Microbial Natural Products in Drug Discovery

 , ,
, ,
Abstract
:1. Introduction
1.1. Historical Overview of Natural Products in Drug Discovery
1.2. Distribution of Natural Products
2. Microbial Sources in Drug Discovery
2.1. Natural Products from Fungal Sources
2.2. Natural Products from Bacterial Sources
2.3. Natural Products from Algae
2.4. Natural Products from Microbial Community Interactions
3. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ebada, S.S.; Edrada, R.A.; Lin, W.; Proksch, P. Methods for isolation, purification and structural elucidation of bioactive secondary metabolites from marine invertebrates. Nat. Protoc. 2008, 3, 1820. [Google Scholar] [CrossRef] [PubMed]
- Baker, W.; van den Broek, A.; Camon, E.; Hingamp, P.; Sterk, P.; Stoesser, G.; Tuli, M.A. The EMBL nucleotide sequence database. Nucleic Acids Res. 2000, 28, 19–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, B. A new golden age of natural products drug discovery. Cell 2015, 163, 1297–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Naggar, M.E.; Soliman, R.A.; Morsy, O.M.; Abdel-Aziz, M.S. Nanoemulsion of Capsicum fruit extract as an eco-friendly antimicrobial agent for production of medical bandages. Biocatal. Agric. Biotechnol. 2020, 23, 101516. [Google Scholar] [CrossRef]
- El-Sayed, S.M.; El-Naggar, M.E.; Hussein, J.; Medhat, D.; El-Banna, M. Effect of Ficus carica L. leaves extract loaded gold nanoparticles against cisplatin-induced acute kidney injury. Colloids Surf. B Biointerfaces 2019, 184, 110465. [Google Scholar] [CrossRef]
- Sharaf, S.; El-Naggar, M.E. Wound dressing properties of cationized cotton fabric treated with carrageenan/cyclodextrin hydrogel loaded with honey bee propolis extract. Int. J. Biol. Macromol. 2019, 133, 583–591. [Google Scholar] [CrossRef]
- Sharaf, S.; El-Naggar, M.E. Eco-friendly technology for preparation, characterization and promotion of honey bee propolis extract loaded cellulose acetate nanofibers in medical domains. Cellulose 2018, 25, 5195–5204. [Google Scholar] [CrossRef]
- Hussein, R.A.; Salama, A.A.A.; El Naggar, M.E.; Ali, G.H. Medicinal impact of microalgae collected from high rate algal ponds; phytochemical and pharmacological studies of microalgae and its application in medicated bandages. Biocatal. Agric. Biotechnol. 2019, 20, 101237. [Google Scholar] [CrossRef]
- Busia, K.; Kasilo, O.M.J. Overview of traditional medicine in ECOWAS member states. Afr. Heal. Monit. WHO AFRO 2010, 13, 16–24. [Google Scholar]
- Barton, D. Comprehensive Natural Products Chemistry; Newnes: Burlington, MA, USA, 1999; ISBN 0080912834. [Google Scholar]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. The influence of natural products upon drug discovery. Nat. Prod. Rep. 2000, 17, 215–234. [Google Scholar] [CrossRef] [Green Version]
- Zhong, G.; Wan, F. An outline on the early pharmaceutical development before Galen. Chin. J. Med. Hist. 1999, 29, 178–182. [Google Scholar]
- Cheng, Z.F.; Zhen, C. The Cheng Zhi-Fan Collectanea of Medical History; Peking Univ. Med. Press: Beijing, China, 2004. [Google Scholar]
- Petrovska, B.B. Historical review of medicinal plants’ usage. Pharmacogn. Rev. 2012, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Patwardhan, B.; Mashelkar, R.A. Traditional medicine-inspired approaches to drug discovery: Can Ayurveda show the way forward? Drug Discov. Today 2009, 14, 804–811. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Courtwright, D.T. Forces of Habit: Drugs and the Making of the Modern World; Harvard University Press: Cambridge, MA, USA, 2009; ISBN 0674029909. [Google Scholar]
- Carmichael, W.W. Cyanobacteria secondary metabolites-The cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Méndez, C.; Salas, J.A. Altering the glycosylation pattern of bioactive compounds. Trends Biotechnol. 2001, 19, 449–456. [Google Scholar] [CrossRef]
- Brusotti, G.; Cesari, I.; Dentamaro, A.; Caccialanza, G.; Massolini, G. Isolation and characterization of bioactive compounds from plant resources: The role of analysis in the ethnopharmacological approach. J. Pharm. Biomed. Anal. 2014, 87, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Mollica, A.; Locatelli, M.; Stefanucci, A.; Pinnen, F. Synthesis and bioactivity of secondary metabolites from marine sponges containing dibrominated indolic systems. Molecules 2012, 17, 6083–6099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haefner, B. Drugs from the deep: Marine natural products as drug candidates. Drug Discov. Today 2003, 8, 536–544. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod. 2004, 67, 1216–1238. [Google Scholar] [CrossRef]
- Locatelli, M.; Governatori, L.; Carlucci, G.; Genovese, S.; Mollica, A.; Epifano, F. Recent application of analytical methods to phase I and phase II drugs development: A review. Biomed. Chromatogr. 2012, 26, 283–300. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.; Kelly, M.; Bowling, J.; Sims, J.; Waters, A.; Hamann, M. Advancement into the Arctic region for bioactive sponge secondary metabolites. Mar. Drugs 2011, 9, 2423–2437. [Google Scholar] [CrossRef] [PubMed]
- Laport, M.S.; Santos, O.C.S.; Muricy, G. Marine sponges: Potential sources of new antimicrobial drugs. Curr. Pharm. Biotechnol. 2009, 10, 86–105. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2012, 29, 144–222. [Google Scholar] [CrossRef]
- Valentin, B.B.; Vinod, V.; Beulah, M.C. Biopotential of secondary metabolites isolated from marine sponge Dendrilla nigra. Asian Pac. J. Trop. Dis. 2011, 1, 299–303. [Google Scholar] [CrossRef]
- Rönsberg, D.; Debbab, A.; Mándi, A.; Wray, V.; Dai, H.; Kurtán, T.; Proksch, P.; Aly, A.H. Secondary metabolites from the endophytic fungus Pestalotiopsis virgatula isolated from the mangrove plant Sonneratia caseolaris. Tetrahedron Lett. 2013, 54, 3256–3259. [Google Scholar] [CrossRef] [Green Version]
- Niu, G.; Li, W. Next-generation drug discovery to combat antimicrobial resistance. Trends Biochem. Sci. 2019, 44, 961–972. [Google Scholar] [CrossRef]
- Saleem, M.; Tousif, M.I.; Riaz, N.; Ahmed, I.; Schulz, B.; Ashraf, M.; Nasar, R.; Pescitelli, G.; Hussain, H.; Jabbar, A. Cryptosporioptide: A bioactive polyketide produced by an endophytic fungus Cryptosporiopsis sp. Phytochemistry 2013, 93, 199–202. [Google Scholar] [CrossRef]
- Charyulu, E.M.; Sekaran, G.; Rajakumar, G.S.; Gnanamani, A. Antimicrobial Activity of Secondary Metabolite from Marine Isolate, Pseudomonas Sp. Against Gram Positive and Negative Bacteria Including MRSA. Indian J. Exp. Biol. 2009, 47, 964–968. [Google Scholar]
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.-K. Pharmaceutically active secondary metabolites of marine actinobacteria. Microbiol. Res. 2014, 169, 262–278. [Google Scholar] [CrossRef]
- Fenical, W. Chemical studies of marine bacteria: Developing a new resource. Chem. Rev. 1993, 93, 1673–1683. [Google Scholar] [CrossRef]
- Fleming, A. On the antibacterial action of cultures of a penicillium, with special reference to their use in the isolation of B. influenzae. Rev. Infect. Dis. 1980, 129–139. [Google Scholar] [CrossRef]
- Shaaban, K.A.; Shepherd, M.D.; Ahmed, T.A.; Nybo, S.E.; Leggas, M.; Rohr, J. Pyramidamycins AD and 3-hydroxyquinoline-2-carboxamide; cytotoxic benzamides from Streptomyces sp. DGC1. J. Antibiot. 2012, 65, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Clardy, J.; Fischbach, M.A.; Walsh, C.T. New antibiotics from bacterial natural products. Nat. Biotechnol. 2006, 24, 1541–1550. [Google Scholar] [CrossRef]
- Ceniceros, A.; Dijkhuizen, L.; Petrusma, M.; Medema, M.H. Genome-based exploration of the specialized metabolic capacities of the genus Rhodococcus. BMC Genom. 2017, 18, 593. [Google Scholar] [CrossRef]
- Agarwal, V.; Blanton, J.M.; Podell, S.; Taton, A.; Schorn, M.A.; Busch, J.; Lin, Z.; Schmidt, E.W.; Jensen, P.R.; Paul, V.J. Metagenomic discovery of polybrominated diphenyl ether biosynthesis by marine sponges. Nat. Chem. Biol. 2017, 13, 537. [Google Scholar] [CrossRef] [Green Version]
- Ueoka, R.; Bhushan, A.; Probst, S.I.; Bray, W.M.; Lokey, R.S.; Linington, R.G.; Piel, J. Genome-Based Identification of a Plant-Associated Marine Bacterium as a Rich Natural Product Source. Angew. Chem. 2018, 130, 14727–14731. [Google Scholar] [CrossRef]
- Wang, L.; Ravichandran, V.; Yin, Y.; Yin, J.; Zhang, Y. Natural products from mammalian gut microbiota. Trends Biotechnol. 2019, 37, 492–504. [Google Scholar] [CrossRef]
- Chevrette, M.G.; Carlson, C.M.; Ortega, H.E.; Thomas, C.; Ananiev, G.E.; Barns, K.J.; Book, A.J.; Cagnazzo, J.; Carlos, C.; Flanigan, W. The antimicrobial potential of Streptomyces from insect microbiomes. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Stewart, E.J. Growing unculturable bacteria. J. Bacteriol. 2012, 194, 4151–4160. [Google Scholar] [CrossRef] [Green Version]
- Strobel, G.; Daisy, B.; Castillo, U.; Harper, J. Natural products from endophytic microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S. The role of natural product chemistry in drug discovery. J. Nat. Prod. 2004, 67, 2141–2153. [Google Scholar] [CrossRef] [PubMed]
- Flisiak, R.; Horban, A.; Gallay, P.; Bobardt, M.; Selvarajah, S.; Wiercinska-Drapalo, A.; Siwak, E.; Cielniak, I.; Higersberger, J.; Kierkus, J. The cyclophilin inhibitor Debio-025 shows potent anti–hepatitis C effect in patients coinfected with hepatitis C and human immunodeficiency virus. Hepatology 2008, 47, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, N.; Balakumar, S.; Indumathi, P.; Ayyadurai, A.; Vivek, R. Production and optimization of mevastatin using Penicillium citrinum NCIM 768. J. Microb Biochem. Technol. 2012, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- López, J.L.C.; Pérez, J.A.S.; Sevilla, J.M.F.; Fernández, F.G.A.; Grima, E.M.; Chisti, Y. Production of lovastatin by Aspergillus terreus: Effects of the C: N ratio and the principal nutrients on growth and metabolite production. Enzyme Microb. Technol. 2003, 33, 270–277. [Google Scholar] [CrossRef] [Green Version]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach; John Wiley & Sons: Hoboken, NJ, USA, 2002; ISBN 0471496413. [Google Scholar]
- Balba, H. Review of strobilurin fungicide chemicals. J. Environ. Sci. Heal. Part B 2007, 42, 441–451. [Google Scholar] [CrossRef]
- Petrini, O. Fungal endophytes of tree leaves. In Microbial Ecology of Leaves; Springer: Berlin/Heidelberg, Germany, 1991; pp. 179–197. [Google Scholar]
- Hyde, K.D.; Soytong, K. The fungal endophyte dilemma. Fungal Divers. 2008, 33, e173. [Google Scholar]
- Arnold, A.E. Understanding the diversity of foliar endophytic fungi: Progress, challenges, and frontiers. Fungal Biol. Rev. 2007, 21, 51–66. [Google Scholar] [CrossRef]
- Aly, A.H.; Debbab, A.; Kjer, J.; Proksch, P. Fungal endophytes from higher plants: A prolific source of phytochemicals and other bioactive natural products. Fungal Divers. 2010, 41, 1–16. [Google Scholar] [CrossRef]
- Kusari, S.; Verma, V.C.; Lamshoeft, M.; Spiteller, M. An endophytic fungus from Azadirachta indica A. Juss. that produces azadirachtin. World J. Microbiol. Biotechnol. 2012, 28, 1287–1294. [Google Scholar] [CrossRef]
- Zhang, P.; Zhou, P.; Jiang, C.; Yu, H.; Yu, L. Screening of taxol-producing fungi based on PCR amplification from Taxus. Biotechnol. Lett. 2008, 30, 2119. [Google Scholar] [CrossRef]
- Shen, L.; Shi, D.; Song, Y.; Tan, R. Chemical constituents of liquid culture of endophyte IFB-E012 in Artemisia annua. Chin. J. Nat. Med. 2009, 7, 354–356. [Google Scholar] [CrossRef]
- Kour, A.; Shawl, A.S.; Rehman, S.; Sultan, P.; Qazi, P.H.; Suden, P.; Khajuria, R.K.; Verma, V. Isolation and identification of an endophytic strain of Fusarium oxysporum producing podophyllotoxin from Juniperus recurva. World J. Microbiol. Biotechnol. 2008, 24, 1115–1121. [Google Scholar] [CrossRef]
- Weber, D.; Sterner, O.; Anke, T.; Gorzalczancy, S.; Martino, V.; Acevedo, C. Phomol, a new antiinflammatory metabolite from an endophyte of the medicinal plant Erythrina crista-galli. J. Antibiot. 2004, 57, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusari, S.; Pandey, S.P.; Spiteller, M. Untapped mutualistic paradigms linking host plant and endophytic fungal production of similar bioactive secondary metabolites. Phytochemistry 2013, 91, 81–87. [Google Scholar] [CrossRef]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef]
- Kusari, S.; Zuühlke, S.; Spiteller, M. Effect of artificial reconstitution of the interaction between the plant Camptotheca acuminata and the fungal endophyte Fusarium solani on camptothecin biosynthesis. J. Nat. Prod. 2011, 74, 764–775. [Google Scholar] [CrossRef]
- Eyberger, A.L.; Dondapati, R.; Porter, J.R. Endophyte fungal isolates from Podophyllum peltatum produce podophyllotoxin. J. Nat. Prod. 2006, 69, 1121–1124. [Google Scholar] [CrossRef]
- Kusari, S.; Lamshöft, M.; Spiteller, M. Aspergillus fumigatus Fresenius, an endophytic fungus from Juniperus communis L. Horstmann as a novel source of the anticancer pro-drug deoxypodophyllotoxin. J. Appl. Microbiol. 2009, 107, 1019–1030. [Google Scholar] [CrossRef]
- Kusari, S.; Zuühlke, S.; Kosuth, J.; Cellarova, E.; Spiteller, M. Light-independent metabolomics of endophytic Thielavia subthermophila provides insight into microbial hypericin biosynthesis. J. Nat. Prod. 2009, 72, 1825–1835. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berdy, J. The discovery of new bioactive microbial metabolites: Screening and identification. Prog. Ind. Microbiol. 1989, 27, 3–27. [Google Scholar]
- Kämpfer, P. The family Streptomycetaceae, part I: Taxonomy. Prokaryotes 2006, 3, 538–604. [Google Scholar]
- Yarza, P.; Richter, M.; Peplies, J.; Euzeby, J.; Amann, R.; Schleifer, K.-H.; Ludwig, W.; Glöckner, F.O.; Rosselló-Móra, R. The All-Species Living Tree project: A 16S rRNA-based phylogenetic tree of all sequenced type strains. Syst. Appl. Microbiol. 2008, 31, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chater, K.F. Regulation of sporulation in Streptomyces coelicolor A3 (2): A checkpoint multiplex? Curr. Opin. Microbiol. 2001, 4, 667–673. [Google Scholar] [CrossRef]
- Kimura, K.; Kanou, F.; Takahashi, H.; Esumi, Y.; Uramoto, M.; Yoshihama, M. Propeptin, a New Inhibitor of Prolyl Endopeptidase Produced by Microbispora. J. Antibiot. 1997, 50, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.-H.; Wang, Y.; Wang, Z.; Zhou, J.-Q.; Jin, W.-Z.; You, X.-F.; Gao, H.; Zhao, L.-X.; Si, S.-Y.; Li, X. Chemomicin A, a new angucyclinone antibiotic produced by Nocardia mediterranei subsp. kanglensis 1747-64. J. Antibiot. 2007, 60, 211–215. [Google Scholar] [CrossRef]
- Hatsu, M.; Sasaki, T.; Gomi, S.; Kodama, Y.; Sezaki, M.; Inouye, S.; Kondo, S. A new tetracycline antibiotic with antitumor activity. J. Antibiot. 1992, 45, 325–330. [Google Scholar] [CrossRef]
- Maskey, R.P.; Kock, I.; Helmke, E.; Laatsch, H. Isolation and Structure Determination of Phenazostatin D, a New Phenazine froma Marine Actinomycete Isolate Pseudonocardia sp. B6273. Z. Für Naturforsch. B 2003, 58, 692–694. [Google Scholar] [CrossRef]
- Tanaka, Y.; Komaki, H.; Yazawa, K.; Mikami, Y.; Nemoto, A.; Tojyo, T.; Kadowaki, K.; Shigemori, H.; Kobayashi, J. Brasilinolide A, a new macrolide antibiotic produced by Nocardia brasiliensis: Producing strain, isolation and biological activity. J. Antibiot. 1997, 50, 1036–1041. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, I.; Takahashi, K.-I.; Asano, K.; Kawamoto, I.; Yasuzawa, T.; Ashizawat, T.; Tomita, F.; Nakano, H. DC92-B, a new antitumor antibiotic from Actinomadura. J. Antibiot. 1988, 41, 1151–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemriss, S.; Laurent, F.; Couble, A.; Casoli, E.; Lancelin, J.M.; Saintpierre-Bonaccio, D.; Rifai, S.; Fassouane, A.; Boiron, P. Screening of nonpolyenic antifungal metabolites produced by clinical isolates of actinomycetes. Can. J. Microbiol. 2003, 49, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Maeda, A.; Nagai, H.; Yazawa, K.; Tanaka, Y.; Imai, T.; Mikami, Y.; Kuramochi, T.; Yamazaki, C. Three new reduced anthracycline related compounds from pathogenic Nocardia brasiliensis. J. Antibiot. 1994, 47, 976–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, H.; Sakai, H.-I.; Kohsaka, M.; Konomi, T.; Hosoda, J.; Kubochi, Y.; Iguchi, E.; Imanaka, H. Nocardicin A, a new monocyclic β-lactam antibiotic. J. Antibiot. 1976, 29, 492–500. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, Y.; Shirasaki, S.; Shiba, S.; Kawasaki, T.; Matsuo, Y.; Adachi, K.; Shizuri, Y. Piericidins C 7 and C 8, New Cytotoxic Antibiotics Produced by a Marine Streptomyces sp. J. Antibiot. 2007, 60, 196–200. [Google Scholar] [CrossRef] [Green Version]
- Radzom, M.; Zeeck, A.; Antal, N.; Fiedler, H.-P. Fogacin, a novel cyclic octaketide produced by Streptomyces strain Tü 6319. J. Antibiot. 2006, 59, 315–317. [Google Scholar] [CrossRef]
- Doull, J.L.; Singh, A.K.; Hoare, M.; Ayer, S.W. Conditions for the production of jadomycin B byStreptomyces venezuelae ISP5230: Effects of heat shock, ethanol treatment and phage infection. J. Ind. Microbiol. 1994, 13, 120–125. [Google Scholar] [CrossRef]
- Singh, M.P.; Petersen, P.J.; Weiss, W.J.; Kong, F.; Greenstein, M. Saccharomicins, novel heptadecaglycoside antibiotics produced by Saccharothrix espanaensis: Antibacterial and mechanistic activities. Antimicrob. Agents Chemother. 2000, 44, 2154–2159. [Google Scholar] [CrossRef] [Green Version]
- Imai, H.; Suzuki, K.-I.; Morioka, M.; Numasaki, Y.; Kadota, S.; Nagai, K.; Sato, T.; Iwanami, M.; Saito, T. Okilactomycin, a novel antibiotic produced by a Streptomyces species. J. Antibiot. 1987, 40, 1475–1482. [Google Scholar] [CrossRef] [Green Version]
- Bibb, M.J. Regulation of secondary metabolism in streptomycetes. Curr. Opin. Microbiol. 2005, 8, 208–215. [Google Scholar] [CrossRef]
- Gesheva, V.; Ivanova, V.; Gesheva, R. Effects of nutrients on the production of AK-111-81 macrolide antibiotic by Streptomyces hygroscopicus. Microbiol. Res. 2005, 160, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Weber, T.; Welzel, K.; Pelzer, S.; Vente, A.; Wohlleben, W. Exploiting the genetic potential of polyketide producing streptomycetes. J. Biotechnol. 2003, 106, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Corbin, D.R.; Greenplate, J.T.; Wong, E.Y.; Purcell, J.P. Cloning of an insecticidal cholesterol oxidase gene and its expression in bacteria and in plant protoplasts. Appl. Environ. Microbiol. 1994, 60, 4239–4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopwood, D.A. Forty years of genetics with Streptomyces: From in vivo through in vitro to in silico. Microbiology 1999, 145, 2183–2202. [Google Scholar] [CrossRef] [Green Version]
- Brautaset, T.; Sekurova, O.N.; Sletta, H.; Ellingsen, T.E.; Strøm, A.R.; Valla, S.; Zotchev, S.B. Biosynthesis of the polyene antifungal antibiotic nystatin in Streptomyces noursei ATCC 11455: Analysis of the gene cluster and deduction of the biosynthetic pathway. Chem. Biol. 2000, 7, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Raja, A.; Prabakarana, P. Actinomycetes and drug-an overview. Am. J. Drug Discov. Dev. 2011, 1, 75–84. [Google Scholar] [CrossRef]
- Meanwell, R.J.L.; Shama, G. Production of streptomycin from chitin using Streptomyces griseus in bioreactors of different configuration. Bioresour. Technol. 2008, 99, 5634–5639. [Google Scholar] [CrossRef] [Green Version]
- Dulmage, H.T. The production of neomycin by Streptomyces fradiae in synthetic media. Appl. Microbiol. 1953, 1, 103. [Google Scholar] [CrossRef] [Green Version]
- Yanai, K.; Murakami, T. The kanamycin biosynthetic gene cluster from Streptomyces kanamyceticus. J. Antibiot. 2004, 57, 351–354. [Google Scholar] [CrossRef] [Green Version]
- Petković, H.; Cullum, J.; Hranueli, D.; Hunter, I.S.; Perić-Concha, N.; Pigac, J.; Thamchaipenet, A.; Vujaklija, D.; Long, P.F. Genetics of Streptomyces rimosus, the oxytetracycline producer. Microbiol. Mol. Biol. Rev. 2006, 70, 704–728. [Google Scholar] [CrossRef] [Green Version]
- Vitayakritsirikul, V.; Jaemsaeng, R.; Lohmaneeratana, K.; Thanapipatsiri, A.; Daduang, R.; Chuawong, P.; Thamchaipenet, A. Improvement of chloramphenicol production in Streptomyces venezuelae ATCC 10712 by overexpression of the aroB and aroK genes catalysing steps in the shikimate pathway. Antonie Van Leeuwenhoek 2016, 109, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Williamson, J.M.; Inamine, E.; Wilson, K.E.; Douglas, A.W.; Liesch, J.M.; Albers-Schönberg, G. Biosynthesis of the beta-lactam antibiotic, thienamycin, by Streptomyces cattleya. J. Biol. Chem. 1985, 260, 4637–4647. [Google Scholar] [PubMed]
- Zhanel, G.G.; Homenuik, K.; Nichol, K.; Noreddin, A.; Vercaigne, L.; Embil, J.; Gin, A.; Karlowsky, J.A.; Hoban, D.J. The glycylcyclines. Drugs 2004, 64, 63–88. [Google Scholar] [CrossRef] [PubMed]
- Chapman, T.M.; Perry, C.M. Everolimus. Drugs 2004, 64, 861–872. [Google Scholar] [CrossRef]
- Broggini, M.; Marchini, S.; Fontana, E.; Moneta, D.; Fowst, C.; Geroni, C. Brostallicin: A new concept in minor groove DNA binder development. Anticancer. Drugs 2004, 15, 1–6. [Google Scholar] [CrossRef]
- Salas, J.A.; Méndez, C. yGenetic manipulation of antitumor-agent biosynthesis to produce novel drugs. Trends Biotechnol. 1998, 16, 475–482. [Google Scholar] [CrossRef]
- Khosla, C.; Zawada, R.J.X. Generation of polyketide libraries via combinatorial biosynthesis. Trends Biotechnol. 1996, 14, 335–341. [Google Scholar] [CrossRef]
- Watve, M.G.; Tickoo, R.; Jog, M.M.; Bhole, B.D. How many antibiotics are produced by the genus Streptomyces? Arch. Microbiol. 2001, 176, 386–390. [Google Scholar] [CrossRef]
- Qin, S.; Xing, K.; Jiang, J.-H.; Xu, L.-H.; Li, W.-J. Biodiversity, bioactive natural products and biotechnological potential of plant-associated endophytic actinobacteria. Appl. Microbiol. Biotechnol. 2011, 89, 457–473. [Google Scholar] [CrossRef]
- Janso, J.E.; Carter, G.T. Biosynthetic potential of phylogenetically unique endophytic actinomycetes from tropical plants. Appl. Environ. Microbiol. 2010, 76, 4377–4386. [Google Scholar] [CrossRef] [Green Version]
- Kupchan, S.M. Maytansine, a novel antileukemic ansa macrolide from Maytenus ovatus. J. Am. Chem. Soc. 1972, 94, 1354–1356. [Google Scholar] [CrossRef] [PubMed]
- Powell, R.G.; Weisleder, D.; Smith Jr, C.R.; Kozlowski, J.; Rohwedder, W.K. Treflorine, trenudine, and N-methyltrenudone: Novel maytansinoid tumor inhibitors containing two fused macrocyclic rings. J. Am. Chem. Soc. 1982, 104, 4929–4934. [Google Scholar] [CrossRef]
- Suwanborirux, K.; Chang, C.-J.; Spjut, R.W.; Cassady, J.M. Ansamitocin P-3, a maytansinoid, fromClaopodium crispifolium andAnomodon attenuatus or associated actinomycetes. Experientia 1990, 46, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Higashide, E.; Asai, M.; Ootsu, K.; Tanida, S.; Kozai, Y.; Hasegawa, T.; Kishi, T.; Sugino, Y.; Yoneda, M. Ansamitocin, a group of novel maytansinoid antibiotics with antitumour properties from Nocardia. Nature 1977, 270, 721–722. [Google Scholar] [CrossRef]
- Taechowisan, T.; Lu, C.; Shen, Y.; Lumyong, S. 4-arylcoumarin inhibits immediate-type allergy. Food Agric. Immunol. 2007, 18, 203–211. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Nassar, A.H.; Hardy, G.E.S.J.; Sivasithamparam, K. Plant growth promotion and biological control of Pythium aphanidermatum, a pathogen of cucumber, by endophytic actinomycetes. J. Appl. Microbiol. 2009, 106, 13–26. [Google Scholar] [CrossRef]
- Shimizu, M.; Yazawa, S.; Ushijima, Y. A promising strain of endophytic Streptomyces sp. for biological control of cucumber anthracnose. J. Gen. Plant. Pathol. 2009, 75, 27–36. [Google Scholar] [CrossRef]
- Merzaeva, O.V.; Shirokikh, I.G. The production of auxins by the endophytic bacteria of winter rye. Appl. Biochem. Microbiol. 2010, 46, 44–50. [Google Scholar] [CrossRef]
- Wohlleben, W.; Mast, Y.; Muth, G.; Röttgen, M.; Stegmann, E.; Weber, T. Synthetic biology of secondary metabolite biosynthesis in actinomycetes: Engineering precursor supply as a way to optimize antibiotic production. FEBS Lett. 2012, 586, 2171–2176. [Google Scholar] [CrossRef]
- Medema, M.H.; Breitling, R.; Takano, E. Synthetic biology in Streptomyces bacteria. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 497, pp. 485–502. ISBN 0076-6879. [Google Scholar]
- Zaehner, H.; Fiedler, H.-P. The need for new antibiotics: Possible ways forward. In Proceedings of the Symposia-Society for General MIcrobiology; Cambridge University Press: Cambridge, UK, 1995; Volume 1, p. 67. [Google Scholar]
- Nakagawa, A.; Omura, S. Biosynthesis of bioactive microbial metabolites and its application to the structural studies and production of hybrid compounds. J. Antibiot. 1996, 49, 717–741. [Google Scholar] [CrossRef] [Green Version]
- Moellering, R.C. Linezolid: The first oxazolidinone antimicrobial. Ann. Intern. Med. 2003, 139, 864. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Report and Proceedings of a WHO Meeting in Berlin, Germany, 13–17 October 1997. WHO EM CL 2001, 97, 4. [Google Scholar]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.-M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3 (2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, H.; Ishikawa, J.; Hanamoto, A.; Shinose, M.; Kikuchi, H.; Shiba, T.; Sakaki, Y.; Hattori, M.; Ōmura, S. Complete genome sequence and comparative analysis of the industrial microorganism Streptomyces avermitilis. Nat. Biotechnol. 2003, 21, 526. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.; Mathivanan, N. Screening of marine actinomycetes isolated from the Bay of Bengal, India for antimicrobial activity and industrial enzymes. World J. Microbiol. Biotechnol. 2009, 25, 2103–2111. [Google Scholar] [CrossRef]
- Xiao, J.; Luo, Y.; Xie, S.; Xu, J. Serinicoccus profundi sp. nov., an actinomycete isolated from deep-sea sediment, and emended description of the genus Serinicoccus. Int. J. Syst. Evol. Microbiol. 2011, 61, 16–19. [Google Scholar] [CrossRef]
- Oh, D.-C.; Gontang, E.A.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Salinipyrones and pacificanones, mixed-precursor polyketides from the marine actinomycete Salinispora pacifica. J. Nat. Prod. 2008, 71, 570–575. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.R.; Williams, P.G.; Oh, D.-C.; Zeigler, L.; Fenical, W. Species-specific secondary metabolite production in marine actinomycetes of the genus Salinispora. Appl. Environ. Microbiol. 2007, 73, 1146–1152. [Google Scholar] [CrossRef] [Green Version]
- Maloney, K.N.; MacMillan, J.B.; Kauffman, C.A.; Jensen, P.R.; DiPasquale, A.G.; Rheingold, A.L.; Fenical, W. Lodopyridone, a structurally unprecedented alkaloid from a marine actinomycete. Org. Lett. 2009, 11, 5422–5424. [Google Scholar] [CrossRef] [Green Version]
- Asolkar, R.N.; Kirkland, T.N.; Jensen, P.R.; Fenical, W. Arenimycin, an antibiotic effective against rifampin-and methicillin-resistant Staphylococcus aureus from the marine actinomycete Salinispora arenicola. J. Antibiot. 2010, 63, 37–39. [Google Scholar] [CrossRef] [Green Version]
- Dias, D.A.; Urban, S.; Roessner, U. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Straight, P.D. Antibiotic discovery through microbial interactions. Curr. Opin. Microbiol. 2019, 51, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Zhanel, G.G.; Walters, M.; Noreddin, A.; Vercaigne, L.M.; Wierzbowski, A.; Embil, J.M.; Gin, A.S.; Douthwaite, S.; Hoban, D.J. The ketolides. Drugs 2002, 62, 1771–1804. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.M.; Ibbotson, T. Biapenem. Drugs 2002, 62, 2221–2234. [Google Scholar] [CrossRef]
- Keating, G.M.; Perry, C.M. Ertapenem. Drugs 2005, 65, 2151–2178. [Google Scholar] [CrossRef] [PubMed]
- Pastores, G.M.; Barnett, N.L.; Kolodny, E.H. An open-label, noncomparative study of miglustat in type I Gaucher disease: Efficacy and tolerability over 24 months of treatment. Clin. Ther. 2005, 27, 1215–1227. [Google Scholar] [CrossRef]
- Fenton, C.; Keating, G.M.; Curran, M.P. Daptomycin. Drugs 2004, 64, 445–455. [Google Scholar] [CrossRef]
- Sugiura, T.; Ariyoshi, Y.; Negoro, S.; Nakamura, S.; Ikegami, H.; Takada, M.; Yana, T.; Fukuoka, M. Phase I/II study of amrubicin, a novel 9-aminoanthracycline, in patients with advanced non-small-cell lung cancer. Investig. New Drugs 2005, 23, 331–337. [Google Scholar] [CrossRef]
- Gupta, A.K.; Chow, M. Pimecrolimus: A review. J. Eur. Acad. Dermatol. Venereol. 2003, 17, 493–503. [Google Scholar] [CrossRef]
- Guiry, M.D. How many species of algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef]
- Abdel-Razek, A.S.; Hamed, A.; Frese, M.; Sewald, N.; Shaaban, M. Penicisteroid C: New polyoxygenated steroid produced by co-culturing of Streptomyces piomogenus with Aspergillus niger. Steroids 2018, 138, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Leal, M.C.; Munro, M.H.G.; Blunt, J.W.; Puga, J.; Jesus, B.; Calado, R.; Rosa, R.; Madeira, C. Biogeography and biodiscovery hotspots of macroalgal marine natural products. Nat. Prod. Rep. 2013, 30, 1380–1390. [Google Scholar] [CrossRef] [PubMed]
- Vuong, Q.V. Utilisation of Bioactive Compounds from Agricultural and Food Production Waste; CRC Press: Boca Raton, FL, USA, 2017; ISBN 1351647598. [Google Scholar]
- Tringali, C.; Oriente, G.; Piattelli, M.; Geraci, C.; Nicolosi, G.; Breitmaier, E. Crenuladial, an antimicrobial diterpenoid from the brown alga Dilophus ligulatus. Can. J. Chem. 1988, 66, 2799–2802. [Google Scholar] [CrossRef]
- Topcu, G.; Aydogmus, Z.; Imre, S.; Gören, A.C.; Pezzuto, J.M.; Clement, J.A.; Kingston, D.G.I. Brominated Sesquiterpenes from the Red Alga Laurencia o btusa. J. Nat. Prod. 2003, 66, 1505–1508. [Google Scholar] [CrossRef] [PubMed]
- Butler, G.L. Algae and pesticides. In Residue Reviews; Springer: Berlin/Heidelberg, Germany, 1977; pp. 19–62. [Google Scholar]
- Zuck, K.M.; Shipley, S.; Newman, D.J. Induced production of N-formyl alkaloids from Aspergillus fumigatus by co-culture with Streptomyces peucetius. J. Nat. Prod. 2011, 74, 1653–1657. [Google Scholar] [CrossRef]
- Cueto, M.; Jensen, P.R.; Kauffman, C.; Fenical, W.; Lobkovsky, E.; Clardy, J. Pestalone, a new antibiotic produced by a marine fungus in response to bacterial challenge. J. Nat. Prod. 2001, 64, 1444–1446. [Google Scholar] [CrossRef]
- Oh, D.-C.; Jensen, P.R.; Kauffman, C.A.; Fenical, W. Libertellenones A–D: Induction of cytotoxic diterpenoid biosynthesis by marine microbial competition. Bioorg. Med. Chem. 2005, 13, 5267–5273. [Google Scholar] [CrossRef]
- Oh, D.-C.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Induced production of emericellamides A and B from the marine-derived fungus Emericella sp. in competing co-culture. J. Nat. Prod. 2007, 70, 515–520. [Google Scholar] [CrossRef]
- Park, H.B.; Kwon, H.C.; Lee, C.-H.; Yang, H.O. Glionitrin A, an antibiotic-Antitumor metabolite derived from competitive interaction between abandoned mine microbes. J. Nat. Prod. 2009, 72, 248–252. [Google Scholar] [CrossRef]
- Hoshino, S.; Zhang, L.; Awakawa, T.; Wakimoto, T.; Onaka, H.; Abe, I. Arcyriaflavin E, a new cytotoxic indolocarbazole alkaloid isolated by combined-culture of mycolic acid-containing bacteria and Streptomyces cinnamoneus NBRC 13823. J. Antibiot. 2015, 68, 342–344. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, S.; Wakimoto, T.; Onaka, H.; Abe, I. Chojalactones A–C, cytotoxic butanolides isolated from Streptomyces sp. cultivated with mycolic acid containing bacterium. Org. Lett. 2015, 17, 1501–1504. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, S.; Okada, M.; Wakimoto, T.; Zhang, H.; Hayashi, F.; Onaka, H.; Abe, I. Niizalactams A–C, multicyclic macrolactams isolated from combined culture of Streptomyces with mycolic acid-containing bacterium. J. Nat. Prod. 2015, 78, 3011–3017. [Google Scholar] [CrossRef] [PubMed]
- Adnani, N.; Chevrette, M.G.; Adibhatla, S.N.; Zhang, F.; Yu, Q.; Braun, D.R.; Nelson, J.; Simpkins, S.W.; McDonald, B.R.; Myers, C.L. Coculture of marine invertebrate-associated bacteria and interdisciplinary technologies enable biosynthesis and discovery of a new antibiotic, keyicin. ACS Chem. Biol. 2017, 12, 3093–3102. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
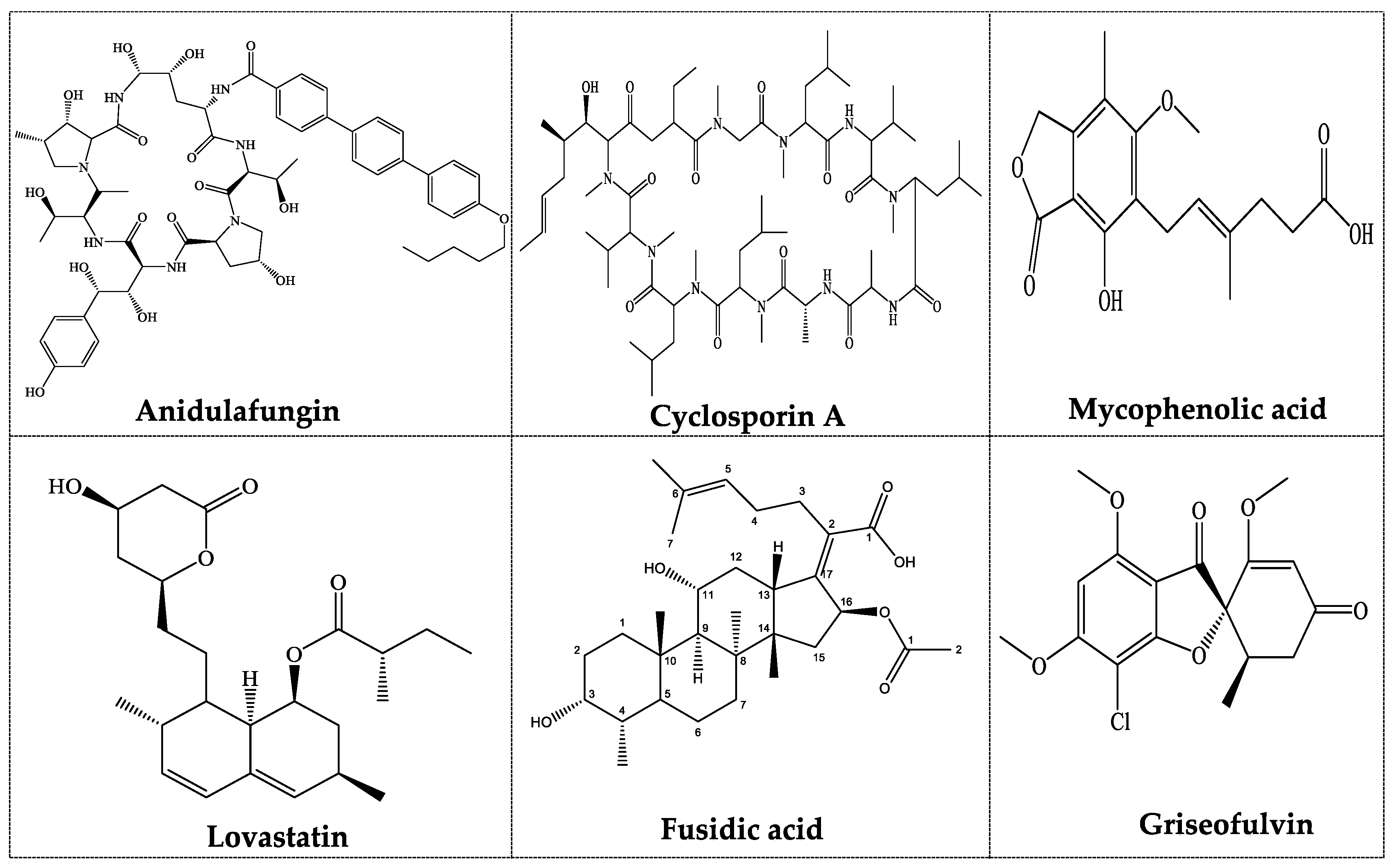
| Compound | Derived Product | Producing Fungus | Application | Reference |
|---|---|---|---|---|
| Compactin | Mevastatin | Penicillium compactum | Cholesterol-lowering agent | [66] |
| Mevacor | Lovastatin | Aspergillus terreus | Cholesterol-lowering agent | [48] |
| Pravastatin | Pravachol | Penicillium compactum Aspergillus spp., Monascus spp. | Cholesterol-lowering agent; Antithrombotic and anti-inflammatory agents | [3,9] |
| Compactin | ML-236B | Penicillium brevicompactum | Antifungal activity | [2] |
| Caspofungin | Cancidas | Glarea lozoyensis | Fungal infections | [18] |
| Anidulafungin | Anidulafungin | Aspergillus nidulans | Fungicidal | [10] |
| Strobilurins | Strobilurins | Stobilurus tenacellus | Agricultural fungicides | [22,50] |
| Taxol | Paclitaxol | Sporangium cellulosum | Antitumor and antifungal activity | [11,12] |
| Cryptocin | Cryptocin | Cryptosporiopsis quercina | Pesticide | [57] |
| Phomol | Phomol | Phomopsis sp. | Anti-inflammatory and antimicrobial activities | [59] |
| Penicillins | Penicillins | Penicillium notatum | Bacterial infections | [13] |
| Cephalosporins | Cephalosporins | Cephalosporium | Bacterial infections | [14] |
| Fusidic Acid | Fucidin | Fusidium coccineum | Bacterial infections | [15] |
| Cyclosporin A | Cyclosporin A | Tolypocladium inflatum | Immunosuppressive agent in organ transplantation | [16] |
| Mycophenolic Acid | Myfortic, CellCept | Penicillium stoloniferum | Immunosuppressive agent in organ transplantation | [17] |
| Fingolimod | Gilenya | Isaria sinclairii | Multiple sclerosis | [19,67] |
| Ergot | Ergotamine | Claviceps purpurea | Migraine headaches | [20,21] |
| Compound | Producing Bacteria | Application | Reference |
|---|---|---|---|
| Tigecycline | Streptomyces aureofaciens | Antibacterial for tetracycline-resistant organisms | [98] |
| Telithromycin | Saccharopolyspora erythraea | Antibacterial activity | [130] |
| Biapenem and Ertapenem | Streptomyces cattleya | Antibacterial activity | [131,132] |
| Everolimus | Streptomyces hygroscopicus | Immunosuppresive agent | [99] |
| Miglustat | Streptomyces lavendulae | Type 1 Gaucher disease | [133] |
| Daptomycin | Streptomyces roseosporus | Skin structure infection | [134] |
| Amrubicin | Streptomyces peuceticus | Antitumor activity | [135] |
| Pimecrolimus | Streptomyces hygroscopicus var ascomyceticus | Anti-inflammatory for skin disease | [135] |
| Gemtuzumab ozogamicin | Micromonospora echinospora sp. calichensis | Acute myeloid lymphoma treatment | [136] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Razek, A.S.; El-Naggar, M.E.; Allam, A.; Morsy, O.M.; Othman, S.I. Microbial Natural Products in Drug Discovery. Processes 2020, 8, 470. https://doi.org/10.3390/pr8040470
Abdel-Razek AS, El-Naggar ME, Allam A, Morsy OM, Othman SI. Microbial Natural Products in Drug Discovery. Processes. 2020; 8(4):470. https://doi.org/10.3390/pr8040470
Chicago/Turabian StyleAbdel-Razek, Ahmed S., Mehrez E. El-Naggar, Ahmed Allam, Osama M. Morsy, and Sarah I. Othman. 2020. "Microbial Natural Products in Drug Discovery" Processes 8, no. 4: 470. https://doi.org/10.3390/pr8040470