
Ultrasound-Assisted Transglutaminase Catalysis of the Cross-Linking and Microstructure of αs-Casein, β-Casein and κ-Casein

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Milk Protein Samples
2.2. Preparation of Milk Samples with/without TGase and UT
2.3. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis
2.4. Gel Staining and Image Analysis
2.5. Particle Size Analysis of Polymerized Individual Caseins with TGase and UT
2.6. Transmission Electron Microscopy (TEM) Analysis
2.7. Statistical Analysis
3. Results and Discussion
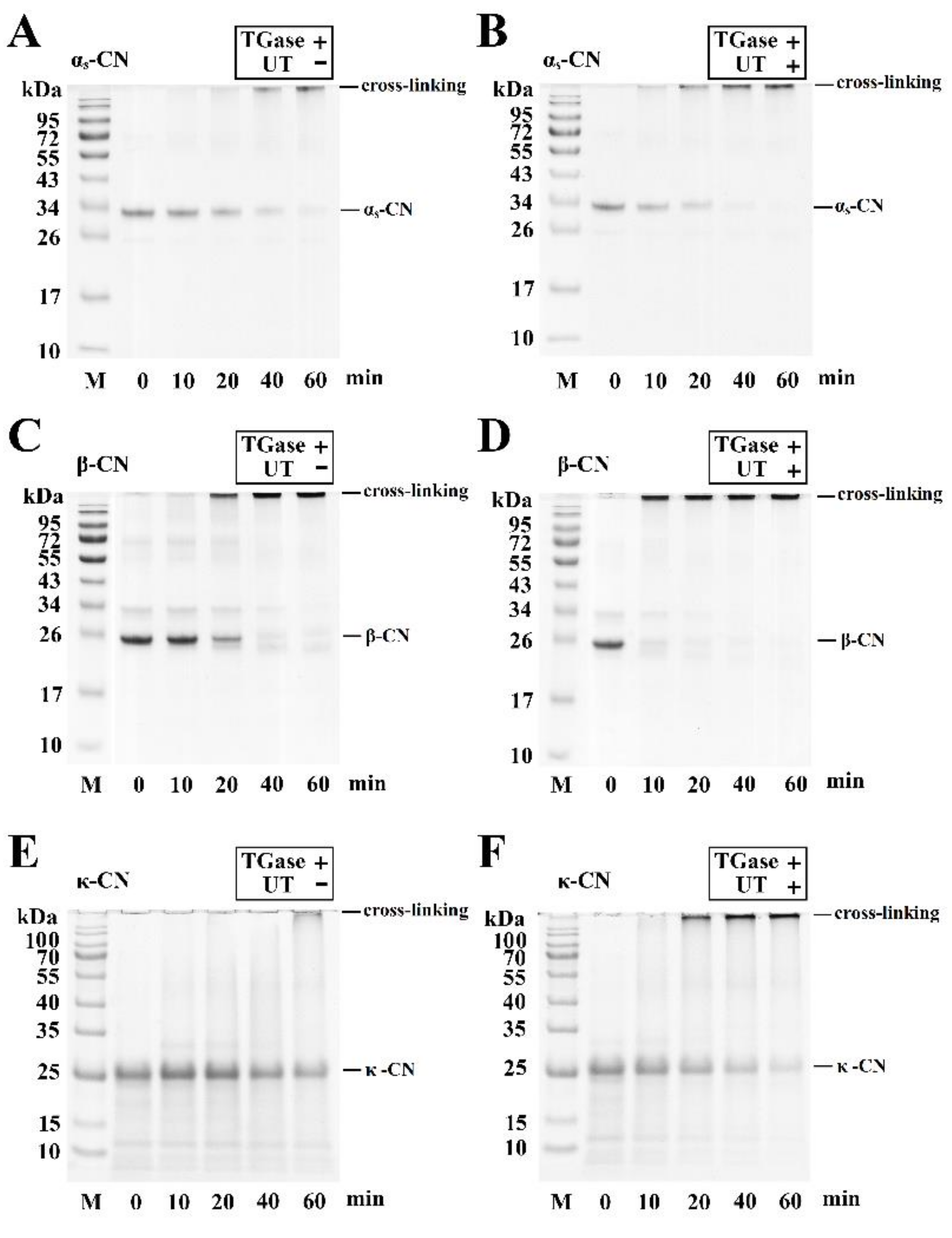
3.1. SDS-PAGE Analysis of TGase and UT on the Cross-Linking of Milk Proteins
3.2. SDS-PAGE Analysis of TGase and UT on the Cross-Linking of Individual Milk Proteins
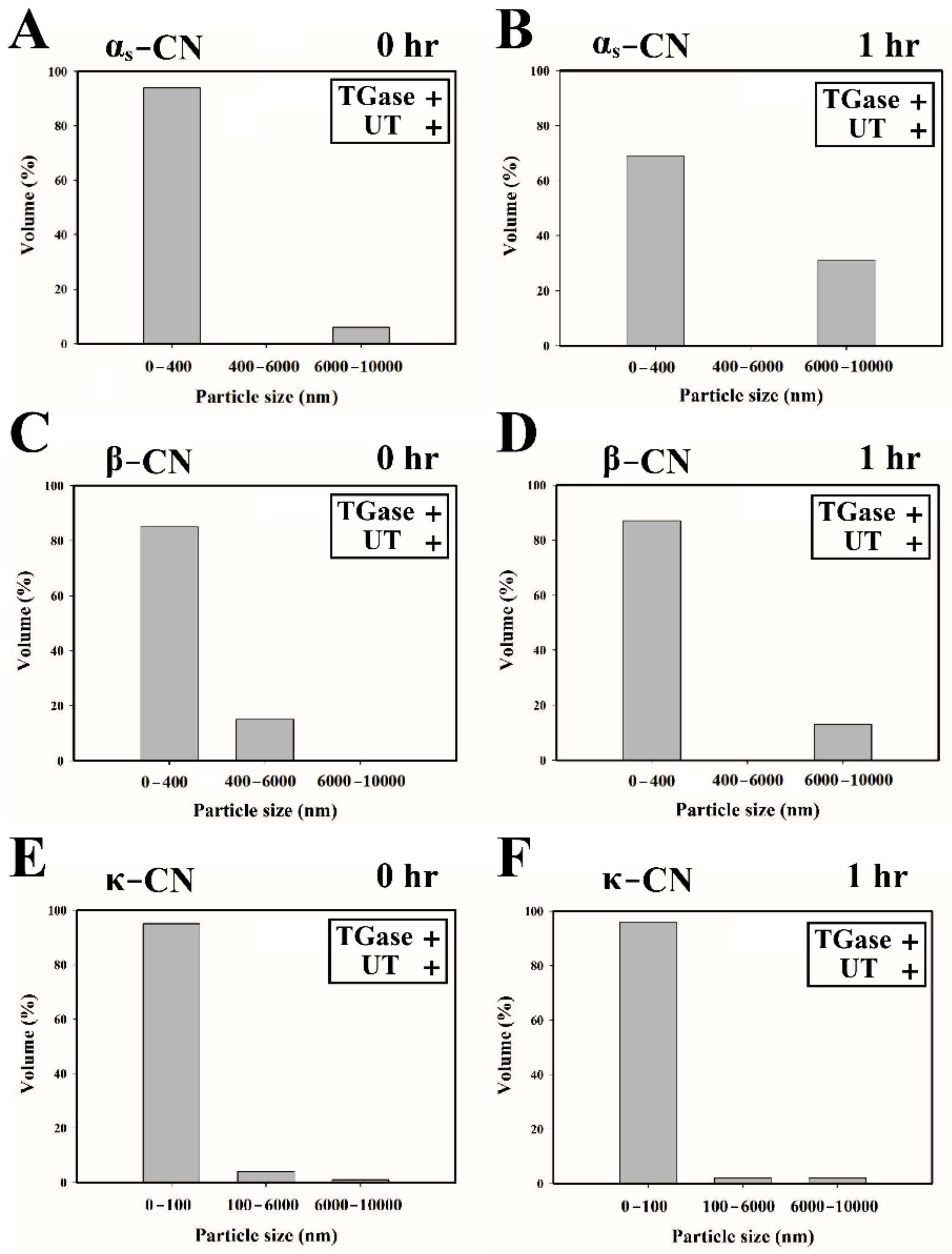
3.3. Particle Size Analysis of Individual Caseins Treated with TGase and UT
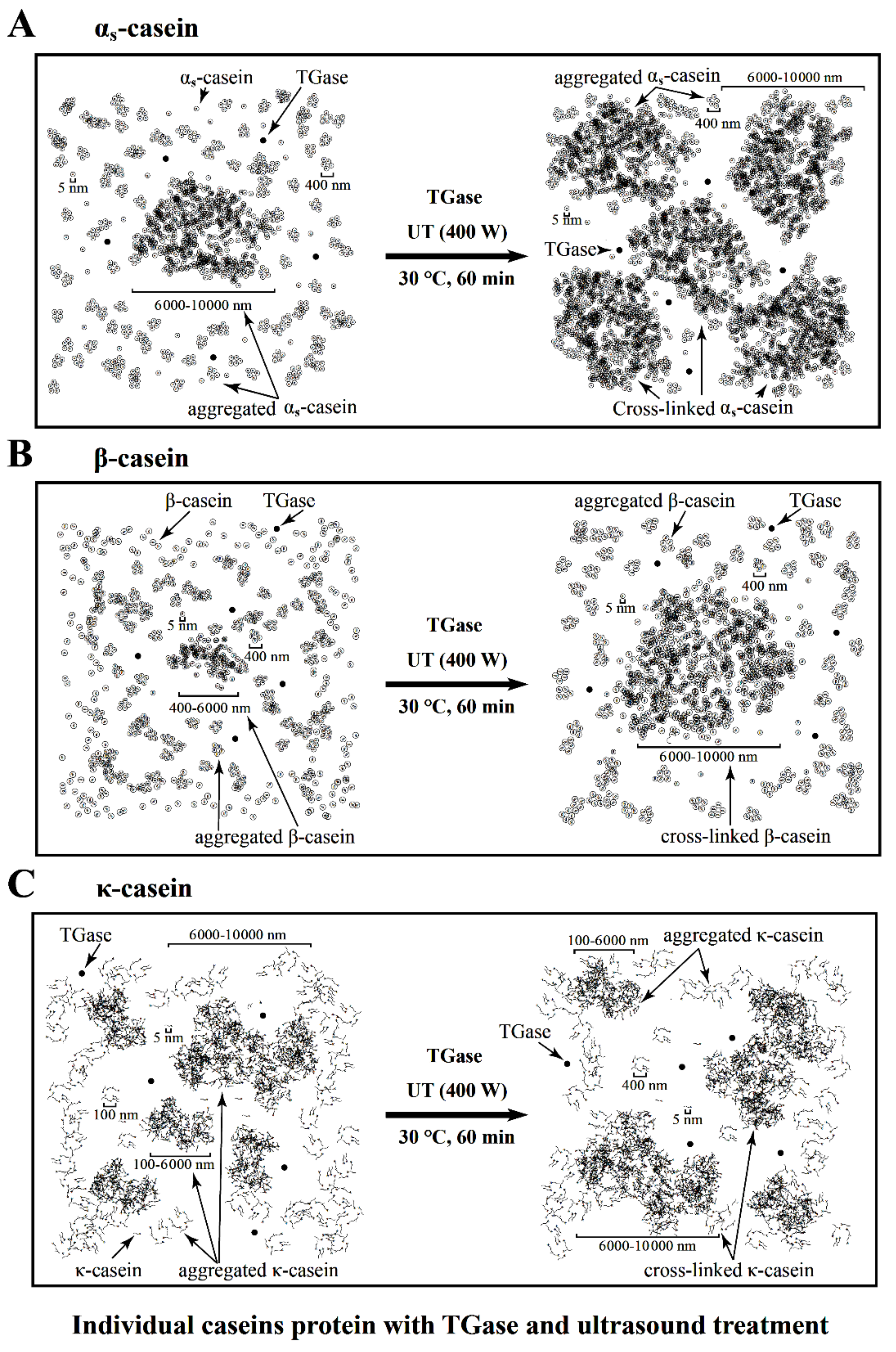
3.4. Microstructures of the Individual Caseins Treated with TGase and UT
3.5. Reaction Scheme for the TGase and UT on the Polymerization of αs-CN, β-CN and κ-CN
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopes, A.S.; Garcia, J.S.; Catharino, R.R.; Santos, L.S.; Eberlin, M.N.; Arruda, M.A.Z. Cloud point extraction applied to casein proteins of cow milk and their identification by mass spectrometry. Anal. Chim. Acta 2007, 590, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Farrell, H.M., Jr.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the proteins of cows’ milk—sixth revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef] [Green Version]
- Jensen, H.; Poulsen, N.; Andersen, K.; Hammershøj, M.; Poulsen, H.; Larsen, L.B. Distinct composition of bovine milk from Jersey and Holstein-Friesian cows with good, poor, or noncoagulation properties as reflected in protein genetic variants and isoforms. J. Dairy Sci. 2012, 95, 6905–6917. [Google Scholar] [CrossRef]
- Wang, C.; Xie, Q.; Wang, Y.; Fu, L. Effect of ultrasound treatment on allergenicity reduction of milk casein via colloid formation. J. Agric. Food Chem. 2020, 68, 4678–4686. [Google Scholar] [CrossRef]
- McDermott, A.; Visentin, G.; De Marchi, M.; Berry, D.P.; Fenelon, M.A.; O’Connor, P.M.; Kenny, O.A.; McParland, S. Prediction of individual milk proteins including free amino acids in bovine milk using mid-infrared spectroscopy and their cor-relations with milk processing characteristics. J. Dairy Sci. 2016, 99, 3171–3182. [Google Scholar] [CrossRef]
- Buettner, K.; Hertel, T.C.; Pietzsch, M. Increased thermostability of microbial transglutaminase by combination of several hot spots evolved by random and saturation mutagenesis. Amino. Acids. 2011, 42, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Giosafatto, C.; Rigby, N.; Wellner, N.; Ridout, M.; Husband, F.; Mackie, A. Microbial transglutaminase-mediated modification of ovalbumin. Food Hydrocoll. 2012, 26, 261–267. [Google Scholar] [CrossRef]
- Özer, B.; Guyot, C.; Kulozik, U. Simultaneous use of transglutaminase and rennet in milk coagulation: Effect of initial milk pH and renneting temperature. Int. Dairy J. 2012, 24, 1–7. [Google Scholar] [CrossRef]
- Kieliszek, M.; Misiewicz, A. Microbial transglutaminase and its application in the food industry. A review. Folia. Microbiol. 2014, 59, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Ridout, M.J.; Paananen, A.; Mamode, A.; Linder, M.B.; Wilde, P.J. Interaction of transglutaminase with adsorbed and spread films of β-casein and κ-casein. Colloids. Surf. B Biointerfaces 2015, 128, 254–260. [Google Scholar] [CrossRef]
- Sayadi, A.; Madadlou, A.; Khosrowshahi, A. Enzymatic cross-linking of whey proteins in low fat Iranian white cheese. Int. Dairy J. 2013, 29, 88–92. [Google Scholar] [CrossRef]
- Moon, J.H.; Hong, Y.H.; Huppertz, T.; Fox, P.F.; Kelly, A.L. Properties of casein micelles cross-linked by transglutami-nase. Int. J. Dairy Technol. 2009, 62, 27–32. [Google Scholar] [CrossRef]
- Kwiatkowska, B.; Bennett, J.; Akunna, J.; Walker, G.; Bremner, D.H. Stimulation of bioprocesses by ultrasound. Biotechnol. Adv. 2011, 29, 768–780. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.-S.; Luo, S.-Z.; Cai, J.; Zhong, X.-Y.; Jiang, S.-T.; Zhao, Y.-Y.; Zheng, Z. Transglutaminase-induced gelation properties of soy protein isolate and wheat gluten mixtures with high intensity ultrasonic pretreatment. Ultrason. Sonochemistry 2016, 31, 590–597. [Google Scholar] [CrossRef]
- Vilkhu, K.; Mawson, R.; Simons, L.; Bates, D. Applications and opportunities for ultrasound assisted extraction in the food industry—A review. Innov. Food Sci. Emerg. Technol. 2008, 9, 161–169. [Google Scholar] [CrossRef]
- Jiang, Z.; Wang, C.; Li, T.; Sun, D.; Gao, H.; Gao, Z.; Mu, Z. Effect of ultrasound on the structure and functional properties of transglutaminase-crosslinked whey protein isolate exposed to prior heat treatment. Int. Dairy J. 2019, 88, 79–88. [Google Scholar] [CrossRef]
- Cui, Q.; Wang, G.; Gao, D.; Wang, L.; Zhang, A.; Wang, X.; Xu, N.; Jiang, L. Improving the gel properties of transgenic mi-crobial transglutaminase cross-linked soybean-whey mixed protein by ultrasonic pretreatment. Process. Biochem. 2020, 91, 104–112. [Google Scholar] [CrossRef]
- Gordon, L.; Pilosof, A.M.R. Application of High-Intensity Ultrasounds to Control the Size of Whey Proteins Particles. Food Biophys. 2010, 5, 203–210. [Google Scholar] [CrossRef]
- Zhang, P.; Hu, T.; Feng, S.; Xu, Q.; Zheng, T.; Zhou, M.; Chu, X.; Huang, X.; Lu, X.; Pan, S.; et al. Effect of high intensity ultrasound on transglutaminase-catalyzed soy protein isolate cold set gel. Ultrason. Sonochemistry 2016, 29, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, S.; Uluko, H.; Liu, L.; Lu, J.; Xue, H.; Kong, F.; Lv, J. Effect of ultrasound pretreatment on rennet-induced coagulation properties of goat’s milk. Food Chem. 2014, 165, 167–174. [Google Scholar] [CrossRef]
- Hinz, K.; Huppertz, T.; Kelly, A.L. Susceptibility of the individual caseins in reconstituted skim milk to cross-linking by transglutaminase: Influence of temperature, pH and mineral equilibria. J. Dairy Res. 2012, 79, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Smolenski, G.; Haines, S.; Kwan, F.Y.-S.; Bond, J.; Farr, V.; Davis, S.R.; Stelwagen, K.; Wheeler, T.T. Characterisation of Host Defence Proteins in Milk Using a Proteomic Approach. J. Proteome Res. 2007, 6, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Moatsou, G.; Hatzinaki, A.; Samolada, M.; Anifantakis, E. Major whey proteins in ovine and caprine acid wheys from in-digenous greek breeds. Int. Dairy J. 2005, 15, 123–131. [Google Scholar] [CrossRef]
- Zhong, Q.; Wang, W.; Hu, Z.; Ikeda, S. Sequential preheating and transglutaminase pretreatments improve stability of whey protein isolate at pH 7.0 during thermal sterilization. Food Hydrocoll. 2013, 31, 306–316. [Google Scholar] [CrossRef]
- Di Pierro, P.; Mariniello, L.; Sorrentino, A.; Giosafatto, C.V.L.; Chianese, L.; Porta, R. Transglutaminase-Induced Chemical and Rheological Properties of Cheese. Food Biotechnol. 2010, 24, 107–120. [Google Scholar] [CrossRef]
- Hsieh, J.-F.; Pan, P.-H. Proteomic Profiling of the Coagulation of Milk Proteins Induced by Chymosin. J. Agric. Food Chem. 2012, 60, 2039–2045. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, S.; Barać, M.; Maćej, O. Whey proteins-properties and possibility of application. Mljekarstvo 2005, 55, 215–233. [Google Scholar]
- Tang, C.-H.; Ma, C.-Y. Modulation of the thermal stability of β-lactoglobulin by transglutaminase treatment. Eur. Food Res. Technol. 2007, 225, 649–652. [Google Scholar] [CrossRef]
- Rahila, M.P.; Kumar, R.; Mann, B.; Koli, P.S. Enzymatic Modification of Milk Proteins for the Preparation of Low Fat Dahi. J. Food Process. Preserv. 2015, 40, 1038–1046. [Google Scholar] [CrossRef]
- Cozzolino, A.; Di Pierro, P.; Mariniello, L.; Sorrentino, A.; Masi, P.; Porta, R. Incorporation of whey proteins into cheese curd by using transglutaminase. Biotechnol. Appl. Biochem. 2003, 38, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, W.A. Eeffect of microbial transglutaminase treatment on soft cheese properties. Mesop. J. Agric. 2009, 37, 19–27. [Google Scholar]
- Chen, C.C.; Chen, L.Y.; Chan, D.S.; Chen, B.Y.; Tseng, H.W.; Hsieh, J.F. Influence of microbial transglutaminase on physico-chemical and cross-linking characteristics of individual caseins. Molecules 2020, 25, 3992. [Google Scholar] [CrossRef] [PubMed]
- Munir, M.; Nadeem, M.; Qureshi, T.M.; Leong, T.S.; Gamlath, C.J.; Martin, G.J.; Ashokkumar, M. Effects of high pressure, microwave and ultrasound processing on proteins and enzyme activity in dairy systems—A review. Innov. Food Sci. Emerg. Technol. 2019, 57, 102192. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Anema, S.G. Effect of ultrasonication on the properties of skim milk used in the formation of acid gels. Innov. Food Sci. Emerg. Technol. 2010, 11, 616–622. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-C.; Chen, L.-Y.; Li, W.-T.; Chang, K.-L.; Tseng, H.-W.; Chen, B.-Y.; Chen, C.-J.; Hsieh, J.-F. Ultrasound-Assisted Transglutaminase Catalysis of the Cross-Linking and Microstructure of αs-Casein, β-Casein and κ-Casein. Processes 2021, 9, 1630. https://doi.org/10.3390/pr9091630
Chen C-C, Chen L-Y, Li W-T, Chang K-L, Tseng H-W, Chen B-Y, Chen C-J, Hsieh J-F. Ultrasound-Assisted Transglutaminase Catalysis of the Cross-Linking and Microstructure of αs-Casein, β-Casein and κ-Casein. Processes. 2021; 9(9):1630. https://doi.org/10.3390/pr9091630
Chicago/Turabian StyleChen, Chun-Chi, Liang-Yu Chen, Wen-Tai Li, Ken-Lin Chang, Hsien-Wei Tseng, Bang-Yuan Chen, Chao-Jung Chen, and Jung-Feng Hsieh. 2021. "Ultrasound-Assisted Transglutaminase Catalysis of the Cross-Linking and Microstructure of αs-Casein, β-Casein and κ-Casein" Processes 9, no. 9: 1630. https://doi.org/10.3390/pr9091630