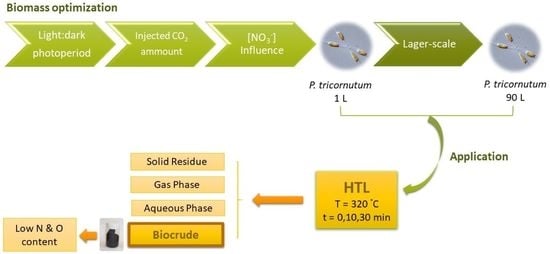
3.1. Lab-Scale Culture
3.1.1. Effect of Photoperiod
The growth of the microalgae under photoperiods 12:12 and 18:6 h of light:dark in the 1 L bioreactor is shown in
Figure 1. Maximum biomass concentrations of 1.56 and 1.73 g/L were obtained for the 12:12 and 18:6 h light:dark photoperiods, respectively. Both biomass concentrations were higher than those obtained by Morais et al. [
45] with the same microalga, who obtained a maximum concentration of 1.3 g/L under 12:12 and 24:0-h light:dark photoperiods at a lower light intensity of 74 µE/(m
2s). However, the volume of the photobioreactor was somewhat higher (2 L) to the one studied in this work. The aeration homogenizes the culture more efficiently in the smaller photobioreactors, which means that all the cells remain for less time in dark areas.
The growth rate was 0.24 and 0.33 d
−1 (
p-value = 0.01) for the 12:12 and 18:6 h light:dark photoperiods, respectively. Therefore, the growth rate raised with the light hours, which has been previously reported. Thus, a similar increase with the light cycle was also observed for
Chlorella vulgaris [
9,
46] and
Nannochloropsis [
47]. In addition, a growth rate of 0.34 d
−1 for
Nannochloropsis at the 18:6 photoperiod was previously reported [
47]. The most notable difference was observed in the day the stationary phase was reached: 8 and 12 days for the 18:6 and 12:12 h photoperiods, respectively. In both photoperiods, the total nitrate consumption was reached after 14 days of cultivation; however, the number of light hours accelerated the consumption of nutrients.
The lipids obtained increased (significantly at a confidence level 0 94%,
p-value = 0.055) from 24% ± 1% to 30% ± 3% (
Table 1) as the light exposure increased from 12 to 18 h, showing a similar trend than that reported for other species in previous studies [
48]. Thus, lipids were mainly accumulated after light exposures above 12 h. In addition to the light time, the amount of lipids depends also on other factors like the CO
2 supply or concentration of nutrients in the culture [
49]. The amount of lipids in the biomass obtained in the culture is frequently analyzed in the literature. However, a complete study of the biomass composition and productivity, which has been scarcely reported previously, is recommended to fully understand their influence in the biocrude production by HTL. Consequently, the concentration of proteins, carbohydrates and ash in the
P. tricornutum biomass were measured and presented also in
Table 1, together with the corresponding biomass and lipid productivities. Unlike what was observed for
Scenedesmus obliquus and
Chlorella [
50,
51], the protein content did not change significantly (
p-value = 0.127) when the photoperiod increased the light times. By contrast, the content of carbohydrates decreased (
p-value = 0.019) as the light hours increased. George et al. [
52] obtained similar results in cultures of
Ankistrodesmus falcatus. Biller and Ross [
53] concluded that there is a trend in the biocrude yield from HTL where lipid contents are the main positive influential factor, followed by protein and carbohydrate contents in this order of influence. Thus, a high content of lipids and proteins favors the production of hydrocarbons after hydrolysis and denitrogenation reactions and hydrolysis and decarboxylation, respectively [
28].
The ash content decreased from 20.27% ± 0.56% to 12.27% ± 0.04% (
p-value = 0.002) with the increase in light exposure. The ash portion varies according to the culture conditions [
54], and in this case, the observed reduction of ash was probably due to the corresponding increase in the photosynthetic efficiency, which, in turn, means a higher biomass production with the light cycle. A high ash content may inhibit the transformation of the microalgae in HTL and has a negative effect on the biocrude properties [
55].
Interestingly, the biomass productivity increased at higher light photoperiods, which was observed previously for the microalga
S. obliquus [
50]. The biomass productivity achieved 123.87 ± 0.06 and 200.02 ± 15.18 mg/(L·d) (
p-value = 0.013) at the 12:12 and 18:6 h light:dark photoperiods, respectively. As the lipid content and biomass productivity raised with the light, the lipid productivity also increased with the hours of light (25.06 ± 0.01 to 52.67 ± 4.00 mg/(L·d) (
p-value < 0.007) for the 12:12 and 18:6 h photoperiods, respectively). Therefore, the results showed a greater efficiency in the production of both biomass and lipids when the culture was exposed to 18 h of light.
One of the objectives of the liquefaction process is to obtain a biofuel with a low content of heteroatoms, mainly O and N.
Table 2 summarizes the biomass elemental analysis at the different photoperiods. The O and N contents of the biomass obtained at both photoperiods did not show significant differences, which was related with the biochemical composition of the biomass obtained. The contents of these heteroatoms were lower than the reported values of other microalgae, such as
Arthrospira platensis [
56],
C. vulgaris [
53] and
Dunaliella tertiolecta [
33].
Therefore, biomass grown under a larger number of light hours seems a priori more adequate to HTL due to its high biomass and lipid productivities, high protein and lipid contents and lower ash content. Thus, the 18:6 h photoperiod was chosen to produce the P. tricornutum biomass for HTL.
3.1.2. Influence of CO2 Injection
The growth curve (
Figure 2) shows the results obtained in the essays with and without an additional CO
2 injection in the
P. tricornutum culture. It can be observed that the exponential phase was extended for two more days when a continuous flow of additional CO
2 was injected. This caused a remarkable increase in the total biomass production, reaching a concentration above 2.61 g/L with a continuous injection of CO
2 compared to 1.73 g/L obtained in the absence of the supplemental CO
2. A similar notable increase was observed previously for the other diatom [
17]. Total nitrate depletion was reached at the end of both experiments (
Figure 2), as observed in the study of the effect of the photoperiod.
Table 3 shows the biochemical composition and biomass and lipid productivities of
P. tricornutum microalgae grown with and without supplemental CO
2 injections. The results show that the contents of lipids and proteins did not change significantly (
p-values: 0.171 and 0.121, respectively) when a flow of pure CO
2 was bubbled in the culture. This could be due to the fact that the metabolic flow involving the biosynthesis of these types of biomolecules is not altered if the carbon source is maintained above a minimum threshold. Other authors, however, reported an increase in lipid and protein accumulations when CO
2 was injected in the culture medium up to a maximum concentration of 10% [
17]. Carbohydrates increased with the injection of CO
2 into the culture medium, from 3.73% ± 1.02% to 12.54% ± 0.68% (
p-value = 0.0005). This is in accordance with a previous study for the microalga
S. obliquus [
57] where the availability of CO
2 in the culture medium favored the production of carbohydrates during the dark phase.
The ash concentration decreased with the CO
2 supplement from 12.27% ± 0.04% to 5.15% ± 1.46% (
p-value = 0.014). The acid nature of CO
2 in the solution controls the pH of the medium at lower values with the supplementation of carbon dioxide, avoiding the precipitation of insoluble salts of the medium during the growth [
58].
However, the productivity of the biomass remained without significant changes (
p-value = 0.353), regardless of whether the additional CO
2 was injected or not. Although an inhibition by an excess of CO
2 to the culture medium was previously reported [
59], the results of the biomass productivity did not indicate an inhibition effect by the presence of the extra CO
2. These similar values of biomass productivities may be due to the fact that the microalgae grew faster and took two more days to reach the stationary phase in the presence of extra amounts of CO
2.
The CO
2 supplemented to the microalgae system led to a significant rise in lipid productivity, from 52.67 ± 4.00 to 69.80 ± 1.68 mg/(L·d) (
p-value = 0.010), despite the similar biomass productivities obtained. A similar increase with CO
2 was observed for
C. vulgaris [
60].
The elemental composition of the microalgal biomass (
Table 4) slightly changed when the CO
2 availability increased. The results only showed a small increase in the O content (
p-value = 0.033) and a small decrease in the N content (
p-value = 0.041). N is mainly contained in proteins, which remained approximately constant with the additional CO
2 injection. Therefore, the above observed results were likely due to the remarkable increase in carbohydrates, rich in O, in the biomass cultured with extra CO
2. Therefore, there is no significant modifications regardless of the presence of a specific CO
2 injection in the culture medium, because the carbon content from air allows a nonlimiting growth of cells [
61]. Similar results have been reported for
C. vulgaris [
59].
Therefore, an extra CO2 injection was selected to obtain the P. tricornutum biomass for the subsequent larger-scale culture and biocrude production by HTL because of the high lipid productivity, together with a lower ash content achieved in the experiments with a direct supplement of CO2.
3.1.3. Effect of Initial Nitrate Concentration
Based on the previous cultures with 18:6 h of a light:dark photoperiod and CO
2 injection, the initial nitrate content was reduced with respect to the original culture medium (11.8 mM) to stress the microalga and evaluate its growth, composition and productivity. The results show that the decrease in nitrate negatively affected the biomass concentration (
Figure 3), since the cell growth was inhibited by the lack of a readily available N source. Similar results were observed earlier [
62,
63]. Culture mediums with nitrate reductions of 50% implied a 40% decrease in biomass growth (from 2.61 g/L to 1.56 g/L). In the culture media without nitrates, the biomass growth decreased to 0.86 g/L. Our results indicate that nitrates can be considered as essential nutrients for
P. tricornutum growth [
62], and therefore, there is a need to have an initial nitrate concentration of at least 11.8 mM in the media.
The original medium (nitrate concentration of 11.8 mM) reached the stationary phase on day 10, whereas the total nitrates uptake was reached on day 13. On the other hand, the culture with a 50% of nitrates reduction reached the stationary phase on day 9 because of the nitrogen limitation. The concentration of nitrates was exhausted on day 8, and therefore, cells could not reproduce. In the medium without them, the stationary phase was reached on day 6 because of the inhibitory effect of the lack of a nitrogen source.
Table 5 shows the effect of the initial nitrate concentration in the culture in the biomass composition. The final lipid content was not significantly affected by the reduction of the initial concentration of the nitrogen source. Although microalgae are subjected to stress in the absence of nitrates, favoring the accumulation of lipids in the cells [
64,
65], the harvesting and characterization of the biomass was carried out after 14 days of cultivation. At that time, nitrate was depleted from the media in all the experiments, so that all cultures were similarly stressed in terms of N availability. The concentration of proteins decreased from 49.16% ± 1.59% (nitrate concentration of 11.8 mM) to 33.47% ± 2.38% (lack of nitrate). The presence of nitrate in the medium is the main source of N assimilation for the microalgae and, therefore, essential for protein formation [
62].
The amount of carbohydrates was significantly higher (22.65% ± 2.12%) for the cultivation performed in the absence of an initial supply of nitrate than that obtained when the initial concentrations of nitrate were 11.8 and 9.6 mM (12.54% ± 0.68% and 11.96% ± 0.01%, respectively). Nutrient reductions such as nitrate drive the microalgae to accumulate energy-rich reserve compounds—essentially, lipids and carbohydrates [
64].
The ash concentration increased with the reduction of nitrate in the culture from 5.15% ± 1.46% (11.8 mM) to 9.3%5 ± 0.78% (5.9 mM) and 8.97% ± 0.62% (absence of an initial supply of nitrate). Therefore, the reduction of the N source negatively affected the biomass composition, increasing the ash content, which is not adequate for the following HTL stage. The lower ash content obtained with the higher initial nitrate concentration (11.8 mM) was due to the better photosynthetic efficiency and, therefore, higher biomass production at this nitrate concentration.
Regarding the biomass and lipid productivity,
Table 5 shows about a 2.5-fold increase in these values at the higher initial nitrate concentration. Although the composition of the biomass is affected at different levels, the main effect of the availability of large amounts of N was exerted on the growth of microalgae, boosting the values of the specific growth rate and, hence, the productivity associated to the biochemical components of the cells.
From the results obtained in the elemental analysis of the biomass (
Table 6), the main remarkable effect of the decrease in the amount of N in the growth medium was a significant reduction in the N content of the biomass when the culture was not supplemented with an initial nitrate concentration. The rest of the elements underwent changes that, although statistically significant in some cases, did not turn out to be very noteworthy.
According to the drastic reduction in biomass and lipid productivities and the increase of ash content in the absence of nitrates, the importance of the nitrate presence was confirmed for the adequate P. tricornutum growth. In this sense, an initial concentration of 11.8 mM of nitrate in the growth medium was chosen to continue the study.
3.2. Culture Scaling
Scaling tests (
Figure 4) show that the microalga did not require an adaptation phase, regardless of the reactor used, and began to grow rapidly in the culture medium. The stationary phase was reached four days later in the 90 L volume reactor compared to in the 1 L one. In addition, the specific growth rate was 0.29 and 0.17 d
−1 in the bioreactors of 1 L and 90 L, respectively, which are suitable values for this type and size of bubble column reactors indoors [
66]. The biomass produced in the 90 L bioreactor is close to other productions obtained outdoors in similar column bioreactors for this species (0.96 g/L) [
25], as well as for other pilot plant reactors, such as circular ponds or tubular photobioreactors [
21], indicating a good biomass production. It must be noted that total consumption of nitrate was not reached for the 90 L culture after 14 days.
The value of the biomass productivity obviously decreased with the bioreactor volume because of the observed greater shielding of the microalga, which prevents the light from reaching the microalga cells at higher volumes (
Table 7). Thus, the biomass productivities were 210.54 ± 5.08 and 56.01 ± 4.45 mg/L·d for the 1 L and 90 L bioreactors, respectively (
p-value < 0.0001). In addition, a light stress reduction at the higher volume, produced by the increase in dark areas, affected the lipid productivity, decreasing from a value of 69.8 ± 1.68 for the 1 L reactor to 9.85 ± 0.78 mg/(L·d) for the 90 L (
p-value < 0.0001). The high biomass and lipid productivities obtained at the 1 L bioreactor can be achieved using optimal culture conditions, which only have a remarkable positive effect at this small-scale cultivation. In this sense, the productivities obtained on a larger scale, although lower, results are adequate for the production of this microalgae and its subsequent use in the production of biofuels.
Finally, the elemental analysis of the biomass produced in both reactors hardly showed significant differences (
Table 8). A slight but significant decrease of C and H (
p-value < 0.0001) was observed in the biomass grown at a photoreactor of 90 L, related to a higher concentration of ashes and a lower concentration of lipids and carbohydrates. In addition, the concentration of N was slightly lower (
p-value = 0.0001) in the biomass cultivated in the bioreactor of 90 L (7.50% ± 0.02%) in comparison to the biomass obtained in the 1 L reactor (8.52% ± 0.02%) despite the higher concentration of proteins found in the former biomass. Consequently, a little higher (
p-value = 0.0004) concentration of O was obtained in the biomass grown in a 90 L culture (37.29% ± 0.22%) than that in the corresponding biomass cultivated in the 1 L bioreactor (30.10% ± 0.05%), which is probably connected with the observed increase in proteins as the size of the photobioreactor increased. In this sense, the elemental distribution in the biomass obtained did not change substantially, despite the differences of the biomass analysis obtained (
Table 7). Although a small size photobioreactor seems to be more adequate to obtain a biomass for biofuel production, according to the above results of the biomass analysis, the development of the HTL process on an industrial scale requires high amounts of biomass and, therefore, the use of large-scale photoreactors.
3.3. Hydrothermal Liquefaction Process
The yield of the different fractions obtained from the HTL process at 320 °C (biocrude, aqueous phase, gas phase and solid residue) for the three reaction times and both culture scales evaluated are shown in
Figure 5. The biomass grown in the 1 L photobioreactor (B-1L) exhibited a higher biocrude yield compared to the biomass produced at 90 L (B-90L) in the HTL process at all reaction times (
p-value ≤ 0.05), which was mainly due to the fact that the former had a higher lipid content (
Table 7) that contributed to the increase in the yield of this fraction [
53]. The yield of the biocrude fraction obtained from the B-1L at 10 min was similar (36.64% ± 4.93%) to those obtained by Christensen et al. (38.8% ± 1.3% at 350 °C and 15 min) for commercial
P. tricornutum at harsher operating conditions [
32], while those of B-90L were lower (25.61–30.03%).
The biocrude yields produced using the B-1L were significantly unaffected at different reaction times (33.99% ± 1.67%–36.64% ± 4.93%,
p-value > 0.05). Christensen et al. [
32] obtained similar yields at 300 and 325 °C, which are closer to the value of 320 °C used in this work. Similarly, the biocrude yield from B-90L did not change with time (25.61% ± 0.27%–30.03% ± 0.95%). Therefore, the reaction time did not have a significant effect on the biocrude yields at the high temperature used in this work (320 °C).
The yield of the aqueous fraction decreased with the reaction time using B-1L but remained approximately constant over time with B-90L. However, the yields of the aqueous fractions were higher (
p-values < 0.05) for the B-90L because of the lower biocrude yields obtained in this case. More water-insoluble molecules were produced at longer reaction times, which may be due to decarboxylation, deamination, dehydration, oligomerization and condensation reactions, thus producing a change in the product distribution and a decrease in the yield of the aqueous phase [
67].
The yield of the gas phase was nearly constant (30.51% ± 0.40% to 35.96% ± 1.98%) for B-90L. A similar behavior were reported previously for HTL carried out at 350 °C and similar reaction times with
Nannochloropsis [
53,
68]. However, there was a significant increase in the gas phase with time in the 1 L culture (28.50% ± 5.54% to 41.03% ± 1.77%) related to the higher content of C and, to a lesser extent, of carbohydrates in this biomass, which produces higher gas yields. The production of gaseous compounds is related to the yields of the biocrude and aqueous phases in the reaction mechanism proposed by Sheehan and Savage for
Nannochloropsis [
30].
The solid residues represented the smallest fractions of the products obtained in the HTL process, their yields varying from 2.59% to 4.74%, in agreement with bibliographic results, where solid fraction yields below 10% are usual for the HTL of microalgae [
29]. The low yields achieved for the solid residue are the cause for the high transformation efficiency of HTL, showing liquefaction yields (sum of biocrude and aqueous and gas phases) over 95% in all cases [
29].
3.4. Analysis of Biocrude
Table 9 shows the elemental composition, the higher heating value (HHV) and the energy recovery (ER) in the biocrude obtained by HTL using B-1L and B-90L at different reaction times.
A significant decrease in the O amount was observed in all the biocrude phases with respect to the starting biomass. The O content varied with time from 9.79% ± 0.20% to 6.86% ± 0.31% and from 9.78% ± 0.10% to 9.20% ± 0.06% in the biocrudes obtained in the HTL of the B-1L and B-90L, respectively, whereas this heteroatom content was 30.10% ± 0.05% and 37.29% ± 0.22% in the starting biomass obtained in the same bioreactors. The O concentration was moderately higher for the biocrudes from B-90L, particularly at longer HTL times. These results indicated the presence of decarboxylation reactions during the HTL process that intensified while increasing the reaction time [
69]. Furthermore, these values were lower than the O contents in the biocrude obtained in the HTL with other species, such as
Tetraselmis (12.3%),
Scenedesmus almeriensis (9.6%),
Chlorella (30.38%),
Nannochloropsis gaditana (14.49%) [
27] or
Scenedesmus (10.5%) [
70].
The N content in the biocrudes were very similar in all cases and lower (5.42% ± 0.03% to 5.39% ± 0.04%) than the corresponding N amount in the two initial biomasses (8.52% ± 0.01% and 7.50% ± 0.02% for the biomasses obtained in the reactors with 1 and 90 L, respectively) because of the denitrogenation reactions during HTL [
27]. These values were comparable to those obtained for the same species at 325 °C (5.62%) [
32].
The observed decrease in N and O amounts in the biocrudes is typical of the HTL process of microalgae, which causes the contents of C (74.46% ± 1.41%–77.52% ± 0.03%) and H (9.37% ± 0.09%–9.76% ± 0.07%) to increase with respect to the corresponding C and H amounts in the raw biomass. The concentrations of C and H changed within the range 53.13% ± 0.06%–47.14% ± 0.22% and 7.33% ± 0.01%–6.94% ± 0.03% for the 1 and 90 L cultivated microalgae, respectively. As noted above, the composition of the initial biomass hardly interfered with the C and H contents in the biocrudes, since these values were very similar. Interestingly, the amounts of C and H of the biocrudes described herein were considerably higher than those recently reported for the same microalga [
33], where the biocrudes with slightly lower N percentages and appreciably higher concentrations of O were obtained. However, the elemental composition indicates that the biocrudes cannot be directly used as transport fuel, and, consequently, a subsequent hydrotreating stage would be required to reduce the contents of N and O and, therefore, improve the chemical composition of the biocrudes to fulfil the standard regulations concerning the presence of these heteroatoms in the commercial fuel.
The calculated HHV values of the biocrudes for each of the HTL reactions of both biomasses were similar (38.27 ± 1.52–39.71 ± 0.50 MJ/kg), within the range obtained from other microalgae (30–43 MJ/kg) [
28] and close to petroleum crude oil (43 MJ/kg) [
71]. In addition, the values of HHV were significantly higher with respect to the corresponding values in the initial microalgal biomass (27.25–26.81 MJ/kg), due to the decrease in the O content and the increase of C and H contents in the biocrudes. The calorific value obtained were higher than that found by López-Barreiro et al. (30.3 and 35.9 MJ/kg) for biocrude from HTL produced at similar temperatures with
P. tricornutum cultured in bubble columns of 25 L [
33]. The presence of nutrients from the culture medium in HTL increased the biocrude yields and the contents of C and O in comparison to the HTL of commercial microalga, being responsible for the increase in calorific value [
32]. Furthermore, the HHV achieved in the present work were similar to those obtained by Christensen et al. [
32] for temperatures around 400 °C.
The energy recovered in the biocrude obtained from the B-90L were lower (38.61–41.98%) than the corresponding values for the B-1L (49.11–51.47%) because of the previously observed lower biocrude yields in the HTL with the B-90L.
The biofuel properties were significantly affected by their H/C, N/C and O/C ratios, as it is well known. The above ratios for the biocrudes, along with those of a petroleum diesel, a biodiesel and a biocrude obtained from the microalgae
N. gaditana [
72], were plotted in Van Krevelen diagrams (
Figure 6) for comparison purposes. The results showed that the O/C and N/C ratios decreased in our biocrudes with respect to the initial biomass, which were due to the denitrogenation and decarboxylation reactions taking place during the HTL process [
27].
The O/C ratios of the biocrudes decreased by 82.3% and 85.6% with respect to the initial biomass cultivated in the 1 and 90 L photobioreactors, respectively. The O/C ratios obtained (0.066–0.099) (
Figure 6a) were, in all cases, within the usual range found in the literature (0.0–0.3) [
73], being lower than those obtained for biodiesel (0.11) and
N. gaditana-derived biocrude (0.28). A decrease in O/C ratio from 0.099 to 0.066 was observed with time in the HTL of the B-1L. Besides, the lowest O/C ratio (0.066) and, therefore, the closest to the reference diesel (0.014) was achieved in the biocrude obtained in the HTL at 30 min using the B-1L.
In the same way, the N/C ratios of the biocrudes (
Figure 6b) were very similar to each other (~0.06), regardless of the biomass used. Therefore, the scaling-up did not seem to affect the fuel properties of the biocrudes significantly. A large decrease of 56.6% and 54.8% in the N/C ratios was observed with respect to the raw biomasses obtained in the 1 and 90 L cultivations, respectively. The N/C of the biocrudes decreased slightly with time, being lower than those found for the biocrude produced with the microalga
N. gaditana (0.084) but far from the N/C values of the
N. gaditana biodiesel (0.0004) and the conventional diesel (0.0048) [
72]. Furthermore, all the values obtained were between the limits found in the literature (0.056–0.1) [
73].
Based on the O/C and N/C ratios of the biocrudes, the biomass of
P. tricornutum is a promising feedstock for HTL compared to the microalga
N. gaditana. However, the H/C ratio decreased from 1.70 for the initial microalga to a range between 1.48–1.56 for the biocrudes when using B-1L. A similar decrease occurred with B-90L from 1.75 for the biomass to values around 1.5 for the biocrudes. These decreases were due to the high yields obtained from the aqueous phases. The H/C ratios of the
P. tricornutum-derived biocrudes were higher than the H/C ratio for a biocrude from
N. gaditana (1.25) [
72]. Therefore, the H/C values of the biocrude from
P. tricornutum were closer to those of the reference diesel (1.84). All these values were within the range 1.37–1.62 for biocrudes from microalgae with high protein and lipid contents [
7].
3.5. Analysis of the Aqueous Phase
The aqueous phase is one of the fractions of the HTL process that are part of the liquefied phase [
74]. The elemental analysis of these fractions (data not shown) led to a C content between 7.93% ± 0.15% and 7.99% ± 0.06% using the B-1L, whereas the C content was slightly higher (8.43% ± 0.16% to 9.48% ± 0.66%) with the biomass cultivated in the larger bioreactor. The relatively low amounts of C in the aqueous layer were due to the breakdown of macromolecules into smaller ones that are soluble in water. However the heteroatom (N and O) amounts were relatively high, mainly because of the hydrolysis of carbohydrates and proteins and the subsequent decarboxylation and deamination, which produced N and O compounds soluble in water media [
28]. The N content in the aqueous phase decreased with time using both microalgae, due to the fact that the deamination reactions are promoted at longer times [
27], but the values were lower in the case of the B-1L (11.57% ± 0.25% to 8.13% ± 0.04%) in comparison to those obtained with the B-90L (13.48% ± 0.66% to 12.56% ± 0.91%). The opposite trend with time was observed in the O content in the aqueous phase due to the increased decarboxylation of organic molecules [
27]. Thus, the O content increased with the HTL time in both cases, showing lower values (29.8% ± 0.7% to 33.6% ± 0.5%) when the B-1L was used as compared with B-90L (37.5% ± 0.4% to 30.7% ± 0.4%).
The pH values obtained in the aqueous phase were, in all cases, around 8 (7.95–8.77), consistent with the slightly alkaline values found in the literature [
29,
75], due to the formation of soluble basic compounds in this phase. A slight increase in pH was observed over time for both biomasses, being somewhat higher in the aqueous layer obtained in the HTL of the B-90L (8.33–8.77) than those obtained in the same layer from the B-1L (7.95–8.35). This may be due to the higher amount of N in the aqueous layer in the former, since nitrates were not completely consumed by the microalga cultivated in the 90 L bioreactor. In this sense, the basic composition of the aqueous phase was mainly due to the high portions of NH
3 and N compounds [
76].
Another key factor in this aqueous phase is the total organic carbon. These values indicated a high formation of new organic compounds soluble in water. The TOC values obtained in this work (880.4–956.2 ppm) were within the bibliographic range (300–1146 ppm) [
77].
3.6. Analysis of the Gaseous Fraction
Gaseous fractions are also included in the liquefied phase. The gaseous phases obtained by HTL from the different cultivated biomasses had a similar composition. These fractions were mainly composed of CO
2 (>80 mol%) in all conditions, as usually reported for the microalgal HTL [
74]. The CO
2 content of the gaseous fraction was higher than 98 mol% for the HTL reactions using the biomass grown in a 90 L culture. Here, the gas fraction also contained small amounts of linear saturated and monounsaturated light hydrocarbons (C1–C4) (<1 mol%), H
2 (<2 mol%) and CO (<1 mol%). In the HTL using B-1L, the CO
2 content was lower (79.82–97.19 mol%). In this case, a methane concentration within the range 8–17 mol% was obtained, which may be due to their high content of lipids. Christensen et al. reported a similar methane content when they performed the HTL at higher temperatures with a biomass with lower concentration of lipids [
32]. Additionally, the gas fraction contained small amounts of linear saturated and monounsaturated light hydrocarbons (C2–C4) (<1 mol%), H
2 (<2 mol%) and CO (<1 mol%). The large concentration of CO
2 of the gas fraction from HTL makes this stream suitable for recirculation towards the cultivation stage in photobioreactors, providing the inorganic carbon source necessary for its development [
72].

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}