
Kinase Fusions in Spitz Melanocytic Tumors: The Past, the Present, and the Future

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
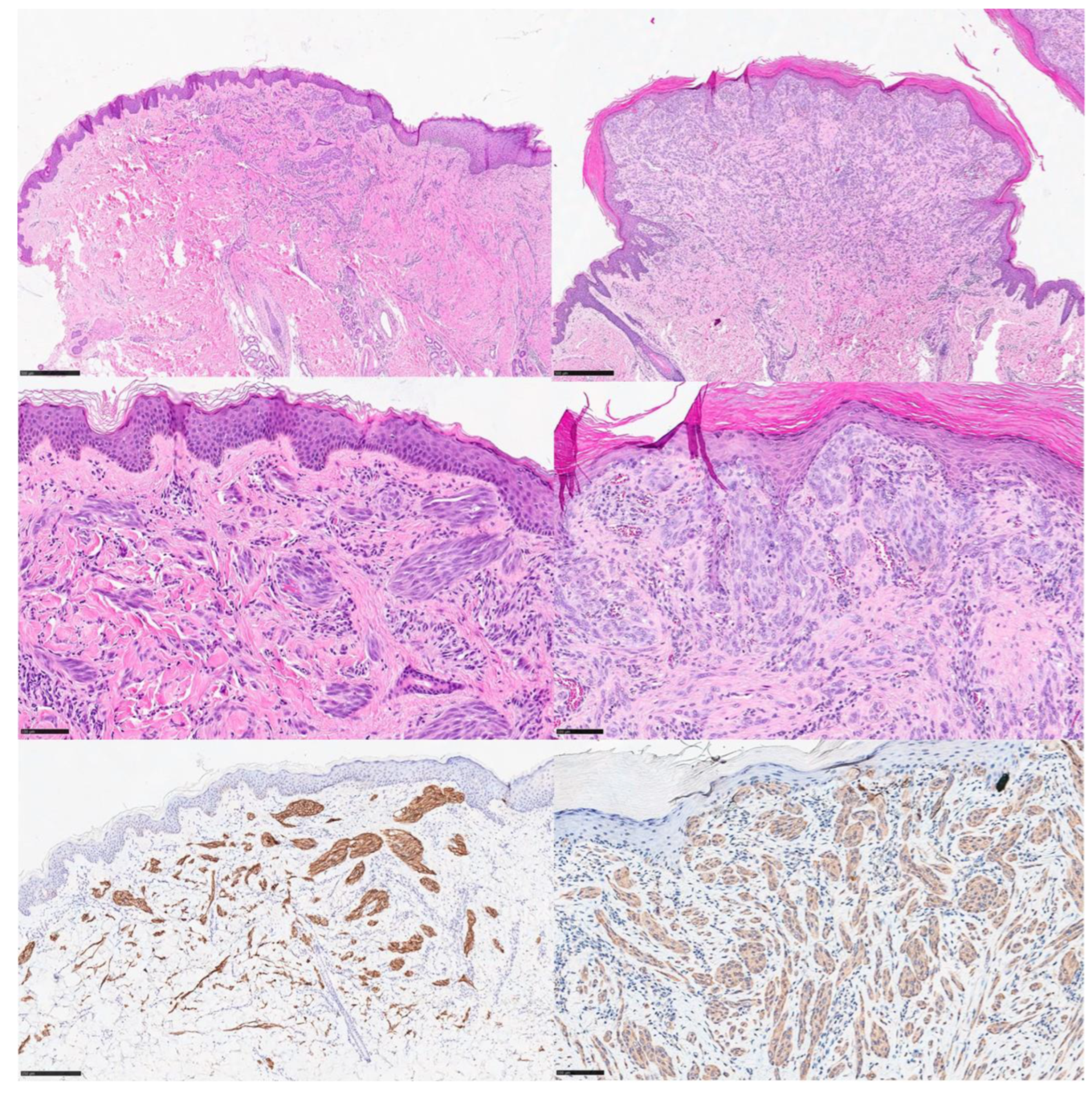
3.1. ALK-Rearranged Spitz Tumors
3.2. ROS1-Rearranged Spitz Tumors
3.3. NTRK-Rearranged Spitz Tumors
3.3.1. NTRK1-Rearranged Spitz Tumors
3.3.2. NTRK2-Rearranged Spitz Tumors
3.3.3. NTRK3-Rearranged Spitz Tumors
3.4. RET-Rearranged Spitz Tumors
3.5. MET-Rearranged Spitz Tumors
3.6. Serine–Threonine Kinase-Rearranged Spitz Tumors
3.6.1. BRAF-Rearranged Spitz Tumors
3.6.2. RAF1-Rearranged Spitz Tumors
3.6.3. MAP3K8-Rearranged Spitz Tumors
4. Conclusions and Future Directions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Brown, A.; Sawyer, J.D.; Neumeister, M.W. Spitz Naevus: Review and Update. Clin. Plast. Surg. 2021, 48, 677–686. [Google Scholar] [CrossRef]
- Yeh, I.; Busam, K.J. Spitz melanocytic tumours—A review. Histopathology 2022, 80, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, T.; He, J.; Yelensky, R.; Esteve-Puig, R.; Botton, T.; Yeh, I.; Lipson, D.; Otto, G.; Brennan, K.; Murali, R.; et al. Kinase fusions are frequent in Spitz tumours and spitzoid melanomas. Nat. Commun. 2014, 5, 3116. [Google Scholar] [CrossRef]
- WHO Classification of Tumours Editorial Board. Skin Tumours, WHO Classification of Tumours Series; 5th ed.; International Agency for Research on Cancer: Lyon, France, 2023; Volume 12, Available online: https://tumourclassification.iarc.who.int/chapters/64 (accessed on 26 December 2023).
- Shalin, S.C. A review of kinase fusions in melanocytic tumors. Lab. Investig. 2017, 97, 158–165. [Google Scholar] [CrossRef]
- Du, X.; Shao, Y.; Qin, H.; Tai, Y.; Gao, H. ALK-rearrangement in non-small-cell lung cancer (NSCLC). Thorac. Cancer 2018, 9, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Dickson, B.C.; Swanson, D.; Charames, G.S.; Fletcher, C.D.; Hornick, J.L. Epithelioid fibrous histiocytoma: Molecular characterization of ALK fusion partners in 23 cases. Mod. Pathol. 2018, 31, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Antonescu, C.R.; Suurmeijer, A.J.; Zhang, L.; Sung, Y.-S.; Jungbluth, A.A.; Travis, W.D.; Al-Ahmadie, H.; Fletcher, C.D.; Alaggio, R. Molecular Characterization of Inflammatory Myofibroblastic Tumors with Frequent ALK and ROS1 Gene Fusions and Rare Novel RET Rearrangement. Am. J. Surg. Pathol. 2015, 39, 957–967. [Google Scholar] [CrossRef]
- Yeh, I.; de la Fouchardiere, A.; Pissaloux, D.; Mully, T.W.; Garrido, M.C.; Vemula, S.S.; Busam, K.J.; LeBoit, P.E.; McCalmont, T.H.; Bastian, B.C. Clinical, Histopathologic, and Genomic Features of Spitz Tumors with ALK Fusions. Am. J. Surg. Pathol. 2015, 39, 581–591. [Google Scholar] [CrossRef]
- Busam, K.J.; Kutzner, H.; Cerroni, L.; Wiesner, T. Clinical and Pathologic Findings of Spitz Nevi and Atypical Spitz Tumors with ALK Fusions. Am. J. Surg. Pathol. 2014, 38, 925–933. [Google Scholar] [CrossRef]
- Wu, G.; Barnhill, R.L.; Lee, S.; Li, Y.; Shao, Y.; Easton, J.; Dalton, J.; Zhang, J.; Pappo, A.; Bahrami, A. The landscape of fusion transcripts in spitzoid melanoma and biologically indeterminate spitzoid tumors by RNA sequencing. Mod. Pathol. 2016, 29, 359–369. [Google Scholar] [CrossRef]
- Brown, R.A.; Wang, J.Y.; Raghavan, S.S.; Zhang, J.; Wan, D.C.; Born, D.; Koo, M.; Hazard, F.K.; Novoa, R.A.; Rieger, K.E. ALK -positive compound Spitz nevus with extensive perineural and intraneural neurotropism. J. Cutan. Pathol. 2021, 48, 154–159. [Google Scholar] [CrossRef]
- Wiesner, T.; Kutzner, H.; Cerroni, L.; Mihm, M.C.; Busam, K.J., Jr.; Murali, R. Genomic aberrations in spitzoid melanocytic tumours and their implications for diagnosis, prognosis and therapy. Pathology 2016, 48, 113–131. [Google Scholar] [CrossRef]
- Saraggi, D.; Salmaso, R.; Zamuner, C.; Munari, G.; Lanza, C.; Alaibac, M.S.; Bassetto, F.; Rugge, M.; Montesco, M.C.; Cerroni, L.; et al. Prevalence of ALK gene alterations among the spectrum of plexiform spitzoid lesions. J. Am. Acad. Dermatol. 2018, 79, 728–735. [Google Scholar] [CrossRef]
- Bahrani, E.; Kunder, C.A.; Teng, J.M.; Brown, R.A.; Rieger, K.E.; Novoa, R.A.; Cloutier, J.M. Spitz nevus with EHBP1-ALK fusion and distinctive membranous localization of ALK. J. Cutan. Pathol. 2022, 49, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Uguen, A.; De Braekeleer, M. ROS1 fusions in cancer: A review. Futur. Oncol. 2016, 12, 1911–1928. [Google Scholar] [CrossRef] [PubMed]
- Gerami, P.; Kim, D.; Compres, E.V.; Zhang, B.; Khan, A.U.; Sunshine, J.C.; Quan, V.L.; Busam, K. Clinical, morphologic, and genomic findings in ROS1 fusion Spitz neoplasms. Mod. Pathol. 2021, 34, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Hagstrom, M.; Fumero-Velázquez, M.; Dhillon, S.; Olivares, S.; Gerami, P. An update on genomic aberrations in Spitz naevi and tumours. Pathology 2023, 55, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Amatu, A.; Sartore-Bianchi, A.; Bencardino, K.; Pizzutilo, E.; Tosi, F.; Siena, S. Tropomyosin receptor kinase (TRK) biology and the role of NTRK gene fusions in cancer. Ann. Oncol. 2019, 30 (Suppl. S8), viii5–viii15. [Google Scholar] [CrossRef]
- Yeh, I.; Busam, K.J.; McCalmont, T.H.; LeBoit, P.E.; Pissaloux, D.; Alberti, L.; de la Fouchardière, A.; Bastian, B.C. Filigree-like Rete Ridges, Lobulated Nests, Rosette-like Structures, and Exaggerated Maturation Characterize Spitz Tumors with NTRK1 Fusion. Am. J. Surg. Pathol. 2019, 43, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Truzzi, F.; Marconi, A.; Lotti, R.; Dallaglio, K.; French, L.E.; Hempstead, B.L.; Pincelli, C. Neurotrophins and Their Receptors Stimulate Melanoma Cell Proliferation and Migration. J. Investig. Dermatol. 2008, 128, 2031–2040. [Google Scholar] [CrossRef] [PubMed]
- Khotskaya, Y.B.; Holla, V.R.; Farago, A.F.; Shaw, K.R.M.; Meric-Bernstam, F.; Hong, D.S. Targeting TRK family proteins in cancer. Pharmacol. Ther. 2017, 173, 58–66. [Google Scholar] [CrossRef]
- Yin, L.; Shi, C.; He, X.; Qiu, Y.; Chen, H.; Chen, M.; Zhang, Z.; Chen, Y.; Zhou, Y.; Zhang, H. NTRK-rearranged spindle cell neoplasms: A clinicopathological and molecular study of 13 cases with peculiar characteristics at one of the largest institutions in China. Pathology 2023, 55, 362–374. [Google Scholar] [CrossRef]
- Cappellesso, R.; Nozzoli, F.; Marino, F.Z.; Simi, S.; Castiglione, F.; De Giorgi, V.; Cota, C.; Senetta, R.; Scognamiglio, G.; Anniciello, A.M.; et al. NTRK Gene Fusion Detection in Atypical Spitz Tumors. Int. J. Mol. Sci. 2021, 22, 12332. [Google Scholar] [CrossRef] [PubMed]
- Forschnern, A.; Forchhammer, S.; Bonzheim, I. NTRK gene fusions in melanoma: Detection, prevalence and potential thera-peutic implications. J. Dtsch. Dermatol. Ges. 2020, 18, 1387–1392. [Google Scholar] [CrossRef]
- Amin, S.M.; Haugh, A.M.; Lee, C.Y.; Zhang, B.; Bubley, J.A.; Merkel, E.A.; Verzì, A.E.; Gerami, P. A Comparison of Morphologic and Molecular Features of BRAF, ALK, and NTRK1 Fusion Spitzoid Neoplasms. Am. J. Surg. Pathol. 2017, 41, 491–498. [Google Scholar] [CrossRef]
- Pozzo, C.A.D.; Cappellesso, R. The Morpho-Molecular Landscape of Spitz Neoplasms. Int. J. Mol. Sci. 2022, 23, 4211. [Google Scholar] [CrossRef]
- Kervarrec, T.; Pissaloux, D.; Tirode, F.; Samimi, M.; Jacquemus, J.; Castillo, C.; de la Fouchardière, A. Morphologic features in a series of 352 Spitz melanocytic proliferations help predict their oncogenic drivers. Virchows Arch. 2022, 480, 369–382. [Google Scholar] [CrossRef]
- Kiuru, M.; Jungbluth, A.; Kutzner, H.; Wiesner, T.; Busam, K.J. Spitz Tumors: Comparison of Histological Features in Rela-tionship to Immunohistochemical Staining for ALK and NTRK1. Int. J. Surg. Pathol. 2016, 24, 200–206. [Google Scholar] [CrossRef]
- Šekoranja, D.; Pižem, J.; Luzar, B. An Update on Molecular Genetic Aberrations in Spitz Melanocytic Proliferations: Correlation with Morphological Features and Biological Behavior. Acta Medica Acad. 2021, 50, 157–174. [Google Scholar] [CrossRef]
- Hondelink, L.M.; Schrader, A.M.; Aghmuni, G.A.; Solleveld-Westerink, N.; Cleton-Jansen, A.-M.; van Egmond, D.; Boot, A.; Ouahoud, S.; Khalifa, M.N.; Lam, S.W.; et al. The sensitivity of pan-TRK immunohistochemistry in solid tumours: A meta-analysis. Eur. J. Cancer 2022, 173, 229–237. [Google Scholar] [CrossRef]
- Goto, K.; Pissaloux, D.; Tirode, F.; de la Fouchardière, A. Spitz nevus with a novel TFG-NTRK2 fusion: The first case report of NTRK2-rearranged Spitz/Reed nevus. J. Cutan. Pathol. 2021, 48, 1193–1196. [Google Scholar] [CrossRef]
- Mansour, B.; Vanecek, T.; Kastnerova, L.; Nosek, D.; Kazakov, D.V.; Donati, M. Spitz Tumor With SQSTM1::NTRK2 Fusion: A Clinicopathological Study of 5 Cases. Am. J. Dermatopathol. 2023, 45, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Yeh, I.; Tee, M.K.; Botton, T.; Shain, A.H.; Sparatta, A.J.; Gagnon, A.; Vemula, S.S.; Garrido, M.C.; Nakamaru, K.; Isoyama, T.; et al. NTRK3 kinase fusions in Spitz tumours. J. Pathol. 2016, 240, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Knezevich, S.R.; Garnett, M.J.; Pysher, T.J.; Beckwith, J.B.; Grundy, P.E.; Sorensen, P.H. ETV6-NTRK3 gene fusions and trisomy 11 establish a histogenetic link between mesoblastic nephroma and congenital fibrosarcoma. Cancer Res. 1998, 58, 5046–5048. [Google Scholar]
- Tognon, C.; Knezevich, S.R.; Huntsman, D.; Roskelley, C.D.; Melnyk, N.; Mathers, J.A.; Becker, L.; Carneiro, F.; MacPherson, N.; Horsman, D.; et al. Expression of the ETV6-NTRK3 gene fusion as a primary event in human secretory breast carcinoma. Cancer Cell 2002, 2, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Skálová, A.; Vanecek, T.; Sima, R.; Laco, J.; Weinreb, I.; Perez-Ordonez, B.; Starek, I.; Geierova, M.; Simpson, R.H.; Passador-Santos, F.; et al. Mammary Analogue Secretory Carcinoma of Salivary Glands, Containing the ETV6-NTRK3 Fusion Gene: A Hitherto Undescribed Salivary Gland Tumor Entity. Am. J. Surg. Pathol. 2010, 34, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.A.; Taube, J.M.; Su, A.; Binder, S.W.; Kazakov, D.V.; Michal, M.; Westra, W.H. Secretory Carcinoma of the Skin Harboring ETV6 Gene Fusions: A Cutaneous Analogue to Secretory Carcinomas of the Breast and Salivary Glands. Am. J. Surg. Pathol. 2017, 41, 62–66. [Google Scholar] [CrossRef]
- Leeman-Neill, R.J.; Kelly, L.M.; Liu, P.; Brenner, A.V.; Little, M.P.; Bogdanova, T.I.; Evdokimova, V.N.; Hatch, M.; Zurnadzy, L.Y.; Nikiforova, M.N.; et al. ETV6-NTRK3 is a common chromosomal rearrangement in radiation-associated thyroid cancer. Cancer 2014, 120, 799–807. [Google Scholar] [CrossRef]
- Kralik, J.M.; Kranewitter, W.; Boesmueller, H.; Marschon, R.; Tschurtschenthaler, G.; Rumpold, H.; Wiesinger, K.; Erdel, M.; Petzer, A.L.; Webersinke, G. Characterization of a newly identified ETV6-NTRK3 fusion transcript in acute myeloid leukemia. Diagn. Pathol. 2011, 6, 19. [Google Scholar] [CrossRef]
- Forghieri, F.; Morselli, M.; Potenza, L.; Maccaferri, M.; Pedrazzi, L.; Paolini, A.; Bonacorsi, G.; Artusi, T.; Giacobbi, F.; Corradini, G.; et al. Chronic eosinophilic leukaemia with ETV6-NTRK3 fusion transcript in an elderly patient affected with pancreatic carcinoma. Eur. J. Haematol. 2011, 86, 352–355. [Google Scholar] [CrossRef]
- Yamamoto, H.; Yoshida, A.; Taguchi, K.; Kohashi, K.; Hatanaka, Y.; Yamashita, A.; Mori, D.; Oda, Y. ALK,ROS1 and NTRK3 gene rearrangements in inflammatory myofibroblastic tumours. Histopathology 2016, 69, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Diaz, A.K.; Paugh, B.S.; Rankin, S.L.; Ju, B.; Li, Y.; Zhu, X.; Qu, C.; Chen, X.; Zhang, J.; et al. The genomic landscape of diffuse intrinsic pontine glioma and pediatric non-brainstem high-grade glioma. Nat. Genet. 2014, 46, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Hechtman, J.F.; Zehir, A.; Yaeger, R.; Wang, L.; Middha, S.; Zheng, T.; Hyman, D.M.; Solit, D.; Arcila, M.E.; Borsu, L.; et al. Identification of Targetable Kinase Alterations in Patients with Colorectal Carcinoma That are Preferentially Associated with Wild-Type RAS/RAF. Mol. Cancer Res. 2016, 14, 296–301. [Google Scholar] [CrossRef] [PubMed]
- VandenBoom, T.; Quan, V.L.B.; Zhang, B.; Garfield, E.M.B.; Kong, B.Y.; Isales, M.C.; Panah, E.; Igartua, C.; Taxter, T.; Beaubier, N.; et al. Genomic Fusions in Pigmented Spindle Cell Nevus of Reed. Am. J. Surg. Pathol. 2018, 42, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- de la Fouchardière, A.; Tee, M.K.; Peternel, S.; Valdebran, M.; Pissaloux, D.; Tirode, F.; Busam, K.J.; LeBoit, P.E.; McCalmont, T.H.; Bastian, B.C.; et al. Fusion partners of NTRK3 affect subcellular localization of the fusion kinase and cytomorphology of melanocytes. Mod. Pathol. 2021, 34, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.L.; Evans, R.D.; Sivarasa, K.; Ramalho, J.S.; Briggs, D.A.; Hume, A.N. The adaptor protein melanophilin regulates dynamic myosin-Va:cargo interaction and dendrite development in melanocytes. Mol. Biol. Cell 2019, 30, 742–752. [Google Scholar] [CrossRef]
- Pastural, E.; Barrat, F.J.; Dufourcq-Lagelouse, R.; Certain, S.; Sanal, O.; Jabado, N.; Seger, R.; Griscelli, C.; Fischer, A.; Basile, G.D.S. Griscelli disease maps to chromosome 15q21 and is associated with mutations in the Myosin-Va gene. Nat. Genet. 1999, 16, 289–292. [Google Scholar] [CrossRef]
- Lee, S.; Barnhill, R.L.; Dummer, R.; Dalton, J.; Wu, J.; Pappo, A.; Bahrami, A. TERT Promoter Mutations Are Predictive of Aggressive Clinical Behavior in Patients with Spitzoid Melanocytic Neoplasms. Sci. Rep. 2015, 5, 11200. [Google Scholar] [CrossRef]
- Lee, C.Y.; Sholl, L.M.; Zhang, B.; Merkel, E.A.; Amin, S.M.; Guitart, J.; Gerami, P. Atypical Spitzoid Neoplasms in Childhood: A Molecular and Outcome Study. Am. J. Dermatopathol. 2017, 39, 181–186. [Google Scholar] [CrossRef]
- Quan, V.L.; Zhang, B.; Zhang, Y.; Mohan, L.S.; Shi, K.; Wagner, A.; Kruse, L.; Taxter, T.; Beaubier, N.; White, K.; et al. Integrating Next-Generation Sequencing with Morphology Improves Prognostic and Biologic Classification of Spitz Neoplasms. J. Investig. Dermatol. 2020, 140, 1599–1608. [Google Scholar] [CrossRef]
- Staubitz, J.I.; Schad, A.; Springer, E.; Rajalingam, K.; Lang, H.; Roth, W.; Hartmann, N.; Musholt, T.J. Novel rearrangements involving the RET gene in papillary thyroid carcinoma. Cancer Genet. 2019, 230, 13–20. [Google Scholar] [CrossRef]
- Peng, P.; Zheng, Y.; Lv, J. TBC1D32-RET: A Novel RET Oncogenic Fusion in Lung Adenocarcinoma. J. Thorac. Oncol. 2019, 14, e7–e9. [Google Scholar] [CrossRef]
- Skálová, A.; Ptáková, N.; Santana, T.; Agaimy, A.; Ihrler, S.; Uro-Coste, E.; Thompson, L.D.; Bishop, J.A.; Baněčkova, M.; Rupp, N.J.; et al. NCOA4-RET and TRIM27-RET Are Characteristic Gene Fusions in Salivary Intraductal Carcinoma, Including Invasive and Metastatic Tumors: Is “Intraductal” Correct? Am. J. Surg. Pathol. 2019, 43, 1303–1313. [Google Scholar] [CrossRef]
- Paratala, B.S.; Chung, J.H.; Williams, C.B.; Yilmazel, B.; Petrosky, W.; Williams, K.; Schrock, A.B.; Gay, L.M.; Lee, E.; Dolfi, S.C.; et al. RET rearrangements are actionable alterations in breast cancer. Nat. Commun. 2018, 9, 4821. [Google Scholar] [CrossRef]
- Kim, S.Y.; Oh, S.O.; Kim, K.; Lee, J.; Kang, S.; Kim, K.-M.; Lee, W.; Kim, S.T.; Nam, D.N. NCOA4-RET fusion in colorectal cancer: Therapeutic challenge using patient-derived tumor cell lines. J. Cancer 2018, 9, 3032–3037. [Google Scholar] [CrossRef]
- Antonescu, C.R.; Dickson, B.C.; Swanson, D.B.; Zhang, L.; Sung, Y.-S.B.; Kao, Y.-C.; Chang, W.-C.; Ran, L.; Pappo, A.; Bahrami, A.; et al. Spindle Cell Tumors with RET Gene Fusions Exhibit a Morphologic Spectrum Akin to Tumors with NTRK Gene Fusions. Am. J. Surg. Pathol. 2019, 43, 1384–1391. [Google Scholar] [CrossRef]
- Kim, D.B.; Compres, E.V.B.; Zhang, B.; Khan, A.U.M.; Sunshine, J.C.; Quan, V.L.; Gerami, P. A Series of RET Fusion Spitz Neoplasms with Plaque-Like Silhouette and Dyscohesive Nesting of Epithelioid Melanocytes. Am. J. Dermatopathol. 2021, 43, 243–251. [Google Scholar] [CrossRef]
- Yeh, I.; Botton, T.; Talevich, E.; Shain, A.H.; Sparatta, A.J.; de la Fouchardiere, A.; Mully, T.W.; North, J.P.; Garrido, M.C.; Gagnon, A.; et al. Activating MET kinase rearrangements in melanoma and Spitz tumours. Nat. Commun. 2015, 6, 7174. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.S.; Wang, K.; Chmielecki, J.; Gay, L.; Johnson, A.; Chudnovsky, J.; Yelensky, R.; Lipson, D.; Ali, S.M.; Elvin, J.A.; et al. The distribution of BRAF gene fusions in solid tumors and response to targeted therapy. Int. J. Cancer 2015, 138, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.B.; Benton, S.B.; Zhang, B.; Olivares, S.B.; Asadbeigi, S.; Busam, K.; Gerami, P. Benign and Intermediate-grade Melanocytic Tumors with BRAF Mutations and Spitzoid Morphology: A Subset of Melanocytic Neoplasms Distinct from Melanoma. Am. J. Surg. Pathol. 2021, 46, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.F.; Milante, R.; Pissaloux, D.; Tirode, F.; Bastian, B.C.; de la Fouchardière, A.; Yeh, I. Spectrum of Melanocytic Tumors Harboring BRAF Gene Fusions: 58 Cases with Histomorphologic and Genetic Correlations. Mod. Pathol. 2023, 36, 100149. [Google Scholar] [CrossRef] [PubMed]
- Donati, M.; Kastnerova, L.; Ptakova, N.; Michal, M.; Kazakov, D.V. Polypoid Atypical Spitz Tumor with a Fibrosclerotic Stroma, CLIP2-BRAF Fusion, and Homozygous Loss of 9p21. Am. J. Dermatopathol. 2020, 42, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Khan, A.U.; Compres, E.V.; Zhang, B.; Sunshine, J.C.; Quan, V.L.; Gerami, P. BRAF fusion Spitz neoplasms; clinical morphological, and genomic findings in six cases. J. Cutan. Pathol. 2020, 47, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Perron, E.; Pissaloux, D.; Neub, A.; Hohl, D.; Tartar, M.D.; Mortier, L.; Alberti, L.; de la Fouchardiere, A. Unclassified sclerosing malignant melanomas with AKAP9-BRAF gene fusion: A report of two cases and review of BRAF fusions in melanocytic tumors. Virchows Arch. 2018, 472, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.F.; Bastian, B.C.; Maguiness, S.; Giubellino, A.; Vemula, S.S.; McCalmont, T.H.; Yeh, I. Multiple desmoplastic Spitz nevi with BRAF fusions in a patient with ring chromosome 7 syndrome. Pigment. Cell Melanoma Res. 2021, 34, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Donati, M.; Nosek, D.; Olivares, S.; Lemahieu, J.; Loontiens, S.; Mansour, B.; Gerami, P.; Kazakov, D.V. Spitz tumor with RAF1 fusion: A report of 3 cases. Ann. Diagn. Pathol. 2023, 67, 152215. [Google Scholar] [CrossRef]
- LeBlanc, R.E.; Lefferts, J.A.; Baker, M.L.; Linos, K.D. Novel LRRFIP2-RAF1 fusion identified in an acral melanoma: A review of the literature on melanocytic proliferations with RAF1 fusions and the potential therapeutic implications. J. Cutan. Pathol. 2020, 47, 1181–1186. [Google Scholar] [CrossRef]
- Lavoie, H.; Therrien, M. Regulation of RAF protein kinases in ERK signalling. Nat. Rev. Mol. Cell Biol. 2015, 16, 281–298. [Google Scholar] [CrossRef]
- Williams, E.A.; Shah, N.; Montesion, M.; Sharaf, R.; Pavlick, D.C.; Sokol, E.S.; Alexander, B.M.; Venstrom, J.M.; Elvin, J.A.; Ross, J.S.; et al. Melanomas with activating RAF1 fusions: Clinical, histopathologic, and molecular profiles. Mod. Pathol. 2020, 33, 1466–1474. [Google Scholar] [CrossRef]
- Donati, M.; Martinek, P.; Kastnerova, L.; Persichetti, P.; Michal, M.; Kazakov, D.V. RAF1 Gene Fusions as a Possible Driver Mechanism in Rare BAP1-Inactivated Melanocytic Tumors: A Report of 2 Cases. Am. J. Dermatopathol. 2020, 42, 961–966. [Google Scholar] [CrossRef]
- da Silva, V.M.; Martinez-Barrios, E.; Tell-Marti, G.; Dabad, M.; Carrera, C.; Aguilera, P.; Brualla, D.; Esteve-Codina, A.; Vicente, A.; Puig, S.; et al. Genetic Abnormalities in Large to Giant Congenital Nevi: Beyond NRAS Mutations. J. Investig. Dermatol. 2019, 139, 900–908. [Google Scholar] [CrossRef]
- Quan, V.L.; Zhang, B.; Mohan, L.S.; Shi, K.; Isales, M.C.; Panah, E.; Taxter, T.J.; Beaubier, N.; White, K.; Gerami, P. Activating Structural Alterations in MAPK Genes Are Distinct Genetic Drivers in a Unique Subgroup of Spitzoid Neoplasms. Am. J. Surg. Pathol. 2019, 43, 538–548. [Google Scholar] [CrossRef]
- Houlier, A.; Pissaloux, D.; Masse, I.; Tirode, F.; Karanian, M.; Pincus, L.B.; McCalmont, T.H.; LeBoit, P.E.; Bastian, B.C.; Yeh, I.; et al. Melanocytic tumors with MAP3K8 fusions: Report of 33 cases with morphological-genetic correlations. Mod. Pathol. 2020, 33, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Newman, S.; Pappo, A.; Raimondi, S.; Zhang, J.; Barnhill, R.; Bahrami, A. Pathologic Characteristics of Spitz Melanoma with MAP3K8 Fusion or Truncation in a Pediatric Cohort. Am. J. Surg. Pathol. 2019, 43, 1631–1637. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, J.; Higashi, T.; Mukai, H.; Ohuchi, T.; Kakunaga, T. Structure and transforming potential of the human cot oncogene encoding a putative protein kinase. Mol. Cell. Biol. 1991, 11, 4088–4096. [Google Scholar] [PubMed]
- Lee, J.-H.; Lee, J.-H.; Lee, S.H.; Do, S.-I.; Cho, S.-D.; Forslund, O.; Inn, K.-S.; Lee, J.-S.; Deng, F.-M.; Melamed, J.; et al. TPL2 Is an Oncogenic Driver in Keratocanthoma and Squamous Cell Carcinoma. Cancer Res. 2016, 76, 6712–6722. [Google Scholar] [CrossRef] [PubMed]
- Gruosso, T.; Garnier, C.; Abelanet, S.; Kieffer, Y.; Lemesre, V.; Bellanger, D.; Mechta-Grigoriou, F. MAP3K8/TPL-2/COT is a potential predictive marker for MEK inhibitor treatment in high-grade serous ovarian carcinomas. Nat. Commun. 2015, 6, 8583. [Google Scholar] [CrossRef]


{kind=link}
| Type of Kinase Fusions (Most Frequently Reported) | Clinical Features (Most Frequent, Rarely Present) | Histopathology (Usually, Rarely) | Molecular Features (Most Frequent Gene Patterns and Rare Gene Patterns) |
|---|---|---|---|
| ALK-rearranged |
|
| TPM3 DCTN1 GTF3C2, TPR, CLIP1 and NPM1 |
| ROS1-rearranged |
|
| PWWP2A, TPM3, PPFIBP1 MYH9, CAPRINI1, and MYO5A |
| NTRK-rearranged |
|
| NTRK1: LMNA, TPM3, TP53, and KHDRBS1 |
| NTRK2: TGF and SQSTM1 | ||
| NTRK3: ETV6, MYO5A, and MYH9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daruish, M.; Ambrogio, F.; Colagrande, A.; Marzullo, A.; Alaggio, R.; Trilli, I.; Ingravallo, G.; Cazzato, G. Kinase Fusions in Spitz Melanocytic Tumors: The Past, the Present, and the Future. Dermatopathology 2024, 11, 112-123. https://doi.org/10.3390/dermatopathology11010010
Daruish M, Ambrogio F, Colagrande A, Marzullo A, Alaggio R, Trilli I, Ingravallo G, Cazzato G. Kinase Fusions in Spitz Melanocytic Tumors: The Past, the Present, and the Future. Dermatopathology. 2024; 11(1):112-123. https://doi.org/10.3390/dermatopathology11010010
Chicago/Turabian StyleDaruish, Maged, Francesca Ambrogio, Anna Colagrande, Andrea Marzullo, Rita Alaggio, Irma Trilli, Giuseppe Ingravallo, and Gerardo Cazzato. 2024. "Kinase Fusions in Spitz Melanocytic Tumors: The Past, the Present, and the Future" Dermatopathology 11, no. 1: 112-123. https://doi.org/10.3390/dermatopathology11010010