Application of Ultrashort Lasers in Developmental Biology: A Review


Abstract
:1. Introduction
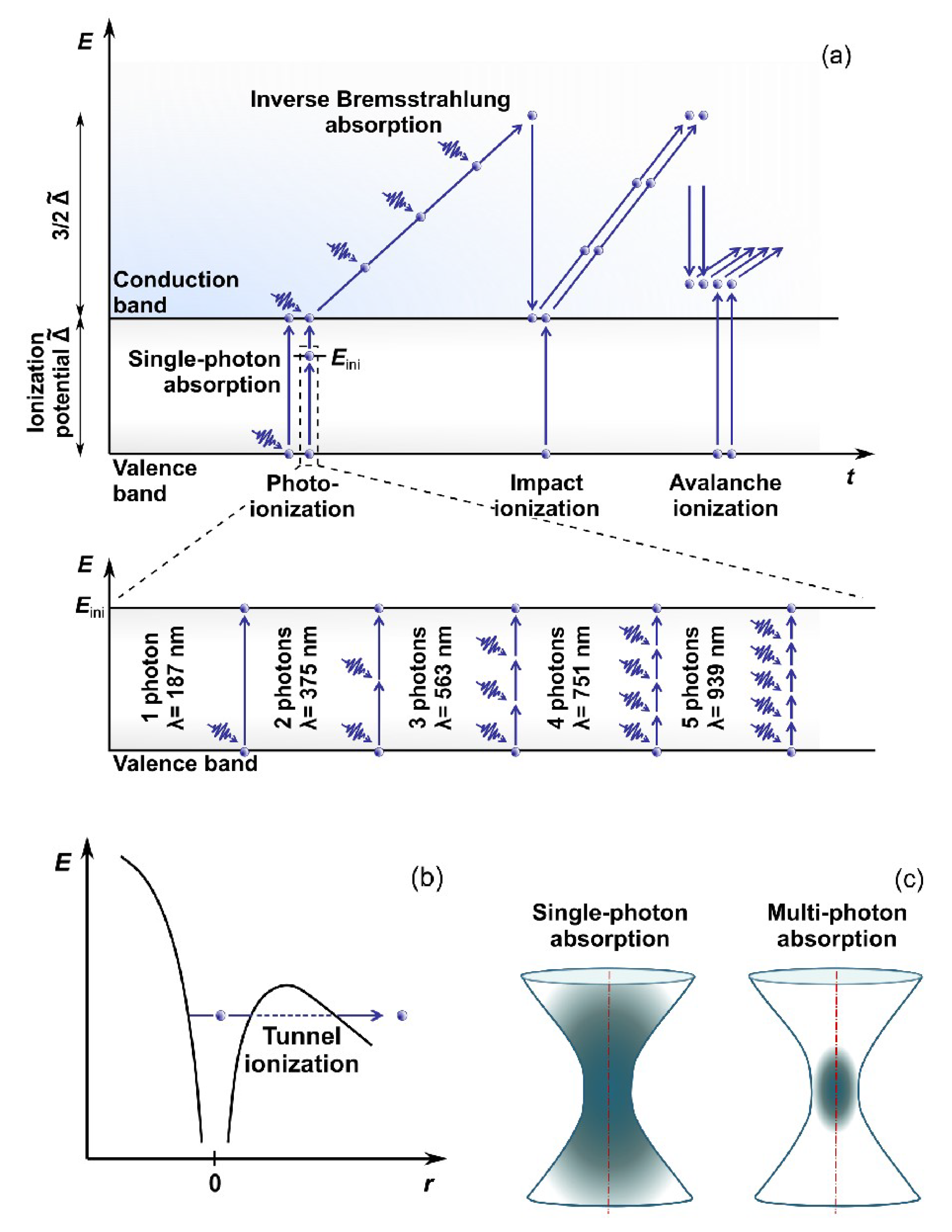
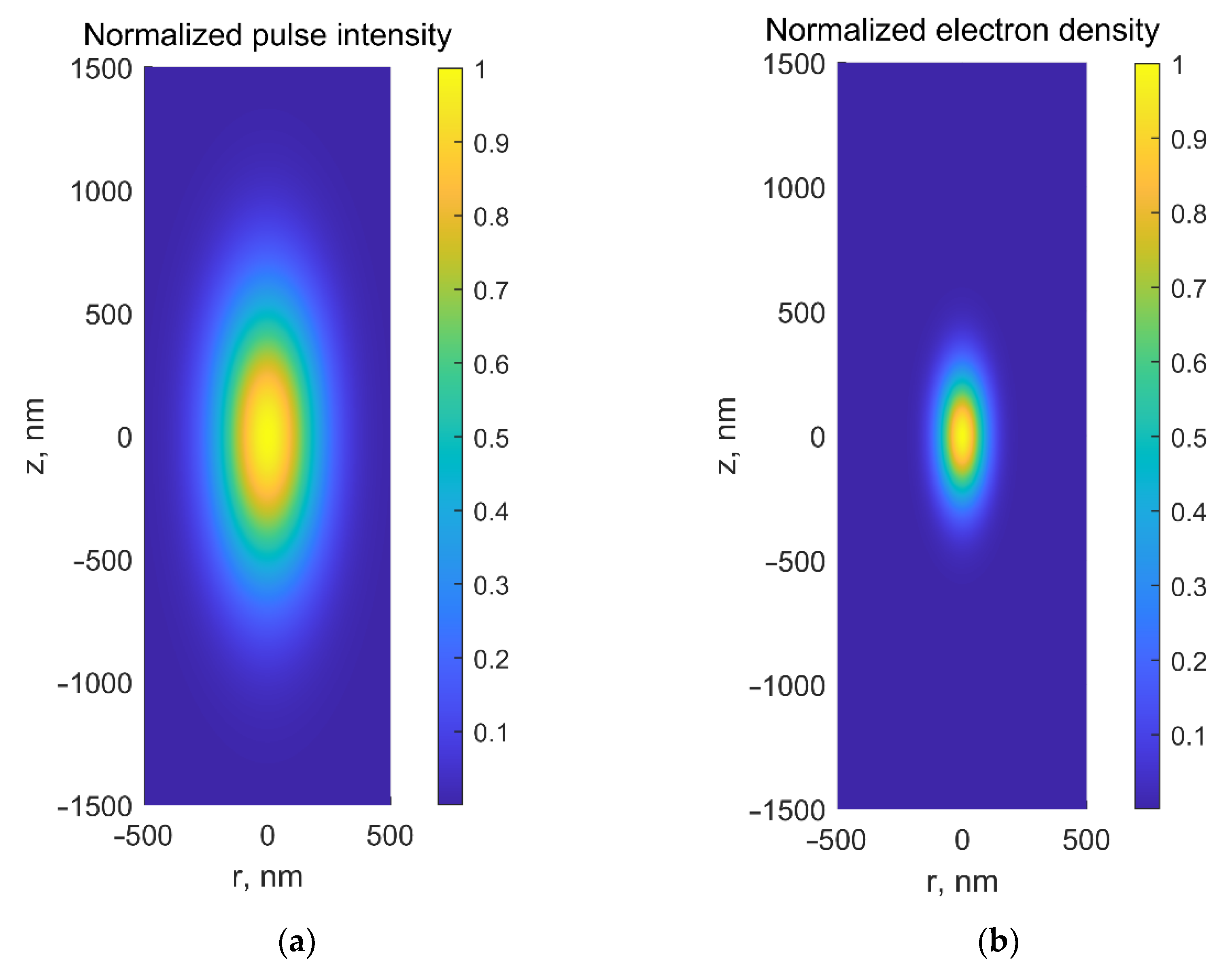
2. Interaction of Ultrashort Laser Pulses with Matter
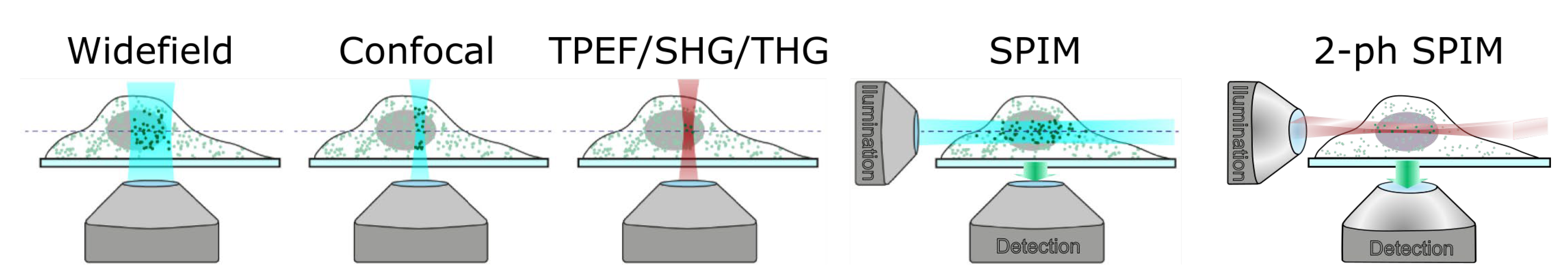
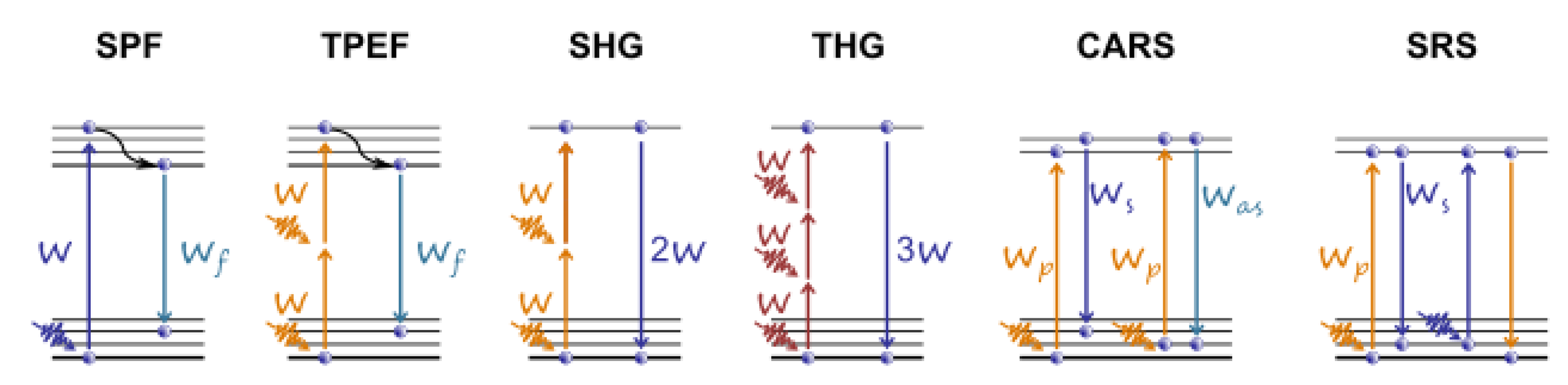
3. Application of Ultrashort Laser Pulses for Nonlinear Microscopy of Embryos
4. Oocyte/Blastomere Enucleation and Embryonic Cell Fusion by Application of Ultrashort Laser Pulses
5. Optoporation and Laser Transfection of Developing Embryos
6. Ultrashort Laser Microsurgery of Externally Developing Embryos
7. Ultrashort Laser-Based Microsurgery and Microscopy of Preimplantation Embryos in Assisted Reproductive Technologies: A Prospects
8. Safety Aspects for Use of Ultrashort Laser Pulses
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Litvinova, K.S.; Rafailov, I.E.; Dunaev, A.V.; Sokolovski, S.G.; Rafailov, E.U. Non-invasive biomedical research and diagnostics enabled by innovative compact lasers. Prog. Quantum Electron. 2017, 56, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A. Laser Techniques in Ophthalmology: A Guide to YAG and Photothermal Laser Treatments in Clinic; CRC Press: Boca Raton, FL, USA, 2022; ISBN 1000538486. [Google Scholar]
- Gianfaldoni, S.; Tchernev, G.; Wollina, U.; Fioranelli, M.; Roccia, M.G.; Gianfaldoni, R.; Lotti, T. An Overview of Laser in Dermatology: The Past, the Present and … the Future (?). Open Access Maced. J. Med. Sci. 2017, 5, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalkhal, E.; Rezaei-Tavirani, M.; Zali, M.R.; Akbari, Z. The Evaluation of Laser Application in Surgery: A Review Article. J. Lasers Med. Sci. 2019, 10, S104–S111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, S.H.; Mazur, E. Surgical applications of femtosecond lasers. J. Biophotonics 2009, 2, 557–572. [Google Scholar] [CrossRef]
- Lehmann, U.; Kreipe, H. Laser-Assisted Microdissection and Isolation of DNA and RNA. In Breast Cancer Research Protocols; Brooks, S.A., Harris, A., Eds.; Humana Press: Totowa, NJ, USA, 2006; pp. 65–75. ISBN 978-1-59259-969-1. [Google Scholar]
- Han, T.-S.; Oshima, M. Laser Microdissection of Cellular Compartments for Expression Analyses in Cancer Models. In Inflammation and Cancer: Methods and Protocols; Jenkins, B.J., Ed.; Springer: New York, NY, USA, 2018; pp. 143–153. ISBN 978-1-4939-7568-6. [Google Scholar]
- Odde, D.J.; Renn, M.J. Laser-guided direct writing for applications in biotechnology. Trends Biotechnol. 1999, 17, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Duan, B. State-of-the-Art Review of 3D Bioprinting for Cardiovascular Tissue Engineering. Ann. Biomed. Eng. 2017, 45, 195–209. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Yue, K.; Aleman, J.; Mollazadeh-Moghaddam, K.; Bakht, S.M.; Yang, J.; Jia, W.; Dell’Erba, V.; Assawes, P.; Shin, S.R.; et al. 3D Bioprinting for Tissue and Organ Fabrication. Ann. Biomed. Eng. 2017, 45, 148–163. [Google Scholar] [CrossRef] [Green Version]
- Berns, M.W.; Aist, J.; Edwards, J.; Strahs, K.; Girton, J.; Mcneill, P.; Rattner, J.B.; Kitzes, M.; Liaw, L.; Siemens, A.; et al. Laser microsurgery in cell and developmental Biology. Science 1981, 213, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Thalhammer, S.; Lahr, G.; Clement-Sengewald, A.; Heckl, W.M.; Burgemeister, R.; Schütze, K. Laser microtools in cell biology and molecular medicine. Laser Phys. 2003, 13, 681–691. [Google Scholar]
- Kohli, V.; Elezzabi, A.Y. Prospects and developments in cell and embryo laser nanosurgery. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2009, 1, 11–25. [Google Scholar] [CrossRef]
- Docchio, F.; Sacchi, C.; Marshall, J. Experimental Investigation of Optical Breakdown Thresholds in Ocular Media under Single Pulse Irradiation with Different Pulse Durations. Lasers Ophthalmol. 1986, 1, 83–93. [Google Scholar]
- Tadir, Y.; Douglas-Hamilton, D.H.; Douglas-Hamilton, D.H.; Douglas-Hamilton, D.H.; Douglas-Hamilton, D.H. Laser Effects in the Manipulation of Human Eggs and Embryos for In Vitro Fertilization. Methods Cell Biol. 2007, 82, 409–431. [Google Scholar] [CrossRef]
- Sacchi, C.A. Laser-induced electric breakdown in water. J. Opt. Soc. Am. B 1991, 8, 337. [Google Scholar] [CrossRef]
- Sander, M.U.; Gudiksen, M.S.; Luther, K.; Troe, J. Liquid water ionization: Mechanistic implications of the H/D isotope effect in the geminate recombination of hydrated electrons. Chem. Phys. 2000, 258, 257–265. [Google Scholar] [CrossRef]
- Elles, C.G.; Jailaubekov, A.E.; Crowell, R.A.; Bradforth, S.E. Excitation-energy dependence of the mechanism for two-photon ionization of liquid H2O and D2O from 8.3to12.4eV. J. Chem. Phys. 2006, 125, 044515. [Google Scholar] [CrossRef] [Green Version]
- Göppert-Mayer, M. Über Elementarakte mit zwei Quantensprüngen. Ann. Phys. 1931, 401, 273–294. [Google Scholar] [CrossRef]
- Keldysh, L.V. Ionization in the field of a strong electromagnetic wave. Sov. Phys. JETP 1965, 20, 1307–1314. [Google Scholar]
- Liang, X.-X.; Zhang, Z.; Vogel, A. Multi-rate-equation modeling of the energy spectrum of laser-induced conduction band electrons in water. Opt. Express 2019, 27, 4672. [Google Scholar] [CrossRef]
- Ilina, I.; Sitnikov, D. From Zygote to Blastocyst: Application of Ultrashort Lasers in the Field of Assisted Reproduction and Developmental Biology. Diagnostics 2021, 11, 1897. [Google Scholar] [CrossRef]
- Vogel, A.; Noack, J.; Hüttman, G.; Paltauf, G. Mechanisms of femtosecond laser nanosurgery of cells and tissues. Appl. Phys. B Lasers Opt. 2005, 81, 1015–1047. [Google Scholar] [CrossRef]
- Linz, N.; Freidank, S.; Liang, X.X.; Vogel, A. Wavelength dependence of femtosecond laser-induced breakdown in water and implications for laser surgery. Phys. Rev. B 2016, 94, 024113. [Google Scholar] [CrossRef] [Green Version]
- Vogel, A.; Venugopalan, V. Mechanisms of Pulsed Laser Ablation of Biological Tissues. Chem. Rev. 2003, 103, 577–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett, B.C.; Dixon, D.A.; Camaioni, D.M.; Chipman, D.M.; Johnson, M.A.; Jonah, C.D.; Kimmel, G.A.; Miller, J.H.; Rescigno, T.N.; Rossky, P.J.; et al. Role of Water in Electron-Initiated Processes and Radical Chemistry: Issues and Scientific Advances. Chem. Rev. 2005, 105, 355–390. [Google Scholar] [CrossRef] [PubMed]
- Nikogosyan, D.N.; Oraevsky, A.A.; Rupasov, V.I. Two-photon ionization and dissociation of liquid water by powerful laser UV radiation. Chem. Phys. 1983, 77, 131–143. [Google Scholar] [CrossRef]
- Boudaiffa, B. Resonant Formation of DNA Strand Breaks by Low-Energy (3 to 20 eV) Electrons. Science 2000, 287, 1658–1660. [Google Scholar] [CrossRef]
- Zalessky, A.; Fedotov, Y.; Yashkina, E.; Nadtochenko, V.; Osipov, A.N. Immunocytochemical localization of XRCC1 and γH2AX foci induced by tightly focused femtosecond laser radiation in cultured human cells. Molecules 2021, 26, 4027. [Google Scholar] [CrossRef]
- Quinto-Su, P.A.; Venugopalan, V. Mechanisms of Laser Cellular Microsurgery. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2007; Volume 82, pp. 111–151. ISBN 0123706483. [Google Scholar]
- Paltauf, G.; Schmidt-Kloiber, H. Microcavity dynamics during laser-induced spallation of liquids and gels. Appl. Phys. A Mater. Sci. Process. 1996, 62, 303–311. [Google Scholar] [CrossRef]
- Berns, M.W.; Cheng, W.K.; Floyd, A.D.; Ohnuki, Y. Chromosome Lesions Produced with an Argon Laser Microbeam without Dye Sensitization. Science 1971, 171, 903–905. [Google Scholar] [CrossRef] [Green Version]
- König, K.; Riemann, I.; Fischer, P.; Halbhuber, K.J. Intracellular nanosurgery with near infrared femtosecond laser pulses. Cell. Mol. Biol. 1999, 45, 195–201. [Google Scholar]
- König, K.; Riemann, I.; Fritzsche, W. Nanodissection of human chromosomes with near-infrared femtosecond laser pulses. Opt. Lett. 2001, 26, 819–821. [Google Scholar] [CrossRef]
- Daniel, J.C. Cleavage of Mammalian Ova inhibited by Visible Light. Nature 1964, 201, 316–317. [Google Scholar] [CrossRef] [PubMed]
- Hirao, Y.; Yanagimachi, R. Detrimental effect of visible light on meiosis of mammalian eggs in vitro. J. Exp. Zool. 1978, 206, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Hegele-Hartung, C.; Schumacher, A.; Fischer, B. Effects of visible light and room temperature on the ultrastructure of preimplantation rabbit embryos: A time course study. Anat. Embryol. 1991, 183, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Handbook of Biological Confocal Microscopy; Pawley, J.B. (Ed.) Springer: Boston, MA, USA, 1995; ISBN 978-1-4757-5350-9. [Google Scholar]
- Tosheva, K.L.; Yuan, Y.; Pereira, P.M.; Culley, S.; Henriques, R. Between life and death: Reducing phototoxicity in Super-Resolution Microscopy. Preprints 2019. [Google Scholar] [CrossRef]
- Denk, W.; Strickler, J.; Webb, W. Two-photon laser scanning fluorescence microscopy. Science 1990, 248, 73–76. [Google Scholar] [CrossRef] [Green Version]
- Barad, Y.; Eisenberg, H.; Horowitz, M.; Silberberg, Y. Nonlinear scanning laser microscopy by third harmonic generation. Appl. Phys. Lett. 1997, 70, 922–924. [Google Scholar] [CrossRef] [Green Version]
- Campagnola, P.J.; Clark, H.A.; Mohler, W.A.; Lewis, A.; Loew, L.M. Second-harmonic imaging microscopy of living cells. J. Biomed. Opt. 2001, 6, 277. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.L.; Xie, X.S. Coherent Anti-Stokes Raman Scattering Microscopy: Chemical Imaging for Biology and Medicine. Annu. Rev. Anal. Chem. 2008, 1, 883–909. [Google Scholar] [CrossRef] [Green Version]
- Parodi, V.; Jacchetti, E.; Osellame, R.; Cerullo, G.; Polli, D.; Raimondi, M.T. Nonlinear Optical Microscopy: From Fundamentals to Applications in Live Bioimaging. Front. Bioeng. Biotechnol. 2020, 8, 585363. [Google Scholar] [CrossRef]
- Boyd, R. Nonlinear Optics, 1st ed.; Academic Press: New York, NY, USA, 1992; ISBN 9781483288239. [Google Scholar]
- Yelin, D.; Silberberg, Y. Laser scanning third-harmonic-generation microscopy in biology. Opt. Express 1999, 5, 169. [Google Scholar] [CrossRef]
- Rehberg, M.; Krombach, F.; Pohl, U.; Dietzel, S. Label-Free 3D Visualization of Cellular and Tissue Structures in Intact Muscle with Second and Third Harmonic Generation Microscopy. PLoS ONE 2011, 6, e28237. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.K. Higher harmonic generation microscopy. Adv. Biochem. Eng. Biotechnol. 2005, 95, 17–56. [Google Scholar] [CrossRef]
- Nandakumar, P.; Kovalev, A.; Volkmer, A. Vibrational imaging based on stimulated Raman scattering microscopy. New J. Phys. 2009, 11, 033026. [Google Scholar] [CrossRef]
- Cheng, J.-X.; Xie, X.S. Coherent Anti-Stokes Raman Scattering Microscopy: Instrumentation, Theory, and Applications. J. Phys. Chem. B 2004, 108, 827–840. [Google Scholar] [CrossRef]
- Wang, S.; Larina, I.V.; Larin, K.V. Label-free optical imaging in developmental biology [Invited]. Biomed. Opt. Express 2020, 11, 2017. [Google Scholar] [CrossRef]
- Truong, T.V.; Supatto, W.; Koos, D.S.; Choi, J.M.; Fraser, S.E. Deep and fast live imaging with two-photon scanned light-sheet microscopy. Nat. Methods 2011, 8, 757–762. [Google Scholar] [CrossRef] [Green Version]
- Lavagnino, Z.; Cella Zanacchi, F.; Ronzitti, E.; Diaspro, A. Two-photon excitation selective plane illumination microscopy (2PE-SPIM) of highly scattering samples: Characterization and application. Opt. Express 2013, 21, 5998. [Google Scholar] [CrossRef]
- Wu, Q.; Kumar, N.; Velagala, V.; Zartman, J.J. Tools to reverse-engineer multicellular systems: Case studies using the fruit fly. J. Biol. Eng. 2019, 13, 33. [Google Scholar] [CrossRef]
- Garcia, M.D.; Udan, R.S.; Hadjantonakis, A.-K.; Dickinson, M.E. Preparation of Postimplantation Mouse Embryos for Imaging: Figure 1. Cold Spring Harb. Protoc. 2011, 2011, pdb.prot5594. [Google Scholar] [CrossRef]
- Lopez, A.L.; Larina, I.V. Second harmonic generation microscopy of early embryonic mouse hearts. Biomed. Opt. Express 2019, 10, 2898. [Google Scholar] [CrossRef]
- Abu-Siniyeh, A.; Al-Zyoud, W. Highlights on selected microscopy techniques to study zebrafish developmental biology. Lab. Anim. Res. 2020, 36, 12. [Google Scholar] [CrossRef] [PubMed]
- Eum, J.; Kwak, J.; Kim, H.; Ki, S.; Lee, K.; Raslan, A.; Park, O.; Chowdhury, M.; Her, S.; Kee, Y.; et al. 3D Visualization of Developmental Toxicity of 2,4,6-Trinitrotoluene in Zebrafish Embryogenesis Using Light-Sheet Microscopy. Int. J. Mol. Sci. 2016, 17, 1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.-Y.; Hsieh, C.-S.; Chu, S.-W.; Lin, C.-Y.; Ko, C.-Y.; Chen, Y.-C.; Tsai, H.-J.; Hu, C.-H.; Sun, C.-K. Noninvasive harmonics optical microscopy for long-term observation of embryonic nervous system development in vivo. J. Biomed. Opt. 2006, 11, 054022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.-Y.; Hovhannisyan, V.; Wu, J.-T.; Lin, C.-W.; Chen, J.-H.; Lin, S.-J.; Dong, C.-Y. Label-free imaging of Drosophila larva by multiphoton autofluorescence and second harmonic generation microscopy. J. Biomed. Opt. 2008, 13, 050502. [Google Scholar] [CrossRef] [PubMed]
- Débarre, D.; Supatto, W.; Pena, A.-M.; Fabre, A.; Tordjmann, T.; Combettes, L.; Schanne-Klein, M.-C.; Beaurepaire, E. Imaging lipid bodies in cells and tissues using third-harmonic generation microscopy. Nat. Methods 2006, 3, 47–53. [Google Scholar] [CrossRef]
- Greenhalgh, C.; Stewart, B.; Cisek, R.; Prent, N.; Major, A.; Barzda, V. Dynamic investigation of Drosophila myocytes with second harmonic generation microscopy. In Photonics North 2006; Mathieu, P., Ed.; SPIE: Bellingham, WA, USA, 2006; Volume 6343, p. 634308. [Google Scholar]
- Greenhalgh, C.; Prent, N.; Green, C.; Cisek, R.; Major, A.; Stewart, B.; Barzda, V. Influence of semicrystalline order on the second-harmonic generation efficiency in the anisotropic bands of myocytes. Appl. Opt. 2007, 46, 1852. [Google Scholar] [CrossRef] [Green Version]
- Débarre, D.; Olivier, N.; Supatto, W.; Beaurepaire, E. Mitigating phototoxicity during multiphoton microscopy of live drosophila embryos in the 1.0–1.2 μm wavelength range. PLoS ONE 2014, 9, e104250. [Google Scholar] [CrossRef] [Green Version]
- Dou, W.; Zhang, D.; Jung, Y.; Cheng, J.X.; Umulis, D.M. Label-free imaging of lipid-droplet intracellular motion in early Drosophila embryos using femtosecond-stimulated Raman loss microscopy. Biophys. J. 2012, 102, 1666–1675. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.; Witvliet, D.; Hernandez-Nunez, L.; Linderman, S.W.; Samuel, A.D.T.; Venkatachalam, V. Imaging whole-brain activity to understand behaviour. Nat. Rev. Phys. 2022, 4, 292–305. [Google Scholar] [CrossRef]
- Memeo, R.; Paiè, P.; Sala, F.; Castriotta, M.; Guercio, C.; Vaccari, T.; Osellame, R.; Bassi, A.; Bragheri, F. Automatic imaging of Drosophila embryos with light sheet fluorescence microscopy on chip. J. Biophotonics 2021, 14, e202000396. [Google Scholar] [CrossRef]
- Karunendiran, A.; Cisek, R.; Tokarz, D.; Barzda, V.; Stewart, B.A. Examination of Drosophila eye development with third harmonic generation microscopy. Biomed. Opt. Express 2017, 8, 4504. [Google Scholar] [CrossRef] [Green Version]
- Karunendiran, A.; Mirsanaye, K.; Stewart, B.A.; Barzda, V. Second Harmonic Generation Properties in Chiral Sarcomeres of Drosophila Larval Muscles. Front. Phys. 2022, 10, 758709. [Google Scholar] [CrossRef]
- Golaraei, A.; Kontenis, L.; Karunendiran, A.; Stewart, B.A.; Barzda, V. Dual- and single-shot susceptibility ratio measurements with circular polarizations in second-harmonic generation microscopy. J. Biophotonics 2020, 13, e201960167. [Google Scholar] [CrossRef]
- Karmenyan, A.V.; Shakhbazyan, A.K.; Sviridova-Chailakhyan, T.A.; Krivokharchenko, A.S.; Chiou, A.E.; Chailakhyan, L.M. Use of picosecond infrared laser for micromanipulation of early mammalian embryos. Mol. Reprod. Dev. 2009, 76, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Kuetemeyer, K.; Lucas-Hahn, A.; Petersen, B.; Lemme, E.; Hassel, P.; Niemann, H.; Heisterkamp, A. Combined multiphoton imaging and automated functional enucleation of porcine oocytes using femtosecond laser pulses. J. Biomed. Opt. 2010, 15, 046006. [Google Scholar] [CrossRef] [PubMed]
- Kuetemeyer, K.; Lucas-Hahn, A.; Petersen, B.; Niemann, H.; Heisterkamp, A. Femtosecond laser-induced fusion of nonadherent cells and two-cell porcine embryos. J. Biomed. Opt. 2011, 16, 088001. [Google Scholar] [CrossRef] [Green Version]
- Ilina, I.V.; Ovchinnikov, A.V.; Sitnikov, D.S.; Rakityanskiy, M.M.; Agranat, M.B.; Khramova, Y.V.; Semenova, M.L. Application of femtosecond laser pulses in biomedical cell technologies. High Temp. 2013, 51, 173–178. [Google Scholar] [CrossRef]
- Ilina, I.V.; Chefonov, O.V.; Agranat, M.B.; Khramova, Y.V.; Ovchinnikov, A.V.; Sitnikov, D.S.; Semenova, M.L. Microsurgery of Cell Membrane with Femtosecond Laser Pulses for Cell Fusion and Optical Injection. ALT Proc. 2012, 1. [Google Scholar] [CrossRef]
- Osychenko, A.A.; Zalessky, A.D.; Kostrov, A.N.; Ryabova, A.V.; Krivokharchenko, A.S.; Nadtochenko, V.A. Femtosecond laser surgery of two-cell mouse embryos: Effect on viability, development, and tetraploidization. J. Biomed. Opt. 2017, 22, 1. [Google Scholar] [CrossRef] [Green Version]
- Krivokharchenko, A.; Karmenyan, A.; Sarkisov, O.; Bader, M.; Chiou, A.; Shakhbazyan, A. Laser Fusion of Mouse Embryonic Cells and Intra-Embryonic Fusion of Blastomeres without Affecting the Embryo Integrity. PLoS ONE 2012, 7, e50029. [Google Scholar] [CrossRef] [Green Version]
- Osychenko, A.; Zalessky, A.; Astafiev, A.; Shakhov, A.; Kostrov, A.; Krivokharchenko, A.; Nadtochenko, V. Femtosecond laser-induced blastomere fusion results in embryo tetraploidy by common metaphase plate formation. Exp. Cell Res. 2020, 389, 111887. [Google Scholar] [CrossRef]
- Gouveia, C.; Huyser, C.; Egli, D.; Pepper, M.S. Lessons learned from somatic cell nuclear transfer. Int. J. Mol. Sci. 2020, 21, 2314. [Google Scholar] [CrossRef] [PubMed]
- Osychenko, A.A.; Zalessky, A.D.; Tochilo, U.A.; Martirosyan, D.Y.; Silaeva, Y.Y.; Nadtochenko, V.A. Femtosecond laser oocyte enucleation as a low-invasive and effective method of recipient cytoplast preparation. Biomed. Opt. Express 2022, 13, 1447. [Google Scholar] [CrossRef] [PubMed]
- Dominko, T.; Chan, A.; Simerly, C.; Luetjens, C.M.; Hewitson, L.; Martinovich, C.; Schatten, G. Dynamic imaging of the metaphase II spindle and maternal chromosomesin bovine oocytes: Implications for enucleation efficiency verification, avoidanceof parthenogenesis, and successful embryogenesis. Biol. Reprod. 2000, 62, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zhao, X.; Xing, Q.; Li, F.; Li, H.; Li, Y.; Chai, L.; Wang, Q.; Zheltikov, A. Femtosecond laser-induced cell fusion. Appl. Phys. Lett. 2008, 92, 093901. [Google Scholar] [CrossRef]
- Katchinskiy, N.; Godbout, R.; Goez, H.R.; Elezzabi, A.Y. Hemifusion of cells using femtosecond laser pulses. In Optical Interactions with Tissue and Cells XXVI; SPIE: Bellingham, WA, USA, 2015; Volume 9321, p. 932105. [Google Scholar] [CrossRef]
- Kohli, V.; Elezzabi, A.Y. Part I. Embryonic surgery using femtosecond laser pulses for the delivery of exogenous materials and the analysis of gene expression. In Optical Interactions with Tissue and Cells XIX; SPIE: Bellingham, WA, USA, 2008; Volume 6854, p. 685413. [Google Scholar] [CrossRef]
- Kohli, V.; Robles, V.; Cancela, M.L.; Acker, J.P.; Waskiewicz, A.J.; Elezzabi, A.Y. An alternative method for delivering exogenous material into developing zebrafish embryos. Biotechnol. Bioeng. 2007, 98, 1230–1241. [Google Scholar] [CrossRef]
- Kohli, V.; Elezzabi, A.Y. Laser surgery of zebrafish (Danio rerio) embryos using femtosecond laser pulses: Optimal parameters for exogenous material delivery, and the laser’s effect on short- and long-term development. BMC Biotechnol. 2008, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Torres-Mapa, M.L.; Antkowiak, M.; Cizmarova, H.; Ferrier, D.E.K.; Dholakia, K.; Gunn-Moore, F.J. Integrated holographic system for all-optical manipulation of developing embryos. Biomed. Opt. Express 2011, 2, 1564. [Google Scholar] [CrossRef] [Green Version]
- Hosokawa, Y.; Ochi, H.; Iino, T.; Hiraoka, A.; Tanaka, M. Photoporation of biomolecules into single cells in living vertebrate embryos induced by a femtosecond laser amplifier. PLoS ONE 2011, 6, e27677. [Google Scholar] [CrossRef] [Green Version]
- Supatto, W.; Débarre, D.; Farge, E.; Beaurepaire, E. Femtosecond pulse-induced microprocessing of live Drosophila embryos. Med. Laser Appl. 2005, 20, 207–216. [Google Scholar] [CrossRef]
- Engelbrecht, C.J.; Greger, K.; Reynaud, E.G.; Kržic, U.; Colombelli, J.; Stelzer, E.H.K. Three-dimensional laser microsurgery in light-sheet based microscopy (SPIM). Opt. Express 2007, 15, 6420. [Google Scholar] [CrossRef]
- Rauzi, M.; Krzic, U.; Saunders, T.E.; Krajnc, M.; Ziherl, P.; Hufnagel, L.; Leptin, M. Embryo-scale tissue mechanics during Drosophila gastrulation movements. Nat. Commun. 2015, 6, 8677. [Google Scholar] [CrossRef]
- Thayil, A.K.N.; Pereira, A.; Mathew, M.; Artigas, D.; Blanco, E.M.; Loza-Alvarez, P. Decrease in laser ablation threshold for epithelial tissue microsurgery in a living Drosophila embryo during dorsal closure. J. Microsc. 2008, 232, 362–368. [Google Scholar] [CrossRef]
- Galbraith, J.A.; Terasaki, M. Controlled Damage in Thick Specimens by Multiphoton Excitation. Mol. Biol. Cell 2003, 14, 1808–1817. [Google Scholar] [CrossRef] [Green Version]
- Supatto, W.; Fraser, S.E.; Vermot, J. An all-optical approach for probing microscopic flows in living embryos. Biophys. J. 2008, 95, L29–L31. [Google Scholar] [CrossRef] [Green Version]
- Rauzi, M.; Lenne, P.-F. Probing Cell Mechanics with Subcellular Laser Dissection of Actomyosin Networks in the Early Developing Drosophila Embryo. In Tissue Morphogenesis: Methods and Protocols; Nelson, C.M., Ed.; Springer: New York, NY, USA, 2015; pp. 209–218. ISBN 978-1-4939-1164-6. [Google Scholar]
- De Medeiros, G.; Kromm, D.; Balazs, B.; Norlin, N.; Günther, S.; Izquierdo, E.; Ronchi, P.; Komoto, S.; Krzic, U.; Schwab, Y.; et al. Cell and tissue manipulation with ultrashort infrared laser pulses in light-sheet microscopy. Sci. Rep. 2020, 10, 1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, S.-Y.; Moon, H.-Y.; Kim, T.G.; Lee, H.S.; Sidhu, M.S.; Kim, C.; Jeon, J.-P.; Jeoung, S.C. Selective disruption of vascular endothelium of zebrafish embryos by ultrafast laser microsurgical treatment. Biomed. Opt. Express 2015, 6, 4694. [Google Scholar] [CrossRef] [Green Version]
- Yalcin, H.C.; Shekhar, A.; Nishimura, N.; Rane, A.A.; Schaffer, C.B.; Butcher, J.T. Two-photon microscopy-guided femtosecond-laser photoablation of avian cardiogenesis: Noninvasive creation of localized heart defects. Am. J. Physiol. Hearth Circ. Physiol. 2010, 299, 1728–1735. [Google Scholar] [CrossRef] [Green Version]
- Yalcin, H.C. Femtosecond laser photodisruption of vitelline vessels of avian embryos as a technique to study embryonic vascular remodeling. Exp. Biol. Med. 2014, 239, 1644–1652. [Google Scholar] [CrossRef]
- Guglielmi, G.; Barry, J.D.; Huber, W.; De Renzis, S. An Optogenetic Method to Modulate Cell Contractility during Tissue Morphogenesis. Dev. Cell 2015, 35, 646–660. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, E.; Quinkler, T.; De Renzis, S. Guided morphogenesis through optogenetic activation of Rho signalling during early Drosophila embryogenesis. Nat. Commun. 2018, 9, 2366. [Google Scholar] [CrossRef] [PubMed]
- Germond, M.; Nocera, D.; Senn, A.; Rink, K.; Delacrétaz, G.; Fakan, S. Microdissection of mouse and human zona pellucida using a 1.48-microns diode laser beam: Efficacy and safety of the procedure. Fertil. Steril. 1995, 64, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Hartshorn, C.; Anshelevich, A.; Wangh, L.J. Laser zona drilling does not induce hsp70i transcription in blastomeres of eight-cell mouse embryos. Fertil. Steril. 2005, 84, 1547–1550. [Google Scholar] [CrossRef] [PubMed]
- Taylor, T.H.; Gilchrist, J.W.; Hallowell, S.V.; Hanshew, K.K.; Orris, J.J.; Glassner, M.J.; Wininger, J.D. The effects of different laser pulse lengths on the embryo biopsy procedure and embryo development to the blastocyst stage. J. Assist. Reprod. Genet. 2010, 27, 663–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malter, H.E.; Schimmel, T.; Cohen, J. Zona dissection by infrared laser: Developmental consequences in the mouse, technical considerations, and controlled clinical trial. Reprod. Biomed. Online 2001, 3, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Douglas-Hamilton, D.H.; Conia, J. Thermal effects in laser-assisted pre-embryo zona drilling. J. Biomed. Opt. 2001, 6, 205. [Google Scholar] [CrossRef]
- Chatzimeletiou, K.; Morrison, E.E.; Panagiotidis, Y.; Prapas, N.; Prapas, Y.; Rutherford, A.J.; Grudzinskas, G.; Handyside, A.H. Comparison of effects of zona drilling by non-contact infrared laser or acid Tyrode’s on the development of human biopsied embryos as revealed by blastomere viability, cytoskeletal analysis and molecular cytogenetics. Reprod. Biomed. Online 2005, 11, 697–710. [Google Scholar] [CrossRef]
- Tucker, M.J.; Ball, G.D. Assisted hatching as a technique for use in human in vitro fertilization and embryo transfer is long overdue for careful and appropriate study. J. Clin. Embryol. 2009, 12, 9–14. [Google Scholar]
- Il’ina, I.V.; Sitnikov, D.S.; Ovchinnikov, A.V.; Agranat, M.B.; Khramova, Y.V.; Semenova, M.L. Noncontact microsurgery and micromanipulation of living cells with combined system femtosecond laser scalpel-optical tweezers. In Biophotonics: Photonic Solutions for Better Health Care III; Popp, J., Drexler, W., Tuchin, V.V., Matthews, D.L., Eds.; SPIE: Bellingham, WA, USA, 2012; Volume 8427, pp. 139–146. [Google Scholar]
- Ilina, I.V.; Khramova, Y.V.; Filatov, M.A.; Semenova, M.L.; Sitnikov, D.S. Application of femtosecond laser scalpel and optical tweezers for noncontact biopsy of late preimplantation embryos. High Temp. 2015, 53, 804–809. [Google Scholar] [CrossRef]
- Sitnikov, D.S.; Ovchinnikov, A.V.; Ilina, I.V.; Chefonov, O.V.; Agranat, M.B. Laser microsurgery of cells by femtosecond laser scalpel and optical tweezers. High Temp. 2014, 52, 803–808. [Google Scholar] [CrossRef]
- McArthur, S.J.; Leigh, D.; Marshall, J.T.; de Boer, K.A.; Jansen, R.P.S. Pregnancies and live births after trophectoderm biopsy and preimplantation genetic testing of human blastocysts. Fertil. Steril. 2005, 84, 1628–1636. [Google Scholar] [CrossRef]
- Ilina, I.V.; Khramova, Y.V.; Ivanova, A.D.; Filatov, M.A.; Silaeva, Y.Y.; Deykin, A.V.; Sitnikov, D.S. Controlled hatching at the prescribed site using femtosecond laser for zona pellucida drilling at the early blastocyst stage. J. Assist. Reprod. Genet. 2021, 38, 517–529. [Google Scholar] [CrossRef]
- Ilina, I.V.; Khramova, Y.V.; Filatov, M.A.; Sitnikov, D.S. Femtosecond laser is effective tool for zona pellucida engraving and tagging of preimplantation mammalian embryos. J. Assist. Reprod. Genet. 2019, 36, 1251–1261. [Google Scholar] [CrossRef]
- Spriggs, M. IVF mixup: White couple have black babies. J. Med. Ethics 2003, 29, 65. [Google Scholar] [CrossRef] [Green Version]
- Bender, L. To err is human’. ART mix-ups: A labor-based, relational. proposal. J. Race Gend. Justice 2006, 9, 443–508. [Google Scholar]
- Ilina, I.V.; Khramova, Y.V.; Filatov, M.A.; Sitnikov, D.S. Application of femtosecond laser microsurgery in assisted reproductive technologies for preimplantation embryo tagging. Biomed. Opt. Express 2019, 10, 2985–2995. [Google Scholar] [CrossRef]
- Squirrell, J.M.; Wokosin, D.L.; White, J.G.; Bavister, B.D. Long-term two-photon fluorescence imaging of mammalian embryos without compromising viability. Nat. Biotechnol. 1999, 17, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Molinari, E.; Evangelista, F.; Racca, C.; Cagnazzo, C.; Revelli, A. Polarized light microscopy-detectable structures of human oocytes and embryos are related to the likelihood of conception in IVF. J. Assist. Reprod. Genet. 2012, 29, 1117–1122. [Google Scholar] [CrossRef]
- Thayil, A.; Watanabe, T.; Jesacher, A.; Wilson, T.; Srinivas, S.; Booth, M. Long-term imaging of mouse embryos using adaptive harmonic generation microscopy. J. Biomed. Opt. 2011, 16, 046018. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.-S.; Chen, S.-U.; Lee, Y.-W.; Yang, Y.-S.; Sun, C.-K. Higher harmonic generation microscopy of in vitro cultured mammal oocytes and embryos. Opt. Express 2008, 16, 11574. [Google Scholar] [CrossRef]
- Kyvelidou, C.; Tserevelakis, G.J.; Filippidis, G.; Ranella, A.; Kleovoulou, A.; Fotakis, C.; Athanassakis, I. Following the course of pre-implantation embryo patterning by non-linear microscopy. J. Struct. Biol. 2011, 176, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, T.; Venturas, M.; Aghvami, S.A.; Yang, X.; Fraden, S.; Sakkas, D.; Needleman, D.J. Combined noninvasive metabolic and spindle imaging as potential tools for embryo and oocyte assessment. Hum. Reprod. 2019, 34, 2349–2361. [Google Scholar] [CrossRef]
- Ma, N.; de Mochel, N.R.; Pham, P.D.; Yoo, T.Y.; Cho, K.W.Y.; Digman, M.A. Label-free assessment of pre-implantation embryo quality by the Fluorescence Lifetime Imaging Microscopy (FLIM)-phasor approach. Sci. Rep. 2019, 9, 13206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomeroy, K.O.; Reed, M.L. The Effect of Light on Embryos and Embryo Culture. J. Reprod. Stem Cell Biotechnol. 2012, 3, 46–54. [Google Scholar] [CrossRef]
- Oh, S.J.; Gong, S.P.; Lee, S.T.; Lee, E.J.; Lim, J.M. Light intensity and wavelength during embryo manipulation are important factors for maintaining viability of preimplantation embryos in vitro. Fertil. Steril. 2007, 88, 1150–1157. [Google Scholar] [CrossRef]
- Turpaev, K.T. Reactive Oxygen Species and Regulation of Gene Expression. Biochemistry 2002, 67, 281–292. [Google Scholar] [CrossRef]
- Leite, R.F.; Annes, K.; Ispada, J.; de Lima, C.B.; Dos Santos, É.C.; Fontes, P.K.; Nogueira, M.F.G.; Milazzotto, M.P. Oxidative Stress Alters the Profile of Transcription Factors Related to Early Development on In Vitro Produced Embryos. Oxid. Med. Cell. Longev. 2017, 2017, 1502489. [Google Scholar] [CrossRef] [Green Version]
- De Assis, P.M.; Castro, L.S.; Siqueira, A.F.P.; De Carvalho Delgado, J.; Dos Santos Hamilton, T.R.; Goissis, M.D.; Mendes, C.M.; Nichi, M.; Visintin, J.A.; Assumpção, M.E.O.D.Á. System for evaluation of oxidative stress on in-vitro-produced bovine embryos. Reprod. Biomed. Online 2015, 31, 577–580. [Google Scholar] [CrossRef] [Green Version]
- Talone, B.; Bazzarelli, M.; Schirato, A.; Dello Vicario, F.; Viola, D.; Jacchetti, E.; Bregonzio, M.; Raimondi, M.T.; Cerullo, G.; Polli, D. Phototoxicity induced in living HeLa cells by focused femtosecond laser pulses: A data-driven approach. Biomed. Opt. Express 2021, 12, 7886. [Google Scholar] [CrossRef]
- Burhans, W.C.; Heintz, N.H. The cell cycle is a redox cycle: Linking phase-specific targets to cell fate. Free Radic. Biol. Med. 2009, 47, 1282–1293. [Google Scholar] [CrossRef]
- Yan, W.; He, H.; Wang, Y.; Wang, Y.; Hu, M.; Wang, C. Controllable generation of reactive oxygen species by femtosecond-laser irradiation. Appl. Phys. Lett. 2014, 104, 083703. [Google Scholar] [CrossRef]
- Baumgart, J.; Kuetemeyer, K.; Lubatschowski, H.; Heisterkamp, A.; Bintig, W.; Ngezahayo, A.; Ertmer, W. Erratum: Repetition rate dependency of reactive oxygen species formation during femtosecond laser-based cell surgery. J. Biomed. Opt. 2010, 14, 054040, Erratum in J. Biomed. Opt. 2010, 15, 059802. [Google Scholar] [CrossRef]
- Baumgart, J.; Bintig, W.; Ngezahayo, A.; Lubatschowski, H.; Heisterkamp, A. Fs-laser-induced Ca^2+ concentration change during membrane perforation for cell transfection. Opt. Express 2010, 18, 2219. [Google Scholar] [CrossRef] [Green Version]
- Davidson, L.M.; Liu, Y.; Griffiths, T.; Jones, C.; Coward, K. Laser technology in the ART laboratory: A narrative review. Reprod. Biomed. Online 2019, 38, 725–739. [Google Scholar] [CrossRef]
- Honguntikar, S.D.; Salian, S.R.; D’Souza, F.; Uppangala, S.; Kalthur, G.; Adiga, S.K. Epigenetic changes in preimplantation embryos subjected to laser manipulation. Lasers Med. Sci. 2017, 32, 2081–2087. [Google Scholar] [CrossRef]
- Sitnikov, D.S.; Ilina, I.V.; Pronkin, A.A. Assessment of the thermal effect of femtosecond and millisecond laser pulses in microsurgery of mammalian embryos. Quantum Electron. 2022, 52, 482. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
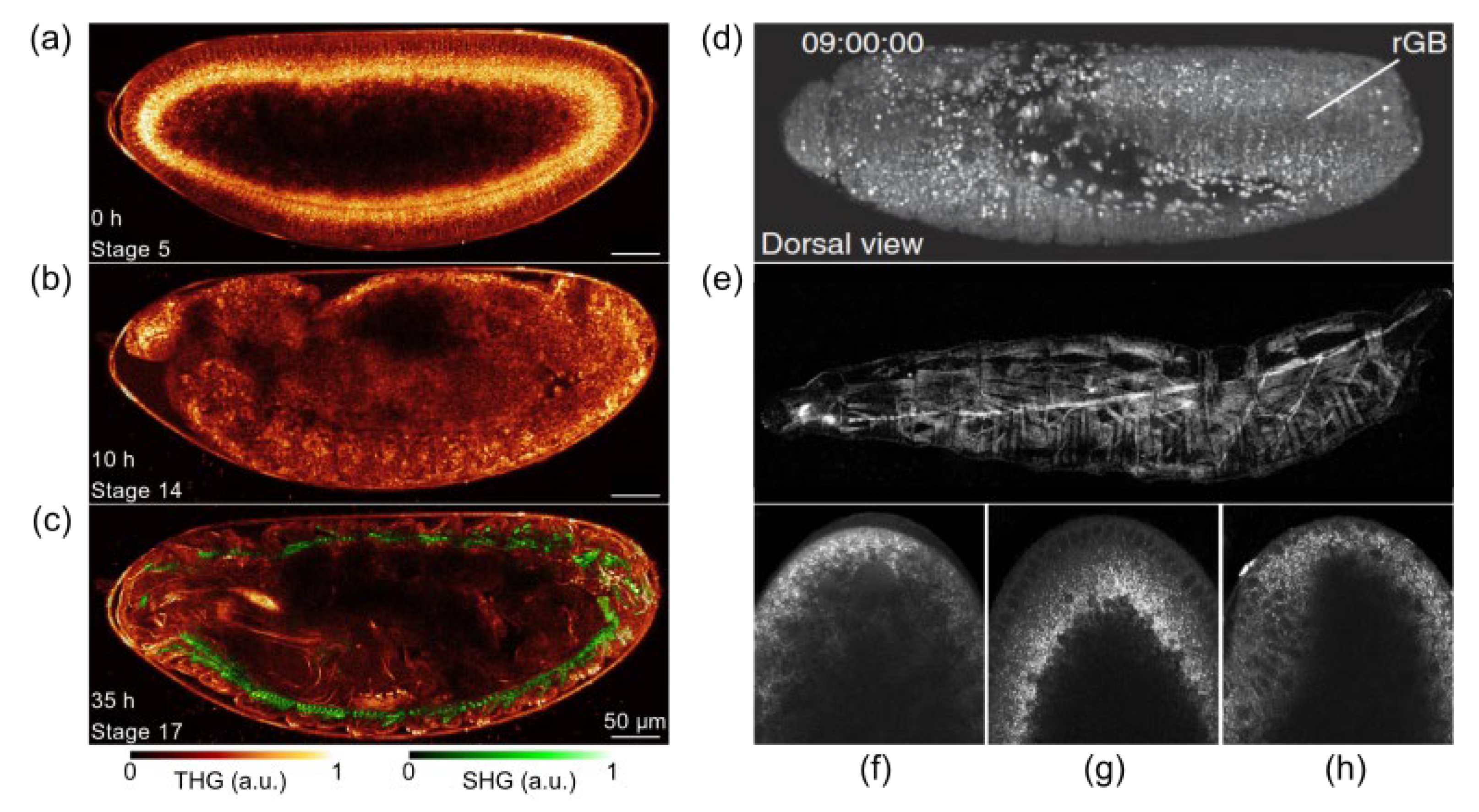{kind=link}
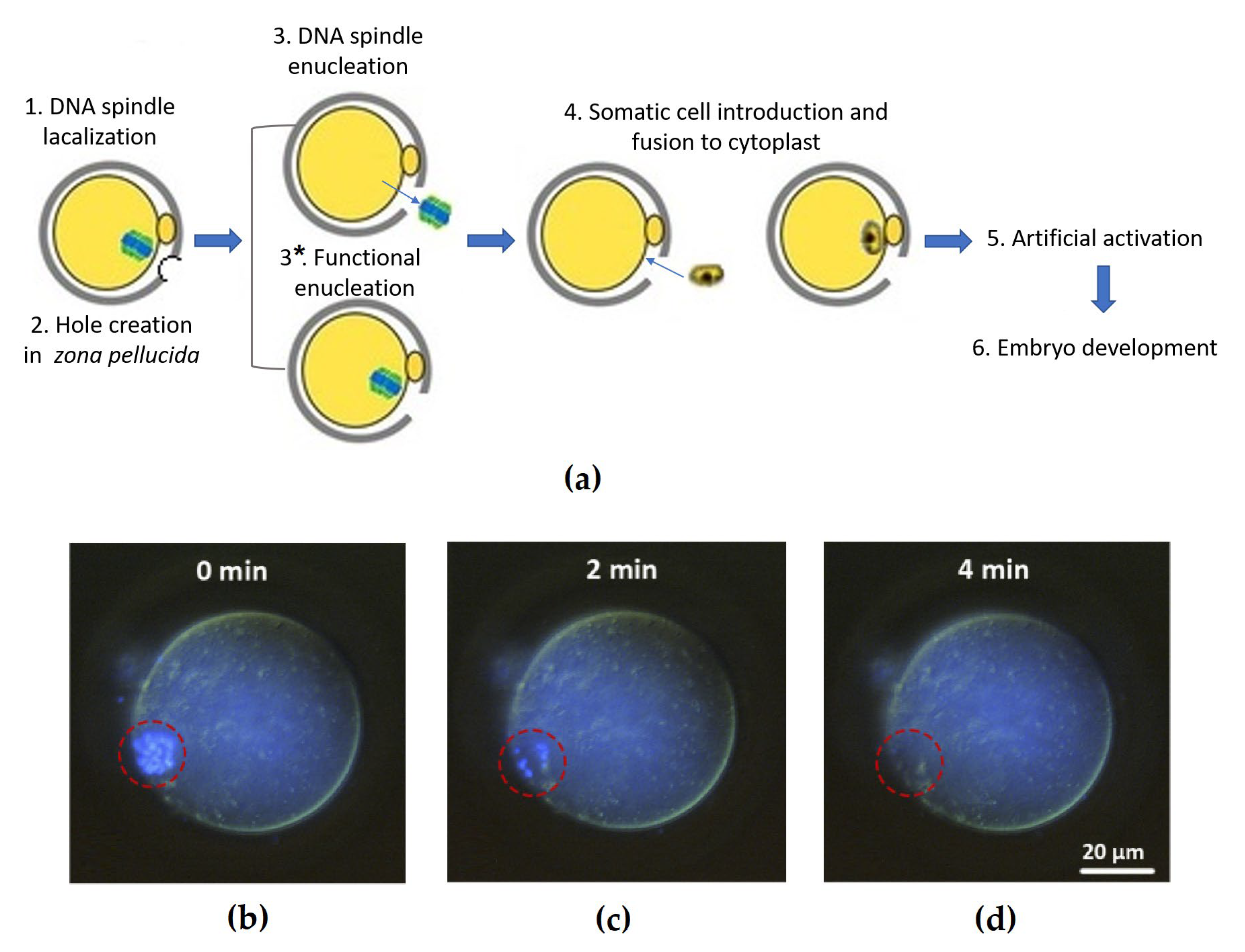{kind=link}
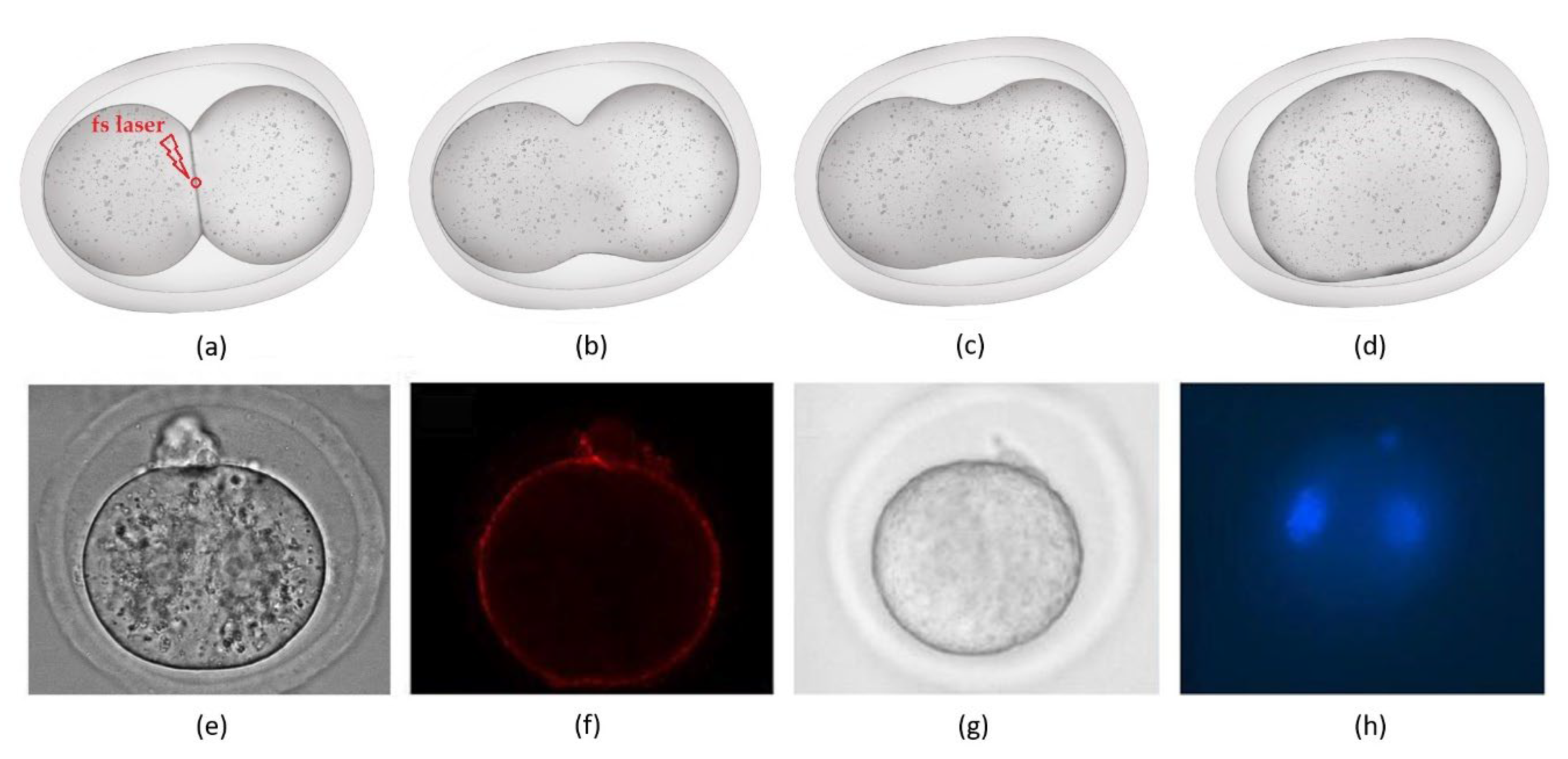{kind=link}
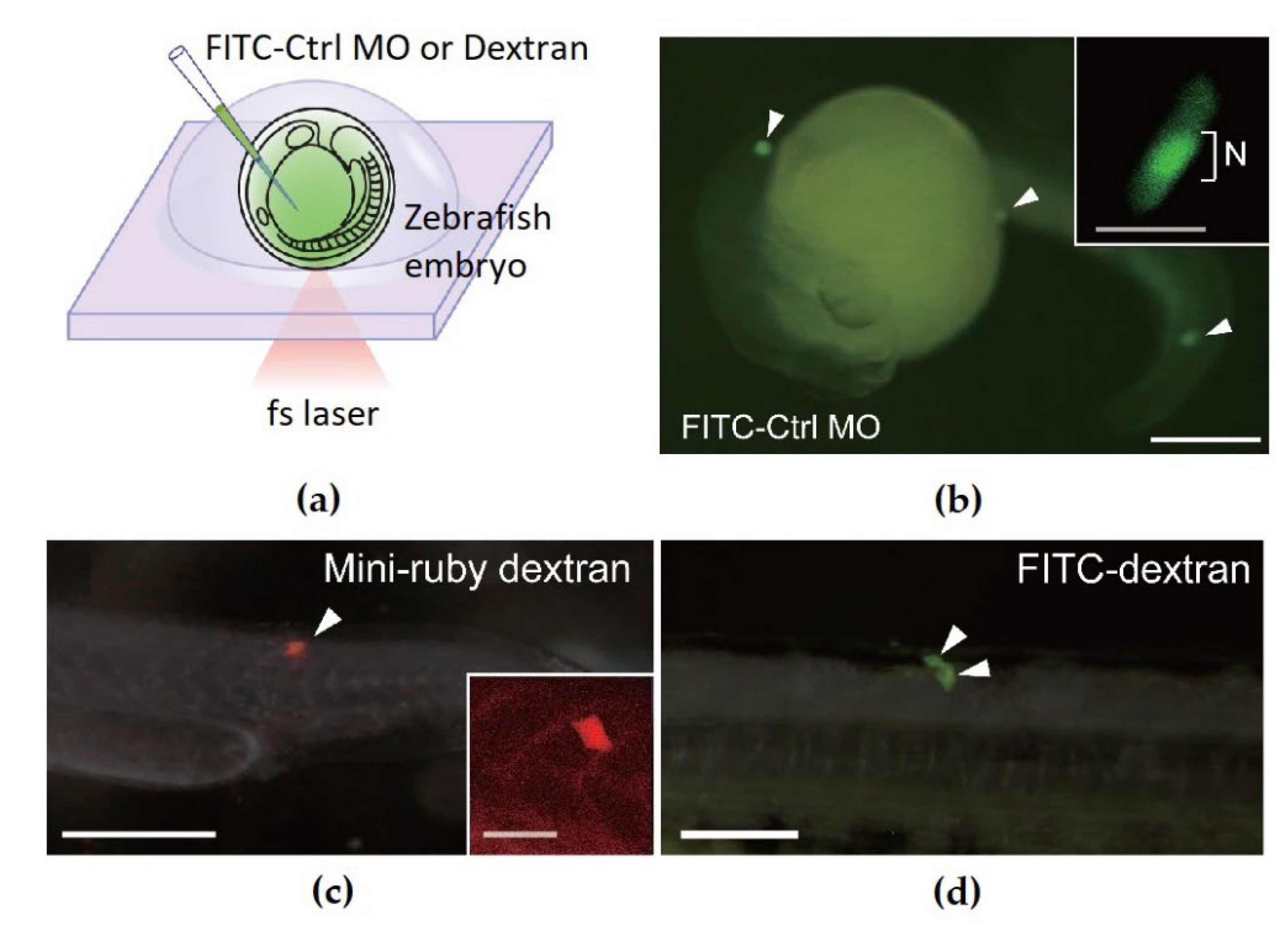{kind=link}
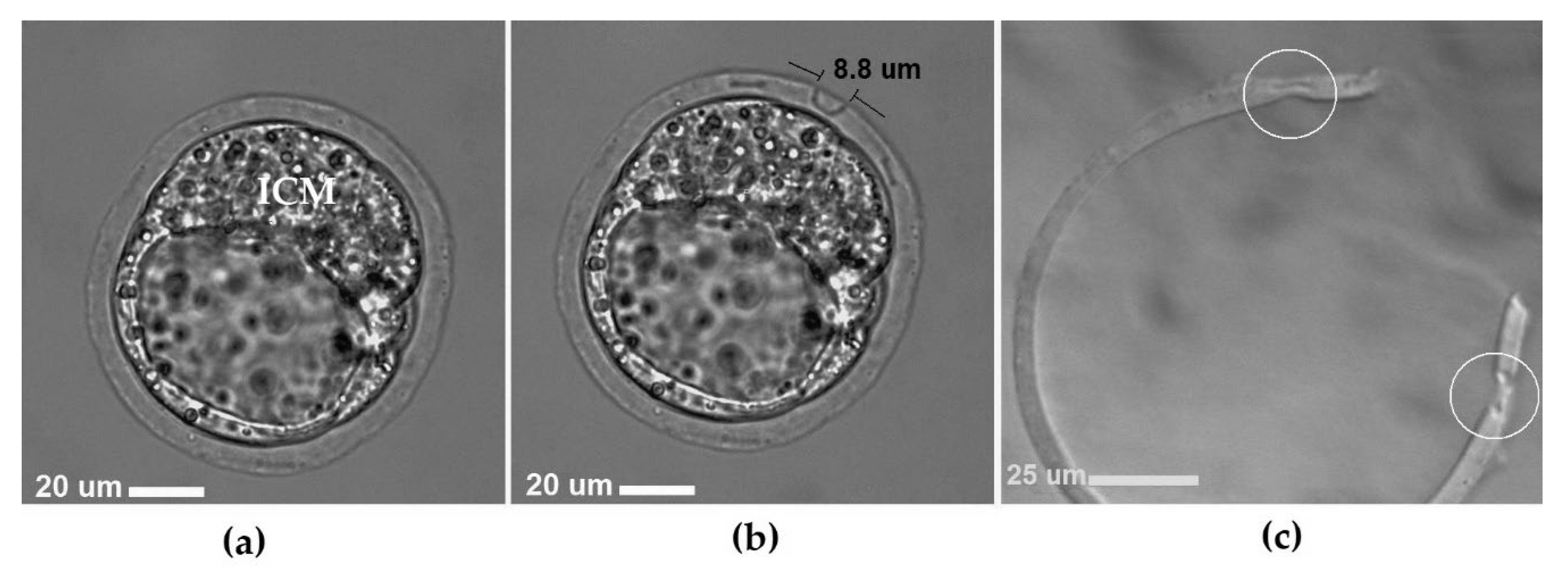{kind=link}
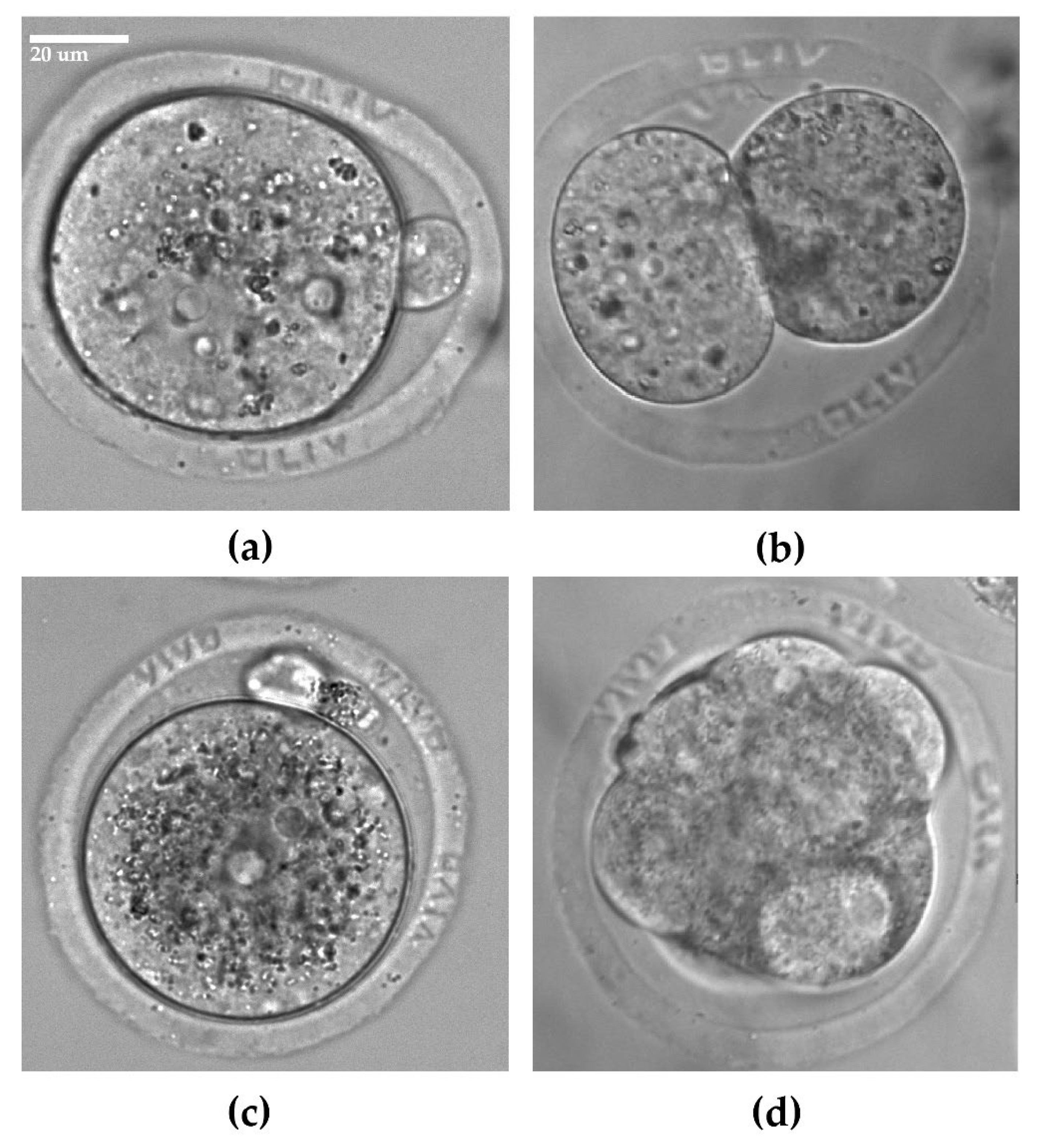{kind=link}
| Type of Oocytes/ Embryos | Type of Manipulation | Laser Exposure Parameters | Efficiency of Enucleation/Fusion | References |
|---|---|---|---|---|
| Mouse | Functional enucleation (nucleoli, cytoplasm, metaphase plate, pronucleus irradiation), Blastomere fusion | Ti:sapphire laser τ = 2 ps, λ = 800 nm, = 80 MHz, Pav = 0.4 W, E = 5 nJ, t = 0.3 s | Efficiency of nuclear inactivation of one of two blastomeres: 75–80% Efficiency of oocyte inactivation after metaphase plate irradiation: 100% Fusion efficiency/Blastocyst formation: 35.8%/33.3%–66.7%/95% (dependent on the site of laser impact on the blastomere contact border) | [71] |
| Porcine | Functional enucleation | Ti:sapphire laser τ ~ 275 fs in the sample, λ = 720 nm, = 1 MHz, E = 2.5 nJ, υ = 100 µm s−1 | Efficiency of enucleation: 96% | [72] |
| Porcine | Blastomere fusion | Ti:sapphire laser τ ~ 275 fs in the sample, λ = 720 nm, = 80 MHz, t = 20 ms, F = 0.36 J cm–2 (Pav = 430 mW), t = 100 ms, F = 0.23 J cm−2 (Pav = 670 mW) | Fusion efficiency/Cell viability/Blastocyst formation: 54%/95%/70% (for t = 20 ms) 44%/73%/43% (for t = 100 ms) | [73] |
| Mouse | Blastomere fusion | Cr:forsterite seed oscillator and regenerative amplifier τ = 100 fs, λ = 620 nm, = 10 Hz, E = 30–50 nJ | Fusion efficiency/Blastocyst formation: 89%/50% | [74,75] |
| Mouse | Blastomere fusion | Ti:sapphire laser τ = 100 fs (at the sample), λ = 800 nm, = 80 MHz, Pav = 80 mW, E = 0.3, 1, 2 nJ, t = 15, 30, 60 ms, 2.0–13.2 × 1011 W cm−2 | Fusion efficiency/Blastocyst formation rate: 29%/49% (for E = 1 nJ, t = 30 ms) | [76] |
| Mouse | Oocyte fusion, fusion of oocyte with blastomere, fusion of 2–3 blastomeres inside 4-cell embryos | Ti:sapphire laser τ = 2 ps, λ = 800 nm, = 80 MHz, Pav = 0.4 W | Oocyte fusion efficiency: 46/7% Oocyte/blastomere fusion efficiency: 21.2% Efficiency of two-blastomere fusion inside 4-cell embryo/Blastocyst formation: 61.5%/78.1% Efficiency of two pairs of blastocyst fusion inside 4-cell embryo/Blastocyst formation: 52.2%/90% Efficiency of three blastomere fusion inside 4-cell embryo/Blastocyst formation: 44.4%/50% | [77] |
| Mouse | Oocyte fusion, Blastomere fusion | Ti:sapphire laser τ = 100 fs in the sample, λ = 690–1000 nm, = 80 MHz, Pav = 0.8 W (before the objective), E = 1 nJ, t = 15 ms (for oocyte fusion), t = 30 ms (for blastomere fusion) | No data | [78] |
| Type of Embryos | Type of Manipulation | Laser Exposure Parameters | References |
|---|---|---|---|
| Zebrafish embryos | Optoinjection and transfection | Ti:sapphire laser τ = sub-10 fs, λ = 800 nm, = 80 MHz, Pav = 40–220 mW, E = 0.5–3 nJ, Ipeak =1011–1012 W cm–2, Pav = 120–160 mW, dwell time: 200–500 ms (for quantum dots introduction) Pav = 40–45 mW, dwell time: 200–500 ms (for transfection) | [84,85] |
| Pomatoceros lamarckii embryos | Optoinjection | Ti:sapphire laser τ = 180 fs, λ = 800 nm, = 80 MHz, Pav = 52–78 mW, t = 10–40 ms | [87] |
| Zebrafish, Chick, Shark and Mouse embryos | Optoinjection and transfection | Ti:sapphire laser τ = 120 fs, λ = 800 nm, = 1 kHz, 50 pulses for exposure E = 100–800 nJ | [88] |
| Embryo/Organ Type | Type of Manipulation | Laser Exposure Parameters | References |
|---|---|---|---|
| Drosophila melanogaster, Oregon-R strain | 3D laser ablations | Ti:sapphire laser oscillator and an optical parametric oscillator τ = 130 fs, λ = 830 nm (for ablation), 920 nm (for TPEF), 1180 nm (for THG) = 76 MHz, Pav = 50–275 mW, Ipeak = , E = 0.7–3.6 nJ Ipeak = (for TPEF) Number of line scans–3, 100 µm long | [89] |
| Drosophila melanogaster embryo (stage 15) | 3D laser ablations | Frequency tripled Nd:YAG laser τ = 470 ps, λ = 355 nm, = 1 kHz, 100 pulses, E = 0.74 ± 0.04 µJ, Ipeak = 744 ± 40 GW cm−2 | [90] |
| Drosophila melanogaster embryo | Local tissue immobilization, actomyosin meshwork ablation | For tissue immobilization: λ = 1030 nm, = 50 MHz, τ = 200 fs t = 40 ms, Pav = 200 mW, For actomyosin meshwork ablation λ = 950 nm | [91,95] |
| Drosophila embryo Zebrafish embryo | Nuclei ablation Neuron dissection Tissue cauterization Optogenetic manipulations | λ = 1025 nm, = 54 MHz, τ = 180–200 fs For nuclei ablation t = 15 ms, 4 times Pav = 800 mW, For soma ablation: t = 9 ms, 5 times Pav = 880 mW For axon ablation: t = 3 ms Pav = 604 mW For tissue cauterization: t = 40 ms Pav = 400 mW Optogenetics: λ = 780 nm = 80 MHz, τ = 150 fs | [96] |
| Drosophila embryo | Epithelial tissue microsurgery | Ti:sapphire laser τ = 160 fs, λ = 870 nm, = 76 MHz, Pav (before the objective) = 20–40 mW (for ablation) Pav (before the objective) = 8 mW (for imaging) E = 0.13–0.15 nJ (for microablation after dorsal closure) E = 0.095 nJ (for microablation during dorsal closure) υ = 0.66 µm ms−1 Number of line scans–160, 13 µm long | [92] |
| Sea urchin embryo, Starfish embryo, Zebrafish embryo | Dendrite ablation, mitotic pole ablation, plasma membrane and nuclear envelope wound | Ti:sapphire laser τ = 100 fs, λ = 800 nm, = 76 MHz, Pav = 60 mW (at the specimen), E = 0.8 nJ | [93] |
| Zebrafish embryo | Ablation of mesodermal cells | τ = 300 fs, λ = 820 nm, = 80 MHz | [94] |
| Zebrafish embryo | Opto-disruption of vascular structure | τ = 150 fs, λ = 800 nm, = 1 kHz F = 5–50 J cm−2 Ipeak = 30–300 × 1012 W cm−2 | [97] |
| Avian embryo heart | Ablation of the superior atrioventricular (AV) cushion and vitelline vessels | τ = 50 fs, = 1 kHz, E = 200 nJ | [98,99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ilina, I.V.; Sitnikov, D.S. Application of Ultrashort Lasers in Developmental Biology: A Review. Photonics 2022, 9, 914. https://doi.org/10.3390/photonics9120914
Ilina IV, Sitnikov DS. Application of Ultrashort Lasers in Developmental Biology: A Review. Photonics. 2022; 9(12):914. https://doi.org/10.3390/photonics9120914
Chicago/Turabian StyleIlina, Inna V., and Dmitry S. Sitnikov. 2022. "Application of Ultrashort Lasers in Developmental Biology: A Review" Photonics 9, no. 12: 914. https://doi.org/10.3390/photonics9120914