94 a | 61.577 c | 18.42 a | 46.86 ab | 50.35 a | 68.56 d |
| E2 | 7.50 b | 69.34 ab | 2.39 c | 45.59 ab | 45.68 abc | 87.17 bc |
| E3 | 7.50 b | 75.25 a | −1.92 d | 43.24 ab | 43.27 abc | 91.34 a |
| 4 weeks | ||||||
| C | 5.89 c | 69.81 ab | −1.32 d | 39.09 b | 40.40 c | 91.36 a |
| E1 | 15.00 a | 61.20 c | 18.33 a | 46.17 ab | 49.68 ab | 68.36 d |
| E2 | 6.22 c | 69.22 ab | 4.41 b | 49.21 a | 49.41 ab | 84.91 c |
| E3 | 6.33 c | 66.78 bc | −1.40 d | 41.68 ab | 41.74 bc | 90.16 ab |
| SEM | 0.156 | 1.484 | 0.409 | 1.853 | 1.8721 | 0.718 |
| Main effects | ||||||
| Diet | ||||||
| C | 5.92 c | 70.84 a | −1.64 c | 39.73 b | 39.78 b | 92.25 a |
| E1 | 14.97 a | 61.39 b | 18.37 a | 46.51 a | 50.02 a | 68.46 c |
| E2 | 6.92 b | 69.28 a | 3.40 b | 47.40 a | 47.54 a | 86.04 b |
| E3 | 6.86 b | 71.02 a | −1.66 c | 42.46 ab | 42.51 b | 90.75 a |
| Time | ||||||
| 2 weeks | 8.97 a | 69.51 a | 4.23 b | 43.70 | 44.62 | 85.05 a |
| 4 weeks | 8.36 b | 66.75 b | 5.00 a | 44.36 | 45.31 | 83.70 b |
| p-Value | ||||||
| Diet | <0.0001 | <0.0001 | <0.0001 | 0.0004 | <0.0001 | <0.0001 |
| Time | <0.0001 | 0.0122 | 0.0106 | 0.6167 | 0.603 | 0.0108 |
| Diet × Time | <0.0001 | 0.0249 | 0.0839 | 0.5176 | 0.5125 | 0.5212 |
| Item | SFA | MUFA | PUFA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n-6 | n-3 | |||||||||||
| C14:0 | C16:0 | C18:0 | C14:1 | C16:1 | C18:1 | C18:2n6 | C20:2n6 | C20:4n6 | C18:3n3 | C22:5n3 | C22:6n3 | |
| 2 weeks | ||||||||||||
| C | 0.32 a | 23.36 ab | 11.00 bc | 0.05 abc | 2.77 cd | 33.04 a | 20.34 a | 0.13 | 3.86 abc | 0.80 c | 0.14 b | 2.15 b |
| E1 | 0.25 bc | 22.76 b | 11.05 bc | 0.06 ab | 3.04 bcd | 33.81 a | 17.79 b | 0.13 | 3.62 bc | 1.68 ab | 0.27 a | 3.44 a |
| E2 | 0.25 bc | 23.19 ab | 11.16 abc | 0.05 ab | 3.59 a | 34.16 a | 17.02 b | 0.12 | 3.48 bc | 1.82 a | 0.20 ab | 3.17 a |
| E3 | 0.27 b | 23.17 ab | 10.55 c | 0.06 a | 3.51 ab | 34.26 a | 17.97 b | 0.14 | 3.33 c | 1.71 ab | 0.24 a | 3.12 a |
| 4 weeks | ||||||||||||
| C | 0.27 b | 23.62 ab | 12.53 a | 0.04 c | 2.15 e | 30.25 b | 21.15 a | 0.12 | 4.31 a | 0.74 c | 0.14 b | 2.32 b |
| E1 | 0.25 bc | 23.72 a | 11.67 abc | 0.05 abc | 2.87 cd | 33.24 a | 17.79 b | 0.09 | 3.65 bc | 1.55 ab | 0.26 a | 3.20 a |
| E2 | 0.24 bc | 23.03 ab | 11.57 abc | 0.04 abc | 3.21 abc | 33.24 a | 18.045 b | 0.10 | 3.65 bc | 1.57 ab | 0.23 ab | 3.23 a |
| E3 | 0.22 c | 23.03 ab | 12.01 ab | 0.04 bc | 2.70 d | 33.03 a | 18.10 b | 0.12 | 4.02 ab | 1.46 b | 0.26 a | 3.26 a |
| SEM | 0.008 | 0.004 | 0.302 | 0.004 | 0.108 | 0.441 | 0.322 | 0.014 | 0.127 | 0.070 | 0.020 | 0.1246 |
| Main effects | ||||||||||||
| Diet | ||||||||||||
| C | 0.29 a | 23.49 | 11.76 | 0.04 | 2.46 c | 31.65 b | 20.75 a | 0.14 b | 4.09 a | 0.77 b | 0.14 b | 2.23 b |
| E1 | 0.25 b | 23.24 | 11.37 | 0.05 | 2.96 b | 33.53 a | 17.79 b | 0.18 a | 3.64 b | 1.61 a | 0.26 a | 3.31 a |
| E2 | 0.24 b | 23.11 | 11.36 | 0.05 | 3.39 a | 33.70 a | 17.53 b | 0.17 ab | 3.56 b | 1.70 a | 0.22 a | 3.20 a |
| E3 | 0.25 b | 23.10 | 11.28 | 0.05 | 3.11 ab | 33.64 a | 18.03 b | 0.13 c | 3.68 b | 1.59 a | 0.25 a | 3.19 a |
| Time | ||||||||||||
| 2 weeks | 0.27 a | 23.12 | 10.94 b | 0.05 a | 3.23 a | 33.82 a | 18.28 b | 0.16 | 3.57 b | 1.57 a | 0.21 | 2.97 |
| 4 weeks | 0.24 b | 23.35 | 11.95 a | 0.04 b | 2.73 b | 33.44 b | 18.77 a | 0.15 | 3.91 a | 1.40 b | 0.22 | 3.00 |
| p-Value | ||||||||||||
| Diet | <0.0001 | 0.0363 | 0.3865 | 0.0967 | <0.0001 | <0.0001 | <0.0001 | 0.3673 | 0.0008 | <0.0001 | <0.0001 | <0.0001 |
| Time | <0.0001 | 0.9116 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0359 | 0.0872 | 0.0006 | 0.001 | 0.5921 | 0.7129 |
| Diet × Time | 0.0069 | 0.9159 | 0.1603 | 0.4694 | 0.028 | 0.0756 | 0.3157 | 0.7799 | 0.0548 | 0.4648 | 0.6387 | 0.3424 |
| Item | ΣSFA | ΣMUFA | ΣUFA | ΣPUFA | of Which: | Σn6/Σn3 | ΣSFA/ΣUFA | ΣPUFA/ΣMUFA | |
|---|---|---|---|---|---|---|---|---|---|
| Σn3 | Σn6 | ||||||||
| 2 weeks | |||||||||
| C | 34.91 b | 36.42 a | 65.09 ab | 28.67 b | 3.30 b | 25.36 a | 7.73 a | 0.54 bc | 0.79 b |
| E1 | 34.31 b | 37.59 a | 65.52 ab | 27.94 bc | 5.61 a | 22.33 bc | 3.99 b | 0.52 bc | 0.75 bc |
| E2 | 34.82 b | 38.36 a | 65.05 ab | 26.68 c | 5.43 a | 22.45 bc | 3.92 b | 0.53 bc | 0.70 c |
| E3 | 34.20 b | 38.41 a | 65.78 a | 27.37 bc | 5.32 a | 22.05 bc | 4.03 b | 0.52 c | 0.72 bc |
| 4 weeks | |||||||||
| C | 36.67 a | 33.09 b | 63.27 c | 30.18 a | 3.44 b | 26.74 a | 7.79 a | 0.58 a | 0.91 a |
| E1 | 35.85 ab | 36.66 a | 64.05 bc | 27.39 bc | 5.22 a | 22.17 bc | 4.26 b | 0.56 ab | 0.75 bc |
| E2 | 35.09 ab | 37.04 a | 64.77 abc | 27.73 bc | 5.28 a | 21.25 c | 4.25 b | 0.54 abc | 0.75 bc |
| E3 | 35.50 ab | 36.31 a | 64.47 abc | 28.15 bc | 5.26 a | 22.90 b | 4.37 b | 0.55 abc | 0.78 b |
| SEM | 0.368 | 0.502 | 0.365 | 0.326 | 0.118 | 0.310 | 0.147 | 0.009 | 0.018 |
| Main effects | |||||||||
| Diet | |||||||||
| C | 35.79 | 34.76 b | 64.18 | 29.42 a | 3.37 b | 26.05 a | 7.76 a | 0.56 | 0.85 a |
| E1 | 35.08 | 37.12 a | 64.79 | 27.66 b | 5.41 a | 22.25 b | 4.12 b | 0.54 | 0.75 b |
| E2 | 34.96 | 37.70 a | 64.91 | 27.21 b | 5.36 a | 21.85 b | 4.09 b | 0.54 | 0.72 b |
| E3 | 34.85 | 37.36 a | 65.12 | 27.76 b | 5.29 a | 22.47 b | 4.20 b | 0.54 | 0.75 b |
| Time | |||||||||
| 2 weeks | 34.56 b | 37.69 a | 65.36 a | 27.66 b | 4.92 | 22.75 b | 4.91 b | 0.53 b | 0.74 b |
| 4 weeks | 35.78 a | 35.77 b | 64.14 b | 28.36 a | 4.80 | 23.56 a | 5.17 a | 0.56 a | 0.80 a |
| p-Value | |||||||||
| Diet | 0.0622 | <0.0001 | 0.0773 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0664 | <0.0001 |
| Time | <0.0001 | <0.0001 | <0.0001 | 0.0042 | 0.1717 | 0.0006 | 0.0202 | <0.0001 | <0.0001 |
| Diet × Time | 0.2008 | 0.0992 | 0.1931 | 0.0199 | 0.183 | 0.0789 | 0.7489 | 0.1929 | 0.0138 |
| Item | Experimental Diets | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| C | E1 | E2 | E3 | |||
| Fat and cholesterol content of the yolk | ||||||
| Fat, % | 26.21 b | 29.16 a | 28.38 a | 28.00 a | 0.323 | 0.0081 |
| Cholesterol (g/100 g dried yolk) | 1.85 a | 1.68 b | 1.64 b | 1.63 b | 0.028 | 0.0081 |
| Lipid oxidative status of the yolk | ||||||
| PV (meq active oxygen/kg) | 0.34 b | 0.23 a | 0.23 a | 0.19 a | 0.018 | 0.0302 |
| CD (µmol/g) | 6.40 | 6.41 | 6.37 | 6.31 | 0.023 | 0.3962 |
| CT (µmol/g) | 2.44 bc | 2.46 b | 2.38 ab | 2.37 a | 0.012 | 0.0181 |
| TBARS (mg MDA/kg) | 0.17 b | 0.14 a | 0.16 ab | 0.14 a | 0.004 | 0.0358 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panaite, T.D.; Nour, V.; Saracila, M.; Turcu, R.P.; Untea, A.E.; Vlaicu, P.A. Effects of Linseed Meal and Carotenoids from Different Sources on Egg Characteristics, Yolk Fatty Acid and Carotenoid Profile and Lipid Peroxidation. Foods 2021, 10, 1246. https://doi.org/10.3390/foods10061246
Panaite TD, Nour V, Saracila M, Turcu RP, Untea AE, Vlaicu PA. Effects of Linseed Meal and Carotenoids from Different Sources on Egg Characteristics, Yolk Fatty Acid and Carotenoid Profile and Lipid Peroxidation. Foods. 2021; 10(6):1246. https://doi.org/10.3390/foods10061246
Chicago/Turabian StylePanaite, Tatiana D., Violeta Nour, Mihaela Saracila, Raluca P. Turcu, Arabela E. Untea, and Petru Al. Vlaicu. 2021. "Effects of Linseed Meal and Carotenoids from Different Sources on Egg Characteristics, Yolk Fatty Acid and Carotenoid Profile and Lipid Peroxidation" Foods 10, no. 6: 1246. https://doi.org/10.3390/foods10061246