Production of Protein Hydrolysate Containing Antioxidant and Angiotensin -I-Converting Enzyme (ACE) Inhibitory Activities from Tuna (Katsuwonus pelamis) Blood

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Materials and Preparation
2.2. Enzymes and Chemicals
2.3. Preparation of TB Hydrolysate (TBH)
2.4. Determination of Degree of Hydrolysis (DH) and Yield
2.5. Antioxidant In Vitro Assays
2.5.1. DPPH Radical Scavenging Activity
2.5.2. ABTS Radical Scavenging Activity
2.5.3. Ferric Reducing Antioxidant Power (FRAP)
2.6. ACE Inhibitory Activity
2.7. Amino Acid Analysis
2.8. Fractionation of Hydrolysates
2.9. Sensory Evaluation of Hydrolysate and Fortified Fruit Jelly
2.10. Experimental Design and Statistical Analysis
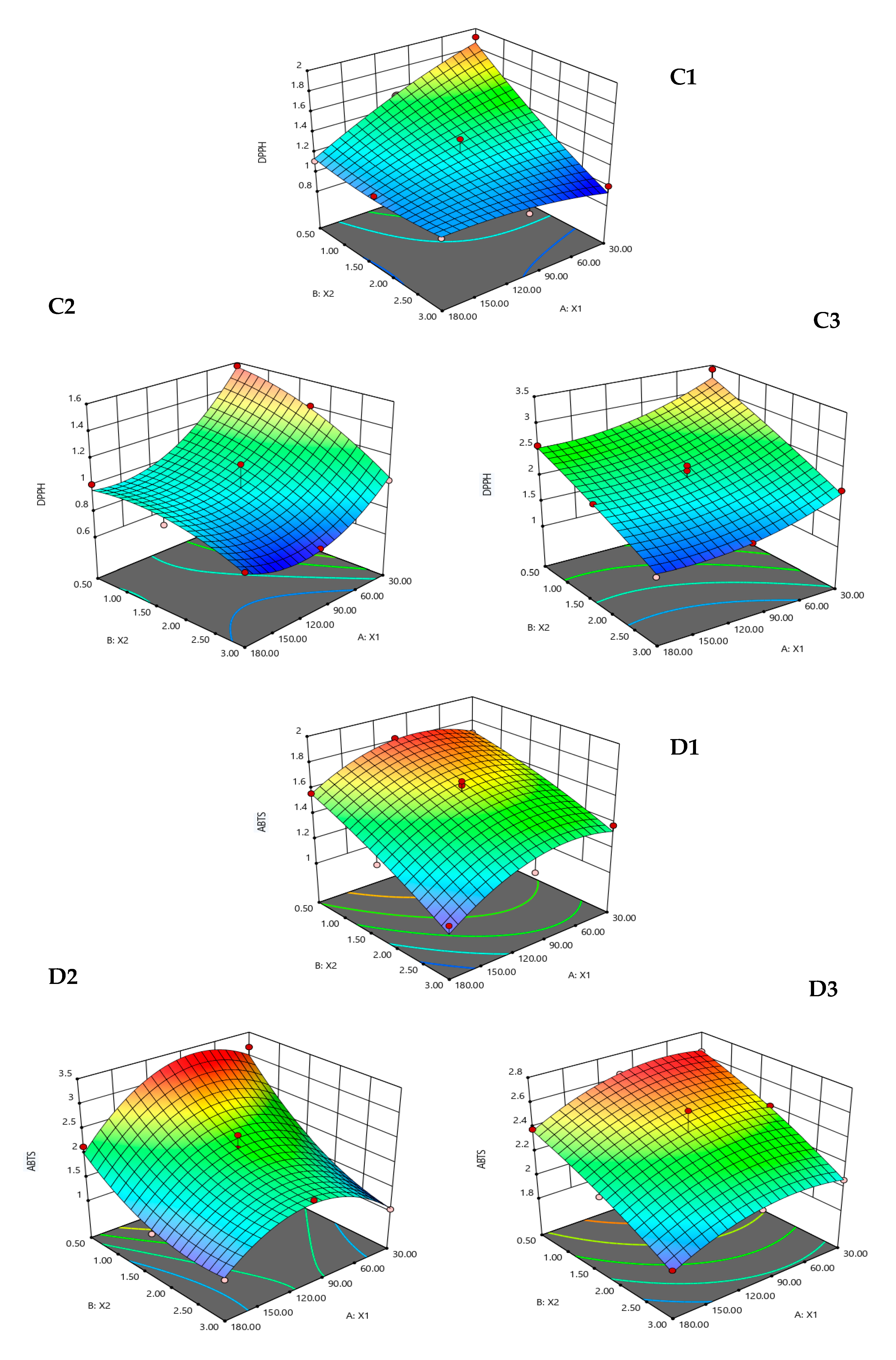
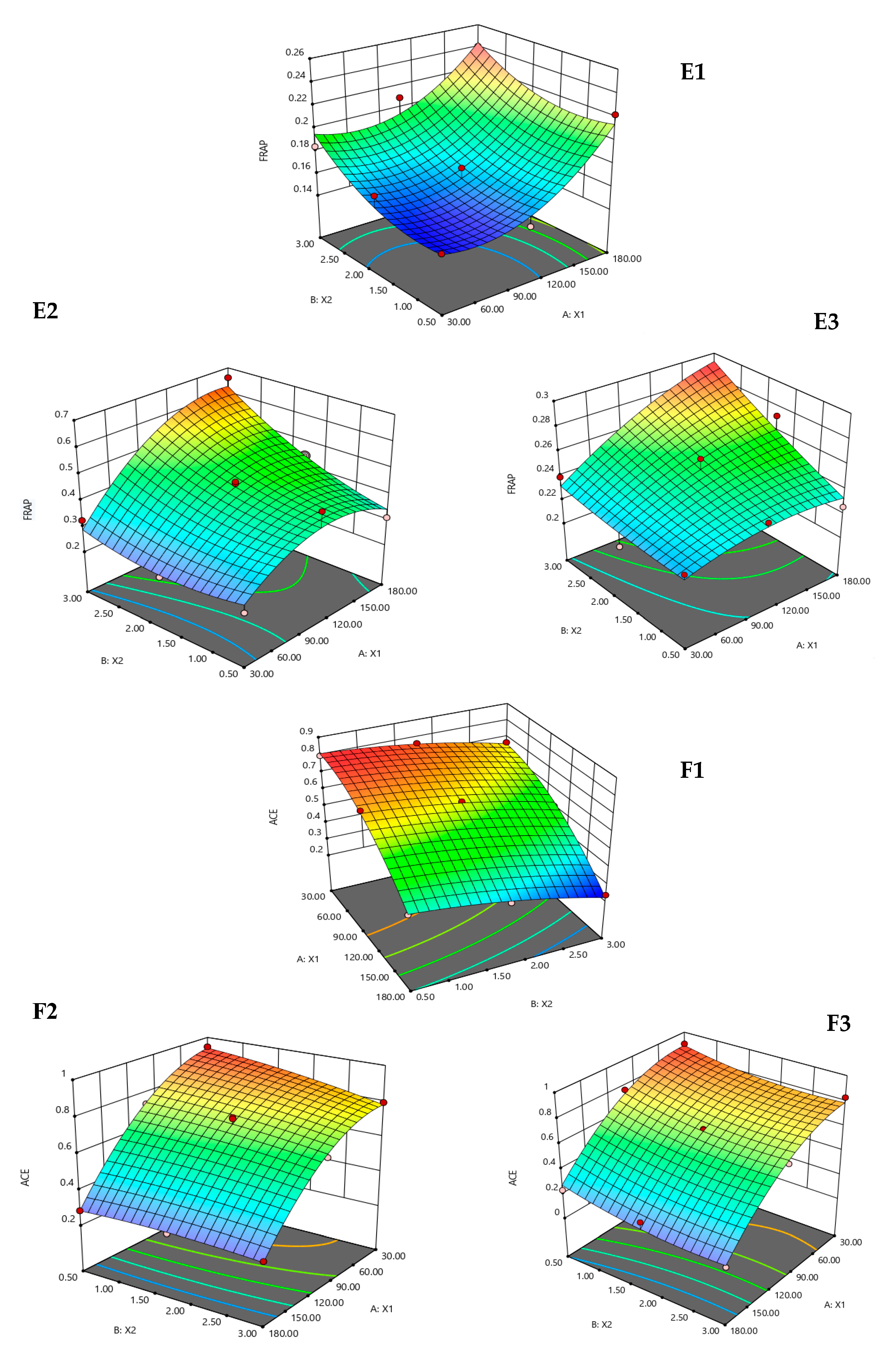
3. Results and Discussion
3.1. Fitting the Models
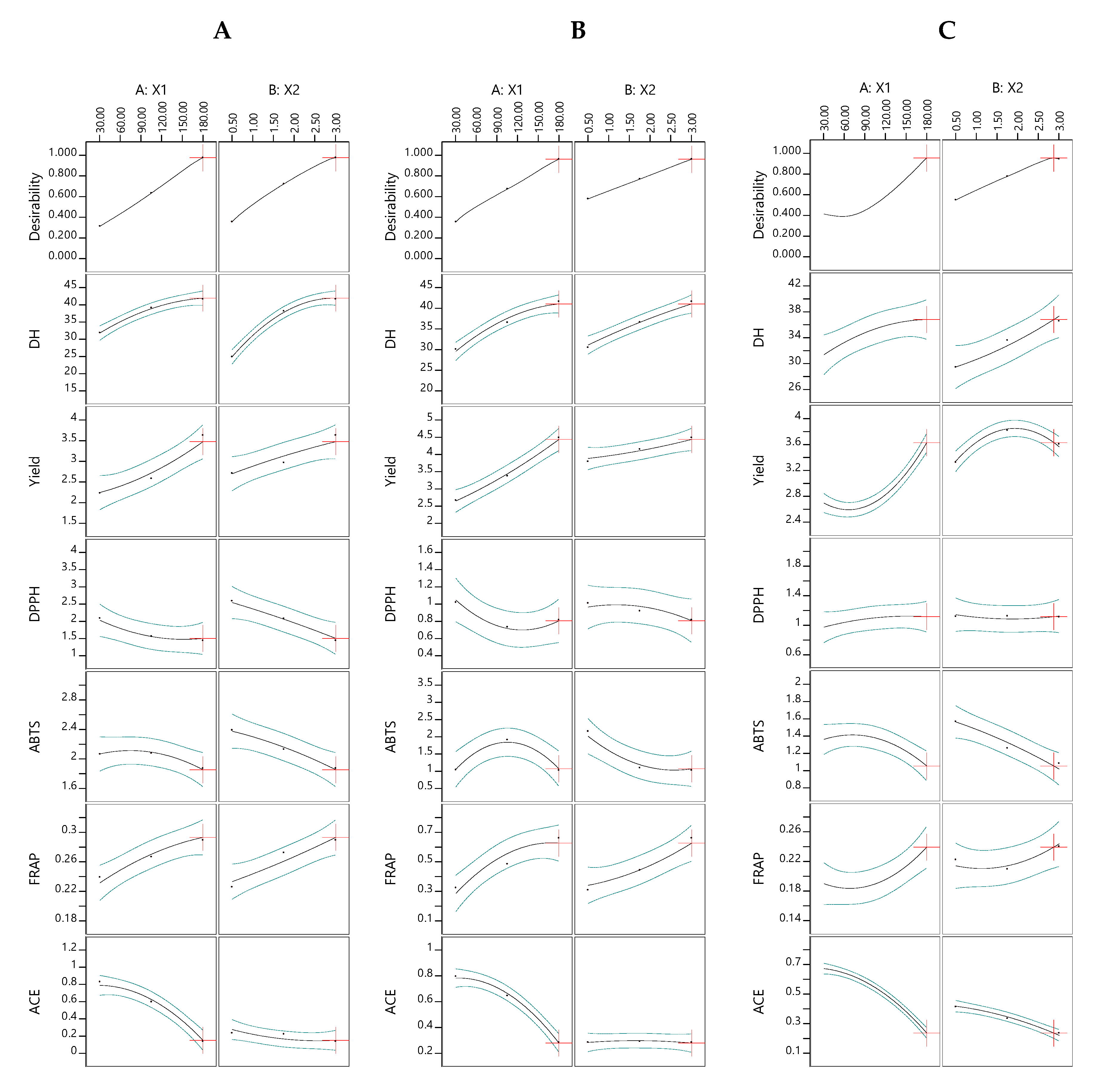
3.2. Effect of the Hydrolysis Conditions on the Responses
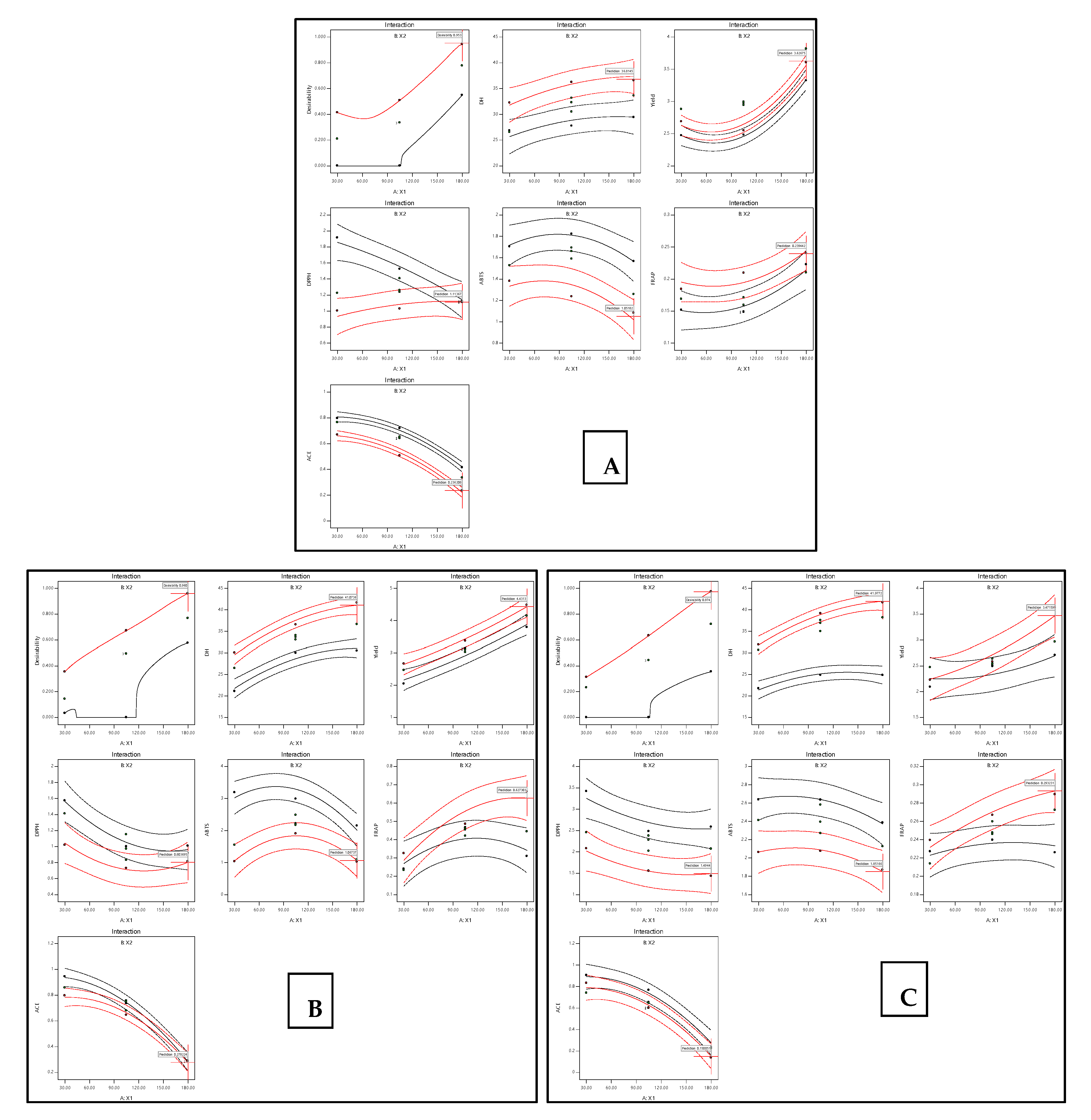
3.3. Optimization of TB Hydrolysis Conditions and Model Validation
3.4. Amino Acid Profile
3.5. Fractionation Using Ultrafiltration Membranes
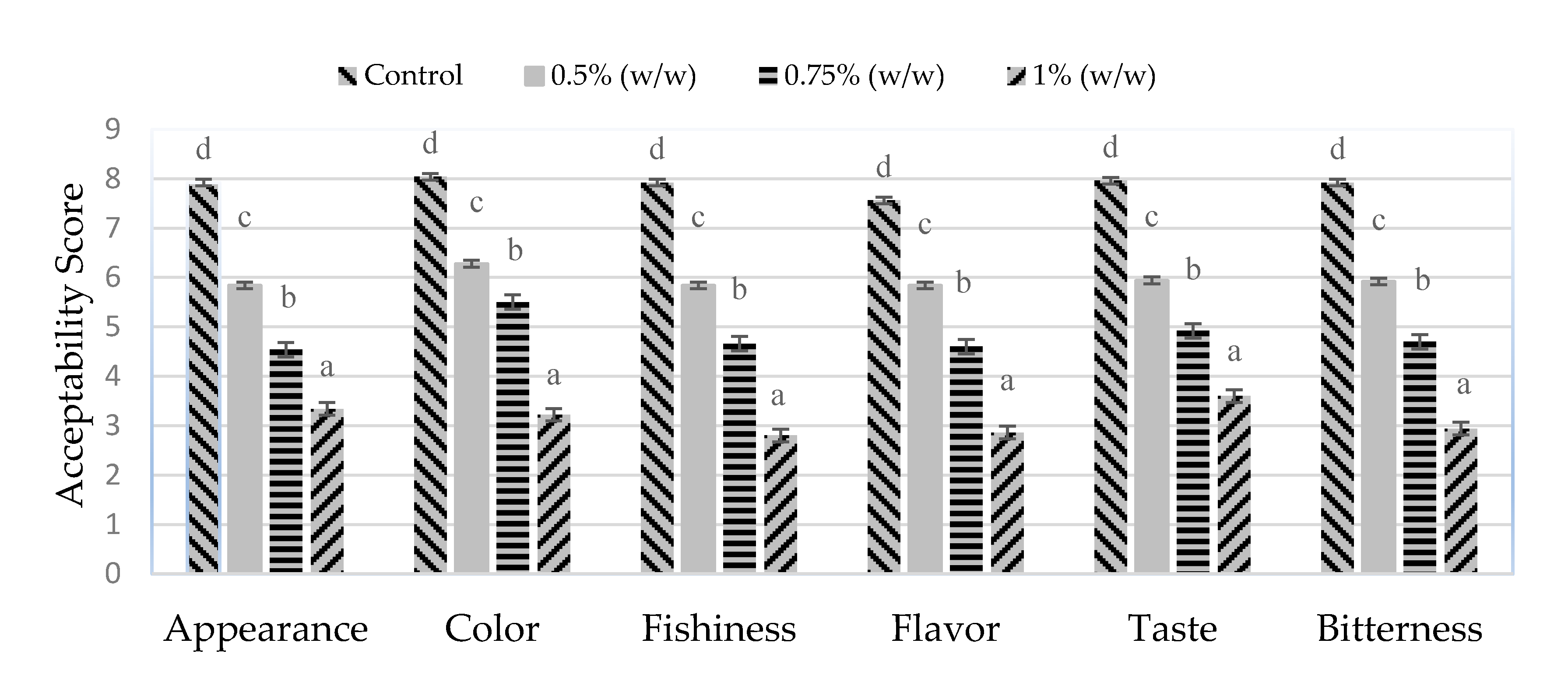
3.6. Sensory Evaluation of Hydrolysate and Fortified Mango Jelly
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mata, W.; Chanmalee, T.; Punyasuk, N.; Thitamadee, S. Simple PCR-RFLP detection method for genus-and species-authentication of four types of tuna used in canned tuna industry. Food Control. 2020, 18, 1211–1215. [Google Scholar] [CrossRef]
- Gamarro, E.G.; Worawattanamateekul, W.; Sentina, J.; Gopal, T.K.S. By-Products of Tuna Processing; GLOBEFISH Research Programme; FAO: Rome, Italy, 2013; Volume 112, 48p. [Google Scholar]
- Jelavic, K.M.; Stepanowska, K.; Grubisic, L.; Segvic Bubic, T.; Katavic, I. Reduced feeding effects to the blood and muscle chemistry of farmed juvenile bluefin tuna in the Adriatic Sea. Aquac. Res. 2012, 43, 317–320. [Google Scholar] [CrossRef]
- Hamzeh, A.; Wongngam, W.; Kiatsongchai, R.; Yongsawatdigul, J. Cellular and chemical antioxidant activities of chicken blood hydrolysates as affected by in vitro gastrointestinal digestion. Poult. Sci. 2019, 98, 6138–6148. [Google Scholar] [CrossRef] [PubMed]
- Wongngam, W.; Mitani, T.; Katayama, S.; Nakamura, S.; Yongsawatdigul, J. Production and characterization of chicken blood hydrolysate with antihypertensive properties. Poult. Sci. 2020, 99, 5163–5174. [Google Scholar] [CrossRef] [PubMed]
- Toldrà, M.; Parés, D.; Saguer, E.; Carretero, C. Hemoglobin hydrolysates from porcine blood obtained through enzymatic hydrolysis assisted by high hydrostatic pressure processing. Innov. Food Sci. Emerg. Technol. 2011, 12, 435–442. [Google Scholar] [CrossRef]
- Fu, Y.; Bak, K.H.; Liu, J.; Gobba, C.D.; Tøstesen, M.; Hansebn, E.T. Protein hydrolysates of porcine hemoglobin and blood: Peptide characteristics in relation to taste attributes and formation of volatile compounds. Food Res. Int. 2019, 121, 28–38. [Google Scholar] [CrossRef]
- Bah, C.S.F.; Carne, A.; McConnell, M.A.; Mros, S.; Bekhit, A.E.D.A. Production of bioactive peptide hydrolysates from deer, sheep, pig and cattle red blood cell fractions using plant and fungal protease preparations. Food Chem. 2016, 202, 458–466. [Google Scholar] [CrossRef]
- Ahn, C.; Lee, K.; Je, J. Enzymatic production of bioactive protein hydrolysates from tuna liver: Effects of enzymes and molecular weight on bioactivity. Int. J. Food Sci. Technol. 2010, 45, 562–568. [Google Scholar] [CrossRef]
- Hemker, A.M.; Nguyen, L.T.; Karwe, M.; Salvi, D. Effects of pressure-assisted enzymatic hydrolysis on functional and bioactive properties of tilapia (Oreochromis niloticus) by-product protein hydrolysates. LWT Food Sci. Technol. 2020, 122, 109003. [Google Scholar] [CrossRef]
- Rivero-Pino, F.; Espejo-Carpio, F.J.; Guadix, E.M. Evaluation of the bioactive potential of foods fortified with fish protein hydrolysates. Food Res. Int. 2020, 137, 109572. [Google Scholar] [CrossRef]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. J. Funct. Foods. 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.H.; Tavano, O.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R. Use of Alcalase in the production of bioactive peptides: A review. Int. J. Biol. Macromol. 2020, 165, 2143–2196. [Google Scholar] [CrossRef] [PubMed]
- Slizyte, R.; Rommi, K.; Mozuraityte, R.; Eck, P.; Five, K.; Rusta, T. Bioactivities of fish protein hydrolysates from defatted salmon backbones. Biotechnol. Rep. 2016, 11, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.K.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: A review. J. Funct. Foods. 2010, 2, 1–9. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, Proximate Analysis, 17th ed.; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Intarasirisawat, R.; Benjakul, S.; Visessanguan, W.; Wu, J. Antioxidative and functional properties of protein hydrolysate from defatted skipjack (Katsuwonous pelamis) roe. Food Chem. 2012, 135, 3039–3048. [Google Scholar] [CrossRef]
- Ahn, C.B.; Kim, J.G.; Je, J.Y. Purification and antioxidant properties of octapeptide from salmon byproduct protein hydrolysate by gastrointestinal digestion. Food Chem. 2014, 147, 78–83. [Google Scholar] [CrossRef]
- Wangtueai, S.; Siebenhandl-Ehn, S.; Haltrich, D. Optimization of the preparation of gelatin hydrolysates with antioxidant activity from lizardfish (Saurida spp.) scales gelatin. Chiang Mai J. Sci. 2016, 43, 68–79. [Google Scholar]
- Kasiwut, J.; Youravong, W.; Sirinupong, N. Angiotensin I-converting enzyme inhibitory peptides produced from tuna cooking juice hydrolysate by continuous enzymatic membrane reactor. Food Biochem. 2019, 43, e13058. [Google Scholar] [CrossRef]
- Yarnpakdee, S.; Benjakul, S.; Kristinsson, H.G.; Maqsood, S. Effect of pretreatment on lipid oxidation and fishy odour development in protein hydrolysates from the muscle of Indian mackerel. Food Chem. 2012, 135, 2474–2482. [Google Scholar] [CrossRef]
- Phanturat, P.; Benjakul, S.; Visessanguan, W.; Roytrakul, S. Use of pyloric caeca extract from bigeye snapper (Priacanthus macracanthus) for the production of gelatin hydrolysate with antioxidative activity. LWT Food Sci. Technol. 2010, 43, 86–97. [Google Scholar] [CrossRef]
- Yarnpakdee, S.; Benjakul, S.; Kristinsson, H.G.; Kishimura, H. Antioxidant and sensory properties of protein hydrolysate derived from Nile tilapia (Oreochromis niloticus) by one-and two-step hydrolysis. J. Food Sci. Technol. 2015, 52, 3336–3349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wangtueai, S.; Phimolsiripol, Y.; Vichasilp, C.; Regenstein, J.M.; Schöenlechner, R. Optimization of gluten-free functional noodles formulation enriched with fish gelatin hydrolysates. LWT Food Sci. Technol. 2020, 133, 109977. [Google Scholar] [CrossRef]
- Forghani, B.; Bakar, J.; Ebrahimpour, A.; Hamid, A.A. Enzyme hydrolysates from Stichopus horrens as a new source for angiotensin-I converting enzyme inhibitory peptides. Evid. Based Complement. Altern. Med. 2012, 10, 1155. [Google Scholar]
- Auwal, S.M.; Zarei, M.; Hamid, A.; Saari, N. Response surface optimisation for the production of Antioxidant hydrolysates from stone fish protein using bromelain. Evid. Based Complement. Altern. Med. 2017, 2017(4), 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bordbar, S.; Ebrahimpour, A.; Zarei, M.; Hamid, A.; Saari, N. Alcalase-generated proteolysates of stone fish (Actinopyga lecanora) flesh as a new source of antioxidant peptides. Int. Food Prop. 2018, 21, 1541–1559. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Shen, S.; Deng, J.; Li, T.; Ding, C. Antioxidant activities and functional properties of tea seed protein hydrolysates (Camellia oleifera Abel) influenced by the degree of enzymatic hydrolysis. Food Sci. Biotechnol. 2014, 23, 2075–2082. [Google Scholar] [CrossRef]
- Ghanbari, R.; Zarei, M.; Ebrahimpour, A.; Abdul-Hamid, A.; Ismail, A.; Saari, N. Angiotensin-I converting enzyme (ACE) inhibitory and anti-oxidant activities of sea cucumber (Actinopyga lecanora) hydrolysates. Int. J. Mol. Sci. 2015, 16, 28870–28885. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Zhao, M.; Shi, J.; Wang, J.; Jiang, Y.; Cui, C.; Kakuda, Y.; Xue, S.J. Purification and identification of antioxidant peptide from grass carp muscle hydrolysates by consecutive chromatography and electrospray ionization-mass spectrometry. Food Chem. 2008, 108, 727–736. [Google Scholar] [CrossRef]
- Tanuja, S.; Haridas, A.; Zynudheen, A.A.; Joshy, C.G. Functional and antioxidative properties of fish protein hydrolysate (FPH) produced from the frame meat of striped cat fish Pangasianodon hypophthalmus (Sauvage, 1878) using alkaline protease alcalase. Indiam. J. Fish. 2014, 61, 82–89. [Google Scholar]
- Dai, Z.Y.; Zhang, Y.P.; Zhang, H.; Lu, Y.B. Preparation and characterization of mussel (Mytilus edulis) protein hydrolysates with angiotensin-I-converting enzyme (ACE) inhibitory activity by enzymatic hydrolysis. J. Food Biochem. 2010, 36, 66–74. [Google Scholar] [CrossRef]
- Wu, H.C.; Chen, H.M.; Shiau, C.Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Nasri, R.; Jridi, M.; Lassoued, I.; Jemil, I.; Ben Slama-Ben Salem, R.; Nasri, M.; Karra-hâabouni, M. The influence of the extent of enzymatic hydrolysis on antioxidative properties and ACE-inhibitory activities of protein hydrolysates from goby (Zosterisessor ophiocephalus) muscle. Appl. Biochem. Biotechnol. 2014, 173, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- Lassoued, I.; Mora, L.; Nasri, R.; Jridi, M.; Toldrá, F.; Aristoy, M.C.; Barkia, A.; Nasri, M. Characterization and comparative assessment of antioxidant and ACE inhibitory activities of thornback ray gelatin hydrolysates. J. Funct. Foods. 2015, 13, 225–238. [Google Scholar] [CrossRef]
- Yang, X.R.; Qiu, Y.T.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Purification and characterization of antioxidant peptides derived from protein hydrolysate of the marine bivalve mollusk Tergillarca granosa. Mar. Drugs 2019, 17, 251. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.; Ahn, C.B.; Nam, K.H.; Kim, Y.K.; Yoon, N.Y.; Je, J.Y. Amino acid composition, antioxidant, and cytoprotective effect of blue mussel (Mytilus edulis) hydrolysate through the inhibition of Caspase-3 activation in oxidative stress-mediated endothelial cell injury. Mar. Drugs 2019, 17, 135. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Sun, J.; Tong, Z.; Lan, X.; Zhao, Z.; Liao, D. Optimization of hydrolysis conditions for the production of angiotensin-I converting enzyme-inhibitory peptides and isolation of a novel peptide from lizardfish (Saurida elongate) muscle protein hydrolysate. Mar. Drugs 2012, 10, 1066–1080. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO. Food and Agriculture Organization/World Health Organization; (FAO/WHO) Evaluation of Protein Quality; Joint Report; FAO Food Nutrition: Rome, Italy, 1991. [Google Scholar]
- Silva, V.M.; Park, K.J.; Hubinger, M.D. Optimization of the enzymatic hydrolysis of mussel meat. J. Food Sci. 2010, 75, C36–C42. [Google Scholar] [CrossRef]
- Zheng, Z.; Wei, X.; Shang, T.; Huang, Y.; Hu, C.; Zhang, R. Bioconversion of duck blood cell: Process optimization of hydrolytic conditions and peptide hydrolysate characterization. BMC Biotechnol. 2018, 18, 67. [Google Scholar] [CrossRef]
- He, X.Q.; Cao, W.H.; Pan, G.K.; Yang, L.; Zhang, C.H. Enzymatic hydrolysis optimization of Paphia undulata and lymphocyte proliferation activity of the isolated peptide fractions. J. Sci. Food Agric. 2015, 95, 1544–1553. [Google Scholar]
- Rho, S.J.; Lee, J.S.; Chung, Y.I.; Kim, Y.W.; Lee, H.G. Purification and identification of an angiotensin I-converting enzyme inhibitory peptide from fermented soybean extract. Process. Biochem. 2009, 44, 490–493. [Google Scholar] [CrossRef]
- Mirzaei, M.; Mirdamadi, S.; Ehsani, M.R.; Aminlari, M. Production of antioxidant and ACE-inhibitory peptides from Kluyveromyces marxianus protein hydrolysates: Purification and molecular docking. J. Food Drug Anal. 2017, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheih, I.C.; Fang, T.J.; Wu, T.K. Isolation and characterization of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the algae protein waste. Food Chem. 2009, 115, 279–284. [Google Scholar] [CrossRef]
- Auwa, S.M.; Abidin, N.Z.; Zarei, M.; Tan, C.P. Identification, structure-activity relationship and in silico molecular docking analyses of five novel angiotensin I-converting enzyme (ACE)-inhibitory peptides from stone fish (Actinopyga lecanora) hydrolysates. PLoS ONE 2019, 10, 1371. [Google Scholar]
- Gobbetti, M.; Ferranti, P.; Smacchi, E.; Goffredi, F.; Addeo, F. Production of angiotensin-I-converting-enzyme-inhibitory peptides in fermented milks started by Lactobacillus delbrueckii subsp. bulgaricus SS1 and Lactococcus lactis subsp. cremoris FT4. Appl. Environ. Microbiol. 2000, 9, 3898–3904. [Google Scholar] [CrossRef] [Green Version]
- Girgih, A.T.; Udenigwe, C.C.; Aluko, R.E. In vitro antioxidant properties of hemp seed (Cannabis sativa L.) potein hydrolysate fractions. J. Am. Oil Chem. Soc. 2011, 88, 381–389. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Jyothirmayi, T.; Diwan, P.V.; Kumar, B.D. Antioxidant activity and functional properties of enzymatic protein hydrolysates from common carp (Cyprinus carpio) roe egg. J. Food Sci. Technol. 2015, 52, 5817–5825. [Google Scholar] [CrossRef] [Green Version]
- Cheung, Y.W.; Li-Chan, E.C.Y. Enzymatic production of protein hydrolysates from steelhead (Oncorhynchus mykiss) skin gelatin as inhibitors of dipeptidyl-peptidase IV and angiotensin-I converting enzyme. J. Funct. Foods 2017, 28, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Tejano, L.A.; Peralta, J.P.; Yap, E.E.S.; Chang, Y.W. Bioactivities of enzymatic protein hydrolysates derived from Chlorella sorokiniana. Food Sci. Nutr. 2019, 7, 2381–2390. [Google Scholar] [CrossRef] [Green Version]
- Abuine, R.; Rathnayake, A.U.; Byun, H.G. Biological activity of peptides purified from fish skin hydrolysates. J. Fish. Aquat. Sci. 2019, 10, 1186. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Li, Z.R.; Chi, C.F.; Zhang, Q.H.; Luo, H.Y. Preparation and evaluation of antioxidant peptides from ethanol-soluble proteins hydrolysate of Sphyrna lewini muscle. Peptides 2012, 36, 240–250. [Google Scholar] [CrossRef]
- Ko, J.Y.; Kanga, N.; Lee, J.H.; Kim, J.S.; Kim, W.S.; Park, S.J.; Kim, Y.T.; Jeona, Y.J. Angiotensin I-converting enzyme inhibitory peptides from anenzymatic hydrolysate of flounder fish (Paralichthys olivaceus) muscleas a potent anti-hypertensive agent. Process. Biochem. 2016, 51, 535–541. [Google Scholar] [CrossRef]
- Nilsang, S.; Lertsiri, S.; Suphantharika, M.; Assavanig, A. Optimization of enzymatic hydrolysis of fish soluble concentrate by commercial proteases. J. Food Eng. 2005, 70, 571–578. [Google Scholar] [CrossRef]
- Benjakul, S.; Yarnpakdee, S.; Senphan, T.; Halldorsdottir, S.M.; Kristinsson, H.G. Fish protein hydrolysates: Production, bioactivities and applications. In Antioxidants and Functional Components in Aquatic Foods; Kristinsson, H.G., Ed.; Matil Ltd.: Reykjavik, Iceland, 2014; pp. 237–282. [Google Scholar]
- Begum, N.; Raza, A.; Shen, D.; Song, H.; Zhang, Y.; Zhang, L.; Liu, P. Sensory attribute and antioxidant capacity of Maillard reaction products from enzymatic hydrolysate of bovine bone marrow extract. Int. J. Food Sci. Technol. 2019, 57, 1786–1797. [Google Scholar] [CrossRef] [PubMed]
- Yarnpakdee, S.; Benjakul, S.; Nalinanon, S.; Kristinsson, H.G. Lipid oxidation and fishy odour development in protein hydrolysate from Nile tilapia (Oreochromis niloticus) muscle as affected by freshness and antioxidants. Food Chem. 2012, 132, 1781–1788. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Treatments | Factors | Responses | ||||||
|---|---|---|---|---|---|---|---|---|---|
| X1 (min) | X2 (%) | %DH | %Yield | DPPH (IC50) (mg/mL) | ABTS (IC50) (mg/mL) | FRAP (mmol FeSO4/g) | ACE (IC50) (mg/mL) | ||
| Alcalase | 1 | 180 | 0.50 | 29.4 a ± 0.2 | 3.3 b ± 0.1 | 1.11 a ± 0.03 | 1.6 c,d ± 0.1 | 0.22 a,b ± 0.01 | 0.41 c ± 0.04 |
| 2 | 105 | 1.75 | 33.2 b ± 0.2 | 2.9 a,b ± 0.3 | 1.4 b,c ± 0.2 | 1.7 d,e ± 0.1 | 0.15 a ± 0.01 | 0.64 e ± 0.00 | |
| 3 | 105 | 3.00 | 36.2 b ± 0.3 | 2.6 a ± 0.2 | 1.0 a ± 0.1 | 1.2 a,b ± 0.1 | 0.21 b ± 0.01 | 0.5 d ± 0.1 | |
| 4 | 105 | 1.75 | 30.5 a,b ± 0.1 | 3.0 a,b ± 0.2 | 1.3 a,b ± 0.2 | 1.7 d,e ± 0.1 | 0.16 a ± 0.01 | 0.65 e ± 0.00 | |
| 5 | 30 | 3.00 | 32.2 b ± 0.1 | 2.69 a ± 0.04 | 1.0 a ± 0.1 | 1.38 b,c ± 0.03 | 0.18 a,b ± 0.01 | 0.67 e ± 0.01 | |
| 6 | 30 | 0.50 | 26.8 a ± 0.01 | 2.5 a ± 0.2 | 1.9 d ± 0.1 | 1.7 d,e ± 0.1 | 0.15 a ± 0.01 | 0.80 g ± 0.02 | |
| 7 | 105 | 1.75 | 32.3 b ± 0.1 | 3.0 a,b ± 0.5 | 1.2 a,b ± 0.1 | 1.59 d ± 0.02 | 0.17 a ± 0.01 | 0.64 e ± 0.02 | |
| 8 | 180 | 3.00 | 36.6 b ± 0.3 | 3.6 b ± 0.1 | 1.1 a ± 0.3 | 1.1 a ± 0.1 | 0.24 b ± 0.01 | 0.24 a ± 0.01 | |
| 9 | 180 | 1.75 | 33.6 b ± 0.4 | 3.8 b,c ± 0.1 | 1.1 a ± 0.1 | 1.3 a,b ± 0.2 | 0.21 b ± 0.01 | 0.33 b ± 0.00 | |
| 10 | 105 | 0.50 | 27.7 a ± 0.2 | 2.5 a ± 0.3 | 1.52 c ± 0.00 | 1.8 e ± 0.1 | 0.15 a ± 0.01 | 0.7 e,f ± 0.1 | |
| 11 | 30 | 1.75 | 26.5 a ± 0.2 | 2.9 a ± 0.2 | 1.2 a,b ± 0.1 | 1.53 c,d ± 0.03 | 0.17 a ± 0.00 | 0.76 f,g ± 0.04 | |
| Neutrase | 1 | 180 | 0.50 | 30.5 b ± 0.2 | 3.8 b,c ± 0.2 | 1.0 b,c ± 0.1 | 2.2 d ± 0.2 | 0.31 b ± 0.01 | 0.28 a ± 0.00 |
| 2 | 105 | 1.75 | 33.6 b,c ± 0.2 | 3.1 b ± 0.1 | 0.8 a,b ± 0.1 | 2.2 d ± 0.2 | 0.42 c ± 0.04 | 0.76 c ± 0.00 | |
| 3 | 105 | 3.00 | 36.6 b,c ± 0.1 | 3.4 b,c ± 0.6 | 0.7 a ± 0.1 | 1.9 c ± 0.2 | 0.49 c ± 0.00 | 0.65 b ± 0.01 | |
| 4 | 105 | 1.75 | 33.1 b,c ± 0.2 | 3.1 b ± 0.2 | 1.0 a,b,c ± 0.1 | 2.5 f ± 0.1 | 0.5 c ± 0.1 | 0.68 b ± 0.00 | |
| 5 | 30 | 3.00 | 30.0 b ± 0.2 | 2.7 a,b ± 0.1 | 1.0 b,c ± 0.1 | 1.04 a ± 0.02 | 0.3 b ± 0.1 | 0.80 d ± 0.03 | |
| 6 | 30 | 0.50 | 21.1 a ± 0.3 | 2.0 a ± 0.3 | 1.57 d ± 0.04 | 3.2 g ± 0.1 | 0.24 a ± 0.01 | 0.94 f ± 0.00 | |
| 7 | 105 | 1.75 | 34.0 b,c ± 0.1 | 3.0 b ± 0.1 | 1.2 c ± 0.1 | 2.2 d ± 0.1 | 0.46 c ± 0.01 | 0.75 c ± 0.03 | |
| 8 | 180 | 3.00 | 41.7 d ± 0.3 | 4.5 c,d ± 0.1 | 0.8 a,b ± 0.3 | 1.0 a ± 0.1 | 0.66 d ± 0.02 | 0.28 a ± 0.00 | |
| 9 | 180 | 1.75 | 36.6 b,c ± 0.1 | 4.1 c ± 0.3 | 0.9 a,b,c ± 0.1 | 1.09 a ± 0.03 | 0.44 c ± 0.03 | 0.29 a ± 0.00 | |
| 10 | 105 | 0.50 | 30.0 b ± 0.3 | 3.1 b ± 0.6 | 1.0 b,c ± 0.1 | 3.0 g ± 0.1 | 0.47 c ± 0.04 | 0.74 c ± 0.00 | |
| 11 | 30 | 1.75 | 26.5 b ± 0.2 | 2.5 a,b ± 0.2 | 1.4 d ± 0.1 | 1.6 b ± 0.1 | 0.23 a ± 0.04 | 0.9 e ± 0.1 | |
| Flavourzyme | 1 | 180 | 0.50 | 24.8 a ± 0.1 | 2.7 b,c ± 0.2 | 2.6 c ± 0.1 | 2.4 c,d,e ± 0.1 | 0.23 a,b ± 0.02 | 0.23 b ± 0.01 |
| 2 | 105 | 1.75 | 36.9 b ± 0.5 | 2.6 b,c ± 0.1 | 2.3 c ± 0.4 | 2.3 b,c,d ± 0.1 | 0.25 a,b,c ± 0.03 | 0.60 c ± 0.01 | |
| 3 | 105 | 3.00 | 39.2 b,c ± 0.1 | 2.6 b,c ± 0.4 | 1.6 a ± 0.1 | 2.1 a,b ± 0.1 | 0.27 b,c ± 0.05 | 0.60 c ± 0.00 | |
| 4 | 105 | 1.75 | 37.5 b,c ± 0.1 | 2.5 b,c ± 0.2 | 2.98 b,c ± 0.04 | 2.39 c,d,e ± 0.02 | 0.25 a,b,c ± 0.00 | 0.65 d ± 0.01 | |
| 5 | 30 | 3.00 | 31.9 b ± 0.2 | 2.2 b ± 0.1 | 2.1 b ± 0.1 | 2.1 a,b ± 0.1 | 0.24 a,b ± 0.01 | 0.83 g ± 0.02 | |
| 6 | 30 | 0.50 | 21.7 a ± 0.3 | 2.1 a ± 0.2 | 3.4 d ± 0.2 | 2.6 e ± 0.2 | 0.23 a,b ± 0.03 | 0.90 h ± 0.00 | |
| 7 | 105 | 1.75 | 35 b ± 0.5 | 2.5 b,c ± 0.3 | 2.0 b ± 0.2 | 2.6 d,e ± 0.1 | 0.26 a,b,c ± 0.02 | 0.65 d ± 0.00 | |
| 8 | 180 | 3.00 | 41.7 c ± 0.4 | 3.6 c ± 0.1 | 1.4 a ± 0.3 | 1.9 a ± 0.3 | 0.29 c ± 0.00 | 0.14 a ± 0.00 | |
| 9 | 180 | 1.75 | 38.2 b,c ± 0.4 | 3.0 c ± 0.1 | 2.1 b ± 0.1 | 2.1 a,b ± 0.3 | 0.27 b,c ± 0.01 | 0.22 b ± 0.00 | |
| 10 | 105 | 0.50 | 24.9 a ± 0.2 | 2.5 b,c ± 0.1 | 2.5 c ± 0.1 | 2.6 e ± 0.1 | 0.24 a,b ± 0.03 | 0.77 f ± 0.00 | |
| 11 | 30 | 1.75 | 30.7 b ± 0.3 | 2.5 b,c ± 0.1 | 2.5 c ± 0.1 | 2.4 c,d,e ± 0.3 | 0.21 a ± 0.00 | 0.74 e ± 0.01 | |
| Hydrolysates | Responses | Quadratic Polynomial Model | R2 | p-Value |
|---|---|---|---|---|
| Alcalase | %DH | Y1 = 23.1 + 0.07X1 + 0.88X2 − 0.0002X12 + 0.41X22 − 0.01X1X2 | 0.9132 | 0.0110 |
| %Yield | Y2 = 2.33 − 0.01X1 + 0.92X2 + 0.0001X12 − 0.25X22 + 0.0002X1X2 | 0.9881 | <0.0001 | |
| DPPH (IC50) (mg/mL) | Y3 = 2.22 − 0.004X1 − 0.55X2 − 0.00001X12 + 0.03X22 + 0.002X1X2 | 0.9304 | 0.0064 | |
| ABTS (IC50) (mg/mL) | Y4 = 1.62 + 0.01X1 − 0.06X − 0.00003X12 − 0.02X22 − 0.0004X1X2 | 0.9377 | 0.0049 | |
| FRAP (mmol FeSO4/g) | Y5 = 0.17 − 0.001X1 − 0.01X2 + 0.000004X12 + 0.01X22 − 0.00004X1X2 | 0.9151 | 0.0104 | |
| ACE (IC50) (mg/mL) | Y6 = 0.802 + 0.0008X1 + 0.004X − 0.00002X12 − 0.017X22 − 0.0001X1X2 | 0.9956 | <0.0001 | |
| Neutrase | %DH | Y1 = 15.8 + 0.14X1 + 4.11X2 − 0.0004X12 − 0.33X22 + 0.01X1X2 | 0.9846 | 0.0002 |
| %Yield | Y2 = 1.88 + 0.01X1 + 0.07X2 + 0.00002X12 + 0.03X22 + 0.0002X1X2 | 0.9804 | 0.0003 | |
| DPPH (IC50) (mg/mL) | Y3 =1.92 − 0.01X1 − 0.04X2 + 0.00004X12 − 0.06X22 + 0.001X1X2 | 0.9050 | 0.0136 | |
| ABTS (IC50) (mg/mL) | Y4 = 3.26 + 0.02X1 − 1.66X2 − 0.0001X12 + 0.23X22 + 0.003X1X2 | 0.9558 | 0.0021 | |
| FRAP (mmol FeSO4/g) | Y5 = 0.206 + 0.004X1 − 0.100X2 − 0.00002X12 + 0.025X22 + 0.0007X1X2 | 0.9074 | 0.0128 | |
| ACE (IC50) (mg/mL) | Y6 = 0.96 + 0.0004X1 − 0.04X2 − 0.00002X12 − 0.01X22 + 0.0004X1X2 | 0.9915 | <0.0001 | |
| Flavourzyme | %DH | Y1= 12.7 + 0.09X1 + 13.5X2 − 0.0004X12 − 2.83X22 + 0.02X1X2 | 0.9906 | 0.0001 |
| %Yield | Y2 = 2.29 − 0.003X1 + 0.03X2 − 0.00003X12 − 0.03X22 + 0.002X1X2 | 0.9007 | 0.0151 | |
| DPPH (IC50) (mg/mL) | Y3 = 3.82 − 0.01X1 − 0.43X2 + 0.00004X12 − 0.02X22 + 0.001X1X2 | 0.9265 | 0.0073 | |
| ABTS (IC50) (mg/mL) | Y4 = 2.65 + 0.003X1 − 0.14X2 − 0.00002X12 − 0.03X22 + 0.0002X1X2 | 0.9218 | 0.0085 | |
| FRAP (mmol FeSO4/g) | Y5 = 0.216 + 0.0003X1 − 0.01X2 − 0.000001X12 + 0.001X22 + 0.0001X1X2 | 0.8927 | 0.0182 | |
| ACE (IC50) (mg/mL) | Y6 = 0.93 + 0.002X1 − 0.15X2 − 0.00003X12 + 0.03X22 − 0.0001X1X2 | 0.9811 | 0.0003 |
| Hydrolysates | Value | Responses | |||||||
|---|---|---|---|---|---|---|---|---|---|
| X1 (min) | X2 (%) | %DH | %Yield | DPPH (mg/mL) | ABTS (mg/mL) | FRAP (mmol FeSO4/g) | ACE (mg/mL) | ||
| TBHA | Predicated value | 36.58 | 3.65 | 1.11 | 1.07 | 0.24 | 0.24 | ||
| Experimental value | 180 | 2.81 | 40.4 a ± 1 | 3.61 b ± 0.03 | 1.16 b ± 0.03 | 1.0 a ± 0.1 | 0.28 a ± 0.01 | 0.26 b ± 0.02 | |
| Composite desirability | 0.95 | ||||||||
| TBHN | Predicated value | 40.73 | 4.40 | 0.82 | 1.05 | 0.60 | 0.28 | ||
| Experimental value | 180 | 2.89 | 44 b ± 3 | 4.6 c ± 0.1 | 0.84 a ± 0.01 | 1.07 a ± 0.03 | 0.65 c ± 0.01 | 0.28 b ± 0.01 | |
| Composite desirability | 0.96 | ||||||||
| TBHF | Predicated value | 41.96 | 3.44 | 1.56 | 1.89 | 0.29 | 0.15 | ||
| Experimental value | 180 | 2.87 | 46 c ± 3 | 3.5 a ± 0.2 | 1.6 c ± 0.1 | 1.4 b ± 0.1 | 0.31 b ± 0.01 | 0.19 a ± 0.00 | |
| Composite desirability | 0.96 | ||||||||
| Amino Acids | g/100 g of TBHN | Essential Amino Acid Score | FAO/WHO (1991) |
|---|---|---|---|
| Alanine | 2.12 | - | - |
| Aspartic acid + Asparagine | 2.92 | - | - |
| Cystine | <0.02 | - | - |
| Glutamic acid + Glutamine | 3.71 | - | - |
| Glycine | 1.20 | - | - |
| Histidine | 4.03 | 2.26 | 1.9 |
| Hydroxylysine | <0.02 | - | - |
| Hydroxyproline | <0.02 | - | - |
| Isoleucine | 9.21 | 3.29 | 2.8 |
| Leucine | 6.50 | 0.98 | 6.6 |
| Lysine | 9.25 | 1.60 | 5.8 |
| Methionine | <0.02 | <0.01 | 2.5 |
| Phenylalanine | 4.68 | 0.74 | 6.3 |
| Proline | 1.62 | - | - |
| Serine | 0.54 | - | - |
| Threonine | 0.76 | 0.22 | 3.4 |
| Tryptophan | 0.11 | 0.10 | 1.1 |
| Tyrosine | 2.54 | - | - |
| Valine | 8.71 | 2.49 | 3.5 |
| Hydrophobic amino acids | 35.6 | - | - |
| Hydrophilic amino acids | 22.4 | - | - |
| Total amino acids | 57.9 | - | - |
| Molecular Weight (kDa) | DPPH (IC50) (mg/mL) | ABTS (IC50) (mg/mL) | FRAP (mmol FeSO4/g) | ACE (IC50) (mg/mL) | Yield (%) |
|---|---|---|---|---|---|
| >10 | 39 c ± 8 | 0.51 d ± 0.03 | 0.40 a ± 0.04 | 2.7 d ± 0.1 | 39.7 d ± 0.2 |
| 3–10 | 14 b ± 2 | 0.37 c ± 0.03 | 0.45 a ± 0.04 | 2.0 c ± 0.1 | 30.0 c ± 0.1 |
| 1–3 | 12 b ± 1 | 0.29 b ± 0.02 | 0.64 b ± 0.00 | 0.73 b ± 0.02 | 14 b ± 1 |
| <1 | 6.0 a ± 0.3 | 0.13 a ± 0.01 | 0.70 b,c ± 0.01 | 0.37 a ± 0.01 | 5.23 a ± 0.01 |
| Enzymes | Concentration (%) | Score | ||
|---|---|---|---|---|
| Fishy Odor | Fishy Flavor | Bitterness | ||
| Reference | 10 | 15 | 15 | 15 |
| Alcalase | 0.5 | 9.5 b ± 1.0 | 9.4 a ± 0.6 | 6.8 a ± 3.3 |
| 1.0 | 9.9 c ± 3.6 | 11.2 e ± 0.8 | 7.3 b ± 3.9 | |
| Neutrase | 0.5 | 10 e ± 1 | 9.5 b ± 1.0 | 7.3 b ± 3.5 |
| 1.0 | 11 f ± 1 | 11 d ± 1 | 8.2 d ± 3.4 | |
| Flavourzyme | 0.5 | 9.4 a ± 1.4 | 9.9 c ± 1.4 | 7.6 c ± 3.1 |
| 1.0 | 11 d ± 2 | 12 f ± 1 | 8.8 e ± 3.2 | |
| Concentration (%) | DPPH (IC50) (mg/mL) | ABTS (IC50) (mg/mL) | FRAP (mmol FeSO4/g) | ACE (IC50) (mg/mL) |
|---|---|---|---|---|
| Control | 8.7 d ± 0.3 | 9.7 c ± 0.4 | 0.15 d ± 0.00 | ND |
| 0.5 | 4.54 c ± 0.02 | 0.48 b ± 0.03 | 0.65 a ± 0.03 | 3.7 c ± 0.1 |
| 0.75 | 4.1 b ± 0.1 | 0.46 b ± 0.02 | 0.72 b ± 0.02 | 2.7 b ± 0.1 |
| 1 | 3.9 a ± 0.1 | 0.37 a ± 0.01 | 0.77 c ± 0.01 | 1.3 a ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mongkonkamthorn, N.; Malila, Y.; Yarnpakdee, S.; Makkhun, S.; Regenstein, J.M.; Wangtueai, S. Production of Protein Hydrolysate Containing Antioxidant and Angiotensin -I-Converting Enzyme (ACE) Inhibitory Activities from Tuna (Katsuwonus pelamis) Blood. Processes 2020, 8, 1518. https://doi.org/10.3390/pr8111518
Mongkonkamthorn N, Malila Y, Yarnpakdee S, Makkhun S, Regenstein JM, Wangtueai S. Production of Protein Hydrolysate Containing Antioxidant and Angiotensin -I-Converting Enzyme (ACE) Inhibitory Activities from Tuna (Katsuwonus pelamis) Blood. Processes. 2020; 8(11):1518. https://doi.org/10.3390/pr8111518
Chicago/Turabian StyleMongkonkamthorn, Natthaphon, Yuwares Malila, Suthasinee Yarnpakdee, Sakunkhun Makkhun, Joe M. Regenstein, and Sutee Wangtueai. 2020. "Production of Protein Hydrolysate Containing Antioxidant and Angiotensin -I-Converting Enzyme (ACE) Inhibitory Activities from Tuna (Katsuwonus pelamis) Blood" Processes 8, no. 11: 1518. https://doi.org/10.3390/pr8111518