
The Effect of Bacillus coagulans Induced Interactions among Intestinal Bacteria, Metabolites, and Inflammatory Molecules in Improving Natural Skin Aging
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Measurement of TEWL and Skin Hydration Levels
2.3. Evaluation of Wrinkles
2.4. Preparation and Staining of the Dorsal Skin Sections
2.5. Measuring the Plasma Levels of Hyaluronic Acid, IL-6, Tumor Necrosis Factor-α, IL-10, and Skin Levels of Transforming Growth Factor-β, Hyaluronic Acid, Ionized Calcium-Binding Adapter Molecule 1, CC-Chemokine Receptor 7, and Cluster of Differentiation 163
2.6. Microbiome Analysis of Intestinal Flora
2.7. Stool Metabolome Analysis
2.8. Statistical Analyses
3. Results
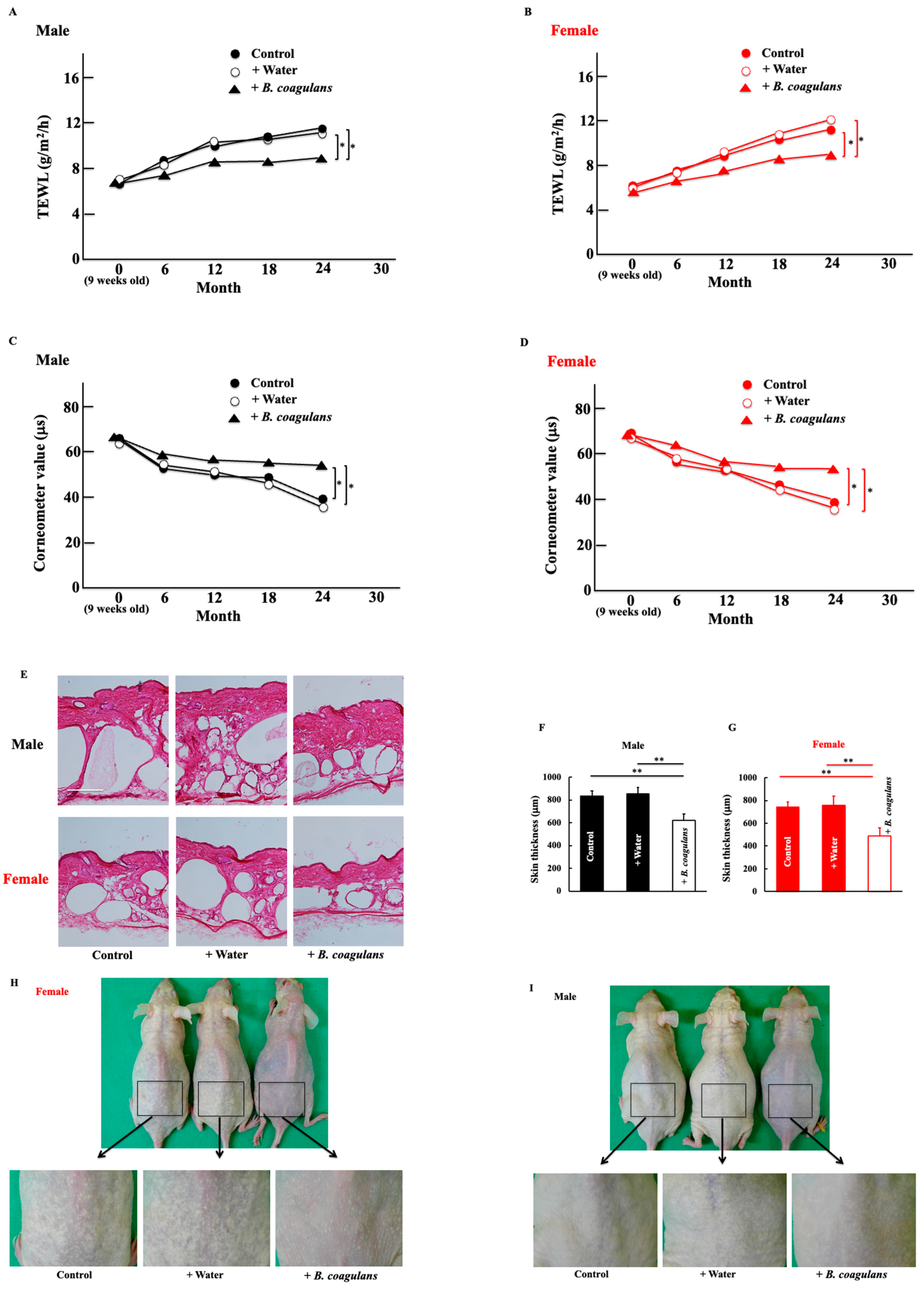
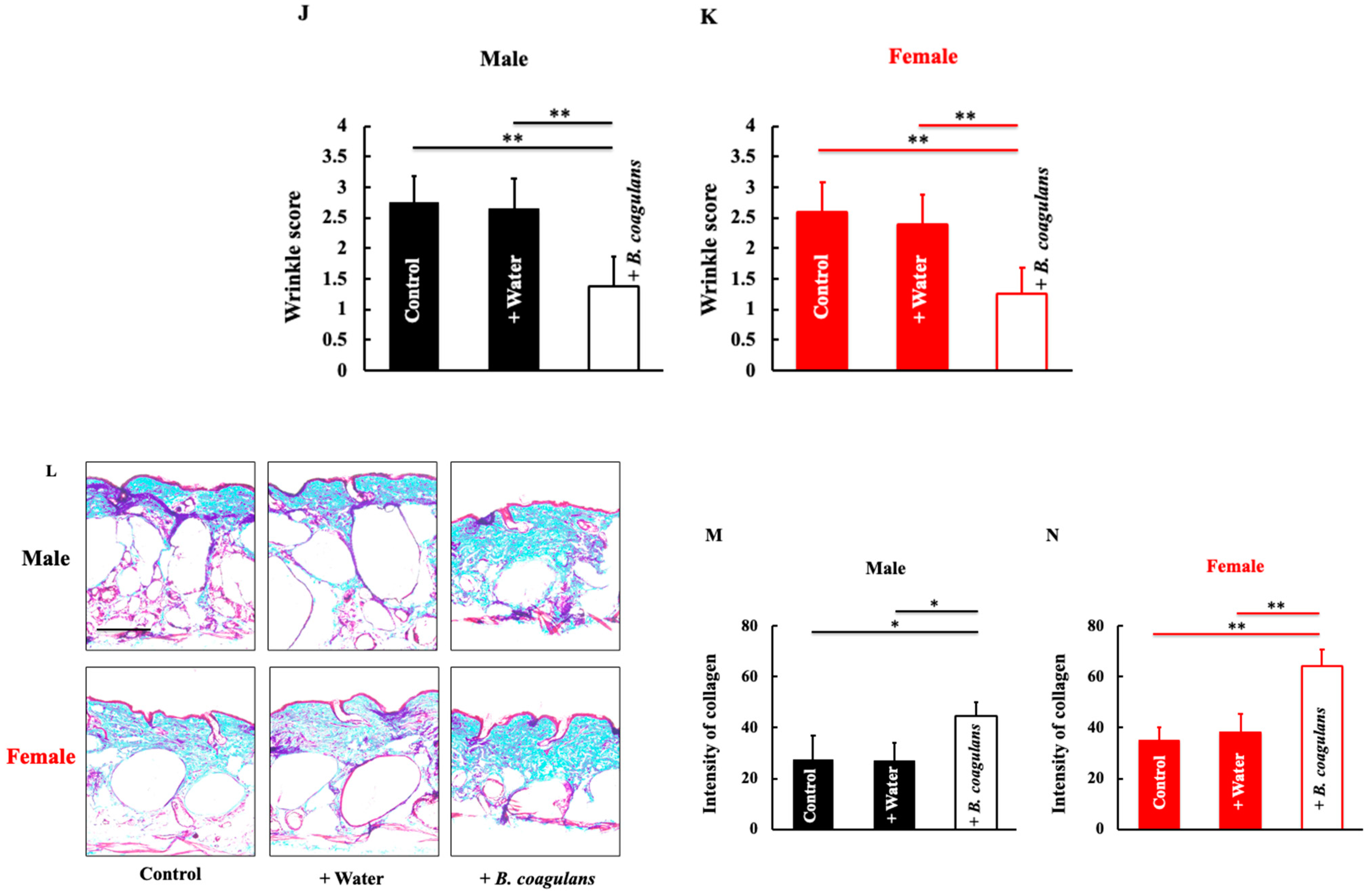
3.1. Effects of B. coagulans Administration on TEWL, Skin Hydration, and Skin Conditions in Aging Mice
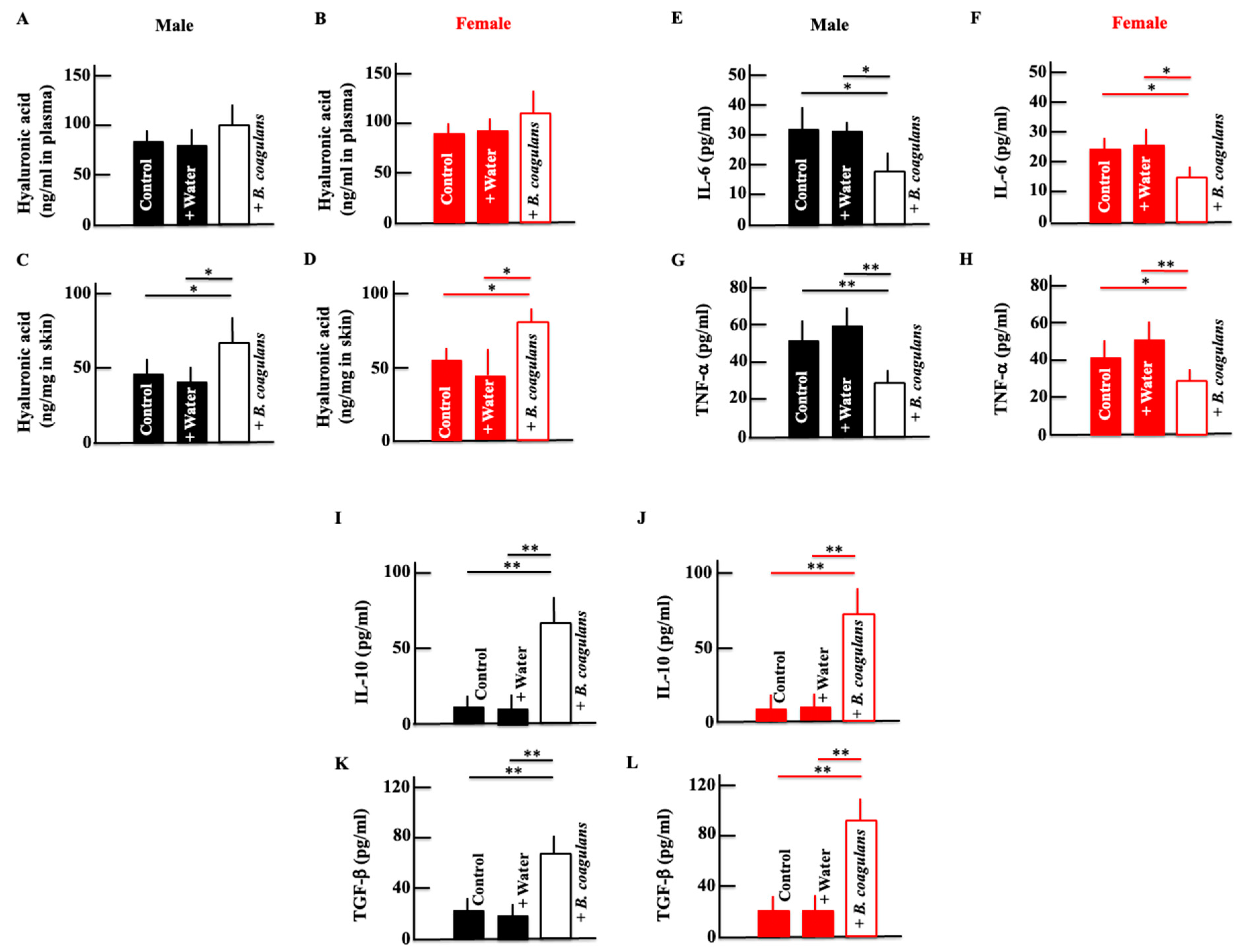
3.2. Effects of B. coagulans Administration on the Levels of Hyaluronic Acid, IL-6, TNF-α, IL-10, and TGF-β in Aging Mice
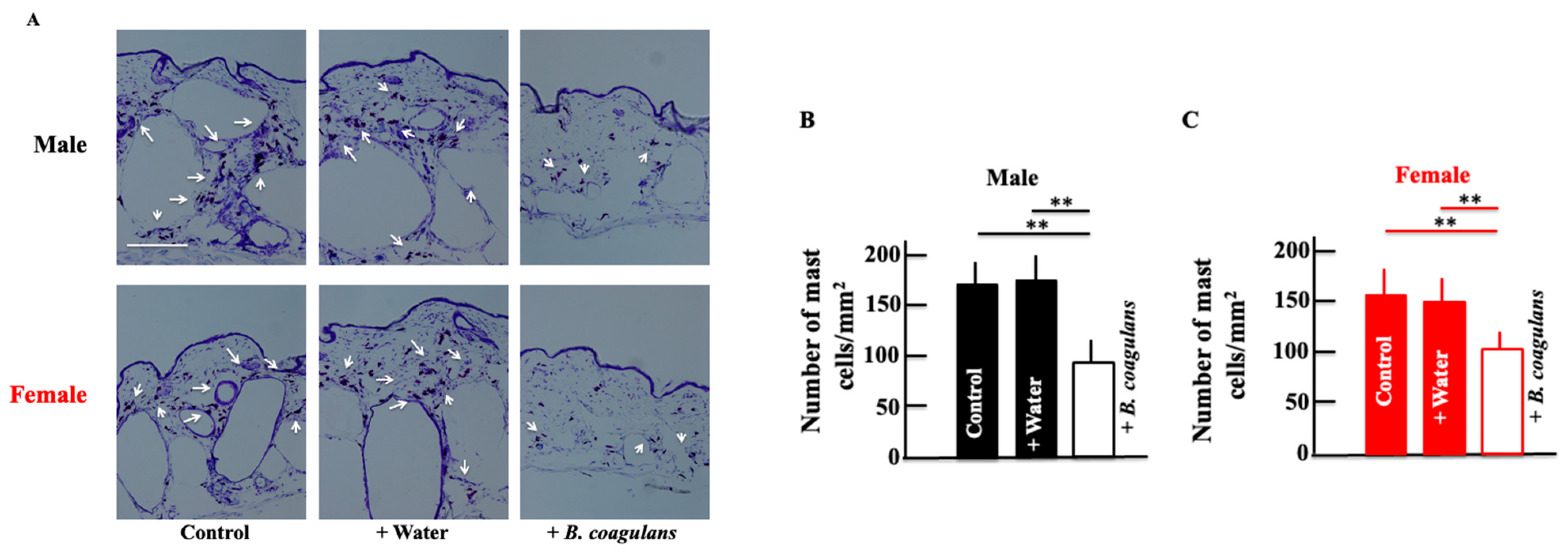
3.3. Effects of B. coagulans Administration on the Expression of Mast Cells in Aging Mice
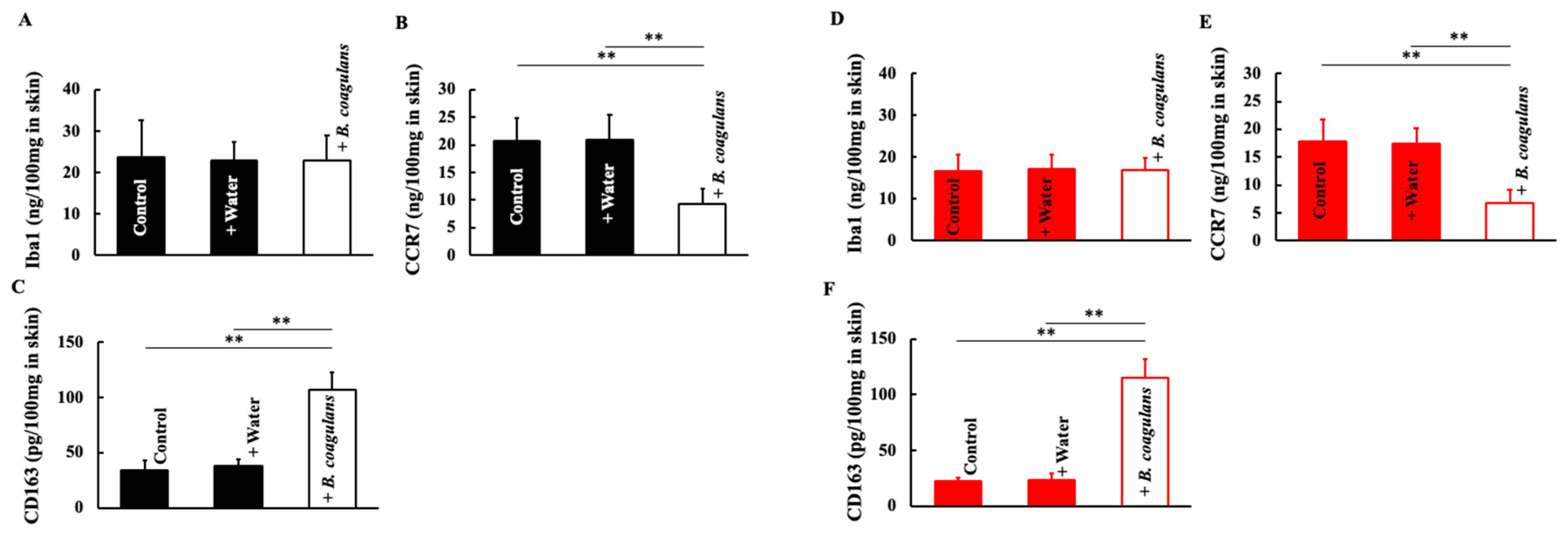
3.4. Effects of B. coagulans Administration on Macrophage Levels in Aging Mice
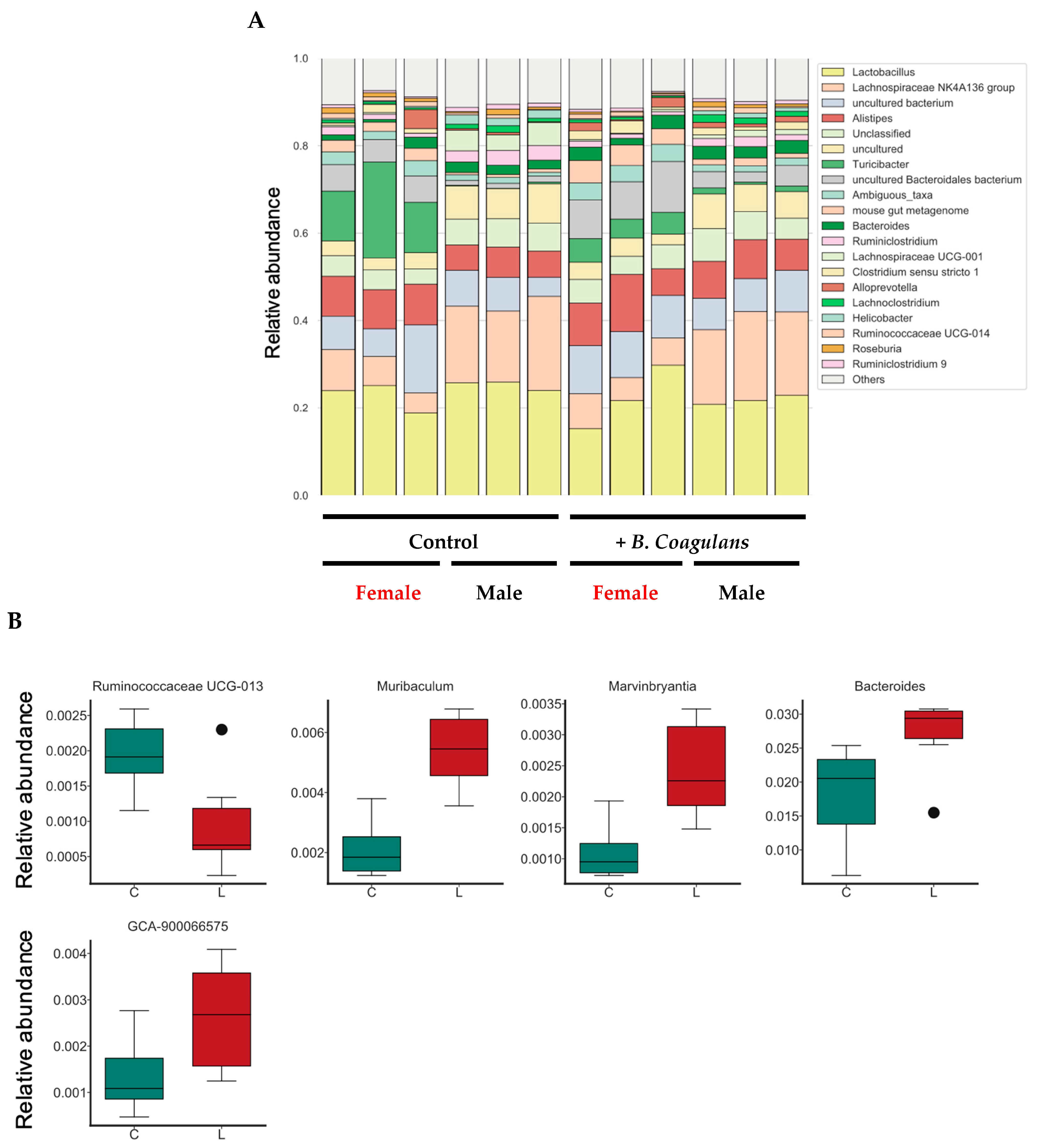
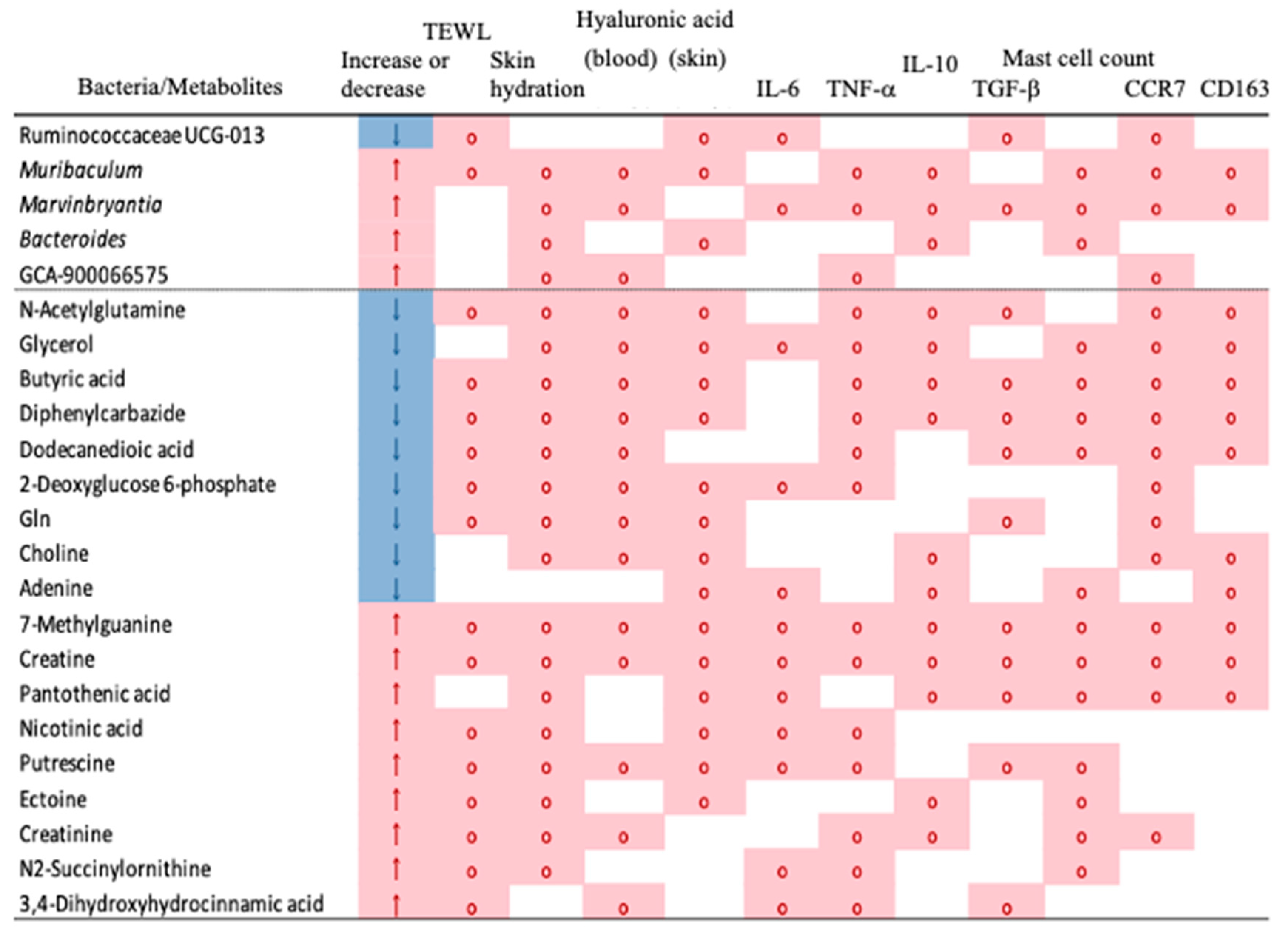
3.5. Effects of B. coagulans Administration on the Gut Microbiota Profile in Aging Mice
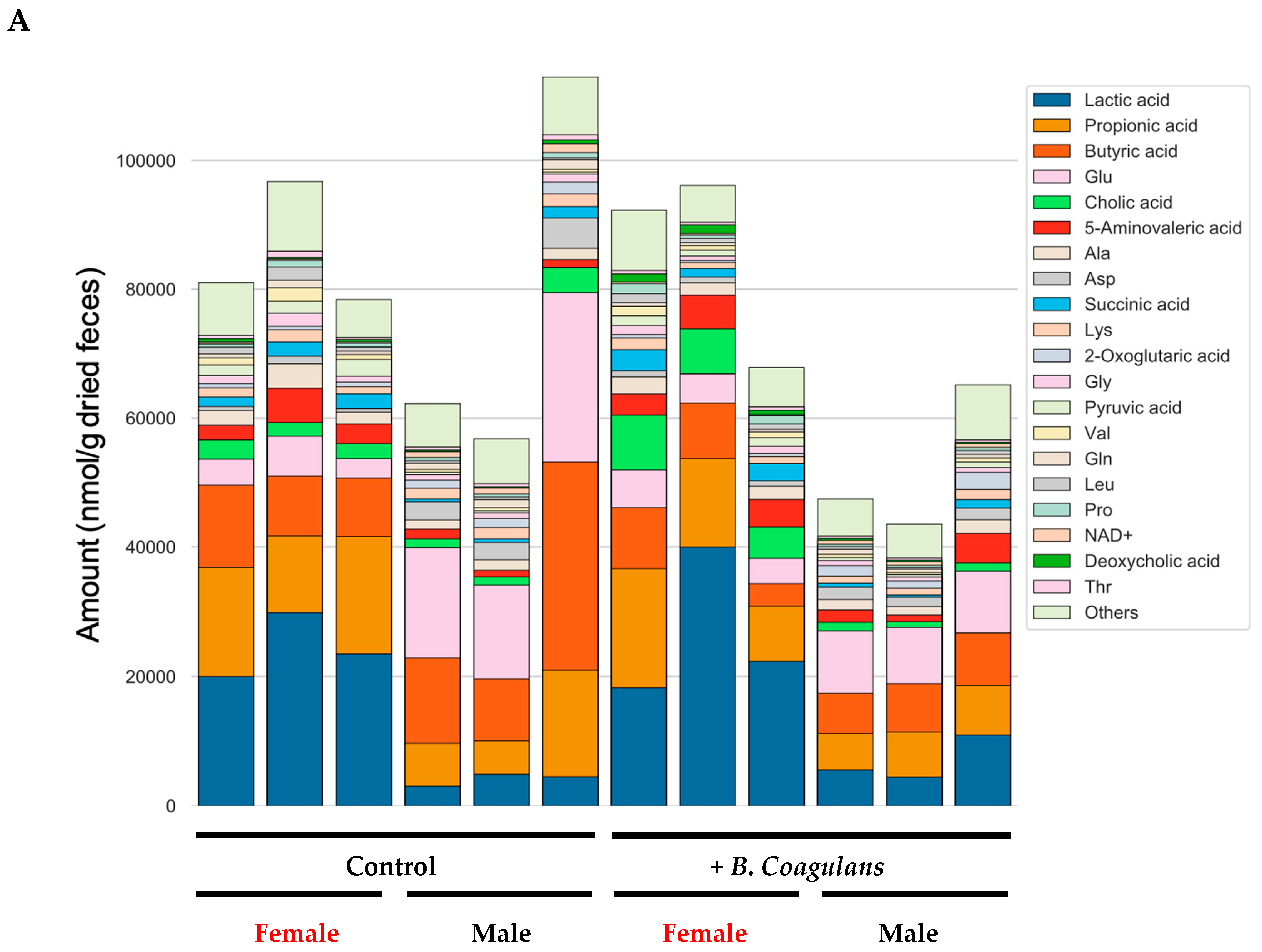
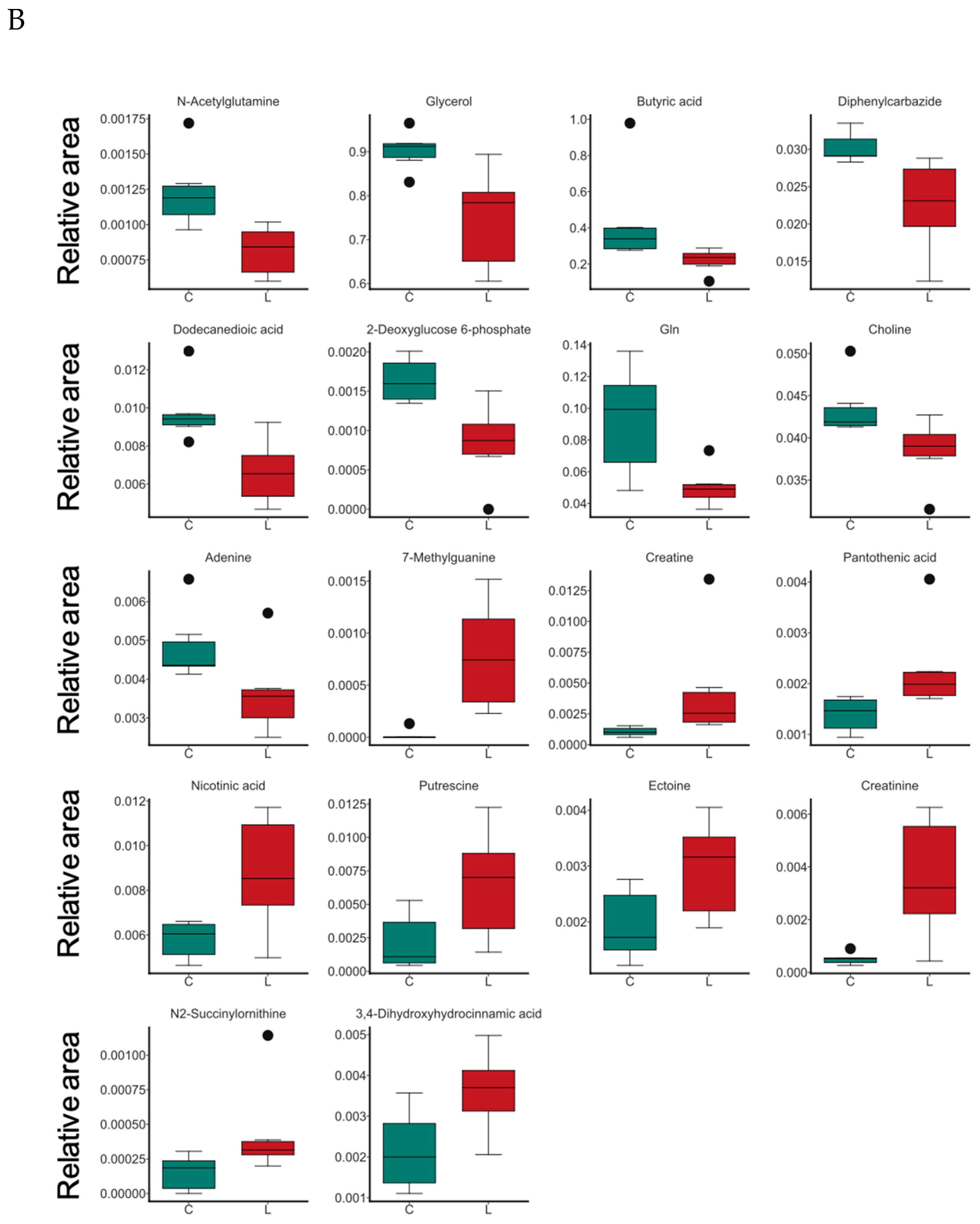
3.6. Effects of B. coagulans Administration on Metabolites in Aging Mice
4. Discussion
- 1.
- Increased absorption of hyaluronic acid from the intestine
- 2.
- Induction of IL-10-producing cells
- 3.
- Suppression of intestinal permeability
- 4.
- Increased pantothenic acid levels
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J. Introduction to skin aging. J Tissue Viability 2017, 26, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Yonei, Y.; Ichihashi, M.; Takabe, W. Age-related diseases of the skin and anti-aging medicine. Nihon Rinsho. Jap. J. Clin. Med. 2016, 74, 1541–1547. [Google Scholar]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Intrinsic and extrinsic factors in skin aging: A review. Int. J. Cosmet. Sci. 2008, 30, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Tatsumi, K.; Shinozuka, N.; Nakayama, K.; Sekiya, N.; Kuriyama, T.; Fukuchi, Y. Hochuekkito improves systemic inflammation and nutritional status in elderly patients with chronic obstructive pulmonary disease. J. Am. Geriatr. Soc. 2009, 57, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Kaya, G.; Saurat, J.H. Dermatoporosis: A chronic cutaneous indufficiency/fragility syndrome. Dermatology 2007, 215, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, K.; Orita, K.; Yamate, Y.; Kobayashi, H. Role of Momordica charantia in preventing the natural aging process of skin and sexual organs in mice. Dermatol. Ther. 2022, 33, e14243. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, K.; Yamate, Y.; Sugiyama, D.; Matsuda, K.; Iizuka, Y.; Yamaguchi, T. Ameliorative effect of tranexamic acid on physiological skin aging and its sex difference in mice. Arch. Dermatol. Res. 2019, 311, 545–553. [Google Scholar] [CrossRef]
- Curtis, H.J. The biology of aging. BNL 854, (T-340). Tech. Rep. Brookhaven Natl. Lab. 1964, 18, 1–11. [Google Scholar]
- Gershon, H.; Gershon, D. Detection of inactive enzyme molecules in aging organisms. Nature 1970, 227, 1214–1217. [Google Scholar] [CrossRef]
- Harman, D. Aging; a theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Takada, T.; Shimizu, K.; Moriyama, K.; Kawakami, K.; Hirano, K.; Okajima, O.; Nomoto, K. Effects of a probiotic fermented milk beverage containing Lactobacillus casei strain Shirota on defecation frequency, intestinal microbiota, and the intestinal environment of healthy individuals with soft stools. J. Biosci. Bioeng. 2010, 110, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, J.; Goretzki, A.; Meier, C.; Wolfheimer, S.; Lin, Y.J.; Rainer, H.; Krause, M.; Wedel, S.; Spies, G.; Fuhrer, F.; et al. Modulation of dendritic cell metabolism by an MPLA-adjuvanted allergen product for specific immunotherapy. Front Immunol. 2022, 13, 916491. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Park, S.J.; Kim, J.; Hong, U.; Lee, J. Effect of lactic acid strains isolated from kimchi on atopic dermatitis and immunomodulation in NC/Nga mice. Prev. Nutr. Food Sci. 2021, 26, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Yamashita, M.; Takeuchi, N.; Kabuki, T.; Hattori, M.; Yoshida, T. Lactobacillus helveticas SBT2171 alleviates allergic symptoms in a murine model for pollen allergy. Biosci. Biotechnol. Biochem. 2019, 83, 2298–2306. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kang, J.; Choi, S.; Park, H.; Hwang, E.; Kang, Y.G.; Kim, A.R.; Holzapfel, W.; Ji, Y. Cholesterol-lowering effect of Lactobacillus rhamnosus BFE5264 and its influence on the gut microbiome and propionate level in a murine model. PLoS ONE 2018, 13, e0203150. [Google Scholar] [CrossRef]
- Lau, A.S.; Liong, M.T. Lactic acid bacteria and bifidobacterial-inhibited Staphylococcus epidermadis. Wounds 2014, 26, 121–131. [Google Scholar] [PubMed]
- Jeong, J.H.; Lee, C.Y.; Chung, D.K. Probiotic lactic acid bacteria and skin health. Crit. Rev. Food Sci. Nutr. 2016, 56, 2331–2337. [Google Scholar] [CrossRef]
- Morifuji, M.; Kitade, M.; Fukasawa, T.; Yamaji, T.; Ichihashi, M. Exopolysaccharides isolated from milk fermented with lactic acid bacteria prevent ultraviolet-induced skin damage in hairless mice. Int. J. Mol. Sci. 2017, 18, 146. [Google Scholar] [CrossRef]
- Chen, H.; Li, Y.; Xie, X.; Chen, M.; Xue, L.; Wang, J.; Ye, Q.; Wu, S.; Yang, R.; Zhao, J.; et al. Exploration on the molecular machanisms underlying the anti-photoaging effect of Limosilactobacillus fermentum XJC60. Front. Cell. Infect. Microbiol. 2022, 12, 838060. [Google Scholar]
- Fu, H.; Zhang, Y.; An, Q.; Wang, D.; You, S.; Zhao, D.; Zhang, J.; Wang, C.; Li, M. Anti-phatoaging effect of Rhodiola resea fermented by Lactobacillus plantarum on UVA-damaged fibroblasts. Nutrients 2022, 14, 2324. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, K.; Kubo, S.; Tsuji, K.; Sugiyama, D.; Iizuk, Y.; Hamana, H. Bacillus. coagulans (species of lactic acid-forming Bacillus bacteria) ameliorates azoxymethane and dextran sodium sulfate-induced colon cancer in mice. J. Funct. Foods 2023, 100, 10406. [Google Scholar] [CrossRef]
- Yamaguchi, Y. Periostin in skin tissue and skin-related disease. Allergol. Int. 2014, 63, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Amamoto, R.; Shimamoto, K.; Park, S.; Matsumoto, H.; Shimizu, K.; Katto, M.; Tsuji, H.; Matsubara, S.; Shephard, R.J.; Aoyagi, Y. Yearly changes in the composition of gut microbiota in the elderly, and the effect of lactobacilli intake on these changes. Sci. Rep. 2021, 11, 12765. [Google Scholar] [CrossRef] [PubMed]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The gut microbiome as a major regulator of the gut-skin axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Erturk-Hasdemir, D.; Ochoa-Reparaz, J.; Reinecker, H.C.; Kasper, D.L. Plasmacytoid dendritic cells mediate anti-inflammatory responses to a gut commensal molecule via both innate and adaptive mechanisms. Cell Host Microbe. 2014, 15, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Li, Z.; Liu, T.; Yang, N.; Li, Y.; He, J.; Qian, M.; Kuang, Z.; Zhang, W.; Ni, C.; et al. Prebiotics regulation of intestinal microbiota attenuates cognitive dysfunction induced by surgery stimulation in APP/PS1 mice. Aging Dis. 2020, 11, 1029–1045. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Hiramoto, K.; Koyama, M.; Ooi, K. Impairment of skin barrier function via cholinergic signal transduction in a dextran sulphate sodium-induced colitis mouse model. Exp. Dermatol. 2015, 24, 779–784. [Google Scholar] [CrossRef]
- Bissett, D.L.; Hannon, D.P.; Orr, T.V. An animal model of solar-aged skin: Histological, physical, and visible changes in UV-irradiated hair- less mouse skin. Photochem. Photobiol. 1987, 3, 367–378. [Google Scholar] [CrossRef]
- Murakami, S.; Goto, Y.; Ito, K.; Hayasaka, S.; Kurihara, S.; Soga, T.; Tomita, M.; Fukuda, S. The consumption of bicarbonate-rich mineral water improves glycemic control. Evid. Based Complement. Alternat. Med. 2015, 2015, 824395. [Google Scholar] [CrossRef]
- Hiramoto, K.; Sugiyama, D.; Takahashi, Y.; Mafune, E. The amelioration effect of tranexamic acid in wrinkles induced by skin dryness. Biomed. Pharmacother. 2016, 80, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Takamori, K. Peripheral itch sensitization in atopic dermatitis. Allergol. Int. 2022, 71, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Elbasiony, E.; Mittal, S.K.; Foulsham, W.; Cho, W.K.; Chauhan, S.K. Epithelium derived IL-33 activates mast cells to initiate neutrophil recruitment following corneal injury. Ocul. Surf. 2020, 18, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Veerappan, A.; O’Connor, N.J.; Brazin, J.; Reid, A.C.; Jung, A.; McGee, D.; Summers, B.; Branch-Elliman, D.; Stiles, B.; Worgall, S.; et al. Mast cells: A pivotal role in pulmonary fibrosis. DNA Cell Biol. 2013, 32, 206–218. [Google Scholar] [CrossRef]
- Metz, M.; Grimbaldeston, M.A.; Nakae, S.; Piliponsky, A.M.; Tsai, M.; Galli, S.J. Mast cells in the promotion and limitation of chronic inflammation. Immunol. Rev. 2007, 217, 304–328. [Google Scholar] [CrossRef] [PubMed]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast cell: A multi-functional master cell. Front Immunol. 2016, 6, 620. [Google Scholar] [CrossRef] [PubMed]
- Mendeley, K.S.; Pedersen, M.; Bruunsgaard, H. Inflammatory mediators in the elderly. Exp. Gerontol. 2004, 39, 687–699. [Google Scholar]
- Elbasony, E.; Cho, W.J.; Singh, A.; Mittal, S.K.; Zoukhri, D.; Chauhan, S.K. Increased activity of lacrimal gland mast cells are associated with corneal epitheliopathy in aged mice. NPJ Aging 2023, 9, 2. [Google Scholar] [CrossRef]
- Lin, S.; Huang, H.; Ling, M.; Zhang, C.; Yang, F.; Fan, Y. Development and validation of a novel diagnostic model for musculoskeletal aging (sarcopenia) based of cuproptosis-related genes associated with immunity. Am. J. Transl. Res. 2022, 14, 8523–8538. [Google Scholar]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Yoshimoto, S.; Mitsuyama, E.; Yoshida, K.; Odamaki, T.; Xiao, J.Z. Enriched metabolites that potentially promote age-associated diseases in subjects with an elderly-type gut microbiota. Gut Microbes. 2021, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, D.; Kusama, M.; Onda, M.; Nakahata, N. The effect of pantothenic acid deficiency on keratinocyte growth factor and collagen in fibroblasts. J. Pharmacol. Sci. 2011, 115, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, K.; Hosomi, K.; Sawane, K.; Kunisawa, J. Methabolism of dietary and microbial vitamin B family in the regulation of host immunity. Front. Nutr. 2019, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Timmons, J.; Chang, E.T.; Wang, J.Y.; Rao, J.N. Polyamines and gut mucosal homeostasis. J. Gastrointest. Dig. Syst. 2012, 2 (Suppl. 7), 001. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Kurihara, S.; Takahashi, D.; Ohashi, W.; Nakamura, Y.; Kimura, S.; Onuki, M.; Kume, A.; Sasazawa, Y.; Hase, K.; et al. Symbiotic polyamine metabolism regulates epithelial proliferation and macrophage differentiation in the colon. Nat. Commun. 2021, 12, 2105. [Google Scholar] [CrossRef] [PubMed]
- Oe, M.; Mitsugi, K.; Odanaka, W.; Yoshida, H.; Matsuoka, R.; Seino, S.; Kanemitsu, T.; Masuda, Y. Dietary hyaluronic acid migrates into the skin of rats. Sci. World J. 2014, 2014, 378024. [Google Scholar] [CrossRef]
- Kawai, K.; Kamochi, R.; Oiki, S.; Murata, K.; Hashimoto, W. Probiotics in human gut microbiota can degrade host glycosaminoglycans. Sci. Rep. 2018, 8, 10674. [Google Scholar] [CrossRef]
- Levkovich, T.; Poutahidis, T.; Smillie, C.; Varian, B.J.; Ibrahim, Y.M.; Lakritz, J.R.; Alm, E.J.; Erdman, S.E. Probiotic bacteria induce a “glow of health”. PLOS ONE 2013, 8, e53867. [Google Scholar] [CrossRef]
- Iizuka, T.; Sakai, H.; Moriyama, H.; Suto, N.; Nagai, M.; Bagchi, D. Vasorelaxant effects of forsythide isolated from the leaves of Forsythia viridissima on NE-induced aortal contraction. Phytomedicine 2009, 16, 386–390. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hiramoto, K.; Kubo, S.; Tsuji, K.; Sugiyama, D.; Iizuka, Y.; Hamano, H. The Effect of Bacillus coagulans Induced Interactions among Intestinal Bacteria, Metabolites, and Inflammatory Molecules in Improving Natural Skin Aging. Dermatopathology 2023, 10, 287-302. https://doi.org/10.3390/dermatopathology10040037
Hiramoto K, Kubo S, Tsuji K, Sugiyama D, Iizuka Y, Hamano H. The Effect of Bacillus coagulans Induced Interactions among Intestinal Bacteria, Metabolites, and Inflammatory Molecules in Improving Natural Skin Aging. Dermatopathology. 2023; 10(4):287-302. https://doi.org/10.3390/dermatopathology10040037
Chicago/Turabian StyleHiramoto, Keiichi, Sayaka Kubo, Keiko Tsuji, Daijiro Sugiyama, Yasutaka Iizuka, and Hideo Hamano. 2023. "The Effect of Bacillus coagulans Induced Interactions among Intestinal Bacteria, Metabolites, and Inflammatory Molecules in Improving Natural Skin Aging" Dermatopathology 10, no. 4: 287-302. https://doi.org/10.3390/dermatopathology10040037