The Effect of 5-Aminolevulinic Acid Photodynamic Therapy in Promoting Pyroptosis of HPV-Infected Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Preparation and Administration of MCC950
2.3. Cell Viability Assay
2.4. Quantitative Real-Time PCR (qRT-PCR)
2.5. Western Blot
2.6. Intracellular Generation of ROS
2.7. Lactate Dehydrogenase (LDH) Release Assay
2.8. Hoechst 33342/PI Staining
2.9. Statistical Analysis
3. Results
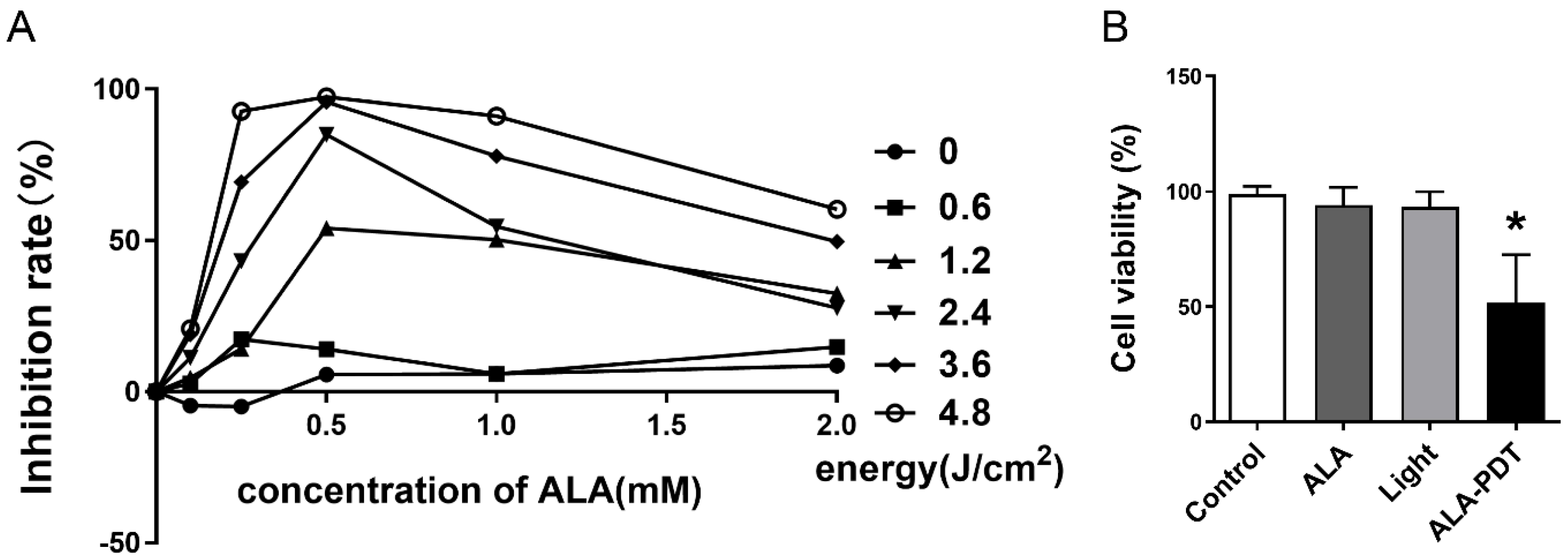
3.1. Different Gradients of ALA and Energy Suppress HeLa Cell Proliferation
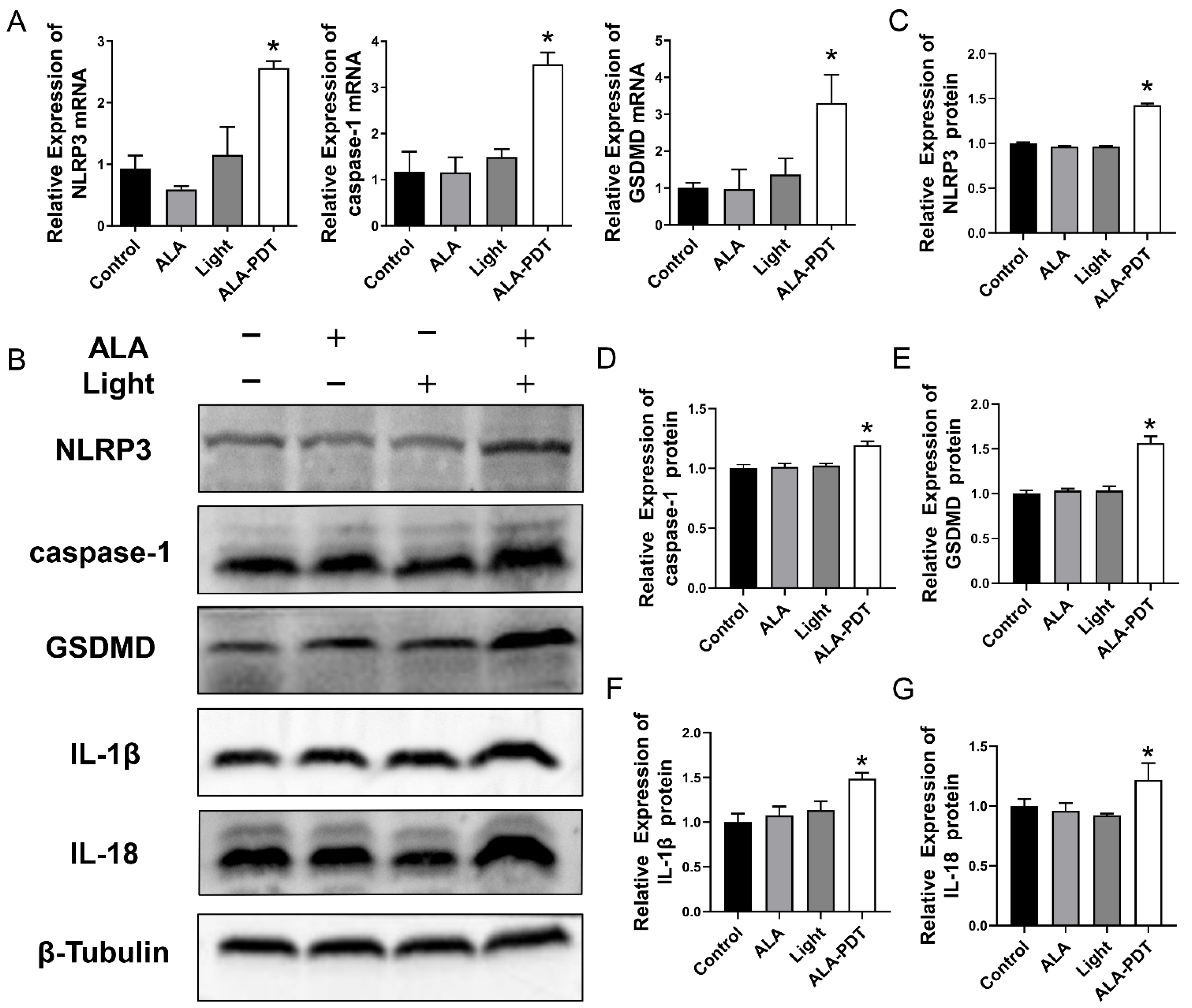
3.2. ALA-PDT Induces Pyroptosis in HeLa Cells
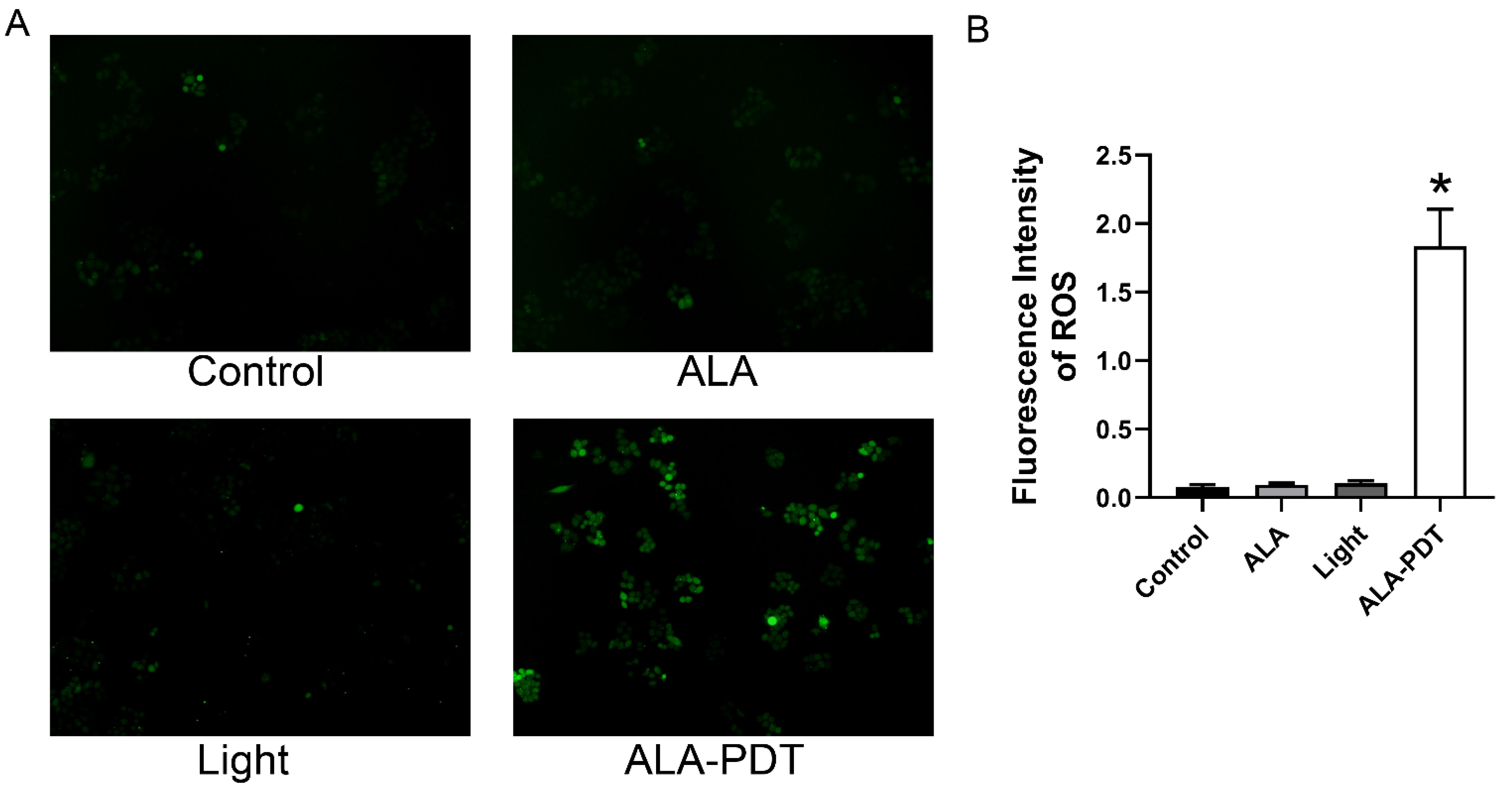
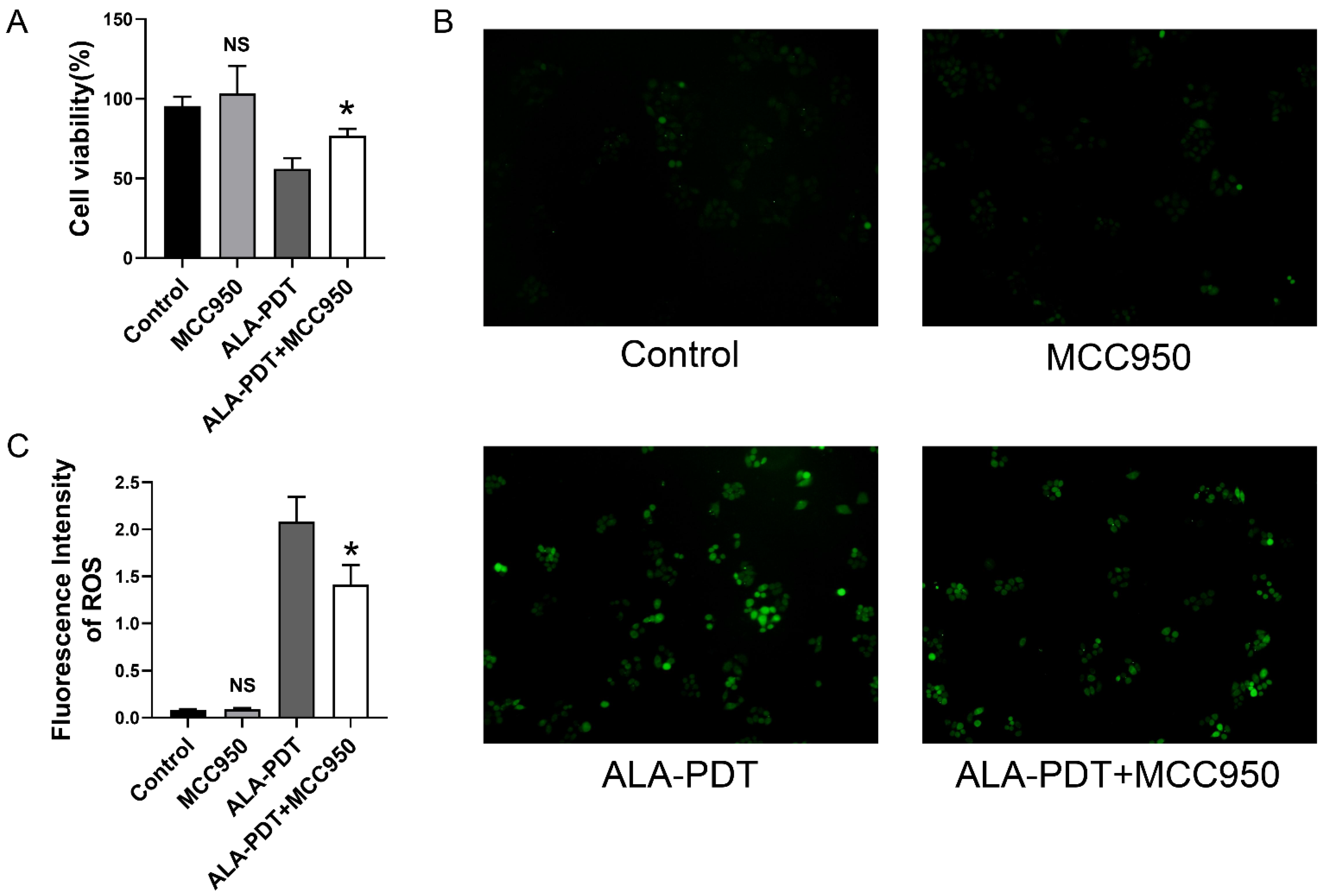
3.3. ALA-PDT Enhances the ROS Production
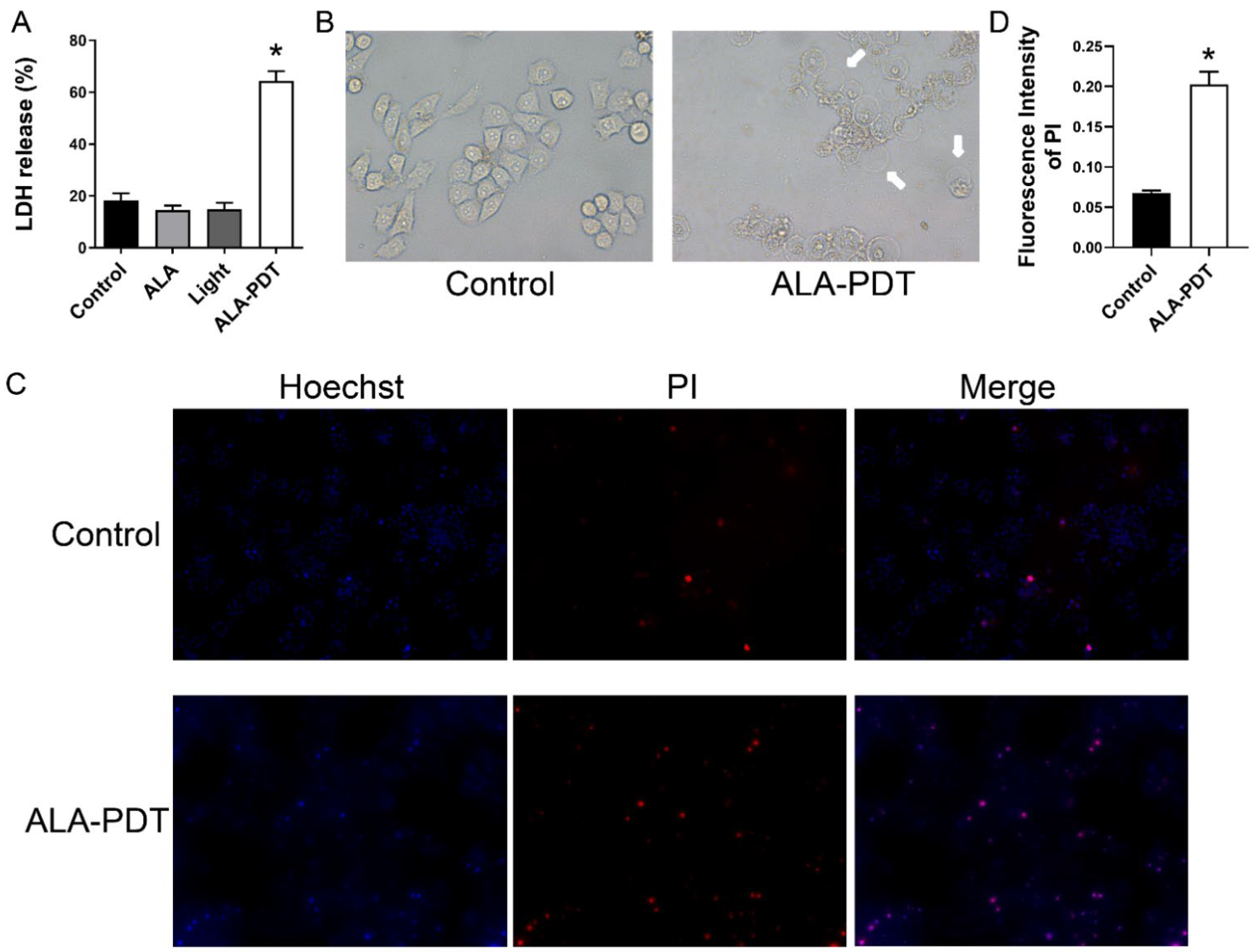
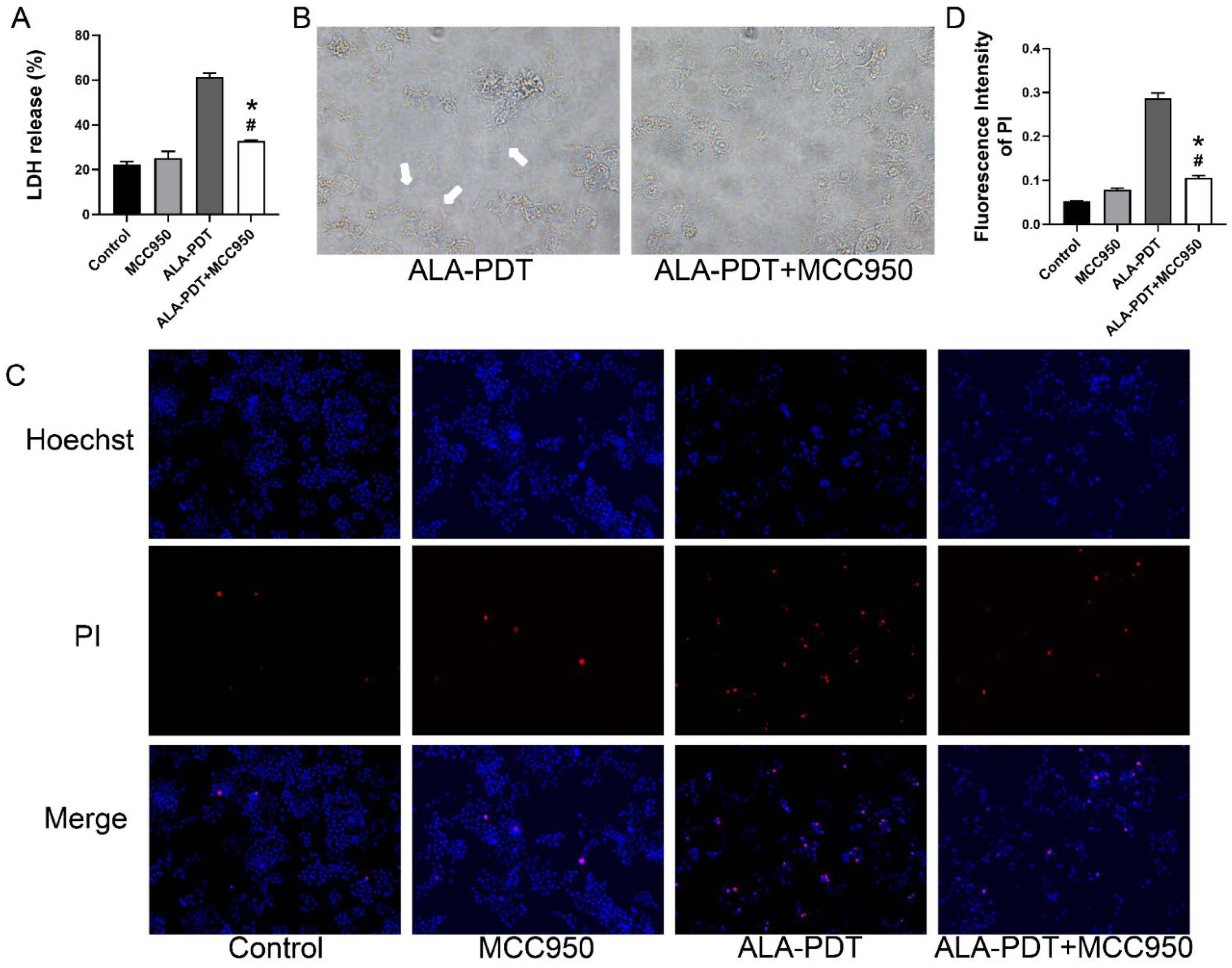
3.4. ALA-PDT Induces Destruction of Cell Membrane of HeLa Cells
3.5. MCC950 Alleviates the ALA-PDT-Induced Cell Death and ROS Production
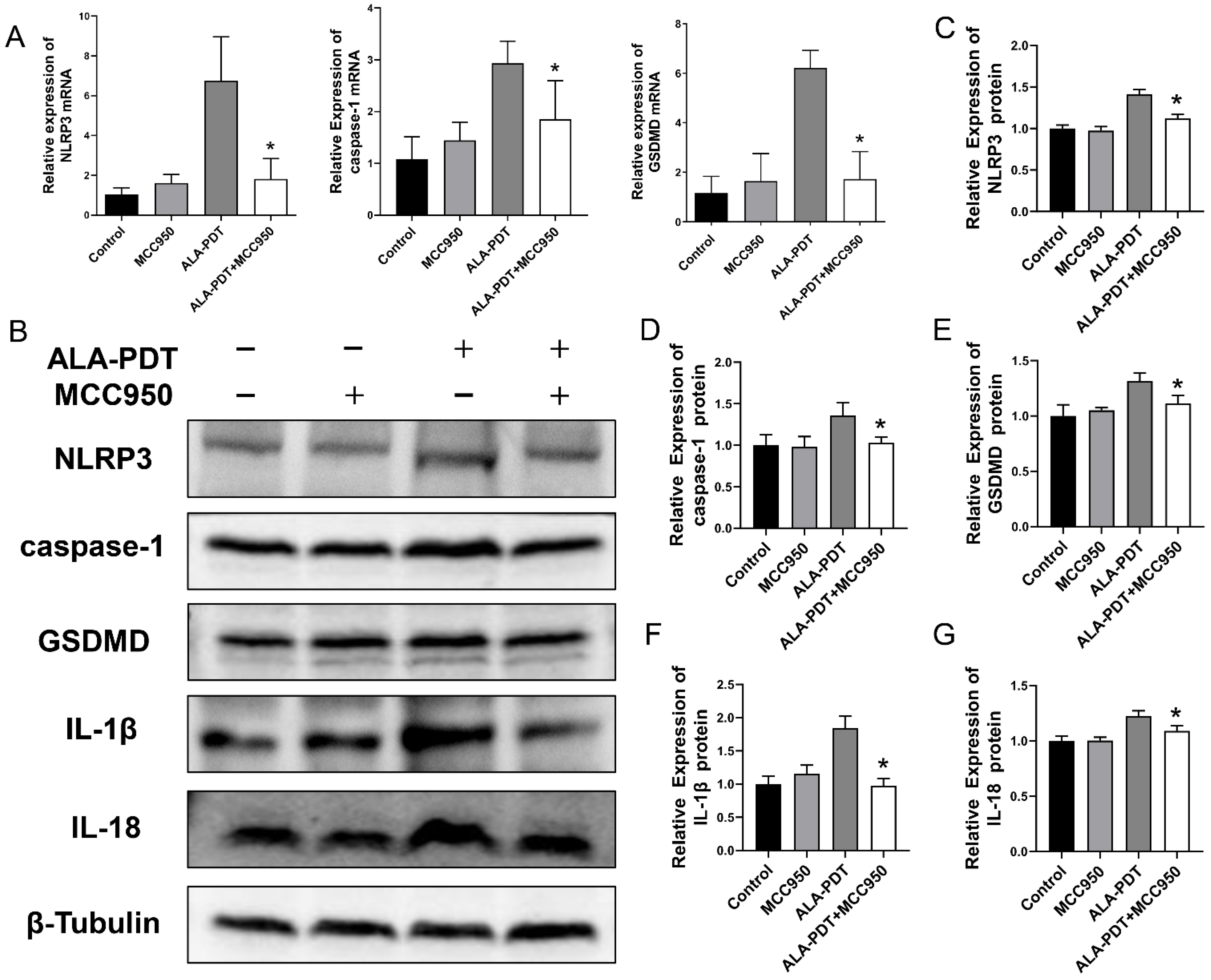
3.6. MCC950 Suppressed ALA-PDT-Induced Pyroptosis in HeLa Cells
3.7. MCC950 Abrogated the Pyroptotic Morphology in ALA-PDT-Treated Cells
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grennan, D. Genital warts. JAMA 2019, 321, 520. [Google Scholar] [CrossRef] [PubMed]
- Serrano, B.; Brotons, M.; Bosch, F.X.; Bruni, L. Epidemiology and burden of hpv-related disease. Best. Pract. Res. Clin. Obstet. Gynaecol. 2018, 47, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Ni, G.; Wang, T.; Mounsey, K.; Cavezza, S.; Pan, X.; Liu, X. Genital warts treatment: Beyond imiquimod. Hum. Vacc. Immunother. 2018, 14, 1815–1819. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Lei, S.; Pan, K.; Chen, T.; Lin, J.; Ni, G.; Liu, J.; Zeng, X.; Chen, Q.; Dan, H. Application of photodynamic therapy in immune-related diseases. Photodiagnosis Photodyn. Ther. 2021, 34, 102318. [Google Scholar] [CrossRef]
- Chilakamarthi, U.; Giribabu, L. Photodynamic therapy: Past, present and future. Chem. Rec. 2017, 17, 775–802. [Google Scholar] [CrossRef]
- van Straten, D.; Mashayekhi, V.; de Bruijn, H.S.; Oliveira, S.; Robinson, D.J. Oncologic photodynamic therapy: Basic principles, current clinical status and future directions. Cancers 2017, 9, 19. [Google Scholar] [CrossRef]
- Kwiatkowski, S.; Knap, B.; Przystupski, D.; Saczko, J.; Kędzierska, E.; Knap-Czop, K.; Kotlińska, J.; Michel, O.; Kotowski, K.; Kulbacka, J. Photodynamic therapy-mechanisms, photosensitizers and combinations. Biomed. Pharmacother. 2018, 106, 1098–1107. [Google Scholar] [CrossRef]
- Xie, J.; Wang, S.; Li, Z.; Ao, C.; Wang, J.; Wang, L.; Peng, X.; Zeng, K. 5-aminolevulinic acid photodynamic therapy reduces hpv viral load via autophagy and apoptosis by modulating ras/raf/mek/erk and pi3k/akt pathways in hela cells. J. Photochem. Photobiol. B 2019, 194, 46–55. [Google Scholar] [CrossRef]
- Li, Z.; Teng, M.; Wang, Y.; Wang, Q.; Feng, Y.; Xiao, Z.; Li, C.; Zeng, K. The mechanism of 5-aminolevulinic acid photodynamic therapy in promoting endoplasmic reticulum stress in the treatment of hr-hpv-infected hela cells. Photodermatol. Photoimmunol. Photomed. 2021, 37, 348–359. [Google Scholar] [CrossRef]
- Donohoe, C.; Senge, M.O.; Arnaut, L.G.; Gomes-Da-Silva, L.C. Cell death in photodynamic therapy: From oxidative stress to anti-tumor immunity. Biochim. Biophys. Acta Rev. Cancer 2019, 1872, 188308. [Google Scholar] [CrossRef]
- Shi, J.; Gao, W.; Shao, F. Pyroptosis: Gasdermin-mediated programmed necrotic cell death. Trends Biochem. Sci. 2017, 42, 245–254. [Google Scholar] [CrossRef]
- Kovacs, S.B.; Miao, E.A. Gasdermins: Effectors of pyroptosis. Trends Cell Biol. 2017, 27, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Lamkanfi, M.; Dixit, V.M. Mechanisms and functions of inflammasomes. Cell 2014, 157, 1013–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vande, W.L.; Lamkanfi, M. Pyroptosis. Curr. Biol. 2016, 26, R568–R572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Zhang, T.; Ma, X.; Yang, Q.C.; Yang, L.L.; Yang, S.C.; Liang, M.; Xu, Z.; Sun, Z.J. Microenvironment-responsive prodrug-induced pyroptosis boosts cancer immunotherapy. Adv. Sci. 2021, 8, e2101840. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.X.; Zhu, W.T.; Hu, J.H.; Yang, W.; Liu, P.; Liu, Q.H.; Bai, Y.X.; Xie, R. Curcumin-loaded poly(l-lactide-co-glycolide) microbubble-mediated sono-photodynamic therapy in liver cancer cells. Ultrasound. Med. Biol. 2020, 46, 2030–2043. [Google Scholar] [CrossRef]
- Li, L.; Song, D.; Qi, L.; Jiang, M.; Wu, Y.; Gan, J.; Cao, K.; Li, Y.; Bai, Y.; Zheng, T. Photodynamic therapy induces human esophageal carcinoma cell pyroptosis by targeting the pkm2/caspase-8/caspase-3/gsdme axis. Cancer Lett. 2021, 520, 143–159. [Google Scholar] [CrossRef]
- Wu, M.; Liu, X.; Chen, H.; Duan, Y.; Liu, J.; Pan, Y.; Liu, B. Activation of pyroptosis by membrane-anchoring aie photosensitizer design: New prospect for photodynamic cancer cell ablation. Angew. Chem. Int. Ed. Engl. 2021, 60, 9093–9098. [Google Scholar] [CrossRef]
- Broekgaarden, M.; Weijer, R.; van Gulik, T.M.; Hamblin, M.R.; Heger, M. Tumor cell survival pathways activated by photodynamic therapy: A molecular basis for pharmacological inhibition strategies. Cancer Metastasis Rev. 2015, 34, 643–690. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; He, Y.; Ming, H.; Lei, S.; Leng, Y.; Xia, Z.Y. Lipopolysaccharide (lps) aggravates high glucose- and hypoxia/reoxygenation-induced injury through activating ros-dependent nlrp3 inflammasome-mediated pyroptosis in h9c2 cardiomyocytes. J. Diabetes Res. 2019, 2019, 8151836. [Google Scholar] [CrossRef] [Green Version]
- Sun, B.; Cai, Y.; Li, Y.; Li, J.; Liu, K.; Li, Y.; Yang, Y. The nonstructural protein np1 of human bocavirus 1 induces cell cycle arrest and apoptosis in hela cells. Virology 2013, 440, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Wang, H.W.; Yuan, K.H.; Li, F.L.; Huang, Z. Combination of photodynamic therapy and immunomodulation for skin diseases--update of clinical aspects. Photochem. Photobiol. Sci. 2011, 10, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Zhang, X.; Ma, C.; Yu, N.; Wang, J.; Xia, L.; Ge, X.; Liu, M.; Duan, A. Clinical analysis of five methods used to treat condylomata acuminata. Dermatology 2013, 227, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Lu, X.N.; Tang, H.; Zhang, Z.; Fan, J.; Xu, J.H. Evaluation of photodynamic therapy using topical aminolevulinic acid hydrochloride in the treatment of condylomata acuminata: A comparative, randomized clinical trial. Photodermatol. Photoimmunol. Photomed. 2009, 25, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Buytaert, E.; Dewaele, M.; Agostinis, P. Molecular effectors of multiple cell death pathways initiated by photodynamic therapy. Biochim. Biophys. Acta 2007, 1776, 86–107. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.T.; Chien, L.T.; Lin, Y.H.; Chien, H.F.; Chen, C.T. 5-ala mediated photodynamic therapy induces autophagic cell death via amp-activated protein kinase. Mol. Cancer 2010, 9, 91. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.; Ma, X.; Song, Y.; Chen, Q.; Jiao, Q.; Zhou, L. Targeting regulated cell death in tumor nanomedicines. Theranostics 2022, 12, 817–841. [Google Scholar] [CrossRef]
- Orning, P.; Weng, D.; Starheim, K.; Ratner, D.; Best, Z.; Lee, B.; Brooks, A.; Xia, S.; Wu, H.; Kelliher, M.A.; et al. Pathogen blockade of tak1 triggers caspase-8-dependent cleavage of gasdermin d and cell death. Science 2018, 362, 1064–1069. [Google Scholar] [CrossRef] [Green Version]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef]
- Wallach, D.; Kang, T.B.; Dillon, C.P.; Green, D.R. Programmed necrosis in inflammation: Toward identification of the effector molecules. Science 2016, 352, f2154. [Google Scholar] [CrossRef]
- Zeng, C.; Wang, R.; Tan, H. Role of pyroptosis in cardiovascular diseases and its therapeutic implications. Int. J. Biol. Sci. 2019, 15, 1345–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin d causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, C.; Fernandes-Alnemri, T.; Mayes, L.; Alnemri, D.; Cingolani, G.; Alnemri, E.S. Cleavage of dfna5 by caspase-3 during apoptosis mediates progression to secondary necrotic/pyroptotic cell death. Nat. Commun. 2017, 8, 14128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gao, W.; Shi, X.; Ding, J.; Liu, W.; He, H.; Wang, K.; Shao, F. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 2017, 547, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yin, B.; Li, D.; Wang, G.; Han, X.; Sun, X. Gsdme mediates caspase-3-dependent pyroptosis in gastric cancer. Biochem. Biophys. Res. Commun. 2018, 495, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Elliott, E.I.; Sutterwala, F.S. Initiation and perpetuation of nlrp3 inflammasome activation and assembly. Immunol. Rev. 2015, 265, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Bai, H.; Ma, X.; Shen, M.; Li, R.; Qiu, D.; Li, S.; Gao, L. Blockade of the nlrp3/caspase-1 axis attenuates ketamine-induced hippocampus pyroptosis and cognitive impairment in neonatal rats. J Neuroinflammation 2021, 18, 239. [Google Scholar] [CrossRef]
- Moossavi, M.; Parsamanesh, N.; Bahrami, A.; Atkin, S.L.; Sahebkar, A. Role of the nlrp3 inflammasome in cancer. Mol. Cancer 2018, 17, 158. [Google Scholar] [CrossRef] [Green Version]
- Shao, B.Z.; Xu, Z.Q.; Han, B.Z.; Su, D.F.; Liu, C. Nlrp3 inflammasome and its inhibitors: A review. Front. Pharmacol. 2015, 6, 262. [Google Scholar] [CrossRef] [Green Version]
- Coll, R.C.; Robertson, A.A.; Chae, J.J.; Higgins, S.C.; Muñoz-Planillo, R.; Inserra, M.C.; Vetter, I.; Dungan, L.S.; Monks, B.G.; Stutz, A.; et al. A small-molecule inhibitor of the nlrp3 inflammasome for the treatment of inflammatory diseases. Nat. Med. 2015, 21, 248–255. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Yang, L.; Li, L.; Fan, Z. Nahs alleviated cell apoptosis and mitochondrial dysfunction in remote lung tissue after renal ischemia and reperfusion via nrf2 activation-mediated nlrp3 pathway inhibition. Biomed. Res. Int. 2021, 2021, 5598869. [Google Scholar] [CrossRef] [PubMed]
- Ni, B.; Pei, W.; Qu, Y.; Zhang, R.; Chu, X.; Wang, Y.; Huang, X.; You, H. Mcc950, the nlrp3 inhibitor, protects against cartilage degradation in a mouse model of osteoarthritis. Oxid. Med. Cell. Longev. 2021, 2021, 4139048. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, J.; Peng, X.; Wang, S.; Xu, M.; Liu, H.; Zhong, Y.; Chen, X.; Wang, Q.; Huang, X.; Zeng, K. The Effect of 5-Aminolevulinic Acid Photodynamic Therapy in Promoting Pyroptosis of HPV-Infected Cells. Photonics 2022, 9, 408. https://doi.org/10.3390/photonics9060408
Wei J, Peng X, Wang S, Xu M, Liu H, Zhong Y, Chen X, Wang Q, Huang X, Zeng K. The Effect of 5-Aminolevulinic Acid Photodynamic Therapy in Promoting Pyroptosis of HPV-Infected Cells. Photonics. 2022; 9(6):408. https://doi.org/10.3390/photonics9060408
Chicago/Turabian StyleWei, Junxiao, Xiaoming Peng, Sijia Wang, Meinian Xu, Hui Liu, Yixiu Zhong, Xi Chen, Qi Wang, Xiaowen Huang, and Kang Zeng. 2022. "The Effect of 5-Aminolevulinic Acid Photodynamic Therapy in Promoting Pyroptosis of HPV-Infected Cells" Photonics 9, no. 6: 408. https://doi.org/10.3390/photonics9060408