
Cordyceps militaris: An Overview of Its Chemical Constituents in Relation to Biological Activity


Abstract
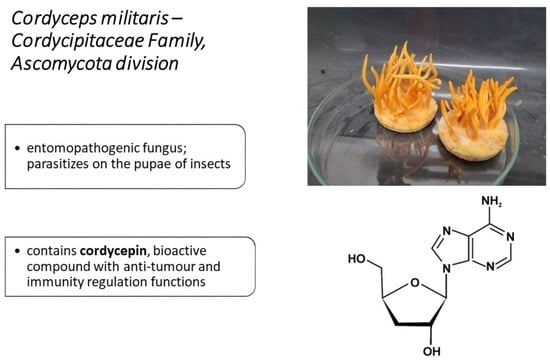
:
1. Introduction
2. Materials and Methods
3. Bioactive Compounds from C. militaris
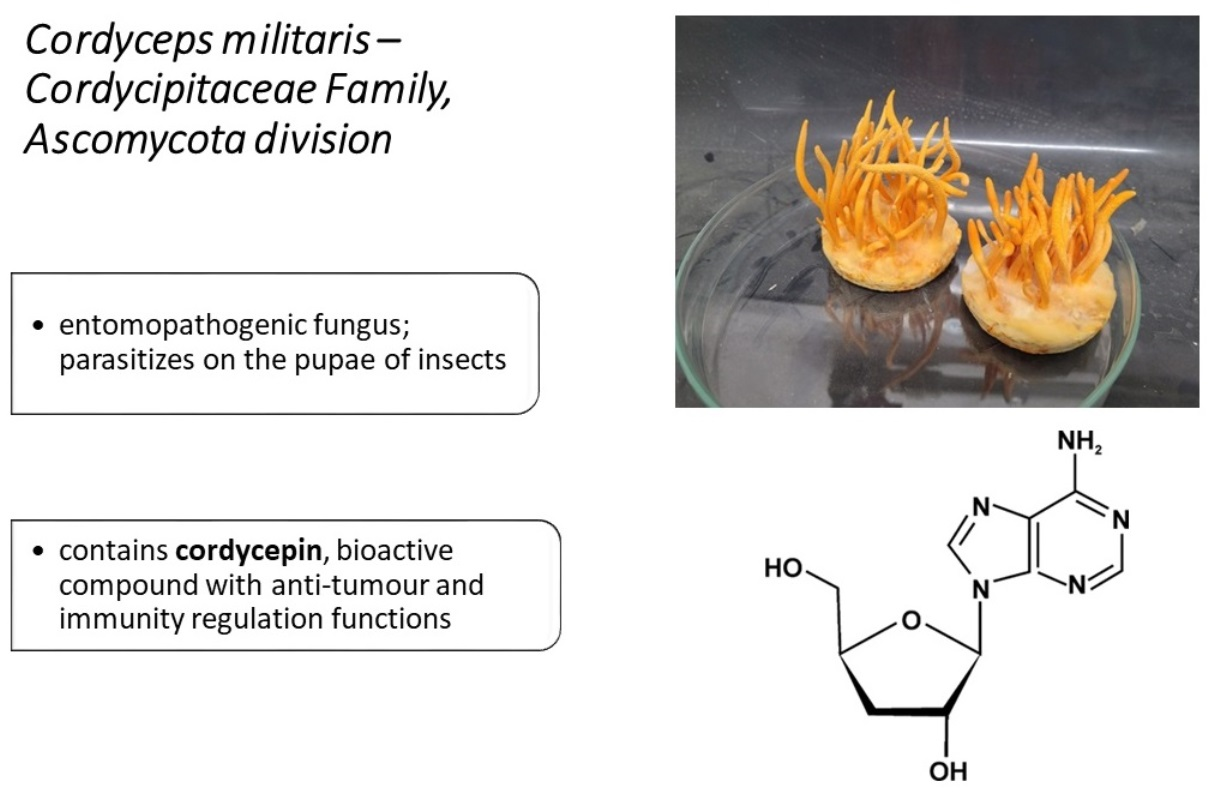
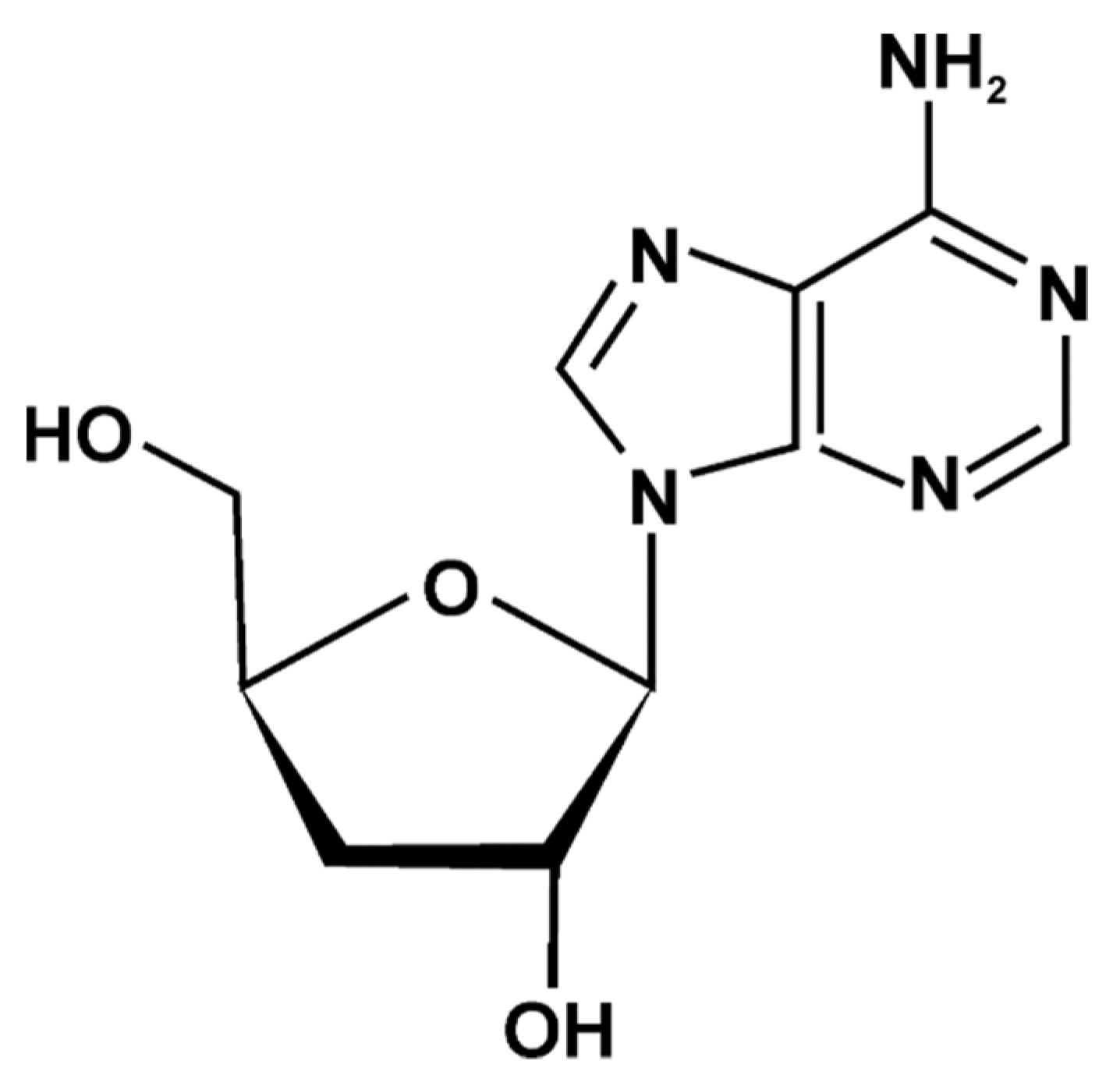
3.1. Nucleosides
3.2. Carbohydrates
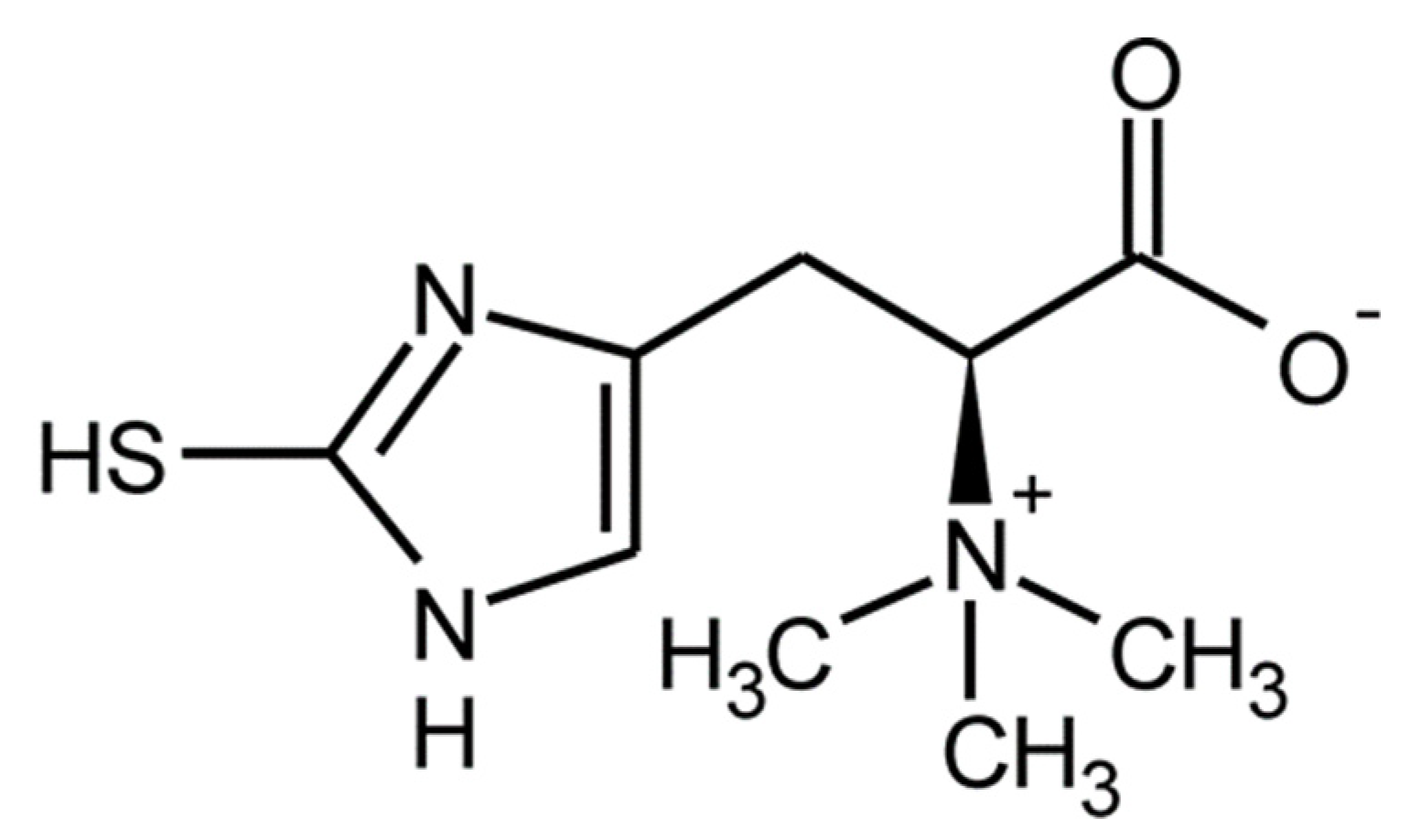
3.3. Amino Acids
3.4. Carotenoids
3.5. Statins
3.6. Phenolic Compounds
3.7. Other Bioactive Compounds in C. militaris Fruiting Bodies
4. Biological Activity
4.1. Ergogenic and Anti-Fatigue Activity
In Vivo Research
4.2. Immunostimulating Activity
4.2.1. In Vitro Research
4.2.2. In Vivo Research
4.3. Antitumor Activity
4.3.1. In Vitro Research
4.3.2. In Vivo Research
4.4. Antioxidant Activity
4.4.1. In Vitro Research
4.4.2. In Vivo Research
4.5. Anti-Inflammatory Activity
4.5.1. In Vitro Research
4.5.2. In Vivo Research
4.6. Hypoglycemic Activity
4.7. Antimicrobial Activity
4.8. Effect of C. militaris on the Endocrine System
4.9. Effect of C. militaris on the Respiratory System
4.10. Effect of C. militaris on the Locomotor System
4.11. Effect of C. militaris on the Nervous System
4.12. Effect of C. militaris on the Cardiovascular System
4.13. Other Biological Activity of C. militaris
5. Safety Assessment and Toxicology
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Das, S.K.; Masuda, M.; Sakurai, A.; Sakakibara, M. Medicinal Uses of the Mushroom Cordyceps militaris: Current State and Prospects. Fitoterapia 2010, 81, 961–968. [Google Scholar] [CrossRef]
- Holliday, J. Cordyceps: A Highly Coveted Medicinal Mushroom. In Medicinal Plants and Fungi: Recent Advances in Research and Development; Springer: Singapore, 2017; pp. 59–91. [Google Scholar]
- European Commission. Available online: https://ec.europa.eu/search/ (accessed on 15 August 2021).
- Reis, F.S.; Martins, A.; Vasconcelos, M.H.; Morales, P.; Ferreira, I.C.F.R. Functional Foods Based on Extracts or Compounds Derived from Mushrooms. Trends Food Sci. Technol. 2017, 66, 48–62. [Google Scholar] [CrossRef]
- Cheong, P.C.H.; Tan, C.S.; Fung, S.Y. Medicinal Mushrooms: Cultivation and Pharmaceutical Impact. In Biology of Macrofungi. Fungal Biology; Springer: Cham, switzerland, 2018; pp. 287–304. [Google Scholar]
- Wang, J.; Kan, L.; Nie, S.; Chen, H.; Cui, S.W.; Phillips, A.O.; Phillips, G.O.; Li, Y.; Xie, M. A Comparison of Chemical Composition, Bioactive Components and Antioxidant Activity of Natural and Cultured Cordyceps sinensis. LWT 2015, 63, 2–7. [Google Scholar] [CrossRef]
- Cohen, N.; Cohen, J.; Asatiani, M.D.; Varshney, V.K.; Yu, H.-T.; Yang, Y.-C.; Li, Y.-H.; Mau, J.-L.; Wasser, S.P. Chemical Composition and Nutritional and Medicinal Value of Fruit Bodies and Submerged Cultured Mycelia of Culinary-Medicinal Higher Basidiomycetes Mushrooms. Int. J. Med. Mushrooms 2014, 16, 273–291. [Google Scholar] [CrossRef]
- Chen, S.Y.; Ho, K.J.; Hsieh, Y.J.; Wang, L.T.; Mau, J.L. Contents of Lovastatin, γ-Aminobutyric Acid and Ergothioneine in Mushroom Fruiting Bodies and Mycelia. LWT 2012, 47, 274–278. [Google Scholar] [CrossRef]
- Chan, J.S.L.; Barseghyan, G.S.; Asatiani, M.D.; Wasser, S.P. Chemical Composition and Medicinal Value of Fruiting Bodies and Submerged Cultured Mycelia of Caterpillar Medicinal Fungus Cordyceps militaris CBS-132098 (Ascomycetes). Int. J. Med. Mushrooms 2015, 17, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.F.; Zhang, M.; Li, Z. Influence of Infrared Drying on the Drying Kinetics, Bioactive Compounds and Flavor of Cordyceps militaris. LWT 2019, 111, 790–798. [Google Scholar] [CrossRef]
- Tuli, H.S.; Sandhu, S.S.; Sharma, A.K. Pharmacological and Therapeutic Potential of Cordyceps with Special Reference to Cordycepin. 3 Biotech 2014, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Qin, P.; Li, X.; Yang, H.; Wang, Z.-Y.; Lu, D. Therapeutic Potential and Biological Applications of Cordycepin and Metabolic Mechanisms in Cordycepin-Producing Fungi. Molecules 2019, 24, 2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shawkat, H.; Westwood, M.-M.; Mortimer, A. Mannitol: A Review of Its Clinical Uses. BJA Edu. 2012, 12, 82–85. [Google Scholar] [CrossRef]
- Zhang, J.; Wen, C.; Duan, Y.; Zhang, H.; Ma, H. Advance in Cordyceps militaris (Linn) Link Polysaccharides: Isolation, Structure, and Bioactivities: A Review. Int. J. Biol. Macromol. 2019, 132, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kwon, J.S.; Won, D.P.; Lee, K.E.; Shin, W.C.; Hong, E.K. Study on Macrophage Activation and Structural Characteristics of Purified Polysaccharide from the Liquid Culture Broth of Cordyceps militaris. Carbohydr. Polym. 2010, 82, 982–988. [Google Scholar] [CrossRef]
- Chen, X.; Wu, G.; Huang, Z. Structural Analysis and Antioxidant Activities of Polysaccharides from Cultured Cordyceps militaris. Int. J. Biol. Macromol. 2013, 58, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Hu, S.; Liu, C.; Meng, Q.; Song, J.; Lu, J.; ChenG, Y.; Gao, C.; Liu, Y.; Wang, D.; et al. Purification of Polysaccharides from Cordyceps militaris and Their Anti-Hypoxic Effect. Mol. Med. Rep. 2015, 11, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.Y.; Guo, M.Z.; Liu, F.; Luo, Y.; Chen, L.; Meng, M.; Wang, X.T.; Zhang, Y.M. Preparation and Inhibition on α-D-Glucosidase of Low Molecular Weight Polysaccharide from Cordyceps militaris. Int. J. Biol. Macromol. 2016, 93, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.Y.; Liu, F.; Gao, H.; Sun, H.; Meng, M.; Zhang, Y.M. Synthesis, Characterization and Antioxidant Activity of Selenium Polysaccharide from Cordyceps militaris. Int. J. Biol. Macromol. 2016, 93, 1090–1099. [Google Scholar] [CrossRef]
- Luo, X.; Duan, Y.; Yang, W.; Zhang, H.; Li, C.; Zhang, J. Structural Elucidation and Immunostimulatory Activity of Polysaccharide Isolated by Subcritical Water Extraction from Cordyceps militaris. Carbohydr. Polym. 2017, 157, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Cui, X.; Chen, Z.; Huang, L.; Song, L.; Liu, T.; Lv, W.; Yu, R. Elucidation and Biological Activities of a New Polysaccharide from Cultured Cordyceps militaris. Carbohydr. Polym. 2014, 102, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Jing, Y.; Zhou, Q.; Hu, X.; Zhu, J.; Guo, Z.; Song, L.; Yu, R. Structural Elucidation and Immunostimulatory Activity of a New Polysaccharide from Cordyceps militaris. Food Funct. 2018, 9, 279–293. [Google Scholar] [CrossRef]
- Boonstra, E.; de Kleijn, R.; Colzato, L.S.; Alkemade, A.; Forstmann, B.U.; Nieuwenhuis, S. Neurotransmitters as Food Supplements: The Effects of GABA on Brain and Behavior. Front. Psychol. 2015, 6, 1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepsomali, P.; Groeger, J.A.; Nishihira, J.; Scholey, A. Effects of Oral Gamma-Aminobutyric Acid (GABA) Administration on Stress and Sleep in Humans: A Systematic Review. Front. Neurosci. 2020, 14, 923. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Z.; Liu, X.; Wang, Y.; Mao, F.; Mao, J.; Lu, X.; Jiang, D.; Wan, Y.; Lv, J.-Y.; et al. Study of GABA in Healthy Volunteers: Pharmacokinetics and Pharmacodynamics. Front. Pharmacol. 2015, 6, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamatsu, A.; Yamashita, Y.; Pandharipande, T.; Maru, I.; Kim, M. Effect of Oral γ-Aminobutyric Acid (GABA) Administration on Sleep and Its Absorption in Humans. Food Sci. Biotechnol. 2016, 25, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, B.; Kała, K.; Rojowski, J.; Grzywacz, A.; Opoka, W. Composition and Biological Properties of Agaricus bisporus Fruiting Bodies- a Review. Polish J. Food Nutr. Sci. 2017, 67, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Dubost, N.J.; Ou, B.; Beelman, R.B. Quantification of Polyphenols and Ergothioneine in Cultivated Mushrooms and Correlation to Total Antioxidant Capacity. Food Chem. 2007, 105, 727–735. [Google Scholar] [CrossRef]
- Weigand-Heller, A.J.; Kris-Etherton, P.M.; Beelman, R.B. The Bioavailability of Ergothioneine from Mushrooms (Agaricus bisporus) and the Acute Effects on Antioxidant Capacity and Biomarkers of Inflammation. Prev. Med. 2012, 54, S75–S78. [Google Scholar] [CrossRef] [PubMed]
- Borodina, I.; Kenny, L.C.; McCarthy, C.M.; Paramasivan, K.; Pretorius, E.; Roberts, T.J.; van der Hoek, S.A.; Kell, D.B. The Biology of Ergothioneine, an Antioxidant Nutraceutical. Nutr. Res. Rev. 2020, 33, 190–217. [Google Scholar] [CrossRef] [Green Version]
- Cheah, I.K.; Tang, R.M.Y.; Yew, T.S.Z.; Lim, K.H.C.; Halliwell, B. Administration of Pure Ergothioneine to Healthy Human Subjects: Uptake, Metabolism, and Effects on Biomarkers of Oxidative Damage and Inflammation. Antioxid. Redox Signal. 2017, 26, 193–206. [Google Scholar] [CrossRef]
- Sotgia, S.; Zinellu, A.; Mangoni, A.A.; Pintus, G.; Attia, J.; Carru, C.; McEvoy, M. Clinical and Biochemical Correlates of Serum L-Ergothioneine Concentrations in Community-Dwelling Middle-Aged and Older Adults. PLoS ONE 2014, 9, e84918. [Google Scholar] [CrossRef]
- Cheah, I.K.; Feng, L.; Tang, R.M.Y.; Lim, K.H.C.; Halliwell, B. Ergothioneine Levels in an Elderly Population Decrease with Age and Incidence of Cognitive Decline; a Risk Factor for Neurodegeneration? Biochem. Biophys. Res. Commun. 2016, 478, 162–167. [Google Scholar] [CrossRef]
- Dong, J.Z.; Wang, S.H.; Ai, X.R.; Yao, L.; Sun, Z.W.; Lei, C.; Wang, Y.; Wang, Q. Composition and Characterization of Cordyxanthins from Cordyceps militaris Fruit Bodies. J. Funct. Foods 2013, 5, 1450–1455. [Google Scholar] [CrossRef]
- Muszyńska, B.; Mastej, M.; Sułkowska-Ziaja, K. Biological Function of Carotenoids and Their Occurrence in the Fruiting Bodies of Mushrooms. MIR 2016, 107, 113–122. [Google Scholar]
- Bovier, E.R.; Hammond, B.R. A Randomized Placebo-Controlled Study on the Effects of Lutein and Zeaxanthin on Visual Processing Speed in Young Healthy Subjects. Arch. Biochem. Biophys. 2015, 572, 54–57. [Google Scholar] [CrossRef] [Green Version]
- Renzi-Hammond, L.; Bovier, E.; Fletcher, L.; Miller, L.; Mewborn, C.; Lindbergh, C.; Baxter, J.; Hammond, B. Effects of a Lutein and Zeaxanthin Intervention on Cognitive Function: A Randomized, Double-Masked, Placebo-Controlled Trial of Younger Healthy Adults. Nutrients 2017, 9, 1246. [Google Scholar] [CrossRef] [Green Version]
- Stringham, N.T.; Holmes, P.V.; Stringham, J.M. Supplementation with Macular Carotenoids Reduces Psychological Stress, Serum Cortisol, and Sub-Optimal Symptoms of Physical and Emotional Health in Young Adults. Nutr. Neurosci. 2018, 21, 286–296. [Google Scholar] [CrossRef]
- Watzl, B.; Bub, A.; Briviba, K.; Rechkemmer, G. Supplementation of a Low-Carotenoid Diet with Tomato or Carrot Juice Modulates Immune Functions in Healthy Men. Ann. Nutr. Metab. 2003, 47, 255–261. [Google Scholar] [CrossRef]
- Gul, K.; Tak, A.; Singh, A.K.; Singh, P.; Yousuf, B.; Wani, A.A. Chemistry, Encapsulation, and Health Benefits of β-Carotene-A Review. Cogent Food Agric. 2015, 1, 1018696. [Google Scholar] [CrossRef]
- Druesne-Pecollo, N.; Latino-Martel, P.; Norat, T.; Barrandon, E.; Bertrais, S.; Galan, P.; Hercberg, S. Beta-carotene Supplementation and Cancer Risk: A Systematic Review and Metaanalysis of Randomized Controlled Trials. Int. J. Cancer Res. 2010, 127, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Gajendragadkar, P.R.; Hubsch, A.; Mäki-Petäjä, K.M.; Serg, M.; Wilkinson, I.B.; Cheriyan, J. Effects of Oral Lycopene Supplementation on Vascular Function in Patients with Cardiovascular Disease and Healthy Volunteers: A Randomised Controlled Trial. PLoS ONE 2014, 9, e99070. [Google Scholar] [CrossRef] [PubMed]
- Senkus, K.E.; Tan, L.; Crowe-White, K.M. Lycopene and Metabolic Syndrome: A Systematic Review of the Literature. Adv. Nutr. 2019, 10, 19–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirahmadi, M.; Azimi-Hashemi, S.; Saburi, E.; Kamali, H.; Pishbin, M.; Hadizadeh, F. Potential Inhibitory Effect of Lycopene on Prostate Cancer. Biomed. Pharmacother. 2020, 129, 110459. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.; Sagar, A.; Kanwar, S.S.; Singh, S. Anticancer, Antibacterial and Antioxidant Activities of Cordyceps militaris. Indian J. Exp. Biol. 2019, 57, 15–20. [Google Scholar]
- Choi, E.-J.; Park, B.; Lee, J.; Kim, J. Anti-Atopic Dermatitis Properties of Cordyceps militaris on TNFα/IFNγ-Stimulated HaCaT Cells and Experimentally Induced Atopic Dermatitis in Mice. Phys. Act. Nutr. 2020, 24, 7. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Venturini, B.A.; Baptista, P.; Estevinho, L.M.; Ferreira, I.C.F.R. Chemical Composition and Biological Properties of Portuguese Wild Mushrooms: A Comprehensive Study. J. Agric. Food Chem. 2008, 56, 3856–3862. [Google Scholar] [CrossRef]
- Oesterle, A.; Laufs, U.; Liao, J.K. Pleiotropic Effects of Statins on the Cardiovascular System. Circ. Res. 2017, 120, 229–243. [Google Scholar] [CrossRef] [Green Version]
- Tobert, J.A. Efficacy and Long-Term Adverse Effect Pattern of Lovastatin. Am. J. Cardiol. 1988, 62, J28–J34. [Google Scholar] [CrossRef]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Ekiert, H. Phenolic Acids in Selected Edible Basidiomycota Species: Armillaria mellea, Boletus badius, Boletus edulis, Cantharellus cibarius, Lactarius deliciosus and Pleurotus ostreatus. Acta Sci. Pol. Hortorum Cultus 2013, 12, 107–116. [Google Scholar]
- Çayan, F.; Deveci, E.; Tel-Çayan, G.; Duru, M.E. Identification and Quantification of Phenolic Acid Compounds of Twenty-Six Mushrooms by HPLC–DAD. J. Food Meas. Charact. 2020, 14, 1690–1698. [Google Scholar] [CrossRef]
- Reis, F.S.; Barros, L.; Calhelha, R.C.; Ćirić, A.; van Griensven, L.J.L.D.; Soković, M.; Ferreira, I.C.F.R. The Methanolic Extract of Cordyceps militaris (L.) Link Fruiting Body Shows Antioxidant, Antibacterial, Antifungal and Antihuman Tumor Cell Lines Properties. Food Chem. Toxicol. 2013, 62, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.J.; Lin, C.P.; Mau, J.L.; Li, Y.S.; Tsai, S.Y. Effect of UV-B irradiation on physiologically active substance content and antioxidant properties of the medicinal caterpillar fungus Cordyceps militaris (Ascomycetes). Int J. Med. Mushrooms. 2015, 17, 241–253. [Google Scholar] [CrossRef]
- Awang, M.A.; Daud, N.N.N.N.M.; Ismail, N.I.M.; Cheng, P.G.; Ismail, M.F.; Ramaiya, S.D. Antioxidant and Cytotoxicity Activity of Cordyceps Militaris Extracts against Human Colorectal Cancer Cell Line. J App Pharm Sci. 2021, 11, 105–109. [Google Scholar]
- Singh, R.S.; Bhari, R.; Kaur, H.P. Mushroom Lectins: Current Status and Future Perspectives. Crit. Rev. Biotechnol. 2010, 30, 99–126. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Rouf, R.; Tiralongo, E.; May, T.; Tiralongo, J. Mushroom Lectins: Specificity, Structure and Bioactivity Relevant to Human Disease. Int. J. Mol. Sci. 2015, 16, 7802–7838. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Sun, Y.; Luo, F.; Wang, C. Bioactive Metabolites and Potential Mycotoxins Produced by Cordyceps Fungi: A Review of Safety. Toxins 2020, 12, 410. [Google Scholar] [CrossRef] [PubMed]
- Tsimberidou, A.; Giles, F.; Duvic, M.; Fayad, L.; Kurzrock MD, R. Phase II Study of Pentostatin in Advanced T-cell Lymphoid Malignancies: Update of an MD Anderson Cancer Center Series. Cancer 2004, 100, 342–349. [Google Scholar] [CrossRef]
- Wong, J.H.; Ng, T.B.; Wang, H.; Sze, S.C.W.; Zhang, K.Y.; Li, Q.; Lu, X. Cordymin, an Antifungal Peptide from the Medicinal Fungus Cordyceps militaris. Phytomed. 2011, 18, 387–392. [Google Scholar] [CrossRef]
- Commission Regulation (EU) No 432/2012 Commission Regulation (EU) No 432/2012 of 16 May 2012 Establishing a List of Permitted Health Claims Made on Foods, Other than Those Referring to the Reduction of Disease Risk and to Children’s Development and Health. OJEU 2012, 136, 1–40.
- Dong, J.Z.; Lei, C.; Ai, X.R.; Wang, Y. Selenium Enrichment on Cordyceps militaris Link and Analysis on Its Main Active Components. Appl. Biochem. 2012, 166. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.Z.; Ding, J.; Yu, P.Z.; Lei, C.; Zheng, X.J.; Wang, Y. Composition and Distribution of the Main Active Components in Selenium-Enriched Fruit Bodies of Cordyceps militaris Link. Food Chem. 2013, 137, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Liang, Y.; Zhao, G.; Wu, W.; Li, H.; Guo, Y. Selenium Biofortification and Antioxidant Activity in Cordyceps militaris Supplied with Selenate, Selenite, or Selenomethionine. Biol. Trace Elem. Res. 2019, 187, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Maughan, R.J.; Burke, L.M.; Dvorak, J.; Larson-Meyer, D.E.; Peeling, P.; Phillips, S.M.; Rawson, E.S.; Walsh, N.P.; Garthe, I.; Geyer, H. IOC Consensus Statement: Dietary Supplements and the High-Performance Athlete. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 104–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, M.C.; Cholewa, J.M.; Gerosa-Neto, J.; Gonçalves, D.C.; Caperuto, E.C.; Lira, F.S.; Rossi, F.E. A Single Dose of Oral ATP Supplementation Improves Performance and Physiological Response During Lower Body Resistance Exercise in Recreational Resistance-Trained Males. J. Strength Cond. Res. 2019, 33, 3345–3352. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-F. Effect of Polysaccharide from Cordyceps militaris (Ascomycetes) on Physical Fatigue Induced by Forced Swimming. Int. J. Med. Mushrooms 2016, 18, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Zhao, L.; Yang, F.; Yang, W.; Sun, Y.; Hu, Q. Evaluation of Anti-Fatigue Property of the Extruded Product of Cereal Grains Mixed with Cordyceps militaris on Mice. J. Int. Soc. Sports Nutr. 2017, 14, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Wang, Y.; Teng, M.; Cai, G.; Xu, H.; Guo, H.; Liu, Y.; Wang, D.; Teng, L. Studies on the Antifatigue Activities of Cordyceps militaris Fruit Body Extract in Mouse Model. ECAM 2015, 2015, 174616. [Google Scholar] [PubMed] [Green Version]
- Choi, E.; Oh, J.; Sung, G.-H. Beneficial Effect of Cordyceps militaris on Exercise Performance via Promoting Cellular Energy Production. Mycobiology 2020, 48, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, K.R.; Mock, M.G.; Roelofs, E.J.; Trexler, E.T.; Smith-Ryan, A.E. Chronic Supplementation of a Mushroom Blend on Oxygen Kinetics, Peak Power, and Time to Exhaustion. J. Int. Soc. Sports Nutr. 2015, 12, 1. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, K.R.; Smith-Ryan, A.E.; Roelofs, E.J.; Trexler, E.T.; Mock, M.G. Cordyceps militaris Improves Tolerance to High-Intensity Exercise after Acute and Chronic Supplementation. J. Diet. Suppl. 2017, 14, 42–53. [Google Scholar] [CrossRef]
- Dudgeon, W.D.; Thomas, D.D.; Dauch, W.; Scheett, T.P.; Webster, M.J. The Effects of High and Low-Dose Cordyceps militaris-Containing Mushroom Blend Supplementation After Seven and Twenty-Eight Days. Am. J. Sports Sci. 2018, 6, 1–7. [Google Scholar] [CrossRef]
- Compound Solutions, PeakO2®. Available online: https://compoundsolutions.com/ingredients/peako2/ (accessed on 15 August 2021).
- Zhu, S.; Pan, J.; Zhao, B.; Liang, J.; Ze-Yu, W.; Yang, J. Comparisons on Enhancing the Immunity of Fresh and Dry Cordyceps militaris in Vivo and in Vitro. J. Ethnopharmacol. 2013, 149, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-T.; Huang, K.-S.; Shaw, J.-F.; Chen, J.-R.; Kuo, W.-S.; Shen, G.; Grumezescu, A.M.; Holban, A.M.; Wang, Y.-T.; Wang, J.-S. Trends in the Immunomodulatory Effects of Cordyceps militaris: Total Extracts, Polysaccharides and Cordycepin. Front. Pharmacol 2020, 11, 1824. [Google Scholar] [CrossRef]
- Lee, J.S.; Kwon, J.S.; Yun, J.S.; Pahk, J.W.; Shin, W.C.; Lee, S.Y.; Hong, E.K. Structural Characterization of Immunostimulating Polysaccharide from Cultured Mycelia of Cordyceps militaris. Carbohydr. Polym. 2010, 80, 1011–1017. [Google Scholar] [CrossRef]
- Lee, J.S.; Hong, E.K. Immunostimulating Activity of the Polysaccharides Isolated from Cordyceps militaris. Int. Immunopharmacol. 2011, 11, 1226–1233. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kwon, D.S.; Lee, K.R.; Park, J.M.; Ha, S.-J.; Hong, E.K. Mechanism of Macrophage Activation Induced by Polysaccharide from Cordyceps militaris Culture Broth. Carbohydr. Polym. 2015, 120, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.-H.; Lee, C.-M.; Lee, S.-W.; Seo, S.-Y.; Seo, M.-J.; Kang, B.-W.; Jeong, Y.-K.; Choi, Y.-J.; Yang, K.-M.; Jo, W.-S. Cordycepin-Enriched Cordyceps militaris Induces Immunomodulation and Tumor Growth Delay in Mouse-Derived Breast Cancer. Oncol. Rep. 2013, 30, 1996–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.J.; Baik, H.W.; Kim, S.J.; Lee, S.G.; Ahn, H.Y.; Park, J.S.; Park, S.J.; Jang, E.J.; Park, S.W.; Choi, J.Y. Cordyceps militaris Enhances Cell-Mediated Immunity in Healthy Korean Men. J. Med. Food 2015, 18, 1164–1172. [Google Scholar] [CrossRef]
- Jung, S.J.; Hwang, J.H.; Oh, M.R.; Chae, S.W. Effects of Cordyceps militaris Supplementation on the Immune Response and Upper Respiratory Infection in Healthy Adults: A Randomized, Double-Blind, Placebo-Controlled Study. J. Nutr. Health 2019, 52, 258–267. [Google Scholar] [CrossRef]
- Yoo, H.-S.; Shin, J.; Cho, J.-H.; Son, C.-G.; Lee, Y.-W.; Park, S.; Cho, C.-K. Effects of Cordyceps militaris Extract on Angiogenesis and Tumor Growth. Acta Pharmacol. Sin. 2004, 25, 657–665. [Google Scholar]
- Jin, C.-Y.; Kim, G.-Y.; Choi, Y.-H. Induction of Apoptosis by Aqueous Extract of Cordyceps militaris through Activation of Caspases and Inactivation of Akt in Human Breast Cancer MDA-MB-231 Cells. J. Microbiol. Biotechnol. 2008, 18, 1997–2003. [Google Scholar] [PubMed]
- Park, S.E.; Yoo, H.S.; Jin, C.-Y.; Hong, S.H.; Lee, Y.-W.; Kim, B.W.; Lee, S.H.; Kim, W.-J.; Cho, C.K.; Choi, Y.H. Induction of Apoptosis and Inhibition of Telomerase Activity in Human Lung Carcinoma Cells by the Water Extract of Cordyceps militaris. Food Chem. Toxicol. 2009, 47, 1667–1675. [Google Scholar] [CrossRef]
- Rao, Y.K.; Fang, S.-H.; Wu, W.-S.; Tzeng, Y.-M. Constituents Isolated from Cordyceps militaris Suppress Enhanced Inflammatory Mediator’s Production and Human Cancer Cell Proliferation. J. Ethnopharmacol. 2010, 131, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Kim, W.; Moon, S. Cordycepin Suppresses TNF-alpha-induced Invasion, Migration and Matrix Metalloproteinase-9 Expression in Human Bladder Cancer Cells. Phytother. Res. 2010, 24, 1755–1761. [Google Scholar] [CrossRef]
- Yang, C.H.; Kao, Y.H.; Huang, K.S.; Wang, C.Y.; Lin, L.W. Cordyceps militaris and Mycelial Fermentation Induced Apoptosis and Autophagy of Human Glioblastoma Cells. Cell. Death Discov. 2012, 3, e431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruma, I.; Putranto, E.W.; Kondo, E.; Watanabe, R.; Saito, K.; Inoue, Y.; Yamamoto, K.-I.; Nakata, S.; Kaihata, M.; Murata, H. Extract of Cordyceps militaris Inhibits Angiogenesis and Suppresses Tumor Growth of Human Malignant Melanoma Cells. Int. J. Oncol. 2014, 45, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-C.; Zhu, Z.-Y.; Liu, Y.-L.; Sun, H.-Q. Comparisons of the Anti-Tumor Activity of Polysaccharides from Fermented Mycelia and Cultivated Fruiting Bodies of Cordyceps militaris in Vitro. Int. J. Biol. Macromol. 2019, 130, 307–314. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, W.; Dai, G.; Huang, Y. Cordycepin Suppresses the Migration and Invasion of Human Liver Cancer Cells by Downregulating the Expression of CXCR4. Int. J. Mol. 2020, 45, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Jo, E.; Jang, H.-J.; Yang, K.E.; Jang, M.S.; Huh, Y.H.; Yoo, H.-S.; Park, J.S.; Jang, I.-S.; Park, S.J. Cordyceps militaris Induces Apoptosis in Ovarian Cancer Cells through TNF-α/TNFR1-Mediated Inhibition of NF-ΚB Phosphorylation. BMC Complement. Med. Ther. 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Jo, E.; Jang, H.-J.; Shen, L.; Yang, K.E.; Jang, M.S.; Huh, Y.H.; Yoo, H.-S.; Park, J.; Jang, I.S.; Park, S.J. Cordyceps militaris Exerts Anticancer Effect on Non–Small Cell Lung Cancer by Inhibiting Hedgehog Signaling via Suppression of TCTN3. Integr. Cancer Ther. 2020, 19, 1–14. [Google Scholar] [CrossRef]
- He, Y.T.; Zhang, X.L.; Xie, Y.M.; Xu, Y.X.; Li, J.R. Extraction and Antioxidant Property in Vitro of Cordycepin in Artificially Cultivated Cordyceps militaris. Adv. Mater. Res. 2013, 750–752, 1593–1596. [Google Scholar] [CrossRef]
- Yu, H.M.; Wang, B.-S.; Huang, S.C.; Duh, P.-D. Comparison of Protective Effects between Cultured Cordyceps militaris and Natural Cordyceps sinensis against Oxidative Damage. J. Agric. Food Chem. 2006, 54, 3132–3138. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Yang, W.; Song, L.; Yan, C.; Zhang, Z.; Zhao, Y. Structural Characterization and Antioxidant Activity of a Polysaccharide from the Fruiting Bodies of Cultured Cordyceps militaris. Carbohydr. Polym. 2007, 70, 430–436. [Google Scholar] [CrossRef]
- Yu, R.; Yin, Y.; Yang, W.; Ma, W.; Yang, L.; Chen, X.; Zhang, Z.; Ye, B.; Song, L. Structural Elucidation and Biological Activity of a Novel Polysaccharide by Alkaline Extraction from Cultured Cordyceps militaris. Carbohydr. Polym. 2009, 75, 166–171. [Google Scholar] [CrossRef]
- Fengyao, W.; Hui, Y.; Xiaoning, M.; Junqing, J.; Guozheng, Z.; Xijie, G.; Zhongzheng, G. Structural Characterization and Antioxidant Activity of Purified Polysaccharide from Cultured Cordyceps militaris. Afr. J. Microbiol. Res. 2011, 5, 2743–2751. [Google Scholar] [CrossRef]
- Chen, R.; Jin, C.; Li, H.; Liu, Z.; Lu, J.; Li, S.; Yang, S. Ultrahigh Pressure Extraction of Polysaccharides from Cordyceps militaris and Evaluation of Antioxidant Activity. Sep. Purif. Technol. 2014, 134, 90–99. [Google Scholar] [CrossRef]
- Liu, J.; Feng, C.; Li, X.; Chang, M.; Meng, J.; Xu, L. Immunomodulatory and Antioxidative Activity of Cordyceps militaris Polysaccharides in Mice. Int. J. Biol. Macromol. 2016, 86, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Shrestha, B.; Lim, S.Y.; Yoon, D.H.; Chang, W.C.; Shin, D.-J.; Han, S.K.; Park, S.M.; Park, J.H.; Park, H. il Cordycepin Inhibits Lipopolysaccharide-Induced Inflammation by the Suppression of NF-ΚB through Akt and P38 Inhibition in RAW 264.7 Macrophage Cells. Eur. J. Pharmacol. 2006, 545, 192–199. [Google Scholar] [CrossRef]
- Jo, W.S.; Choi, Y.J.; Mm, H.J.; Lee, J.Y.; Nam, B.H.; Lee, J.D.; Lee, S.W.; Seo, S.Y.; Jeong, M.H. The Anti-Inflammatory Effects of Water Extract from Cordyceps militaris in Murine Macrophage. Mycobiology 2010, 38, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, J.-W.; Jin, C.-Y.; Kim, G.-Y.; Lee, J.-D.; Park, C.; Kim, G.-D.; Kim, W.-J.; Jung, W.-K.; Seo, S.K.; Choi, I.-W. Anti-Inflammatory Effects of Cordycepin via Suppression of Inflammatory Mediators in BV2 Microglial Cells. Int. Immunopharmacol. 2010, 10, 1580–1586. [Google Scholar] [CrossRef]
- Han, E.S.; Oh, J.Y.; Park, H.-J. Cordyceps militaris Extract Suppresses Dextran Sodium Sulfate-Induced Acute Colitis in Mice and Production of Inflammatory Mediators from Macrophages and Mast Cells. J. Ethnopharmacol. 2011, 134, 703–710. [Google Scholar] [CrossRef]
- Won, S.-Y.; Park, E.-H. Anti-Inflammatory and Related Pharmacological Activities of Cultured Mycelia and Fruiting Bodies of Cordyceps militaris. J. Ethnopharmacol. 2005, 96, 555–561. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Baggio, C.H.; Borato, D.G.; Santana-Filho, A.P.; Sassaki, G.L.; Iacomini, M.; van Griensven, L.J.L.D. Anti-Inflammatory Properties of the Medicinal Mushroom Cordyceps militaris Might Be Related to Its Linear (1→ 3)-β-D-Glucan. PLoS ONE 2014, 9, e110266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, C.-P.; Liu, S.-C.; Tang, C.-H.; Chan, Y.; El-Shazly, M.; Lee, C.-L.; Du, Y.-C.; Wu, T.-Y.; Chang, F.-R.; Wu, Y.-C. Anti-Inflammatory Cerebrosides from Cultivated Cordyceps militaris. J. Agric. Food Chem. 2016, 64, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Wang, Y.; Pang, J. Efficacy of Cordyceps militaris Capsules in Treatment of Chronic Bronchitis. Chin. J. New Drugs. 2004, 13, 171–174. [Google Scholar]
- Gai, G.; Jin, S.; Wang, B.; Li, Y.; Li, C. The Efficacy of Cordyceps militaris Capsules in Treatment of Chronic Bronchitis in Comparison with Jinshuibao Capsules. Chin. J. New Drugs. 2004, 13, 169–170. [Google Scholar]
- Sun, J.; Xu, J.; Wang, S.; Hou, Z.; Lu, X.; An, L.; Du, P. A New Cerebroside from Cordyceps militaris with Anti-PTP1B Activity. Fitoterapia 2019, 138, 104342. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Yu, X.; Li, T.; Zhu, Z. Structure and Hypoglycemic Activity of a Novel Exopolysaccharide of Cordyceps militaris. Int. J. Biol. Macromol. 2021, 166, 496–508. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, S.; Du, M. Cordycepin from Cordyceps militaris Prevents Hyperglycemia in Alloxan-Induced Diabetic Mice. Nutr. Res. 2015, 35, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Parunyakul, K.; Srisuksai, K.; Charoenlappanit, S.; Phaonakrop, N.; Roytrakul, S.; Fungfuang, W. Metabolic Impacts of Cordycepin on Hepatic Proteomic Expression in Streptozotocin-Induced Type 1 Diabetic Mice. PLoS ONE 2021, 16, e0256140. [Google Scholar] [CrossRef] [PubMed]
- Montefiori, D.C.; Sobol, R.W.; Li, S.W.; Reichenbach, N.L.; Suhadolnik, R.J.; Charubala, R.; Pfleiderer, W.; Modliszewski, A.; Robinson, W.E.; Mitchell, W.M. Phosphorothioate and Cordycepin Analogues of 2′, 5′-Oligoadenylate: Inhibition of Human Immunodeficiency Virus Type 1 Reverse Transcriptase and Infection in Vitro. Proc. Natl. Acad. Sci. USA 1989, 86, 7191–7194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, W.E.G.; Weiler, B.E.; Charubala, R.; Pfleiderer, W.; Leserman, L.; Sobol, R.W.; Suhadolnik, R.J.; Schroeder, H.C. Cordycepin Analogs of 2′, 5′-Oligoadenylate Inhibit Human Immunodeficiency Virus Infection via Inhibition of Reverse Transcriptase. Biochemistry 1991, 30, 2027–2033. [Google Scholar] [CrossRef]
- Ryu, E.; Son, M.; Lee, M.; Lee, K.; Cho, J.Y.; Cho, S.; Lee, S.K.; Lee, Y.M.; Cho, H.; Sung, G.-H. Cordycepin Is a Novel Chemical Suppressor of Epstein-Barr Virus Replication. Oncoscience 2014, 1, 866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, A.K.; Aggarwal, R. Repurposing Potential of FDA Approved and Investigational Drugs for COVID-19 Targeting SARS-CoV-2 Spike and Main Protease and Validation by Machine Learning Algorithm. Chem. Biol. Drug Des. 2020, 97, 836–853. [Google Scholar] [CrossRef] [PubMed]
- Bibi, S.; Hasan, M.M.; Wang, Y.-B.; Papadakos, S.P.; Yu, H. Cordycepin as a Promising Inhibitor of SARS-CoV-2 RNA Dependent RNA Polymerase (RdRp). Curr. Med. Chem. 2021, 28, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yang, S.; Yang, X.; Chen, Z.; Li, J. Anticarcinogenic Effect and Hormonal Effect of Cordyceps militaris Link. Chin. Med. J. 1997, 22, 111–113. [Google Scholar]
- Hong, I.-P.; Choi, Y.-S.; Woo, S.-O.; Han, S.-M.; Kim, H.-K.; Lee, M.-R.; Nam, S.-H.; Ha, N.-G. Stimulatory Effect of Cordyceps militaris on Testosterone Production in Male Mouse. Kor. J. Mycol. 2011, 39, 148–150. [Google Scholar] [CrossRef] [Green Version]
- Leu, S.-F.; Poon, S.L.; Pao, H.-Y.; Huang, B.-M. The in Vivo and in Vitro Stimulatory Effects of Cordycepin on Mouse Leydig Cell Steroidogenesis. Biosci. Biotechnol 2011, 75, 723–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohn, S.-H.; Lee, S.-C.; Hwang, S.-Y.; Kim, S.-W.; Kim, I.-W.; Michael, B.Y.; Kim, S.-K. Effect of Long-Term Administration of Cordycepin from Cordyceps militaris on Testicular Function in Middle-Aged Rats. Planta Med. 2012, 78, 1620–1625. [Google Scholar] [CrossRef] [PubMed]
- Kopalli, S.R.; Cha, K.-M.; Lee, S.-H.; Hwang, S.-Y.; Lee, Y.-J.; Koppula, S.; Kim, S.-K. Cordycepin, an Active Constituent of Nutrient Powerhouse and Potential Medicinal Mushroom Cordyceps militaris Linn., Ameliorates Age-Related Testicular Dysfunction in Rats. Nutrients 2019, 11, 906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.-H.; Tsai, M.-T.; Chen, Y.-S.; Hou, R.C.-W.; Hung, H.-F.; Li, C.-H.; Wang, H.-K.; Lai, M.-N.; Jeng, K.-C.G. Improvement of Sperm Production in Subfertile Boars by Cordyceps militaris Supplement. Am. J. Chinese Med. 2007, 35, 631–641. [Google Scholar] [CrossRef] [Green Version]
- Kusama, K.; Miyagawa, M.; Ota, K.; Kuwabara, N.; Saeki, K.; Ohnishi, Y.; Kumaki, Y.; Aizawa, T.; Nakasone, T.; Okamatsu, S. Cordyceps militaris Fruit Body Extract Decreases Testosterone Catabolism and Testosterone-Stimulated Prostate Hypertrophy. Nutrients 2021, 13, 50. [Google Scholar] [CrossRef]
- Hsu, C.C.; Lin, Y.A.; Su, B.; Li, J.H.; Huang, H.Y.; Hsu, M.C. No Effect of Cordyceps sinensis Supplementation on Testosterone Level and Muscle Strength in Healthy Young Adults for Resistance Training. Biol. Sport 2011, 28, 107. [Google Scholar] [CrossRef] [Green Version]
- Yue, G.G.-L.; Bik-San Lau, C.; Fung, K.-P.; Leung, P.-C.; Ko, W.-H. Effects of Cordyceps sinensis, Cordyceps militaris and Their Isolated Compounds on Ion Transport in Calu-3 Human Airway Epithelial Cells. J. Ethnopharmacol. 2008, 117, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-H.; Sun, H.-L.; Sheu, J.-N.; Ku, M.-S.; Hu, C.-M.; Chan, Y.; Lue, K.-H. Effects of the Immunomodulatory Agent Cordyceps militaris on Airway Inflammation in a Mouse Asthma Model. Pediatr. Neonatol. 2008, 49, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Fei, X.; Zhang, X.; Zhang, G.; Bao, W.; Zhang, Y.; Zhang, M.; Zhou, X. Cordycepin Inhibits Airway Remodeling in a Rat Model of Chronic Asthma. Biomed. Pharmacother. 2017, 88, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, L.; Cai, T. Cordyceps Polysaccharide Ameliorates Airway Inflammation in an Ovalbumin-Induced Mouse Model of Asthma via TGF-Β1/Smad Signaling Pathway. Respir. Physiol. Neurobiol. 2020, 276, 103412. [Google Scholar] [CrossRef]
- Choi, K.-H.; Yoo, J.-E.; Hwang, G.-S.; Yoo, D.-Y. Effects of Cordyceps militaris (CM) on Osteoclastogenesis and Gene Expression. Obstet. Gynecol. Sci. 2012, 25, 16–26. [Google Scholar]
- Ying, X.; Peng, L.; Chen, H.; Shen, Y.; Yu, K.; Cheng, S. Cordycepin Prevented IL-β-Induced Expression of Inflammatory Mediators in Human Osteoarthritis Chondrocytes. Int. Ort. 2014, 38, 1519–1526. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wang, Z.; Qi, W.; Lei, W.; Zhao, G. Cordycepin (3′-Deoxyadenosine) down-Regulates the Proinflammatory Cytokines in Inflammation-Induced Osteoporosis Model. Inflammation 2014, 37, 1044–1049. [Google Scholar] [CrossRef]
- Kim, J.; Lee, H.; Kang, K.S.; Chun, K.-H.; Hwang, G.S. Cordyceps militaris Mushroom and Cordycepin Inhibit RANKL-Induced Osteoclast Differentiation. J. Med. Food 2015, 18, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.; Radhi, M.; Gowler, P.; Burston, J.J.; Gandhi, R.D.; Thorn, G.J.; Piccinini, A.M.; Walsh, D.A.; Chapman, V.; de Moor, C.H. The Polyadenylation Inhibitor Cordycepin Reduces Pain, Inflammation and Joint Pathology in Rodent Models of Osteoarthritis. Sci. Rep. 2019, 9, 4696. [Google Scholar] [CrossRef]
- He, M.T.; Lee, A.Y.; Kim, J.H.; Park, C.H.; Shin, Y.S.; Cho, E.J. Protective Role of Cordyceps militaris in Aβ 1–42-Induced Alzheimer’s Disease in Vivo. Food Sci. Biotechnol. 2019, 28, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Park, J.; Park, J.; Shin, H.-J.; Kwon, S.; Yeom, M.; Sur, B.; Kim, S.; Kim, M.; Lee, H. Cordyceps militaris Improves Neurite Outgrowth in Neuro2a Cells and Reverses Memory Impairment in Rats. Food Sci. Biotechnol. 2011, 20, 1599–1608. [Google Scholar] [CrossRef]
- Kim, Y.O.; Kim, H.J.; Abu-Taweel, G.M.; Oh, J.; Sung, G.-H. Neuroprotective and Therapeutic Effect of Cordyceps militaris on Ischemia-Induced Neuronal Death and Cognitive Impairments. Saudi J. Biol. Sci. 2019, 26, 1352–1357. [Google Scholar] [CrossRef]
- Yuan, G.; An, L.; Sun, Y.; Xu, G.; Du, P. Improvement of Learning and Memory Induced by Cordyceps Polypeptide Treatment and the Underlying Mechanism. ECAM 2018, 2018, 9419264. [Google Scholar] [CrossRef] [PubMed]
- Veena, R.K.; Carmel, E.J.; Ramya, H.; Ajith, T.A.; Wasser, S.P.; Janardhanan, K.K. Caterpillar Medicinal Mushroom, Cordyceps militaris (Ascomycetes), Mycelia Attenuates Doxorubicin-Induced Oxidative Stress and Upregulates Krebs Cycle Dehydrogenases Activity and ATP Level in Rat Brain. Int. J. Med. Mushrooms 2020, 22, 593–604. [Google Scholar] [CrossRef] [PubMed]
- U.S. National Library of Medicine. Clinical Trials. Available online: https://clinicaltrials.gov/ct2/show/NCT04002219/ (accessed on 15 August 2021).
- Won, K.-J.; Lee, S.-C.; Lee, C.-K.; Lee, H.M.; Lee, S.H.; Fang, Z.; Choi, O.B.; Jin, M.; Kim, J.; Park, T. Cordycepin Attenuates Neointimal Formation by Inhibiting Reactive Oxygen Species–Mediated Responses in Vascular Smooth Muscle Cells in Rats. J. Pharmacol. Sci. 2009, 109, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Lian, Z.-Q.; Zhu, P.; Zhu, H.-B. Lipid-Lowering Effect of Cordycepin (3′-Deoxyadenosine) from Cordyceps militaris on Hyperlipidemic Hamsters and Rats. Acta Pharm. Sin. 2011, 46, 669–676. [Google Scholar]
- Guo, P.; Kai, Q.; Gao, J.; Lian, Z.; Wu, C.; Wu, C.; Zhu, H. Cordycepin Prevents Hyperlipidemia in Hamsters Fed a High-Fat Diet via Activation of AMP-Activated Protein Kinase. J. Pharmacol. Sci. 2010, 113, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, S.; Tamai, M.; Nakajima, S.; Kato, H.; Johno, H.; Nakamura, T.; Kitamura, M. Blockade of Adipocyte Differentiation by Cordycepin. Br. J. Pharmacol. 2012, 167, 561–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Guo, Y.; Su, Y.; Zhang, X.; Luan, H.; Zhang, X.; Zhu, H.; He, H.; Wang, X.; Sun, G. Cordycepin Activates AMP-activated Protein Kinase (AMPK) via Interaction with the Γ1 Subunit. J. Cell. Mol. Med. 2014, 18, 293–304. [Google Scholar] [CrossRef]
- Hu, S.; Wang, J.; Li, F.; Hou, P.; Yin, J.; Yang, Z.; Yang, X.; Li, T.; Xia, B.; Zhou, G. Structural Characterisation and Cholesterol Efflux Improving Capacity of the Novel Polysaccharides from Cordyceps militaris. Int. J. Biol. Macromol. 2019, 131, 264–272. [Google Scholar] [CrossRef]
- Wang, L.; Xu, N.; Zhang, J.; Zhao, H.; Lin, L.; Jia, S.; Jia, L. Antihyperlipidemic and Hepatoprotective Activities of Residue Polysaccharide from Cordyceps militaris SU-12. Carbohydr. Polym. 2015, 131, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhang, M.; Zhang, S.; Wang, Y.; Jiang, X. Structural Characterization of Polysaccharides from Cordyceps militaris and Their Hypolipidemic Effects in High Fat Diet Fed Mice. RSC Adv. 2018, 8, 41012–41022. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.-J.; Cho, J.Y.; Rhee, M.H.; Park, H.-J. Cordycepin (3′-Deoxyadenosine) Inhibits Human Platelet Aggregation in a Cyclic AMP-and Cyclic GMP-Dependent Manner. Eur. J. Pharmacol. 2007, 558, 43–51. [Google Scholar] [CrossRef]
- Park, E.-S.; Kang, D.-H.; Yang, M.-K.; Kang, J.C.; Jang, Y.C.; Park, J.S.; Kim, S.-K.; Shin, H.-S. Cordycepin, 3′-Deoxyadenosine, Prevents Rat Hearts from Ischemia/Reperfusion Injury via Activation of Akt/GSK-3β/P70S6K Signaling Pathway and HO-1 Expression. Cardiovasc. Toxicol. 2014, 14, 1–9. [Google Scholar] [CrossRef]
- Lee, D.-H.; Kim, H.-H.; Lim, D.H.; Kim, J.-L.; Park, H.-J. Effect of Cordycepin-Enriched WIB801C from Cordyceps militaris Suppressing Fibrinogen Binding to Glycoprotein IIb/IIIa. Biomol. Ther. 2015, 23, 60. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Cho, G.-S.; Ryu, S.; Kim, H.J.; Song, H.Y.; Yune, T.Y.; Ju, C.; Kim, W.-K. Post-Ischemic Treatment of WIB801C, Standardized Cordyceps Extract, Reduces Cerebral Ischemic Injury via Inhibition of Inflammatory Cell Migration. J. Ethnopharmacol. 2016, 186, 169–180. [Google Scholar] [CrossRef]
- Choi, E.; Oh, J.; Sung, G.-H. Antithrombotic and Antiplatelet Effects of Cordyceps militaris. Mycobiology 2020, 48, 228–232. [Google Scholar] [CrossRef]
- Ji, Y.; Su, A.; Ma, G.; Tao, T.; Fang, D.; Zhao, L.; Hu, Q. Comparison of Bioactive Constituents and Effects on Gut Microbiota by in Vitro Fermentation between Ophicordyceps sinensis and Cordyceps militaris. J. Funct. Foods 2020, 68, 103901. [Google Scholar] [CrossRef]
- Heo, J.Y.; Baik, H.W.; Kim, H.J.; Lee, J.M.; Kim, H.W.; Choi, Y.S.; Won, J.H.; Hyun Mi Kim, W.I.P.; Kim, C.Y. The Efficacy and Safety of Cordyceps militaris in Korean Adults Who Have Mild Liver Dysfunction. J. Clin. Nutr. 2015, 7, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Zhao-Long, W.; Xiao-Xia, W.; Wei-Ying, C. Inhibitory Effect of Cordyceps sinensis and Cordyceps militaris on Human Glomerular Mesangial Cell Proliferation Induced by Native LDL. Cell Biochem. Funct. 2000, 18, 93–97. [Google Scholar] [CrossRef]
- Sun, T.; Dong, W.; Jiang, G.; Yang, J.; Liu, J.; Zhao, L.; Ma, P. Cordyceps militaris Improves Chronic Kidney Disease by Affecting TLR4/NF-ΚB Redox Signaling Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 7850863. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.-H.; Dubey, N.K.; Li, W.-S.; Liu, M.-C.; Chiang, H.-S.; Leu, S.-J.; Shieh, Y.-H.; Tsai, F.-C.; Deng, W.-P. Cordyceps militaris Treatment Preserves Renal Function in Type 2 Diabetic Nephropathy Mice. PLoS ONE 2016, 11, e0166342. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, G.C.; Wasti, S.S.; Hendrickson, D.L. Murine Safety of Two Species of Entomogenous Fungi, Cordyceps militaris (FRIES) LINK and Paecilomyces Fumoso-Roseus (WIZE) BROWN and SMITH. Appl. Entomol. Zool. 1979, 14, 217–220. [Google Scholar] [CrossRef]
- Jhou, B.-Y.; Fang, W.-C.; Chen, Y.-L.; Chen, C.-C. A 90-Day Subchronic Toxicity Study of Submerged Mycelial Culture of Cordyceps militaris in Rats. Toxicol. Res. 2018, 7, 977–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, K.-C.; Sheu, F. A Novel Protein from Edible Fungi Cordyceps militaris That Induces Apoptosis. J. Food Drug Anal. 2018, 26, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gençcelep, H.; Uzun, Y.; Tunçtürk, Y.; Demirel, K. Determination of Mineral Contents of Wild-Grown Edible Mushrooms. Food Chem. 2009, 113, 1033–1036. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content | |||
|---|---|---|---|
| Bioactive Compound | Mycelium | Fruiting Bodies | References |
| Cordycepin | 1.82 mg/g | 1.10 mg/g | [9] |
| Cordycepin | 1.74 mg/g | 5.28 mg/g (Water extrct) | [7] |
| Cordycepin | 8.37 mg/g (Ethanol extract) | [7] | |
| D-mannitol | 5.2 mg/kg | 4.7 mg/kg | [9] |
| Ergothioneine | 130.6 mg/kg | 782.3 mg/kg | [9] |
| Ergothioneine | 123.4–785.1 mg/kg | 409.8 μg/g | [7,8] |
| GABA | 68.6–180.1 mg/kg | 756.30 μg/g | [7,8] |
| Lovastatin | 37.7–57.3 mg/kg | 2.76 μg/g | [7,8] |
| Vitamins | |||
| Vit. A | 100 mg/kg | 96 mg/kg | [9] |
| Vit. E (tocopherols) | 1.3 mg/kg | 3.6 mg/kg | [9] |
| Vit. B2 (riboflavin) | 0.32 mg/kg | 0.16 mg/kg | [9] |
| Vit. B3 (niacin) | 15.2 mg/kg | 4.9 mg/kg | [9] |
| Vit. C | Not detected | <2 mg/kg | [9] |
| Bioelements | |||
| Magnesium | 3414 mg/kg | 4227 mg/kg | [9] |
| Sulfur | 2558 mg/kg | 5088 mg/kg | [9] |
| Potassium | 12,183 mg/kg | 15,938 mg/kg | [9] |
| Selenium | <0.5 mg/kg | 0.4 mg/kg | [9] |
| Iron | 9 mg/kg | 31 mg/kg | [9] |
| Calcium | 11 mg/kg | 797 mg/kg | [9] |
| Zinc | 10 mg/kg | Not detected | [9] |
| Nutrients | |||
| Protein | 39.5% | 59.8% | [9] |
| Protein | Not analyzed | 29.7% | [7] |
| Fat | 2.2% | 8.8% | [9] |
| Fat | Not analyzed | 2.9% | [7] |
| Carbohydrate | 39.6% | 29.1% | [9] |
| Carbohydrate | Not analyzed | 54.3% | [7] |
| Research Group | Quantity of Raw Material C. militaris | Supplementation Time | Biological Activity | Bioactive Compound | Adverse Events | References |
|---|---|---|---|---|---|---|
| 10 healthy individuals of both sexes aged 19–24 years | 4 g/day 1 | 3 weeks | Ergogenic | Undefined | Undefined | [70] |
| 28 healthy individuals of both sexes aged 18–27 years | 4 g/day 1 | 1–3 weeks | Ergogenic | Undefined | Undefined | [71] |
| 43 healthy individuals of both sexes aged 19–34 years | 1–12 g/day 1 | 1–4 weeks | Ergogenic | Undefined | Gastrointestinal | [72] |
| 79 healthy males aged 19–64 years | 1.5 g/day | 4 weeks | Immunostimulation | Cordycepin | No serious adverse events | [80] |
| 100 healthy volunteers aged 20–70 years | 1.5 g/day | 12 weeks | Immunostimulation | Cordycepin | No serious adverse events | [81] |
| 510 patients with chronic bronchitis | Undefined 2 | Undefined | Anti-inflammatory | Undefined | Undefined | [107] |
| 425 patients with chronic bronchitis | 3 g/day 2 | 8 weeks | Anti-inflammatory | Undefined | Undefined | [108] |
| 57 patients with mild hepatic impairment aged 31–61 years | 1.5 g/day | 8 weeks | Hepatoprotective | Cordycepin | No serious adverse events | [155] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jędrejko, K.J.; Lazur, J.; Muszyńska, B. Cordyceps militaris: An Overview of Its Chemical Constituents in Relation to Biological Activity. Foods 2021, 10, 2634. https://doi.org/10.3390/foods10112634
Jędrejko KJ, Lazur J, Muszyńska B. Cordyceps militaris: An Overview of Its Chemical Constituents in Relation to Biological Activity. Foods. 2021; 10(11):2634. https://doi.org/10.3390/foods10112634
Chicago/Turabian StyleJędrejko, Karol Jerzy, Jan Lazur, and Bożena Muszyńska. 2021. "Cordyceps militaris: An Overview of Its Chemical Constituents in Relation to Biological Activity" Foods 10, no. 11: 2634. https://doi.org/10.3390/foods10112634