
Changes of Metabolites and Gene Expression under Different Feeding Systems Associated with Lipid Metabolism in Lamb Meat


 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
2.2. Analysis of Short-Chain Fatty Acid Composition
2.3. Analysis of Muscle Fatty Acid Composition
2.4. Metabolomics Analysis
2.5. Transcriptomics Analysis
2.6. Data Analysis
3. Results and Discussion
3.1. Fatty Acid Analysis
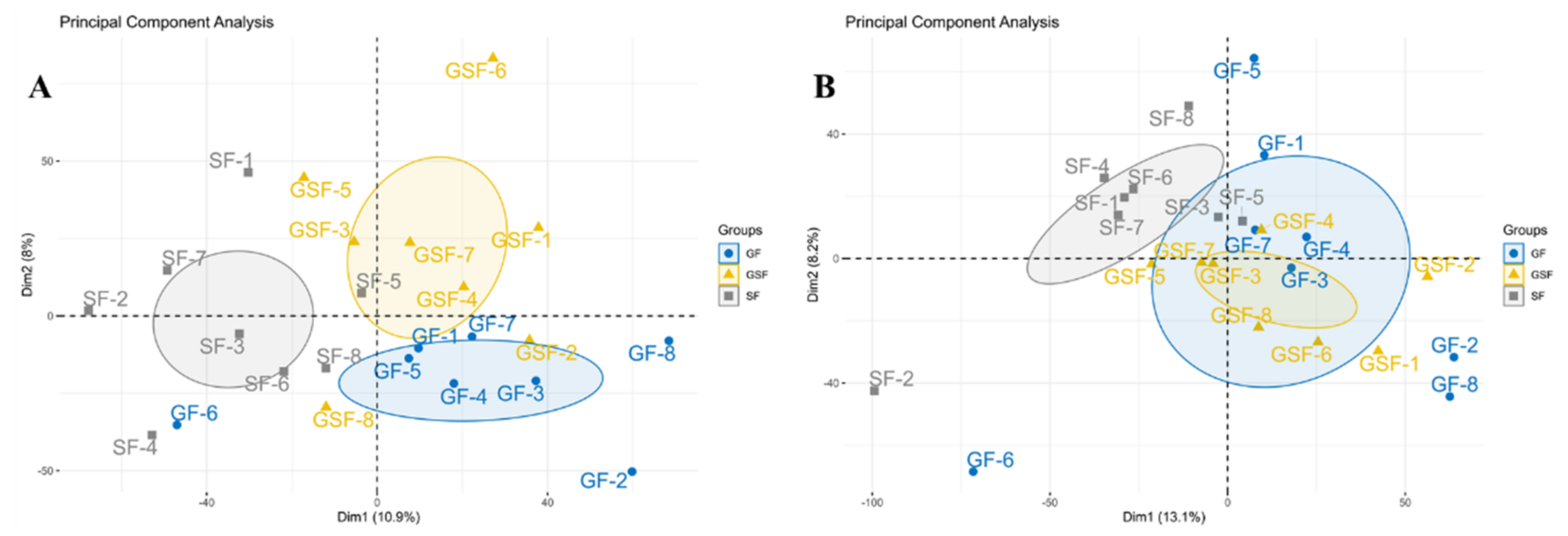
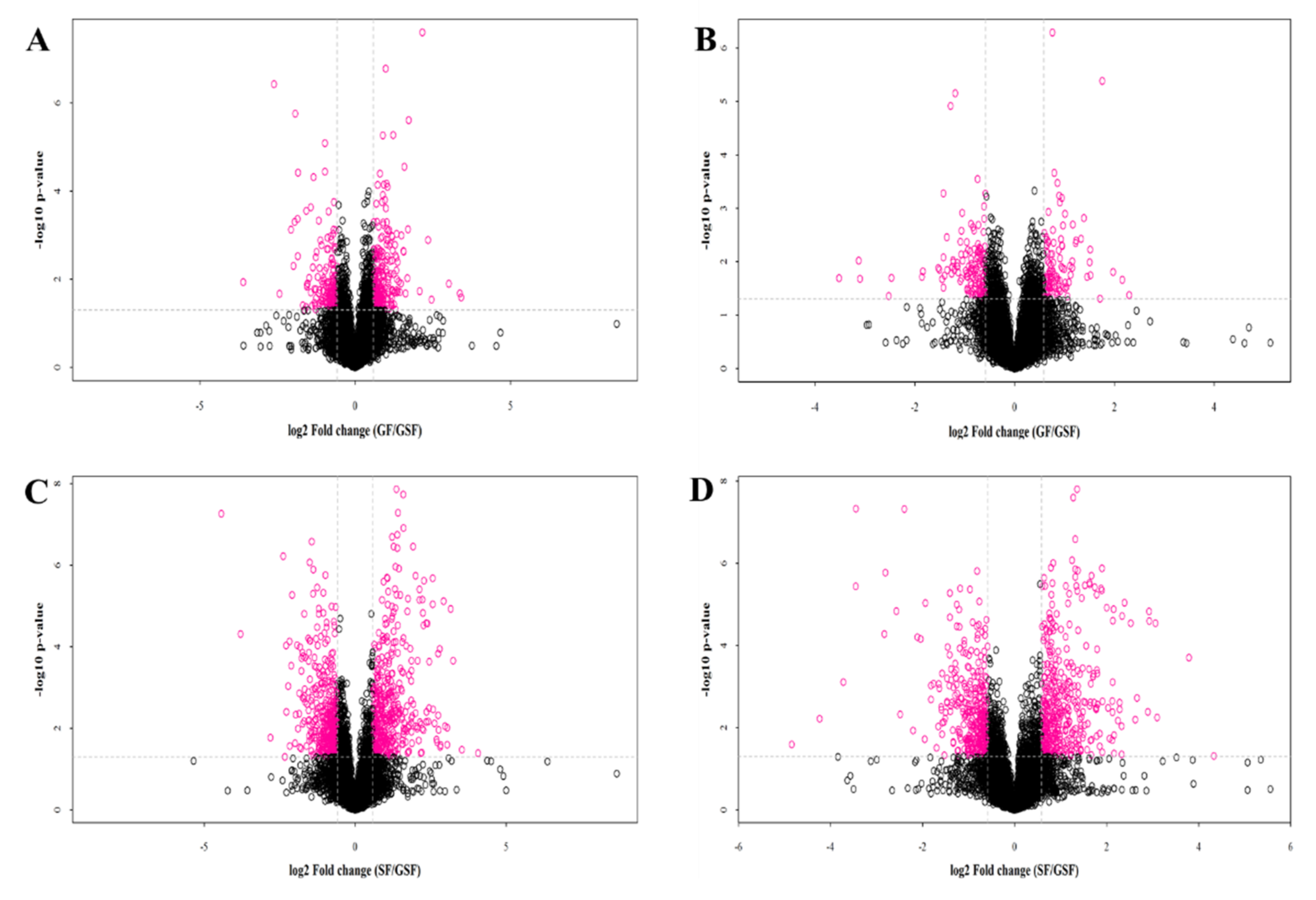
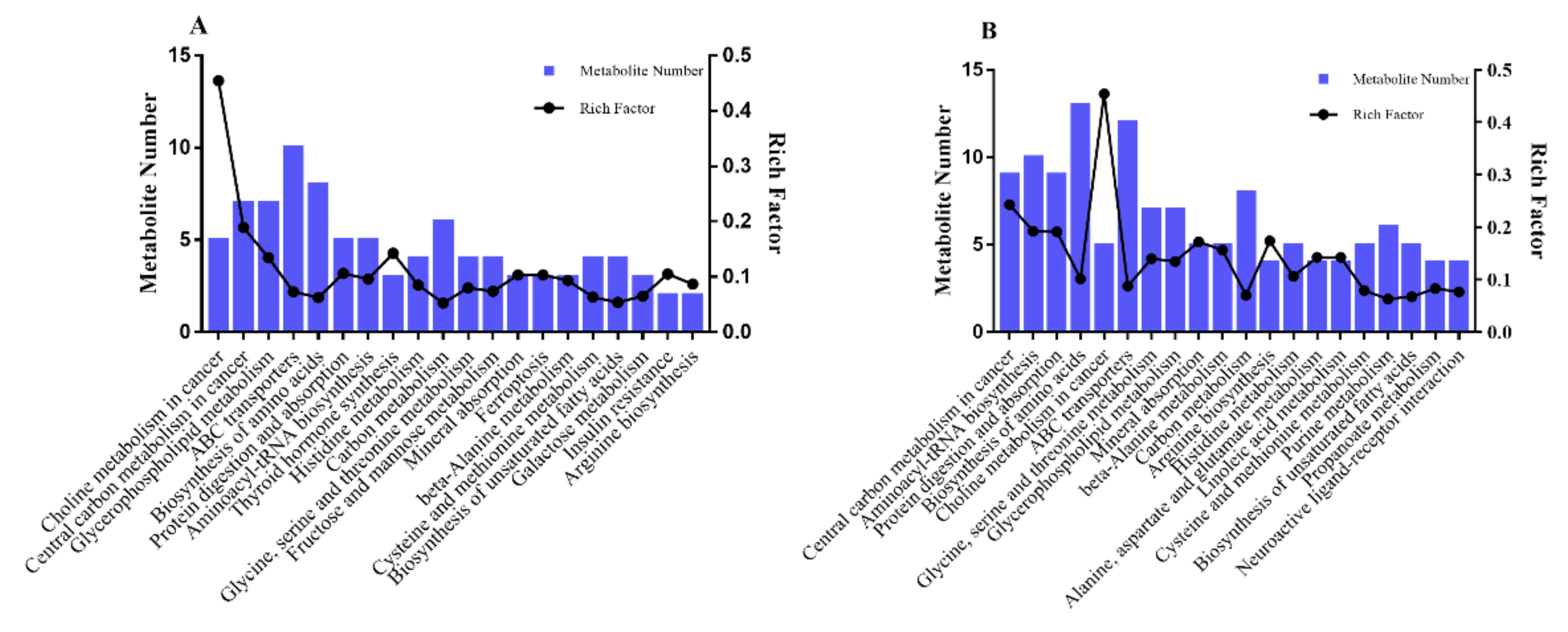
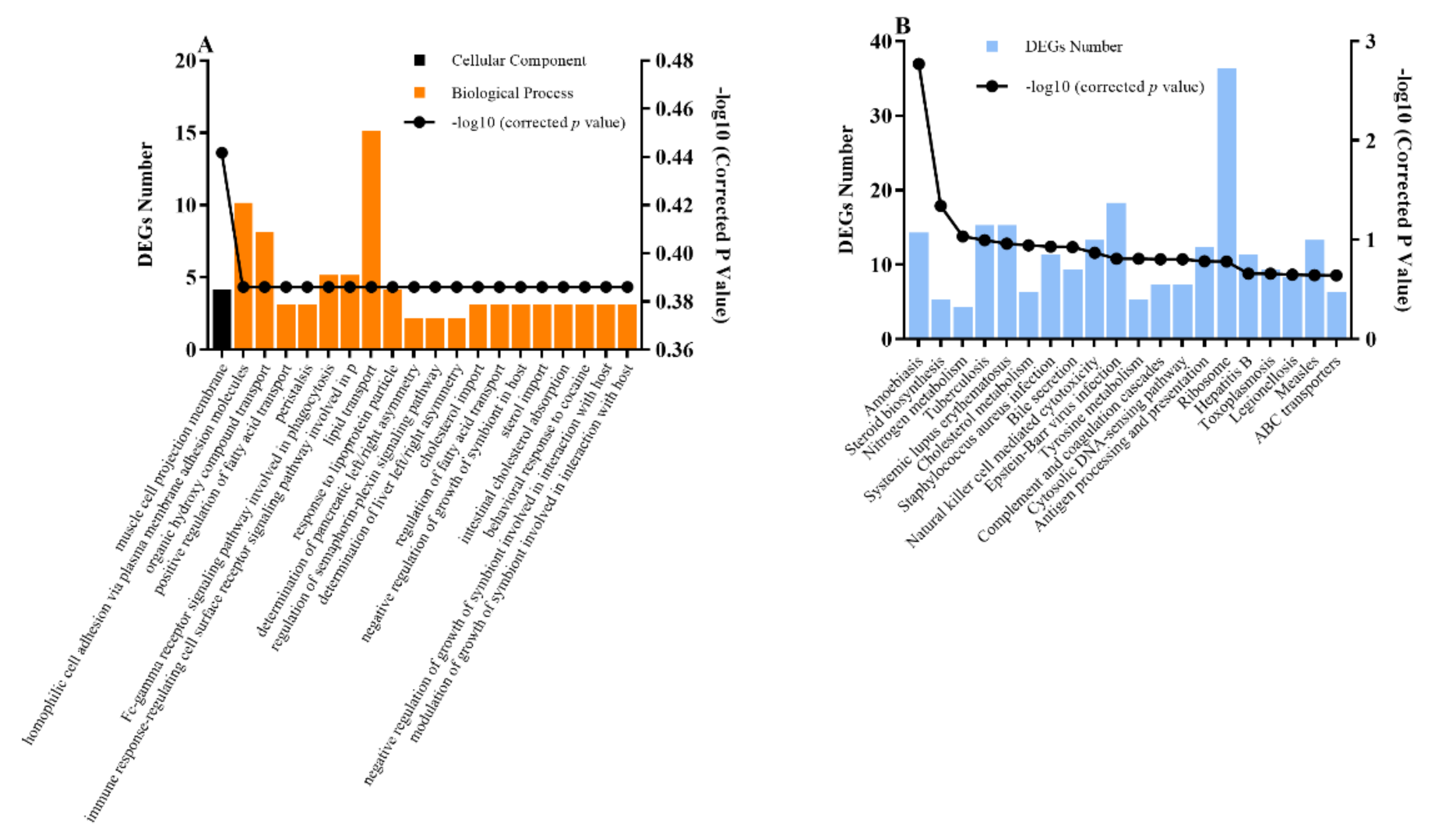
3.2. Bioinformatic Analysis of Metabolites
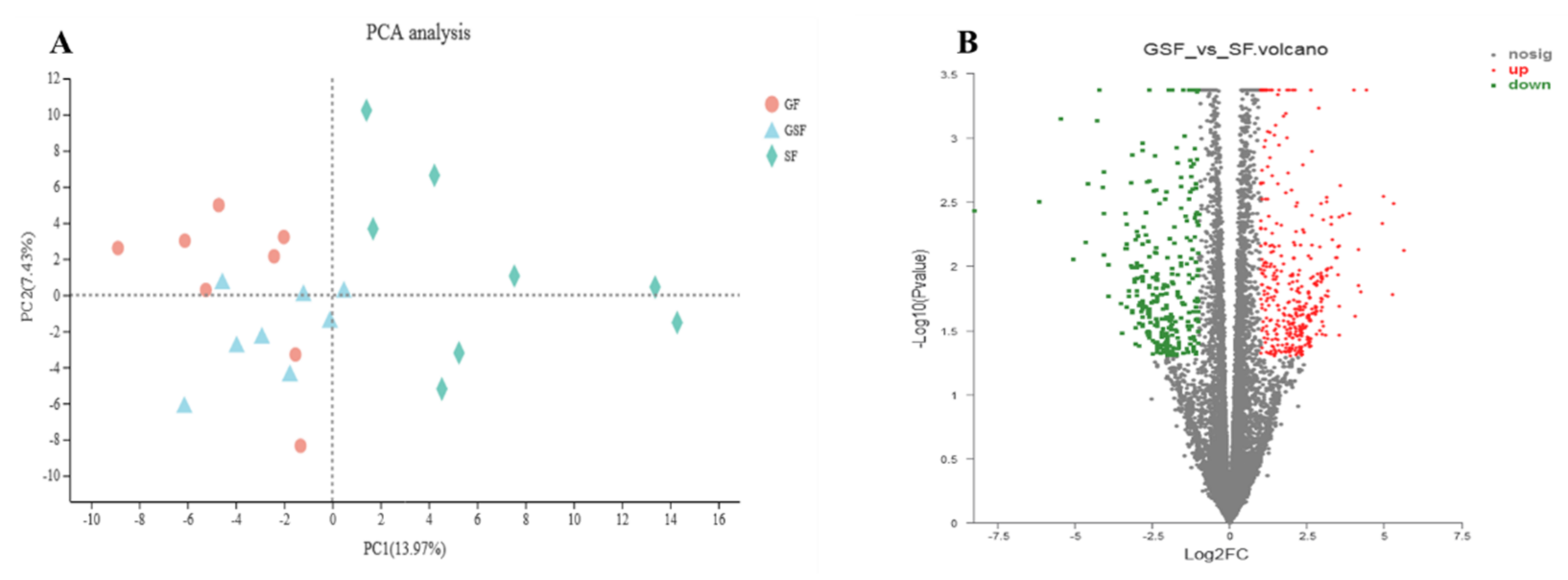
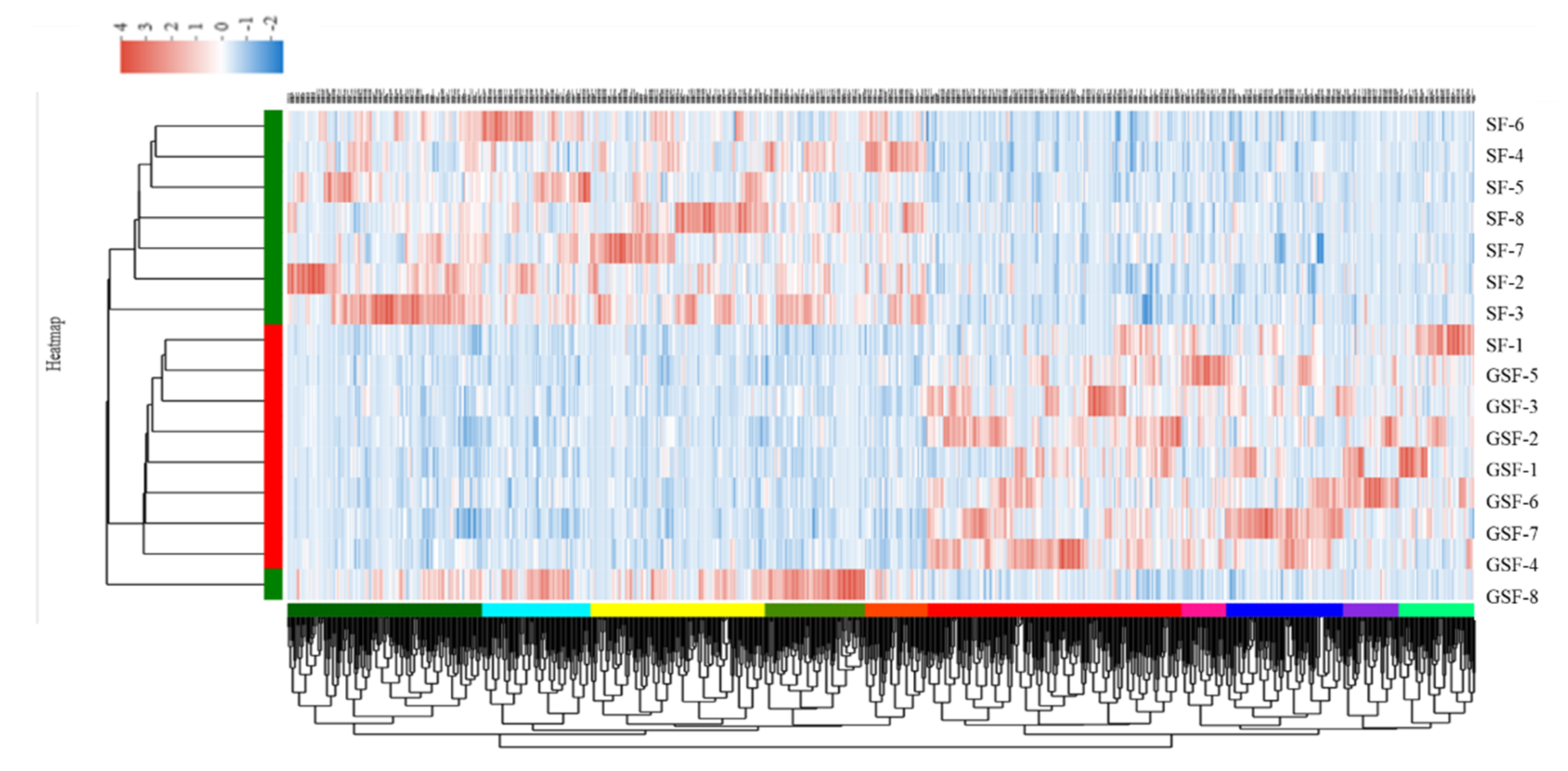
3.3. Bioinformatic Analysis of Transcriptome
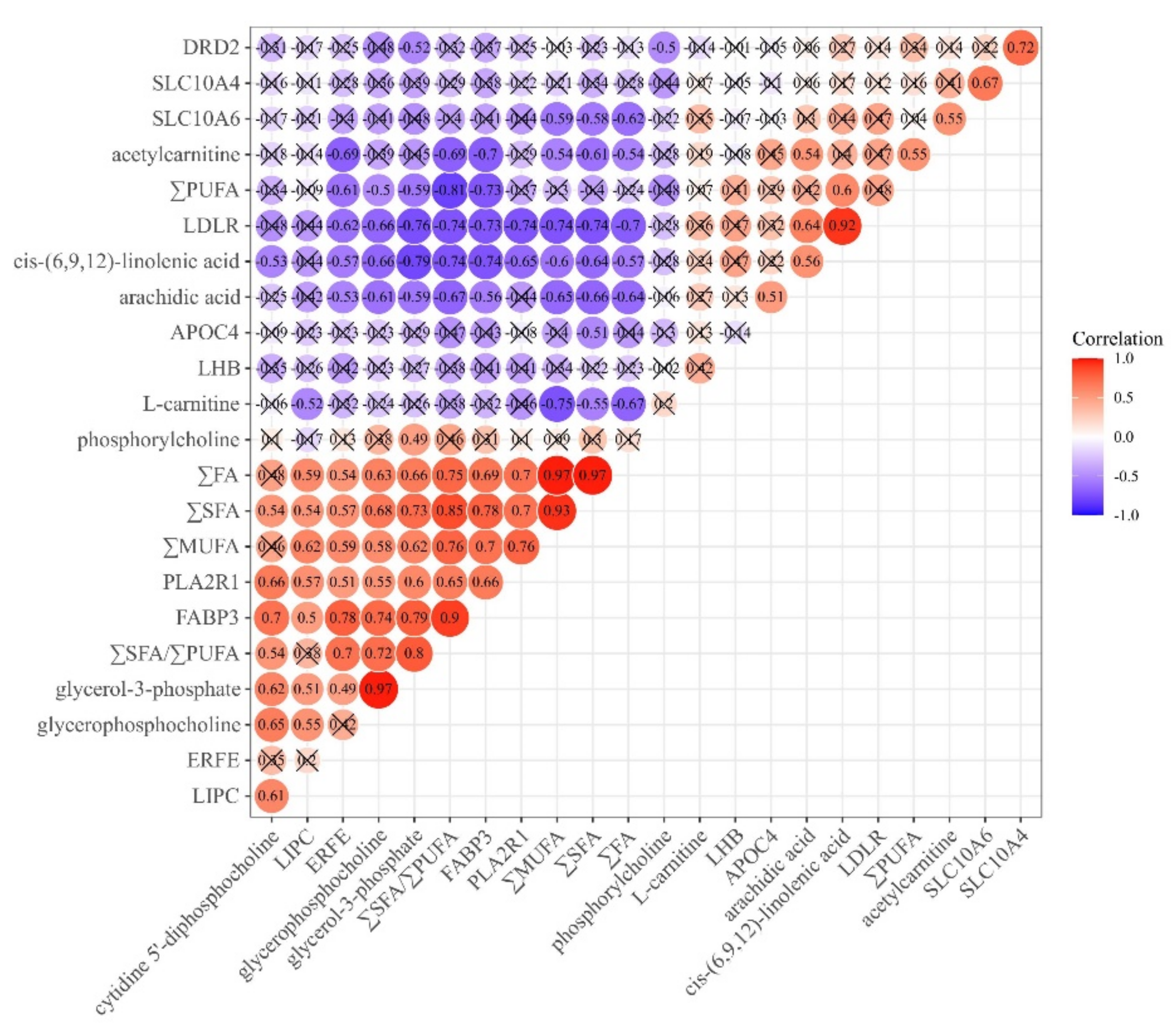
3.4. Correlation Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scollan, N.; Hocquette, J.-F.; Nuernberg, K.; Dannenberger, D.; Richardson, I.; Moloney, A. Innovations in beef production systems that enhance the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 2006, 74, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Knapik, J.; Ropka-Molik, K.; Pieszka, M. Genetic and Nutritional Factors Determining the Production and Quality of Sheep Meat A Review. Ann. Anim. Sci. 2017, 17, 23–40. [Google Scholar] [CrossRef] [Green Version]
- Resconi, V.C.; Bueno, M.; Escudero, A.; Magalhaes, D.; Ferreira, V.; Campo, M.M. Ageing and retail display time in raw beef odour according to the degree of lipid oxidation. Food Chem. 2018, 242, 288–300. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.; Jo, C.; Tariq, M.R. Meat flavor precursors and factors influencing flavor precursors—A systematic review. Meat Sci. 2015, 110, 278–284. [Google Scholar] [CrossRef]
- Li, J.; Tang, C.; Zhao, Q.; Yang, Y.; Li, F.; Qin, Y.; Liu, X.; Yue, X.; Zhang, J. Integrated lipidomics and targeted metabolomics analyses reveal changes in flavor precursors in psoas major muscle of castrated lambs. Food Chem. 2020, 333, 127451. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Wang, X.-C. Identification of important odorants derived from phosphatidylethanolamine species in steamed male Eriocheir sinensis hepatopancreas in model systems. Food Chem. 2019, 286, 491–499. [Google Scholar] [CrossRef]
- Bravo-Lamas, L.; Barron, L.J.; Farmer, L.; Aldai, N. Fatty acid composition of intramuscular fat and odour-active compounds of lamb commercialized in northern Spain. Meat Sci. 2018, 139, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Daszkiewicz, T.; Bak, T.; Denaburski, J. Quality of pork with a different intramuscular fat (IMF) content. Pol. J. Food Nutr. Sci. 2005, 14/55, 31–35. [Google Scholar]
- Beriain, M.J.; Bas, P.; Purroy, A.; Treacher, T. Effect of animal and nutritional factors and nutrition on lamb meat quality. Cah. Options Mediterran. 2000, 52, 75–86. [Google Scholar]
- Chaosap, C.; Chauychuwong, N.; Chauychuwong, R.; Sriprem, C.; Sivapirunthep, P.; Sazili, A. Carcass Composition, Meat Quality, Calpain Activity, Fatty Acid Composition and Ribonucleotide Content in Southern Thai Native Goats and Three-Way Crossbred Goats. Foods 2021, 10, 1323. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, H.; Liu, M.; Zhao, X.; Luo, H.L. Effect of breed on the volatile compound precursors and odor profile at-tributes of lamb meat. Foods 2020, 9, 1178. [Google Scholar] [CrossRef] [PubMed]
- Casey, N.; Webb, E. Managing goat production for meat quality. Small Rumin. Res. 2010, 89, 218–224. [Google Scholar] [CrossRef] [Green Version]
- Enser, M.; Hallett, K.G.; Hewitt, B.; Fursey, G.A.; Wood, J.D.; Harrington, G. Fatty acid content and composition of SUP1 beef and lamb muscle in relation to production system and implications for human nutrition. Meat Sci. 1998, 49, 329–341. [Google Scholar] [CrossRef]
- Nuernberg, K.; Dannenberg, D.; Nuernberg, G.; Ender, K.; Voigt, J.; Scollan, N.D.; Wood, J.D.; Nute, G.R.; Richardson, R.I. Effect of a grass-based and a concentrate feeding system on meat quality characteristics and fatty acid composition of longis-simus muscle in different cattle breeds. Meat Sci. 2005, 94, 137–147. [Google Scholar]
- Luo, Z.; Shen, L.; Jiang, J.; Huang, Y.; Bai, L.; Yu, S.; Yao, X.; Ren, Z.; Yang, Y.; Cao, S. Plasma metabolite changes in dairy cows during parturition identified using untargeted metabolomics. J. Dairy Sci. 2019, 102, 4639–4650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jawor, P.; Ząbek, A.; Wojtowicz, W.; Król, D.; Stefaniak, T.; Młynarz, P. Metabolomic studies as a tool for determining the post-mortem interval (PMI) in stillborn calves. BMC Vet. Res. 2019, 15, 189. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.G.; Guo, Y.P.; Liu, M.; Zhang, C.; Luo, H.L. Effect of desert steppe grazing on slaughtering performance and meat quality of Tan sheep. Pratacultural Sci. 2021, 38, 554–561. [Google Scholar]
- Leary, S.; Underwood, W.; Anthony, R.; Cartner, S.; Corey, D.; Grandin, T.; Greenacre, C.; McCrackin, M.A.; Meyer, R.; Miller, D. AVMA Guidelines for The Euthanasia of Animals: 2013 Edition; American Veterinary Medical Association: Schaumburg, IL, USA, 2013. [Google Scholar]
- Park, B.S.; Park, S.O. Effects of feeding time with betaine diet on growth performance, blood markers, and short chain fatty acids in meat ducks exposed to heat stress. Livest. Sci. 2017, 199, 31–36. [Google Scholar] [CrossRef]
- Hanson, S.W.F.; Olley, J. Application of the Bligh and Dyer method of lipid extraction to tissue homogenates. Biochemi. J. 1963, 89, 101–102. [Google Scholar]
- Morrison, W.R.; Smith, L.M. Preparation of fatty acid methyl esters and dimethyl acetals from lipids with boron fluoride-methanol. J. Lipid Res. 1964, 5, 600–608. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Y.; Zuo, S.; Peng, S.; Wang, Z.; Zhang, Y.; Luo, H. Untargeted and Targeted Metabolomics Profiling of Muscle Reveals Enhanced Meat Quality in Artificial Pasture Grazing Tan Lambs via Rescheduling the Rumen Bacterial Community. J. Agric. Food Chem. 2021, 69, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.; Rinn, J.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential expression of RNA-Seq data at the gene level-the DESeq package. Germany. Eur. Molecular Biol. Lab. (EMBL) 2012, 10, f1000research. [Google Scholar]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Watkins, P.J.; Frank, D.; Singh, T.K.; Young, O.A.; Warner, R.D. Sheep meat flavor and the effect of different feeding systems: A Review. J. Agric. Food Chem. 2013, 61, 3561–3579. [Google Scholar] [CrossRef] [PubMed]
- Almela, E.; Jordán, M.J.; Martínez, C.; Sotomayor, J.A.; Bedia, M.; Bañón, S. Ewe’s Diet (Pasture vs Grain-Based Feed) Affects Volatile Profile of Cooked Meat from Light Lamb. J. Agric. Food Chem. 2010, 58, 9641–9646. [Google Scholar] [CrossRef] [PubMed]
- Larráyoz, P.; Mendía, C.; Torre, P.; Barcina, Y.; Ordóñez, A.I. Sensory profile of flavor and odor characteristics in roncal cheese made from raw ewe’s milk. J. Sens. Stud. 2010, 17, 415–427. [Google Scholar] [CrossRef]
- Najwa, M.H.; Man, Y.C.; Shuhaimi, M.; Abdul, M.Y. Tentative identification of volatile flavor compounds in commercial Budu, a Malaysian fish sauce, using GC-MS. Molecules 2012, 17, 5062–5080. [Google Scholar]
- Scaife, J.R.; Wahle, K.W.; Garton, G.A. Utilization of methylmalonate for the synthesis of branched-chain fatty acids by prep-arations of chicken liver and sheep adipose tissue. Biochem. J. 1978, 176, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.M. Dietary fatty acids and human health. Anim. Res. 2000, 49, 165–180. [Google Scholar] [CrossRef] [Green Version]
- Velasco, S.; Caneque, V.; Lauzurica, S.; Perez, C.; Huidobro, F. Effect of different feeds on meat quality and fatty acid compo-sition of lambs fattened at pasture. Meat Sci. 2004, 66, 457–465. [Google Scholar] [CrossRef]
- Aurousseau, B.; Bauchart, D.; Calichon, E.; Micol, D.; Priolo, A. Effect of grass or concentrate feeding systems and rate of growth on triglyceride and phospholipid and their fatty acids in the M. longissimus thoracis of lambs. Meat Sci. 2004, 66, 531–541. [Google Scholar] [CrossRef]
- Cividini, A.; Levart, A.; Gur, S. Fatty acid composition of lamb meat as affected by production system, weaning and sex. Acta Agric. Slov. 2008, 2, 47–58. [Google Scholar]
- Wang, Z.; Chen, Y.; Luo, H.; Liu, X.; Liu, K. Influence of Restricted Grazing Time Systems on Productive Performance and Fatty Acid Composition of Longissimus dorsi in Growing Lambs. Asian-Australas. J. Anim. Sci. 2015, 28, 1105–1115. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.B.; Gill, C.; Lunt, D.K.; Brooks, M.A. Regulation of Fat and Fatty Acid Composition in Beef Cattle. Asian-Australas. J. Anim. Sci. 2009, 22, 1225–1233. [Google Scholar] [CrossRef]
- Boughalmi, A.; Araba, A. Effect of feeding management from grass to concentrate feed on growth, carcass characteristics, meat quality and fatty acid profile of Timahdite lamb breed. Small Rumin. Res. 2016, 144, 158–163. [Google Scholar] [CrossRef]
- Elmore, J.S.; Campo, M.M.; Enser, M.; Mottram, D.S. Effect of Lipid Composition on Meat-like Model Systems Containing Cysteine, Ribose, and Polyunsaturated Fatty Acids. J. Agric. Food Chem. 2002, 50, 1126–1132. [Google Scholar] [CrossRef]
- Priolo, A.; Micol, D.; Agabriel, J.; Prache, S.; Dransfield, E. Effect of grass or concentrate feeding systems on lamb carcass and meat quality. Meat Sci. 2001, 62, 179–185. [Google Scholar] [CrossRef]
- Dervishi, E.; Serrano, C.; Joy, M.; Serrano, M.; Rodellar, C.; Calvo, J.H. Effect of the feeding system on the fatty acid composi-tion, expression of the Δ9-desaturase, Peroxisome Proliferator-Activated Receptor Alpha, Gamma, and Sterol Regulatory El-ement Binding Protein 1 genes in the semitendinous muscle of light lambs of the Rasa Agragonesa breed. BMC Vet. Res. 2010, 6, 40. [Google Scholar]
- Saylor, P.J.; Karoly, E.D.; Smith, M.R. Prospective Study of Changes in the Metabolomic Profiles of Men during Their First Three Months of Androgen Deprivation Therapy for Prostate Cancer. Clin. Cancer Res. 2012, 18, 3677–3685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralph, F.; Wulf, R.; Muntau, A.C.; Torsten, D.; Jens, K.; Ralf, A.; Andrea, S.F.; Heinz, T.; Bernhard, L.; Bernhard, O. Hepatic carnitine palmitoyltransferase I deficiency: Acylcarnitine profiles in blood spots are highly specific. Clin. Chem. 2001, 47, 1763–1768. [Google Scholar]
- Sder, J.; Wernersson, S.; Dicksved, J.; Hagman, R.; Hglund, K. Indication of metabolic inflexibility to food intake in sponta-neously overweight Labrador retriever dogs. BMC Vet. Res. 2019, 15, 96. [Google Scholar]
- Faris, R.; Weber, M.M.; Seeger, D.R.; Cavazos, D.; Graffenried, L.D.; Murphy, E.J.; Jolly, C.A. Mitochondrial glycer-ol-3-phosphate acyltransferase-dependent phospholipid synthesis modulates phospholipid mass and il-2 production in jurkat T cells. Lipids 2016, 51, 1–11. [Google Scholar] [CrossRef]
- Khatun, I.; Clark, R.W.; Vera, N.B.; Kou, K.; Erion, D.M.; Coskran, T.; Bobrowski, W.F.; Okerberg, C.; Goodwin, B. Charac-terization of a novel intestinal glycerol-3-phosphate acyltransferase pathway and its role in lipid homeostasis. J. Biol. Chem. 2015, 291, 2602–2615. [Google Scholar] [CrossRef] [Green Version]
- Zufferey, R.; Mamoun, C.B. The initial step of glycerolipid metabolism in Leishmania major promastigotes involves a single glycerol-3-phosphate acyltransferase enzyme important for the synthesis of triacylglycerol but not essential for virulence. Mol. Microbiol. 2005, 56, 800–810. [Google Scholar] [CrossRef]
- Cui, Z.; Houweling, M. Phosphatidylcholine and cell death. Biochim. et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2002, 1585, 87–96. [Google Scholar] [CrossRef]
- da Silva, R.P.; Kelly, K.B.; Lewis, E.D.; Leonard, K.-A.; Goruk, S.; Curtis, J.M.; Vine, D.F.; Proctor, S.; Field, C.; Jacobs, R.L. Choline deficiency impairs intestinal lipid metabolism in the lactating rat. J. Nutr. Biochem. 2015, 26, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Michel, V.; Singh, R.K.; Bakovic, M. The impact of choline availability on muscle lipid metabolism. Food Funct. 2010, 2, 53–62. [Google Scholar] [CrossRef]
- Yan, L.; Han, P.; Man, J.; Tian, Y.; Wang, F.; Wang, J. Discovery of lipid profiles of type 2 diabetes associated with hyper-lipidemia using untargeted UPLC Q-TOF/MS-based lipidomics approach. Clin. Chim. Acta 2021, 520, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Cassar-Malek, I.; Jurie, C.; Bernard, C.; Barnola, I.; Micol, D.; Hocquette, J.-F. Pasture-feeding of Charolais steers influences skeletal muscle metabolism and gene expression. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2009, 60, 83–90. [Google Scholar]
- Gallardo, M.A.; Cárcamo, J.G.; Hiller, B.; Nuernberg, G.; Nuernberg, K.; Dannenberger, D. Expression of lipid metabolism related genes in subcutaneous adipose tissue from Chilota lambs grazing on two different pasture types. Eur. J. Lipid Sci. Technol. 2014, 117, 23–30. [Google Scholar] [CrossRef]
- Selim, S.; Elo, K.; Jaakkola, S.; Karikoski, N.; Boston, R.; Reilas, T.; Särkijärvi, S.; Saastamoinen, M.; Kokkonen, T. Relationships among body condition, insulin resistance and subcutaneous adipose tissue gene expression during the grazing season in mares. PLoS ONE 2015, 10, e0125968. [Google Scholar] [CrossRef] [Green Version]
- Berton, M.P.; Fonseca, L.F.S.; Gimenez, D.F.J.; Utembergue, B.L.; Cesar, A.S.M.; Coutinho, L.L.; de Lemos, M.V.A.; Aboujaoude, C.; Pereira, A.S.C.; de O Silva, R.M.; et al. Gene expression profile of intramuscular muscle in Nellore cattle with extreme values of fatty acid. BMC Genom. 2016, 17, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Quach, N.D.; Mock, J.N.; Scholpa, N.E.; Eggert, M.W.; Payré, C.; Lambeau, G.; Arnold, R.D.; Cummings, B.S. Role of the Phospholipase A2 Receptor in Liposome Drug Delivery in Prostate Cancer Cells. Mol. Pharm. 2014, 11, 3443–3451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farid, M.K.; Azizi, F.; He Da Yati, M.; Da Neshpour, M.S.; Shamshiri, A.R.; Siassi, F. Association between cetp taq1b and lipc -514c/t polymorphisms with the serum lipid levels in a group of tehran’s population: A cross sectional study. Lipids Health Dis. 2010, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Talmud, P.J.; Emma, H.; Steve, M.; Michael, O.; Miller, G.J.; Rubin, E.M.; Pennacchio, L.A.; Humphries, S.E. Relative contri-bution of variation within the apoc3/a4/a5 gene cluster in determining plasma triglycerides. Hum. Mol. Genet. 2002, 24, 3039–3046. [Google Scholar] [CrossRef] [Green Version]
- Fisher, C.; Beglova, N.; Blacklow, S.C. Structure of an ldlr-rap complex reveals a general mode for ligand recognition by lipo-protein receptors. Mol. Cell. 2006, 22, 277–283. [Google Scholar] [CrossRef]
- da Silva, T.C.; Polli, J.E.; Swaan, P.W. The solute carrier family 10 (SLC10): Beyond bile acid transport. Mol. Asp. Med. 2013, 34, 252–269. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Liu, X.; Cui, H.; Liu, R.; Wen, J. Transcriptional insights into key genes and pathways controlling muscle lipid metab-olism in broiler chickens. BMC Genom. 2019, 20, 863. [Google Scholar] [CrossRef] [PubMed]
- Mccann, J.C. Influence of Nutrition on the Muscle Transcriptome and Ruminal Microbiome in Cattle. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Urbana, IL, USA, 2016. [Google Scholar]
- Gabrielsson, B.G. Transcriptome profiling and network analysis of effects in metabolic tissues by dietary herring and of fish-oil in mice. In Proceedings of the 10th Nordic Nutrition Conference, Reykjavik, Iceland, 3–5 June 2012. [Google Scholar]
- O’Neill, H.M. Role of AMP-Activated Protein Kinase in Regulating Skeletal Muscle Metabolism during Exercise. Ph.D. Thesis, The University of Melbourne, Melbourne, Australia, 2013. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content (%) | Components | Content (%) |
|---|---|---|---|
| Pellets | |||
| Corn | 25.24 | Dry matter | 93.17 |
| Alfalfa powder | 25.00 | Crude protein | 13.96 |
| Wheat bran | 7.76 | Crude fiber | 22.74 |
| Rapeseed cake | 6.00 | Ether extract | 3.32 |
| Soybean meal | 8.00 | Crude ash | 9.55 |
| NaHCO3 | 1.00 | Calcium | 0.60 |
| Salt | 1.00 | Phosphorus | 0.35 |
| Premix (1) | 1.00 | Metabolic energy (MJ/kg) (2) | 9.39 |
| Hay | |||
| Alfalfa hay | 10.00 | ||
| Corn straw | 15.00 | ||
| Total | 100.00 |
| Item | GSF (n = 8) | GF (n = 8) | SF (n = 8) | SEM | p Value |
|---|---|---|---|---|---|
| Acetic acid (C2:0) | 37.09 | 35.69 | 35.77 | 1.11 | 0.8570 |
| Propionic acid (C3:0) | 0.79 | 0.63 | 0.57 | 0.14 | 0.8093 |
| Butyric acid (C4:0) | 10.94 | 9.71 | 8.62 | 1.93 | 0.8954 |
| Isobutyric acid (iso-C4:0) | 0.52 | 0.44 | 0.41 | 0.03 | 0.3484 |
| Valeric acid (C5:0) | 0.23 | 0.18 | 0.22 | 0.02 | 0.6214 |
| Isovaleric acid (iso-C5:0) | 0.21 | 0.17 | 0.18 | 0.01 | 0.3305 |
| Hexanoic acid (C6:0) | 0.59 | 0.52 | 0.58 | 0.06 | 0.8885 |
| Total SCFA | 50.37 | 47.35 | 46.36 | 3.17 | 0.8760 |
| Item | GSF (n = 8) | GF (n = 8) | SF (n = 8) | SEM | p Value |
|---|---|---|---|---|---|
| ∑SFA | 1276.39 b (46.02) | 1282.97 b (44.44) | 1677.73 a (49.15) | 49.871 | <0.001 |
| ∑MUFA | 1026.83 c (37.02) | 1210.37 b (41.93) | 1349.95 a (39.55) | 37.488 | <0.001 |
| ∑PUFA | 469.28 a (16.92) | 395.98 b (13.72) | 384.72 b (11.27) | 10.830 | <0.001 |
| ∑SFA/∑PUFA | 2.73 c | 3.25 b | 4.38 a | 0.156 | <0.001 |
| ∑FA | 2773.50 b | 2886.77 b | 3412.94 a | 73.841 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Zhao, X.; Li, Z.; Luo, H.; Zhang, H.; Guo, Y.; Zhang, C.; Ma, Q. Changes of Metabolites and Gene Expression under Different Feeding Systems Associated with Lipid Metabolism in Lamb Meat. Foods 2021, 10, 2612. https://doi.org/10.3390/foods10112612
Wang B, Zhao X, Li Z, Luo H, Zhang H, Guo Y, Zhang C, Ma Q. Changes of Metabolites and Gene Expression under Different Feeding Systems Associated with Lipid Metabolism in Lamb Meat. Foods. 2021; 10(11):2612. https://doi.org/10.3390/foods10112612
Chicago/Turabian StyleWang, Bo, Xingang Zhao, Zhen Li, Hailing Luo, Hao Zhang, Yanping Guo, Can Zhang, and Qing Ma. 2021. "Changes of Metabolites and Gene Expression under Different Feeding Systems Associated with Lipid Metabolism in Lamb Meat" Foods 10, no. 11: 2612. https://doi.org/10.3390/foods10112612