
More than Just Wine: The Nutritional Benefits of Grapevine Leaves

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Fatty Acid Quantification and Calculation of Health Lipid Indices
2.3. Pigment Analysis
2.4. Elemental Profiling
2.5. Statistical Analysis
3. Results
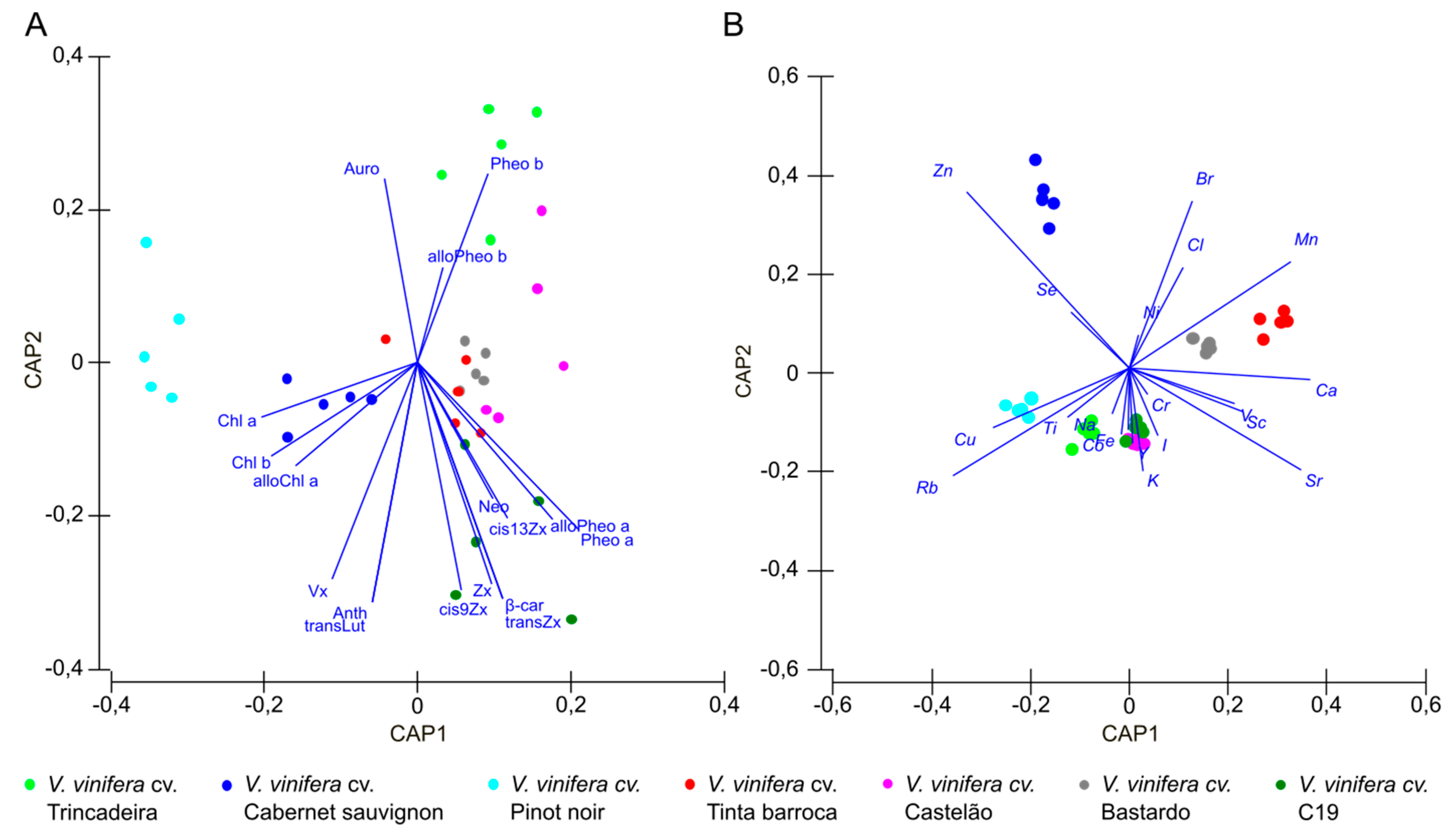
3.1. Pigment Composition
3.2. Elemental Profile
3.3. Fatty Acid Profile
4. Discussion

{kind=link}
|
Elements (μg/g) | Mean Values of Grapevine Leaves | Asparagus [80,81] | Broccoli [82] | Carrot [83] | Cabbage [84] | Chinese Cabbage [84] | Red Cabbage [84] | Savoy Cabbage [84] | Spinach [75] | Watercress [63] |
|---|---|---|---|---|---|---|---|---|---|---|
| Na | 724.00 ± 152.33 | 32 | 360 | NA | 500 | 650 | 270 | 280 | 1200 | 130 |
| Mg | 21.34 ± 32.59 | 170 | 210 | 90 | 1300 | 190 | 160 | 280 | 580 | 270 |
| K | 3515.05 ± 626.60 | 3200 | 3030 | 2400 | 34,500 | 2520 | 2430 | 2300 | 6330 | 2740 |
| Ca | 2139.97 ± 275.87 | 360 | 460 | 340 | 5600 | 1050 | 450 | 350 | 1260 | 1510 |
| Mn | 10.74 ± 0.00 | 2.100 | 1.970 | NA | 1.600 | 1.590 | 2.430 | 1.800 | 87.500 | 2 |
| Fe | 18.77 ± 6.03 | 6.300 | 6.900 | 4 | 53170 | 8 | 8 | 4 | 40 to 350 | 13 |
| Cu | 3.28 ± 0.27 | 2.100 | 0.590 | 0.200 | 0.190 | 0.210 | 0.170 | 0.620 | 1.280 | 2 |
| Zn | 19.89 ± 2.34 | 7.600 | 4.200 | 2 | 1.800 | 1.900 | 2.200 | 2.7 | 5 to 42.500 | 5 |
| Se | 0.01 ± 0.02 | 0.060 | 0.016 | NA | 3 | 0.005 | 0.006 | 0.009 | NA | NA |
| FA | Mean Values of Grapevine Leaves | Asparagus [80,81] | Broccoli [82] | Carrot [83] | Cabbage [84] | Chinese Cabbage [84] | Red Cabbage [84] | Savoy Cabbage [84] | Spinach [75] | Watercress [63] |
|---|---|---|---|---|---|---|---|---|---|---|
| PUFAs (%) | 69.06 ± 3.63 | 37.0 | 25.4 | 69.9 | 25.0 | 69.6 | 70.8 | 64.5 | 64.5 | 81.2 |
| MUFAs (%) | 7.77 ± 1.83 | 22.2 | 16.4 | 8.2 | 25.0 | 10.9 | 10.1 | 14.4 | 14.4 | 2.1 |
| SFAs (%) | 23.17 ± 1.91 | 40.7 | 58.2 | 21.9 | 50.0 | 19.6 | 18.8 | 21.1 | 21.1 | 16.7 |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Organisation of Vine and Wine. 2020 Wine Production. OIV First Estimates. 2020. Available online: https://www.oiv.int/public/medias/7541/en-oiv-2020-world-wine-production-first-estimates.pdf (accessed on 31 July 2021).
- Bordiga, M. Valorization of Wine Making By-Products; Taylor & Francis Group: Boca Raton, FL, USA, 2016. [Google Scholar]
- Albergamo, A.; Costa, R.; Bartolomeo, G.; Rando, R.; Vadalà, R.; Nava, V.; Gervasi, T.; Toscano, G.; Germanò, M.P.; D’angelo, V.; et al. Grape water: Reclaim and valorization of a by-product from the industrial cryoconcentration of grape (Vitis vinifera) must. J. Sci. Food Agric. 2020, 100, 2971–2981. [Google Scholar] [CrossRef]
- Rizzuti, A.; Caliandro, R.; Gallo, V.; Mastrorilli, P.; Chita, G.; Latronico, M. A Combined Approach for Characterisation of Fresh and Brined Vine Leaves by X-Ray Powder Diffraction, NMR Spectroscopy and Direct Infusion High Resolution Mass Spectrometry. Food Chem. 2013, 141, 1908–1915. [Google Scholar] [CrossRef]
- Sat, I.G.; Sengul, M.; Keles, F. Use of Grape Leaves in Canned Food. Pak. J. Nutr. 2002, 1, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Koşar, M.; Küpeli, E.; Malyer, H.; Uylaşer, V.; Türkben, C.; Başer, K.H.C. Effect of Brining on Biological Activity of Leaves of Vitis Vinifera, L. (Cv. Sultani Çekirdeksiz) from Turkey. J. Agric. Food Chem. 2007, 55, 4596–4603. [Google Scholar] [CrossRef]
- Martini, D. Health Benefits of Mediterranean Diet. Nutrients 2019, 11, 1802. [Google Scholar] [CrossRef] [Green Version]
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development; United Nations: New York, NY, USA, 2015. [Google Scholar]
- Goicoechea, N.; Jiménez, L.; Prieto, E.; Gogorcena, Y.; Pascual, I.; Irigoyen, J.J.; Antolín, M.C. Assessment of Nutritional and Quality Properties of Leaves and Musts in Three Local Spanish Grapevine Varieties Undergoing Controlled Climate Change Scenarios. Plants 2021, 10, 1198. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, M.L.; Carpa, R.; Fizeșan, I.; Vlase, L.; Bogdan, C.; Iurian, S.M.; Benedec, D.; Pop, A. Phytochemical Profile and Biological Activities of Tendrils and Leaves Extracts from a Variety of Vitis Vinifera L. Antioxidants 2020, 9, 373. [Google Scholar] [CrossRef]
- Schoedl, K.; Schuhmacher, R.; Forneck, A. Studying the Polyphenols of Grapevine Leaves According to Age and Insertion Level under Controlled Conditions. Sci. Hortic. 2012, 141, 37–41. [Google Scholar] [CrossRef]
- Aguilar, T.; Loyola, C.; de Bruijn, J.; Bustamante, L.; Vergara, C.; von Baer, D.; Mardones, C.; Serra, I. Effect of Thermomaceration and Enzymatic Maceration on Phenolic Compounds of Grape Must Enriched by Grape Pomace, Vine Leaves and Canes. Eur. Food Res. Technol. 2016, 242, 1149–1158. [Google Scholar] [CrossRef]
- Ledesma-Escobar, C.A.; Luque de Castro, M.D. Coverage exploitation of by-products from the agrofood industry. In Green Extraction of Natural Products; Chemat, F., Strube, J., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, German, 2015; pp. 265–306. ISBN 978-3-527-67682-8. [Google Scholar]
- Pari, L.; Suresh, A. Effect of Grape (Vitis vinifera L.) Leaf Extract on Alcohol Induced Oxidative Stress in Rats. Food Chem. Toxicol. 2008, 46, 1627–1634. [Google Scholar] [CrossRef]
- Farhadi, K.; Esmaeilzadeh, F.; Hatami, M.; Forough, M.; Molaie, R. Determination of Phenolic Compounds Content and Antioxidant Activity in Skin, Pulp, Seed, Cane and Leaf of Five Native Grape Cultivars in West Azerbaijan Province, Iran. Food Chem. 2016, 199, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Katalinic, V.; Mozina, S.S.; Generalic, I.; Skroza, D.; Ljubenkov, I.; Klancnik, A. Phenolic Profile, Antioxidant Capacity, and Antimicrobial Activity of Leaf Extracts from Six Vitis vinifera L. Varieties. Int. J. Food Prop. 2013, 16, 45–60. [Google Scholar] [CrossRef]
- Pintać, D.; Četojević-Simin, D.; Berežni, S.; Orčić, D.; Mimica-Dukić, N.; Lesjak, M. Investigation of the Chemical Composition and Biological Activity of Edible Grapevine (Vitis vinifera L.) Leaf Varieties. Food Chem. 2019, 286, 686–695. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.; Bento, A.; Baraldi, I.; Malheiro, R. Selection of Grapevine Leaf Varieties for Culinary Process Based on Phytochemical Composition and Antioxidant Properties. Food Chem. 2016, 212, 291–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, B.; Correia, A.C.; Cosme, F.; Nunes, F.M.; Jordão, A.M. Volatile Components of Vine Leaves from Two Portuguese Grape Varieties (Vitis vinifera L.), Touriga Nacional and Tinta Roriz, Analysed by Solid-Phase Microextraction. Nat. Prod. Res. 2015, 29, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Anđelković, M.; Radovanović, B.; Anđelković, A.M.; Radovanović, V. Phenolic Compounds and Bioactivity of Healthy and Infected Grapevine Leaf Extracts from Red Varieties Merlot and Vranac (Vitis vinifera L.). Plant Foods Hum. Nutr. 2015, 70, 317–323. [Google Scholar] [CrossRef]
- Pantelić, M.M.; Zagorac, D.Č.D.; Ćirić, I.Ž.; Pergal, M.V.; Relić, D.J.; Todić, S.R.; Natić, M.M. Phenolic Profiles, Antioxidant Activity and Minerals in Leaves of Different Grapevine Varieties Grown in Serbia. J. Food Compos. Anal. 2017, 62, 76–83. [Google Scholar] [CrossRef]
- Ali, K.; Maltese, F.; Choi, Y.H.; Verpoorte, R. Metabolic Constituents of Grapevine and Grape-Derived Products. Phytochem. Rev. 2010, 9, 357–378. [Google Scholar] [CrossRef] [Green Version]
- Dani, C.; Oliboni, L.S.; Agostini, F.; Funchal, C.; Serafini, L.; Henriques, J.A.; Salvador, M. Phenolic Content of Grapevine Leaves (Vitis Labrusca Var. Bordo) and Its Neuroprotective Effect against Peroxide Damage. Toxicol. In Vitr. 2010, 24, 148–153. [Google Scholar] [CrossRef]
- Orhan, D.D.; Orhan, N.; Ozcelik, B.; Ergun, F. Biological Activities of Vitis vinifera L. Leaves. Turk. J. Biol. 2009, 33, 341–348. [Google Scholar]
- Fernandes, F.; Ramalhosa, E.; Pires, P.; Verdial, J.; Valentão, P.; Andrade, P.; Bento, A.; Pereira, J.A. Vitis vinifera Leaves towards Bioactivity. Ind. Crop. Prod. 2013, 43, 434–440. [Google Scholar] [CrossRef]
- Lacerda, D.S.; Costa, P.C.; Funchal, C.; Dani, C.; Gomez, R. Benefits of vine leaf on different biological systems. In Grape and Wine Biotechnology; Morata, A., Loira, I., Eds.; IntechOpen: London, UK, 2016; pp. 125–143. [Google Scholar]
- Handoussa, H.; Hanafi, R.; Eddiasty, I.; El-Gendy, M.; El Khatib, A.; Linscheid, M.; Mahran, L.; Ayoub, N. Anti-Inflammatory and Cytotoxic Activities of Dietary Phenolics Isolated from Corchorus Olitorius and Vitis vinifera. J. Funct. Foods 2013, 5, 1204–1216. [Google Scholar] [CrossRef]
- Lima, A.; Pereira, J.A.; Baraldi, I.; Malheiro, R. Cooking Impact in Color, Pigments and Volatile Composition of Grapevine Leaves (Vitis vinifera L. Var. Malvasia Fina and Touriga Franca). Food Chem. 2017, 221, 1197–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maia, M.; Ferreira, A.E.N.; Laureano, G.; Marques, A.P.; Torres, V.M.; Silva, A.B.; Matos, A.R.; Cordeiro, C.; Figueiredo, A.; Silva, M.S. Vitis vinifera “Pinot Noir” Leaves as a Source of Bioactive Nutraceutical Compounds. Food Funct. 2019, 10, 3822–3827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIV. Focus OIV 2017. Distribution of the World’s Grapevine Varieties. 2017. Available online: https://www.oiv.int/public/medias/5888/en-distribution-of-the-worlds-grapevine-varieties.pdf (accessed on 31 July 2021).
- Coutinho, M.P. Melhoramento Da Videira. Seu Aspecto Particular Da Resistência à Plasmopora Viticola. Ann. Junta Nac. Vin. 1950, 2, 13–135. [Google Scholar]
- Coutinho, M.P. Some Vine Clones Resistant to Plasmopora. Vitis 1964, 4, 341–346. [Google Scholar]
- Laureano, G.; Cavaco, A.R.; Matos, A.R.; Figueiredo, A. Fatty Acid Desaturases: Uncovering Their Involvement in Grapevine Defence against Downy Mildew. Int. J. Mol. Sci. 2021, 22, 5473. [Google Scholar] [CrossRef] [PubMed]
- Laureano, G.; Figueiredo, J.; Cavaco, A.R.; Duarte, B.; Caçador, I.; Malhó, R.; Sousa Silva, M.; Matos, A.R.; Figueiredo, A. The Interplay between Membrane Lipids and Phospholipase A Family Members in Grapevine Resistance against Plasmopara Viticola. Sci. Rep. 2018, 8, 14538. [Google Scholar] [CrossRef]
- Garaffo, M.A.; Vassallo-Agius, R.; Nengas, Y.; Lembo, E.; Rando, R.; Maisano, R.; Dugo, G.; Giuffrida, D. Fatty Acids Profile, Atherogenic (IA) and Thrombogenic (IT) Health Lipid Indices, of Raw Roe of Blue Fin Tuna (Thunnus thynnus L.) and Their Salted Product “Bottarga”. Food Nutr. Sci. 2011, 2, 736–743. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Liu, H. Nutritional Indices for Assessing Fatty Acids: A Mini-Review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef]
- Attia, Y.A.; Al-Harthi, M.A.; Korish, M.A.; Shiboob, M.M. Fatty Acid and Cholesterol Profiles and Hypocholesterolemic, Atherogenic, and Thrombogenic Indices of Table Eggs in the Retail Market. Lipids Health Dis. 2015, 14, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, C.E.; Vasconcelos, M.A.; da, S.; de Almeida Ribeiro, M.; Sarubbo, L.A.; Andrade, S.A.C.; de Melo Filho, A.B. Nutritional and Lipid Profiles in Marine Fish Species from Brazil. Food Chem. 2014, 160, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Wołoszyn, J.; Haraf, G.; Okruszek, A.; Wereńska, M.; Goluch, Z.; Teleszko, M. Fatty Acid Profiles and Health Lipid Indices in the Breast Muscles of Local Polish Goose Varieties. Poult. Sci. 2020, 99, 1216–1224. [Google Scholar] [CrossRef] [PubMed]
- Cabrita, M.T.; Gameiro, C.; Utkin, A.B.; Duarte, B.; Caçador, I.; Cartaxana, P. Photosynthetic Pigment Laser-Induced Fluorescence Indicators for the Detection of Changes Associated with Trace Element Stress in the Diatom Model Species Phaeodactylum Tricornutum. Environ. Monit. Assess. 2016, 188, 285. [Google Scholar] [CrossRef] [PubMed]
- Cabrita, M.T.; Duarte, B.; Gameiro, C.; Godinho, R.M.; Caçador, I. Photochemical Features and Trace Element Substituted Chlorophylls as Early Detection Biomarkers of Metal Exposure in the Model Diatom Phaeodactylum Tricornutum. Ecol. Indic. 2018, 95, 1038–1052. [Google Scholar] [CrossRef]
- Feijão, E.; Gameiro, C.; Franzitta, M.; Duarte, B.; Caçador, I.; Cabrita, M.T.; Matos, A.R. Heat Wave Impacts on the Model Diatom Phaeodactylum Tricornutum: Searching for Photochemical and Fatty Acid Biomarkers of Thermal Stress. Ecol. Indic. 2018, 95, 1026–1037. [Google Scholar] [CrossRef]
- Küpper, H.; Seibert, S.; Parameswaran, A. Fast, Sensitive, and Inexpensive Alternative to Analytical Pigment HPLC: Quantification of Chlorophylls and Carotenoids in Crude Extracts by Fitting with Gauss Peak Spectra. Anal. Chem. 2007, 79, 7611–7627. [Google Scholar] [CrossRef]
- Duarte, B.; Caetano, M.; Almeida, P.R.; Vale, C.; Caçador, I. Accumulation and Biological Cycling of Heavy Metal in Four Salt Marsh Species, from Tagus Estuary (Portugal). Environ. Pollut. 2010, 158, 1661–1668. [Google Scholar] [CrossRef]
- Towett, E.K.; Shepherd, K.D.; Cadisch, G. Quantification of Total Element Concentrations in Soils Using Total X-Ray Fluorescence Spectroscopy (TXRF). Sci. Total Environ. 2013, 463–464, 374–388. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.; Gorley, R.N. Primer v6: User Manual/Tutorial; Plymouth Routines in Multivariate Ecological Research PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Duarte, B.; Pedro, S.; Marques, J.C.; Adão, H.; Caçador, I. Zostera Noltii Development Probing Using Chlorophyll a Transient Analysis (JIP-Test) under Field Conditions: Integrating Physiological Insights into a Photochemical Stress Index. Ecol. Indic. 2017, 76, 219–229. [Google Scholar] [CrossRef]
- Duarte, B.; Cabrita, M.T.; Vidal, T.; Pereira, J.L.; Pacheco, M.; Pereira, P.; Canário, J.; Gonçalves, F.J.M.; Matos, A.R.; Rosa, R.; et al. Phytoplankton Community-Level Bio-Optical Assessment in a Naturally Mercury Contaminated Antarctic Ecosystem (Deception Island). Mar. Environ. Res. 2018, 140, 412–421. [Google Scholar] [CrossRef]
- Lila, M.A. Plant Pigments and Human Health. In Annual Plant Reviews Online; Roberts, J.A., Ed.; John Wiley and Sons: Hoboken, NJ, USA, 2018; pp. 248–274. ISBN 978-1-119-31299-4. [Google Scholar]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Covaci, E.; Senila, M.; Leopold, L.F.; Olah, N.-K.; Cobzac, C.; Ivanova-Petropulos, V.; Balabanova, B.; Cadar, O.; Becze, A.; Ponta, M.; et al. Characterization of Lycium Barbarum L. Berry Cultivated in North Macedonia: A Chemometric Approach. J. Berry Res. 2020, 10, 223–241. [Google Scholar] [CrossRef]
- Candela, C.G.; López, L.M.B.; Kohen, V.L. Importance of a Balanced Omega 6/Omega 3 Ratio for the Maintenance of Health: Nutritional Recommendations. Nutr. Hosp. 2011, 26, 323–329. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.I.; Marcone, M.; Baptista, J. Health-Promoting Ingredients from Four Selected Azorean Macroalgae. Food Res. Int. 2016, 89, 432–438. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations; International Organisation of Vine and Wine Intergovernmental Organisation (OIV). Table and Dried Grapes: FAO and OIV FOCUS 2016. 2019. Available online: http://www.fao.org/3/i7042e/i7042e.pdf (accessed on 31 July 2021).
- Gurbuz, Y. Determination of Nutritive Value of Leaves of Several Vitis vinifera Varieties as a Source of Alternative Feedstuff for Sheep Using in Vitro and in Situ Measurements. Small Rumin. Res. 2007, 71, 59–66. [Google Scholar] [CrossRef]
- Sridhar, K.; Charles, A.L. Discrimination of Kyoho Grape (Vitis Labruscana) Skin, Seed and Flesh Antioxidant Activities by Solvent Extraction: Application of Advanced Chemometrics. Int. J. Food Sci. Technol. 2021. [Google Scholar] [CrossRef]
- Urquiaga, I.; D’Acuña, S.; Pérez, D.; Dicenta, S.; Echeverría, G.; Rigotti, A.; Leighton, F. Wine Grape Pomace Flour Improves Blood Pressure, Fasting Glucose and Protein Damage in Humans: A Randomized Controlled Trial. Biol. Res. 2015, 48, 49. [Google Scholar] [CrossRef] [Green Version]
- Harb, J.; Alseekh, S.; Tohge, T.; Fernie, A.R. Profiling of Primary Metabolites and Flavonols in Leaves of Two Table Grape Varieties Collected from Semiarid and Temperate Regions. Phytochemistry 2015, 117, 444–455. [Google Scholar] [CrossRef] [PubMed]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Commercial Dietary Antioxidant Supplements Assayed for Their Antioxidant Activity by Different Methodologies. J. Agric. Food Chem. 2003, 51, 2512–2519. [Google Scholar] [CrossRef] [PubMed]
- Gupta, U.C.; Gupta, S.C. Sources and Deficiency Diseases of Mineral Nutrients in Human Health and Nutrition: A Review. Pedosphere 2014, 24, 13–38. [Google Scholar] [CrossRef]
- Stein, A.J. Global Impacts of Human Mineral Malnutrition. Plant Soil 2010, 335, 133–154. [Google Scholar] [CrossRef]
- Chellan, P.; Sadler, P.J. The Elements of Life and Medicines. Philos. Trans. R. Soc. Math. Phys. Eng. Sci. 2015, 373, 20140182. [Google Scholar] [CrossRef] [PubMed]
- Pinela, J.; Carvalho, A.M.; Ferreira, I.C.F.R. Watercress. In Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Jaiswal, A.K., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 197–219. ISBN 978-0-12-812780-3. [Google Scholar]
- Pejin, B.; Stanimirovic, B.; Vujovic, D.; Djordjevic, J.P.; Velickovic, M.; Tesevic, V. The Natural Product Content of the Selected Cabernet Franc Wine Samples Originating from Serbia: A Case Study of Phenolics. Nat. Prod. Res. 2016, 30, 1762–1765. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Guideline: Potassium Intake for Adults and Children. 2012. Available online: https://www.who.int/publications/i/item/9789241504829 (accessed on 31 July 2021).
- Orwoll, E.S. Overall approach to the evaluation and treatment of osteoporosis in men. In Osteoporosis in Men, 2nd ed.; Orwoll, E.S., Bilezikian, J.P., Vanderschueren, D., Eds.; Academic Press: San Diego, CA, USA, 2010; pp. 589–601. ISBN 978-0-12-374602-3. [Google Scholar]
- Fraga, C.G. Relevance, Essentiality and Toxicity of Trace Elements in Human Health. Mol. Asp. Med. 2005, 26, 235–244. [Google Scholar] [CrossRef]
- World Health Organization. Worldwide Prevalence of Anaemia 1993–2005: WHO Global Database on Anaemia; de Benoist, B., McLean, E., Egli, I., Cogswell, M., Eds.; WHO Press: Geneva, Switzerland, 2008; ISBN 978-92-4-159665-7. [Google Scholar]
- Ancuceanu, R.; Dinu, M.; Hovaneţ, M.V.; Anghel, A.I.; Popescu, C.V.; Negreş, S. A Survey of Plant Iron Content—A Semi-Systematic Review. Nutrients 2015, 7, 10320–10351. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Guideline: Daily Iron Supplementation in Adult Women and Adolescent Girls. 2016. Available online: https://apps.who.int/iris/handle/10665/204761 (accessed on 31 July 2021).
- Institute of Medicine (US) Panel on Micronutrients. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academies Press: Washington, DC, USA, 2001; ISBN 978-0-309-07279-3. [Google Scholar]
- World Health Organization. Zinc in Drinking-Water: Background Document for Development of WHO Guidelines for Drinking-Water Quality. 2003. Available online: https://www.who.int/water_sanitation_health/dwq/chemicals/zinc.pdf (accessed on 31 July 2021).
- Vannice, G.; Rasmussen, H. Position of the Academy of Nutrition and Dietetics: Dietary Fatty Acids for Healthy Adults. J. Acad. Nutr. Diet. 2014, 114, 136–153. [Google Scholar] [CrossRef] [Green Version]
- Institute of Medicine of the National Academies. Dietary Reference Intakes: The Essential Guide to Nutrient Requirements; The National Academies Press: Washington, DC, USA, 2006; ISBN 978-0-309-15742-1. [Google Scholar]
- Murcia, M.A.; Jiménez-Monreal, A.M.; Gonzalez, J.; Martínez-Tomé, M. Spinach. In Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Jaiswal, A.K., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 181–195. ISBN 978-0-12-812780-3. [Google Scholar]
- Pereira, C.; Barros, L.; Carvalho, A.M.; Ferreira, I.C.F.R. Nutritional Composition and Bioactive Properties of Commonly Consumed Wild Greens: Potential Sources for New Trends in Modern Diets. Food Res. Int. 2011, 44, 2634–2640. [Google Scholar] [CrossRef]
- Sánchez-Mata, M.d.C.; Tardío, J. (Eds.) Mediterranean Wild Edible Plants: Ethnobotany and Food Composition Tables; Springer-Verlag: New York, NY, USA, 2016; ISBN 978-1-4939-3327-3. [Google Scholar]
- Kang, M.J.; Lee, E.K.; Lee, S.S. Effects of Two P/S Ratios with Same Peroxidizability Index Value and Antioxidants Supplementation on Serum Lipid Concentration and Hepatic Enzyme Activities of Rats. Clin. Chim. Acta 2004, 350, 79–87. [Google Scholar] [CrossRef]
- Saito, M. Dietary Docosahexaenoic Acid Does Not Promote Tissue Lipid Peroxide Formation to the Extent Expected from the Peroxidizability Index of the Lipids. BioFactors 2000, 13, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, A.; Jaramillo-Carmona, S.; Rodríguez-Arcos, R.; Jiménez-Araujo, A.; Guillén-Bejarano, R. Asparagus. In Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Jaiswal, A.K., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 121–140. ISBN 978-0-12-812780-3. [Google Scholar]
- Determination of Fatty Acid in Asparagus by Gas Chromatography. Available online: http://actavet.org/en/determination-of-fatty-acid-in-asparagus-by-gas-chromatography-13164 (accessed on 26 July 2021).
- Nagraj, G.S.; Chouksey, A.; Jaiswal, S.; Jaiswal, A.K. Broccoli. In Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Jaiswal, A.K., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 5–17. ISBN 978-0-12-812780-3. [Google Scholar]
- Nagraj, G.S.; Jaiswal, S.; Harper, N.; Jaiswal, A.K. Carrot. In Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Jaiswal, A.K., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 323–337. ISBN 978-0-12-812780-3. [Google Scholar]
- Moreb, N.; Murphy, A.; Jaiswal, S.; Jaiswal, A.K. Cabbage. In Nutritional Composition and Antioxidant Properties of Fruits and Vegetables; Jaiswal, A.K., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 33–54. ISBN 978-0-12-812780-3. [Google Scholar]
- Mulabagal, V.; Ngouajio, M.; Nair, A.; Zhang, Y.; Gottumukkala, A.L.; Nair, M.G. In Vitro Evaluation of Red and Green Lettuce (Lactuca Sativa) for Functional Food Properties. Food Chem. 2010, 118, 300–306. [Google Scholar] [CrossRef]
- Bailey, S.; Horton, P.; Walters, R.G. Acclimation of Arabidopsis Thaliana to the Light Environment: The Relationship between Photosynthetic Function and Chloroplast Composition. Planta 2004, 218, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.V.; Rao, L.G. Carotenoids and Human Health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Podsędek, A. Natural Antioxidants and Antioxidant Capacity of Brassica Vegetables: A Review. LWT Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Murillo, A.G.; Hu, S.; Fernandez, M.L. Zeaxanthin: Metabolism, Properties, and Antioxidant Protection of Eyes, Heart, Liver, and Skin. Antioxidants 2019, 8, 390. [Google Scholar] [CrossRef] [Green Version]
- Nwachukwu, I.D.; Udenigwe, C.C.; Aluko, R.E. Lutein and Zeaxanthin: Production Technology, Bioavailability, Mechanisms of Action, Visual Function, and Health Claim Status. Trends Food Sci. Technol. 2016, 49, 74–84. [Google Scholar] [CrossRef]
- Molldrem, K.L.; Li, J.; Simon, P.W.; Tanumihardjo, S.A. Lutein and β-Carotene from Lutein-Containing Yellow Carrots Are Bioavailable in Humans. Am. J. Clin. Nutr. 2004, 80, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Reig, G.; Iglesias, I.; Gatius, F.; Alegre, S. Antioxidant Capacity, Quality, and Anthocyanin and Nutrient Contents of Several Peach Cultivars [Prunus Persica (L.) Batsch] Grown in Spain. J. Agric. Food Chem. 2013, 61, 6344–6357. [Google Scholar] [CrossRef]

| Subspecies (Subsp.) and/or Cultivar (cv.) | VIVC Variety Number | Type of Accession | Origin | Colour of Berry Skin | |
|---|---|---|---|---|---|
| Vitis vinifera cv. | Subsp. sativa cv. Trincadeira | 15,685 | Unknown Natural hybridisation | Portugal | Noir |
| Subsp. sativa cv. Cabernet sauvignon | 1929 | Natural hybridisation Cabernet Franc X Sauvignon | France | Noir | |
| Subsp. sativa cv. Pinot Noir | 9279 | Savagnin Blanc X? | France | Noir | |
| Subsp. sativa cv. Tinta Barroca | 12,462 | Natural hybridisation Marufo X Touriga Nacional | Portugal | Noir | |
| Subsp. sativa cv. Castelão | 2324 | Natural hybridisation Cayetana Blanca X Alfrocheiro Preto | Portugal | Noir | |
| Subsp. sativa cv. Bastardo | 12,668 | Natural hybridisation (? X Savagnin = Traminer | Portugal | Noir | |
| C19 * | Not registered on VIVC | Hybrid crossing (Jaen T X?) X Jaen T) | Portugal | Noir |
| Vitis vinifera cv. | |||||||
|---|---|---|---|---|---|---|---|
| Pigment (µg/g FW) | Trincadeira | Cabernet Sauvignon | Pinot Noir | Tinta Barroca | Castelão | Bastardo | C19 |
| Chlorophyll a | 230.69 ± 46.00 a | 373.49 ± 35.05 b | 374.26 ± 34.95 b | 254.09 ± 65.26 ab | 283.53 ± 67.89 ab | 318.29 ± 35.10 ab | 269.32 ± 74.30 ab |
| Chlorophyll b | 203.95 ± 30.35 a | 281.04 ± 17.48 a | 276.95 ± 16.52 a | 198.53 ± 31.47 a | 214.81 ± 33.91 a | 211.28 ± 20.01 a | 259.17 ± 54.30 a |
| Pheophytin a | 27.62 ± 8.60 c | 33.58 ± 13.87 abc | 13.83 ± 12.58 ac | 57.96 ± 9.53 bc | 51.20 ± 13.19 abc | 48.69 ± 8.10 abc | 75.39 ± 17.21 b |
| Pheophytin b | 20.34 ± 14.17 a | 3.38 × 10−23 ± 6.05 × 10−23 bcde | 5.22 × 10−24 ± 7.67 × 10−24 bcde | 0.16 ±0.35 abcde | 3.44 ± 6.07 abcde | 6.45 × 10−24 ± 1.06 × 10−23 bcde | 1.12 × 10−23 ± 1.94 × 10−23 bcde |
| alloChlorophyll a | 65.19 ± 23.48 ab | 120.56 ± 35.32 b | 116.10 ± 28.55 b | 75.40 ± 20.53 ab | 59.89 ± 40.98 ab | 37.26 ± 11.78 a | 117.88 ± 29.52 b |
| alloPheophytin a | 35.74 ± 30.92 a | 63.33 ± 40.74 ab | 28.33 ± 42.46 ab | 102.95 ± 13.09 ab | 95.66 ± 11.46 ab | 111.80 ±15.26 b | 121.79 ± 29.56 b |
| alloPheophytin b | 3.92 ± 8.77 a | 3.67 × 10−23 ± 7.66 × 10−23 a | 3.25 × 10−24 ± 3.05 × 10−24 a | 3.05 × 10−24 ± 4.43 × 10−24 a | 1.41 × 10−24 ± 1.47 × 10−24 a | 1.56 × 10−23 ± 1.28 × 10−23 a | 5.43 × 10−24 ± 7.65 × 10−24 a |
| Auroxanthin | 61.48 ± 8.17 b | 27.51 ± 18.36 ab | 55.24 ± 12.02 ab | 34.16 ± 15.44 ab | 48.22 ± 27.32 ab | 45.44 ± 20.01 ab | 9.07 ± 10.40 a |
| Antheraxanthin | 10.61 ± 3.10 a | 27.57 ± 4.37 ab | 27.07 ± 6.68 ab | 26.56 ± 2.57 ab | 21.44 ± 4.52 ab | 25.18 ± 2.21 ab | 35.60 ± 5.48 b |
| β-carotene | 7.10 ± 2.70 a | 16.99 ± 2.29 ab | 12.46 ± 4.48 a | 20.38 ± 2.51 ab | 18.19 ± 3.22 ab | 20.32± 1.29 ab | 29.42 ± 4.56 b |
| Trans-Lutein | 10.32 ± 3.02 a | 26.81 ± 4.25 ab | 26.33 ± 6.50 ab | 25.83 ± 2.50 ab | 20.86 ± 4.40 ab | 24.49 ± 2.15 ab | 34.63 ± 5.33 b |
| Neoxanthin | 5.50 ± 4.38 a | 6.62 ± 7.59 a | 9.16 ± 7.91 a | 22.10 ± 3.99 a | 13.87 ± 8.36 a | 14.70 ± 3.16 a | 23.24 ± 10.96 a |
| Violaxanthin | 11.56 ± 4.42 a | 32.80 ± 7.94 abc | 35.07 ± 8.79 bc | 30.39 ± 2.17 abc | 22.50 ± 5.74 ab | 28.47 ± 3.15 abc | 40.58 ± 7.84 c |
| Trans-Zeaxanthin | 7.52 ± 2.86 a | 18.00 ± 2.42 ab | 13.20 ± 4.75 a | 21.59 ± 2.66 ab | 19.27 ± 3.41 ab | 21.54 ± 1.36 ab | 31.17 ± 4.87 b |
| Cis-9-Zeaxanthin | 0.58 ± 1.30 a | 16.65 ± 5.20 abc | 10.09 ± 7.69 ab | 22.41 ± 7.06 bc | 12.12 ± 12.32 abc | 15.82 ± 7.02 abc | 35.37 ± 5.68 c |
| Cis-13-Zeaxanthin | 3.31 ± 3.04 ab | 5.41 ± 5.44 ab | 1.78 ± 3.21 a | 14.17 ± 8.98 ab | 7.61 ± 10.46 ab | 5.01 ± 5.53 ab | 21.95 ± 10.08 b |
| Zeaxanthin | 11.42 ± 3.73 a | 40.05 ± 8.89 abc | 25.07 ± 13.44 ab | 58.17 ± 17.87 bc | 39.00 ± 25.37 abc | 42.37 ± 12.90 abc | 88.49 ± 18.34 c |
| Vitis vinifera cv. | |||||||
|---|---|---|---|---|---|---|---|
| Element (μg/g FW) | Trincadeira | Cabernet Sauvignon | Pinot Noir | Tinta Barroca | Castelão | Bastardo | C19 |
| Na | 491.72 ± 139.27 a | 661.66 ± 74.64 ab | 898.26 ± 288.80 ab | 746.88 ± 74.99 ab | 1033.40 ± 194.20 b | 571.39 ± 198.90 ab | 671.68 ± 95.52 ab |
| Mg | 21.59 ± 33.32 a | 14.64 ± 32.72 a | 28.61 ± 39.30 a | 0.00 a | 55.58 ± 57.97 a | 18.51 ± 41.39 a | 10.48 ± 23.44 a |
| Cl | 21.27 ± 5.69 abc | 29.01 ± 8.31 bc | 9.67 ± 6.19 a | 23.84 ± 6.31 abc | 9.25 ± 3.28 a | 25.08 ± 8.71 abc | 30.02 ± 7.51 c |
| K | 4046.22 ± 821.37 a | 2817.77 ± 336.38 a | 3446.68 ± 1054.71 a | 3224.14 ± 252.62 a | 3818.64 ± 1174.17 a | 3784.25 ± 552.03 a | 3467.66 ± 194.97 a |
| Ca | 2080.56 ± 392.42 abc | 1691.01 ± 106.95 a | 1580.98 ± 431.74 a | 2771.00 ± 327.37 c | 1752.66 ± 379.56 abc | 2513.26 ± 143.36 abc | 2590.35 ± 149.68 bc |
| Sc | 5.84 ± 1.41 abc | 4.61 ± 0.63 ab | 3.59 ± 0.82 a | 6.65 ± 0.72 bc | 4.86 ± 0.74 abc | 6.02 ± 1.10 abc | 8.65 ± 2.17 c |
| Ti | 0.26 ± 0.25 a | 0.12 ± 0.17 a | 2.39 ± 3.94 a | 0.27 ± 0.28 a | 1.10 ± 0.92 a | 0.00 a | 0.30 ± 0.31 a |
| V | 0.17 ± 0.11 ab | 0.12 ± 0.12 ab | 0.03 ± 0.06 a | 0.26 ± 0.16 ab | 0.19 ± 0.13 ab | 0.150 ± 0.102 ab | 0.31 ± 0.04 b |
| Cr | 0.00 a | 0.00 a | 0.02 ± 0.05 a | 0.03 ± 0.06 a | 0.04 ± 0.09 a | 0.00 a | 0.00 a |
| Mn | 9.30 ± 1.81 abc | 11.77 ± 0.69 abcd | 6.50 ± 0.80 a | 13.01 ± 1.48 cd | 9.08 ± 1.29 abc | 13.24 ± 0.50 d | 12.28 ± 0.42 bcd |
| Fe | 19.79 ± 6.83 a | 15.99 ± 1.62 a | 14.85 ± 3.16 a | 15.32 ± 1.06 a | 29.26 ± 27.19 a | 15.40 ± 1.01 a | 20.77 ± 1.33 a |
| Co | 0.16 ± 0.10 a | 0.07 ± 0.06 a | 0.10 ± 0.10 a | 0.06 ± 0.08 a | 0.20 ± 0.23 a | 0.15 ± 0.05 a | 0.10 ± 0.07 a |
| Ni | 0.13 ± 0.04 a | 0.11 ± 0.03 a | 0.10 ± 0.09 a | 0.11 ± 0.07 a | 0.04 ± 0.05 a | 0.12 ± 0.05 a | 0.12 ± 0.04 a |
| Cu | 3.26 ± 0.66 ab | 3.46 ± 0.11 ab | 3.69 ± 0.18 ab | 2.47 ± 0.11 a | 3.06 ± 0.38 ab | 2.49 ± 0.18 a | 4.51 ± 0.30 b |
| Zn | 11.16 ± 2.04 ab | 55.44 ± 5.41 b | 30.70 ± 3.70 ab | 9.41 ± 0.92 a | 9.53 ± 2.43 a | 9.31 ± 1.23 a | 13.66 ± 0.65 ab |
| Se | 0.00 ± 0.00 a | 0.02 ± 0.02 a | 0.02 ± 0.02 a | 0.00 a | 0.01 ± 0.03 a | 0.02 ± 0.04 a | 0.00 a |
| Br | 0.45 ± 0.10 abcd | 1.39 ± 0.40 d | 0.14 ± 0.03 a | 1.18 ± 0.071 cd | 0.23 ± 0.09 abc | 0.61 ± 0.38 abcd | 0.97 ± 0.43 bcd |
| Rb | 6.57 ± 1.46 cd | 5.18 ± 0.29 abcd | 7.16 ± 1.00 d | 2.90 ± 0.15 a | 5.83 ± 1.31 abcd | 4.57 ± 0.98 abcd | 6.04 ± 0.52 bcd |
| Sr | 4.79 ± 1.05 abc | 3.46 ± 0.35 a | 4.33 ± 0.56 ab | 6.39 ± 0.36 c | 5.71 ± 1.02 abc | 5.66 ± 0.96 abc | 6.31 ± 0.35 bc |
| Y | 2.84 ± 0.64 ab | 2.88 ± 0.51 ab | 4.22 ± 0.91 ab | 3.89 ± 0.60 ab | 4.29 ± 0.51 b | 2.72 ± 0.45 a | 3.37 ± 0.31 ab |
| I | 0.34 ± 0.54 a | 0.00 a | 0.27 ± 0.59 a | 0.56 ± 1.24 a | 1.20 ± 2.21 a | 0.20 ± 0.44 a | 1.11 ± 0.63 a |
| Vitis vinifera cv. | |||||||
|---|---|---|---|---|---|---|---|
| Fatty Acids and Derived Parameters | Trincadeira | Cabernet Sauvignon | Pinot Noir | Tinta Barroca | Castelão | Bastardo | C19 |
| C16:0 (%) | 21.22 ± 1.73 | 20.25 ± 0.74 | 21.36 ± 1.69 | 20.25 ± 0.59 | 19.21 ± 1.39 | 19.92 ± 2.09 | 17.92 ± 0.94 |
| C16:1t (%) | 2.64 ± 0.22 | 2.38 ± 0.12 | 2.78 ± 0.30 | 2.79 ± 0.20 | 2.89 ± 0.35 | 5.62 ± 1.51 | 3.96 ± 1.37 |
| C18:0 (%) | 1.95 ± 0.32 | 2.91 ± 1.94 | 2.28 ± 0.73 | 1.64 ± 0.28 | 2.17 ± 0.60 | 2.61 ± 0.93 | 3.97 ± 1.73 |
| C18:1 (%) | 5.14 ± 1.99 | 5.17 ± 0.46 | 5.33 ± 0.60 | 4.05 ± 1.14 | 5.40 ± 1.93 | 5.94 ± 1.98 | 6.99 ± 2.50 |
| C18:2 (%) | 23.71 ± 3.87 | 24.51 ± 1.06 | 24.71 ± 1.35 | 21.15 ± 3.12 | 20.91 ± 3.32 | 22.56 ± 0.73 | 22.88 ± 0.61 |
| C18:3 (%) | 45.35 ± 7.39 | 44.78 ± 1.90 | 43.53 ± 2.13 | 50.13 ± 4.24 | 49.42 ± 5.26 | 43.34 ± 2.90 | 44.27 ± 5.45 |
| PUFAS (%) | 69.06 ± 3.63 | 69.29 ± 2.72 | 68.24 ± 2.16 | 71.28 ± 1.14 | 70.33 ± 2.19 | 65.90 ± 2.59 | 67.15 ± 6.03 |
| SFA (%) | 23.17 ± 1.91 | 23.16 ± 2.43 | 23.65 ± 1.93 | 21.89 ± 0.42 | 21.38 ± 1.27 | 22.54 ± 1.52 | 21.89 ± 2.31 |
| MUFAS (%) | 7.77 ± 1.83 | 7.55 ± 0.35 | 8.11 ± 0.41 | 6.84 ± 0.97 | 8.29 ± 1.63 | 11.56 ± 3.04 | 10.95 ± 3.87 |
| PUFA/SFA (%) | 3.01 ± 0.41 | 3.03 ± 0.39 | 2.91 ± 0.36 | 3.26 ± 0.10 | 3.30 ± 0.28 | 2.93 ± 0.22 | 3.11 ± 0.56 |
| AI | 0.30 ± 0.03 | 0.30 ± 0.04 | 0.31 ± 0.03 | 0.28 ± 0.01 | 0.27 ± 0.02 | 0.29 ± 0.03 | 0.28 ± 0.04 |
| TI | 0.15 ± 0.03 | 0.15 ± 0.02 | 0.16 ± 0.02 | 0.13 ± 0.01 | 0.13 ± 0.02 | 0.15 ± 0.01 | 0.15 ± 0.03 |
| Cox | 12.29 ± 1.19 | 12.25 ± 0.49 | 12.00 ± 0.44 | 13.05 ± 0.58 | 12.88 ± 0.79 | 11.74 ± 0.57 | 11.99 ± 1.21 |
| OS | 5609.32 ± 566.21 | 5588.91 ± 224.36 | 5473.05 ± 202.84 | 5971.40 ± 282.62 | 5891.16 ± 380.86 | 5360.71 ± 271.28 | 5467.96 ± 567.58 |
| h/H | 3.52 ± 0.37 | 3.68 ± 0.24 | 3.47 ± 0.40 | 3.72 ± 0.12 | 3.96 ± 0.35 | 3.64 ± 0.40 | 4.15 ± 0.38 |
| PI | 114.60 ± 10.94 | 114.27 ± 4.57 | 111.97 ± 4.06 | 121.58 ± 5.34 | 119.96 ± 7.30 | 109.53 ± 5.38 | 111.70 ± 11.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maia, M.; Cavaco, A.R.; Laureano, G.; Cunha, J.; Eiras-Dias, J.; Matos, A.R.; Duarte, B.; Figueiredo, A. More than Just Wine: The Nutritional Benefits of Grapevine Leaves. Foods 2021, 10, 2251. https://doi.org/10.3390/foods10102251
Maia M, Cavaco AR, Laureano G, Cunha J, Eiras-Dias J, Matos AR, Duarte B, Figueiredo A. More than Just Wine: The Nutritional Benefits of Grapevine Leaves. Foods. 2021; 10(10):2251. https://doi.org/10.3390/foods10102251
Chicago/Turabian StyleMaia, Marisa, Ana Rita Cavaco, Gonçalo Laureano, Jorge Cunha, José Eiras-Dias, Ana Rita Matos, Bernardo Duarte, and Andreia Figueiredo. 2021. "More than Just Wine: The Nutritional Benefits of Grapevine Leaves" Foods 10, no. 10: 2251. https://doi.org/10.3390/foods10102251