

Process Intensification in Human Pluripotent Stem Cell Expansion with Microcarriers

Abstract
:1. Introduction
2. Materials and Methods
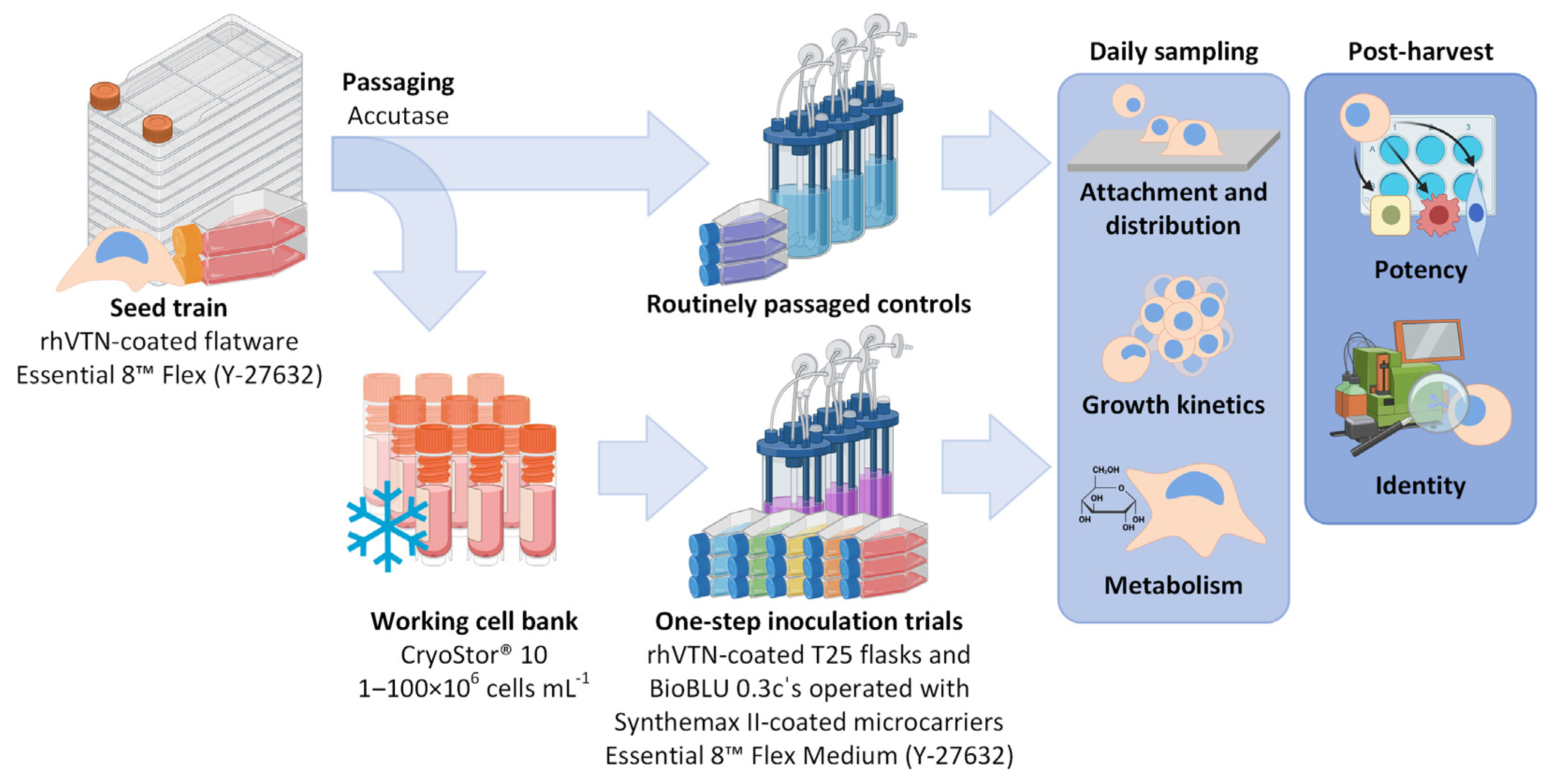
2.1. Cell Line, Cultivation and Cell Banking
2.2. Cell Counting and Medium Component Analysis
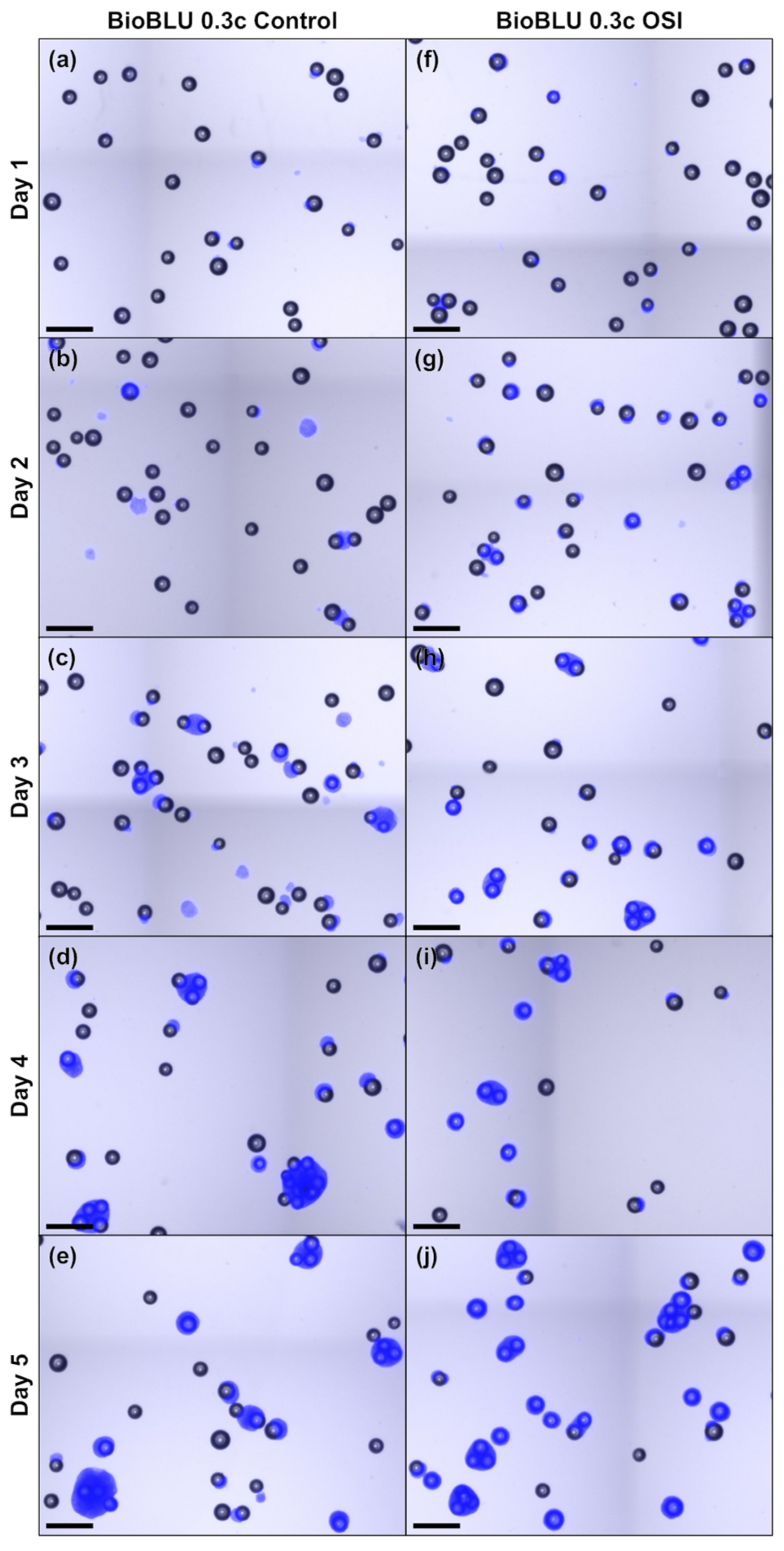
2.3. Microcarrier Aggregate Analysis
2.4. Analysis of Cell Quality and Potency
2.5. Statistical Analysis
3. Results and Discussion
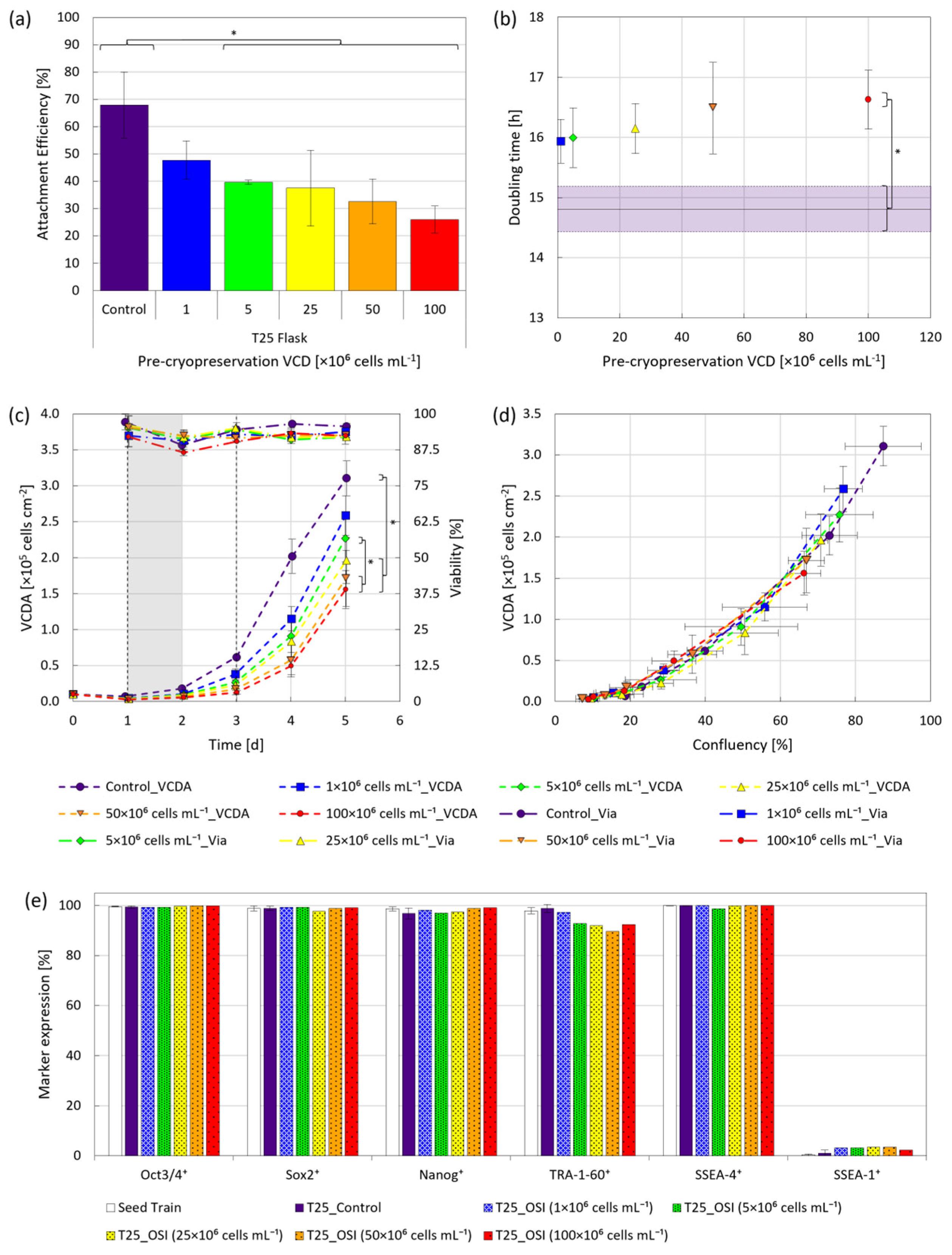
3.1. One-Step Inoculation under Static Conditions
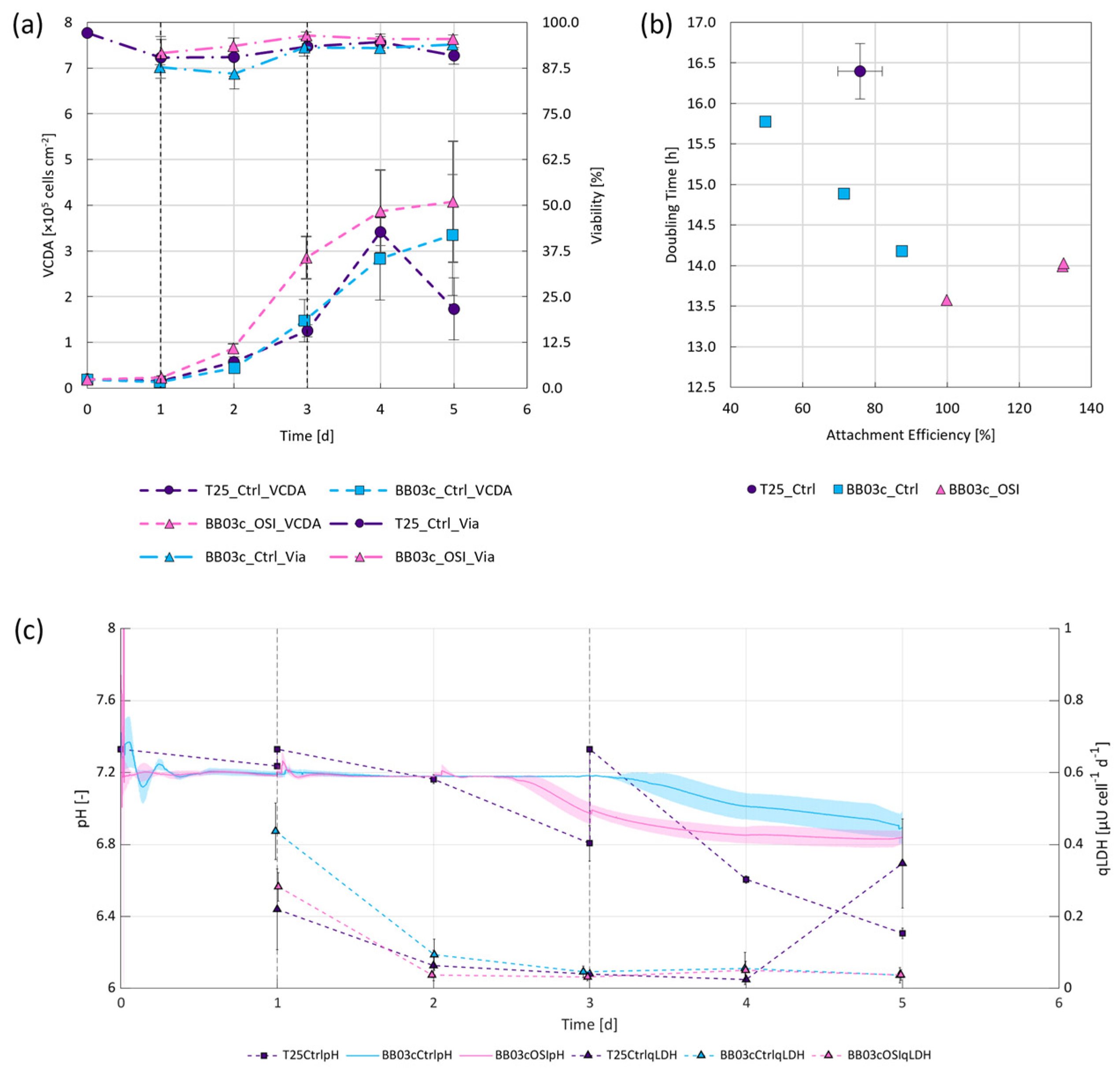
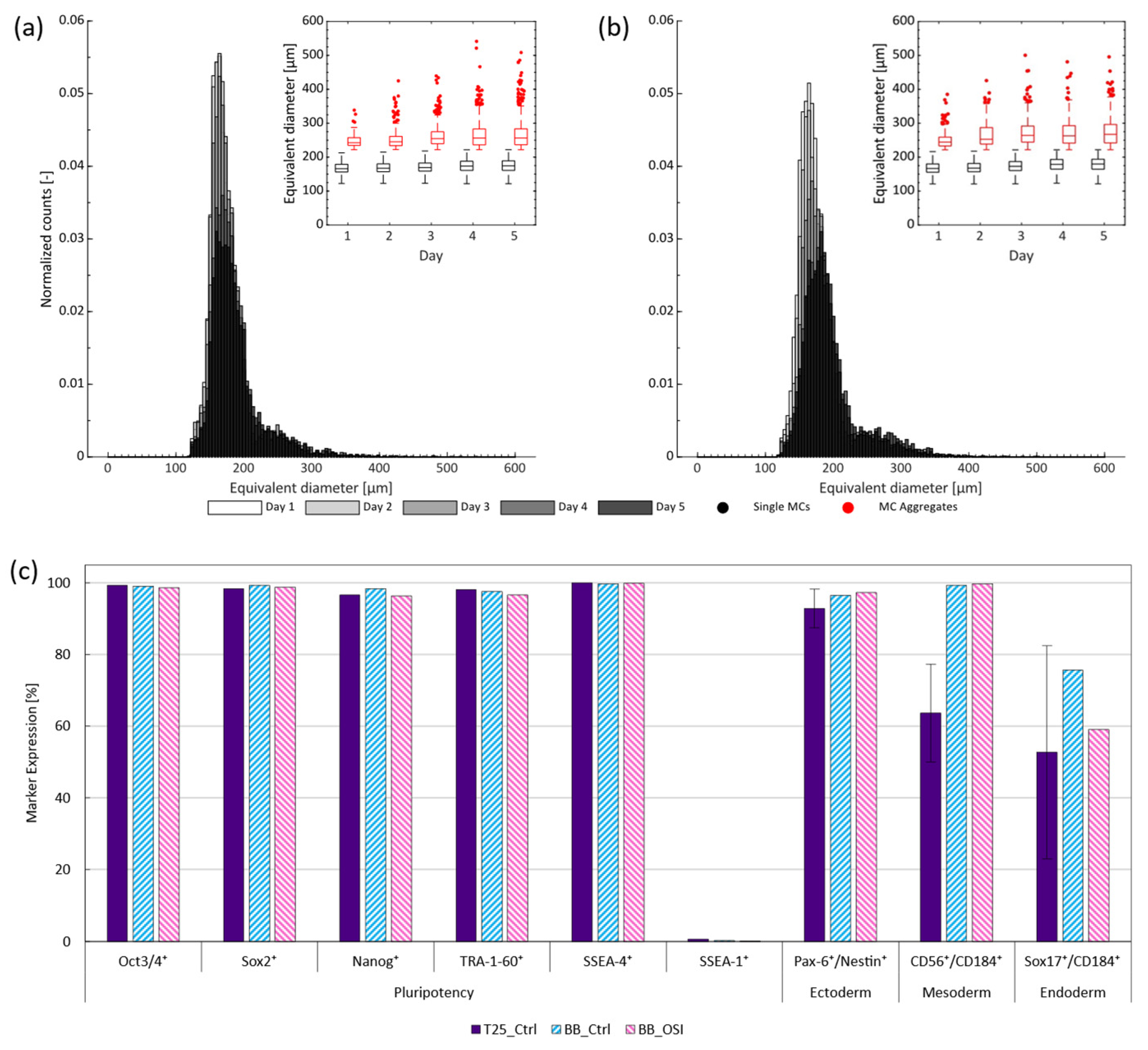
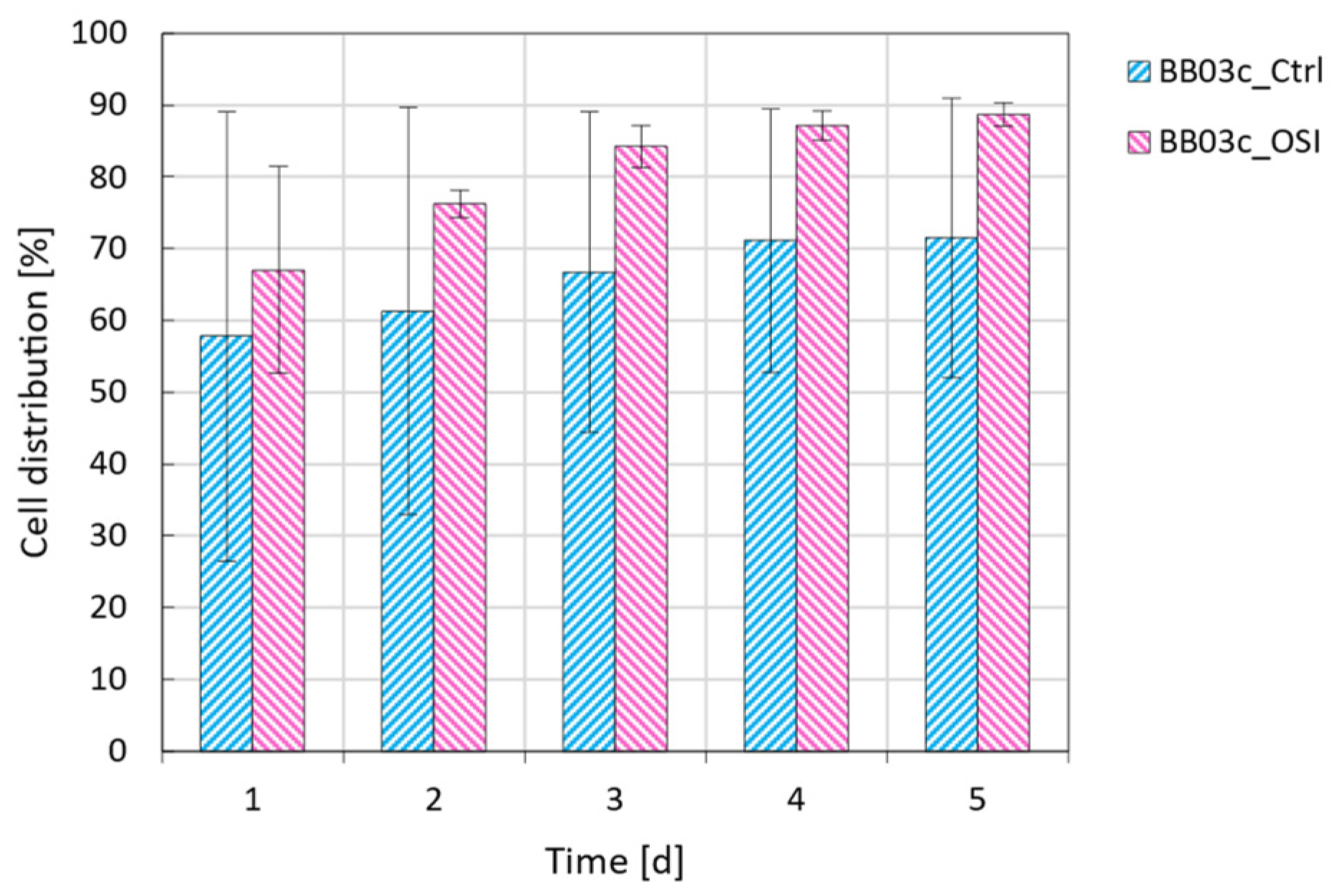
3.2. One-Step Inoculation of Microcarrier-Operated Stirred Bioreactors
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A



References
- Inoue, M.; Yamaguchi, R.; He, C.C.J.; Ikeda, A.; Okano, H.; Kohyama, J. Current Status and Prospects of Regenerative Medicine for Spinal Cord Injury Using Human Induced Pluripotent Stem Cells: A Review. Stem Cell Investig. 2023, 10, 6. [Google Scholar] [CrossRef]
- Tomasik, J.; Jasiński, M.; Basak, G.W. Next Generations of CAR-T Cells—New Therapeutic Opportunities in Hematology? Front. Immunol. 2022, 13, 1034707. [Google Scholar] [CrossRef]
- Li, Y.; Hao, J.; Hu, Z.; Yang, Y.-G.; Zhou, Q.; Sun, L.; Wu, J. Current Status of Clinical Trials Assessing Mesenchymal Stem Cell Therapy for Graft versus Host Disease: A Systematic Review. Stem Cell Res. Ther. 2022, 13, 93. [Google Scholar] [CrossRef]
- Scibona, E.; Morbidelli, M. Expansion Processes for Cell-Based Therapies. Biotechnol. Adv. 2019, 37, 107455. [Google Scholar] [CrossRef]
- Hogrebe, N.J.; Maxwell, K.G.; Augsornworawat, P.; Millman, J.R. Generation of Insulin-Producing Pancreatic β Cells from Multiple Human Stem Cell Lines. Nat. Protoc. 2021, 16, 4109–4143. [Google Scholar] [CrossRef]
- Gunhanlar, N.; Shpak, G.; Van Der Kroeg, M.; Gouty-Colomer, L.A.; Munshi, S.T.; Lendemeijer, B.; Ghazvini, M.; Dupont, C.; Hoogendijk, W.J.G.; Gribnau, J.; et al. A Simplified Protocol for Differentiation of Electrophysiologically Mature Neuronal Networks from Human Induced Pluripotent Stem Cells. Mol. Psychiatry 2018, 23, 1336–1344. [Google Scholar] [CrossRef]
- Weed, L.S.; Mills, J.A. Strategies for Retinal Cell Generation from Human Pluripotent Stem Cells. Stem Cell Investig. 2017, 4, 65. [Google Scholar] [CrossRef]
- Laco, F.; Lam, A.T.-L.; Woo, T.-L.; Tong, G.; Ho, V.; Soong, P.-L.; Grishina, E.; Lin, K.-H.; Reuveny, S.; Oh, S.K.-W. Selection of Human Induced Pluripotent Stem Cells Lines Optimization of Cardiomyocytes Differentiation in an Integrated Suspension Microcarrier Bioreactor. Stem Cell Res. Ther. 2020, 11, 118. [Google Scholar] [CrossRef]
- Cuesta-Gomez, N.; Verhoeff, K.; Dadheech, N.; Dang, T.; Jasra, I.T.; de Leon, M.B.; Pawlick, R.; Marfil-Garza, B.; Anwar, P.; Razavy, H.; et al. Suspension Culture Improves iPSC Expansion and Pluripotency Phenotype. Stem Cell Res. Ther. 2023, 14, 154. [Google Scholar] [CrossRef] [PubMed]
- Manstein, F.; Ullmann, K.; Kropp, C.; Halloin, C.; Triebert, W.; Franke, A.; Farr, C.-M.; Sahabian, A.; Haase, A.; Breitkreuz, Y.; et al. High Density Bioprocessing of Human Pluripotent Stem Cells by Metabolic Control and in Silico Modeling. Stem Cells Transl. Med. 2021, 10, 1063–1080. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Razvi, A.; Anderson-Jenkins, Z.; Sirskyj, D.; Gong, M.; Lavoie, A.-M.; Pigeau, G. Process Development and Scale-up of Pluripotent Stem Cell Manufacturing. Cell Gene Ther. Insights 2020, 6, 1277–1298. [Google Scholar] [CrossRef]
- Pandey, P.R.; Tomney, A.; Woon, M.T.; Uth, N.; Shafighi, F.; Ngabo, I.; Vallabhaneni, H.; Levinson, Y.; Abraham, E.; Friedrich Ben-Nun, I. End-to-End Platform for Human Pluripotent Stem Cell Manufacturing. Int. J. Mol. Sci. 2020, 21, 89. [Google Scholar] [CrossRef]
- Müller, D.; Klein, L.; Lemke, J.; Schulze, M.; Kruse, T.; Saballus, M.; Matuszczyk, J.; Kampmann, M.; Zijlstra, G. Process Intensification in the Biopharma Industry: Improving Efficiency of Protein Manufacturing Processes from Development to Production Scale Using Synergistic Approaches. Chem. Eng. Process. Process Intensif. 2022, 171, 108727. [Google Scholar] [CrossRef]
- Seth, G.; Hamilton, R.W.; Stapp, T.R.; Zheng, L.; Meier, A.; Petty, K.; Leung, S.; Chary, S. Development of a New Bioprocess Scheme Using Frozen Seed Train Intermediates to Initiate CHO Cell Culture Manufacturing Campaigns. Biotechnol. Bioeng. 2013, 110, 1376–1385. [Google Scholar] [CrossRef]
- Clincke, M.-F.; Mölleryd, C.; Samani, P.K.; Lindskog, E.; Fäldt, E.; Walsh, K.; Chotteau, V. Very High Density of Chinese Hamster Ovary Cells in Perfusion by Alternating Tangential Flow or Tangential Flow Filtration in WAVE BioreactorTM—Part II: Applications for Antibody Production and Cryopreservation. Biotechnol. Prog. 2013, 29, 768–777. [Google Scholar] [CrossRef]
- Meiser, I.; Alstrup, M.; Khalesi, E.; Stephan, B.; Speicher, A.M.; Majer, J.; Kwok, C.K.; Neubauer, J.C.; Hansson, M.; Zimmermann, H. Application-Oriented Bulk Cryopreservation of Human iPSCs in Cryo Bags Followed by Direct Inoculation in Scalable Suspension Bioreactors for Expansion and Neural Differentiation. Cells 2023, 12, 1914. [Google Scholar] [CrossRef]
- Heidemann, R.; Mered, M.; Wang, D.Q.; Gardner, B.; Zhang, C.; Michaels, J.; Henzler, H.-J.; Abbas, N.; Konstantinov, K. A New Seed-Train Expansion Method for Recombinant Mammalian Cell Lines. Cytotechnology 2002, 38, 99–108. [Google Scholar] [CrossRef]
- Tao, Y.; Shih, J.; Sinacore, M.; Ryll, T.; Yusuf-Makagiansar, H. Development and Implementation of a Perfusion-Based High Cell Density Cell Banking Process. Biotechnol. Prog. 2011, 27, 824–829. [Google Scholar] [CrossRef]
- Müller, J.; Ott, V.; Eibl, D.; Eibl, R. Seed Train Intensification Using an Ultra-High Cell Density Cell Banking Process. Processes 2022, 10, 911. [Google Scholar] [CrossRef]
- Ninomiya, N.; Shirahata, S.; Murakami, H.; Sugahara, T. Large-Scale, High-Density Freezing of Hybridomas and Its Application to High-Density Culture. Biotechnol. Bioeng. 1991, 38, 1110–1113. [Google Scholar] [CrossRef]
- Lawson, T.; Kehoe, D.E.; Schnitzler, A.C.; Rapiejko, P.J.; Der, K.A.; Philbrick, K.; Punreddy, S.; Rigby, S.; Smith, R.; Feng, Q.; et al. Process Development for Expansion of Human Mesenchymal Stromal Cells in a 50L Single-Use Stirred Tank Bioreactor. Biochem. Eng. J. 2017, 120, 49–62. [Google Scholar] [CrossRef]
- Schirmaier, C.; Jossen, V.; Kaiser, S.C.; Jüngerkes, F.; Brill, S.; Safavi-Nab, A.; Siehoff, A.; van den Bos, C.; Eibl, D.; Eibl, R. Scale-up of Adipose Tissue-Derived Mesenchymal Stem Cell Production in Stirred Single-Use Bioreactors under Low-Serum Conditions. Eng. Life Sci. 2014, 14, 292–303. [Google Scholar] [CrossRef]
- Watanabe, K.; Ueno, M.; Kamiya, D.; Nishiyama, A.; Matsumura, M.; Wataya, T.; Takahashi, J.B.; Nishikawa, S.; Nishikawa, S.; Muguruma, K.; et al. A ROCK Inhibitor Permits Survival of Dissociated Human Embryonic Stem Cells. Nat. Biotechnol. 2007, 25, 681–686. [Google Scholar] [CrossRef]
- Jossen, V.; Eibl, R.; Kraume, M.; Eibl, D. Growth Behavior of Human Adipose Tissue-Derived Stromal/Stem Cells at Small Scale: Numerical and Experimental Investigations. Bioengineering 2018, 5, 106. [Google Scholar] [CrossRef]
- Maldonado, M.; Luu, R.J.; Ramos, M.E.P.; Nam, J. ROCK Inhibitor Primes Human Induced Pluripotent Stem Cells to Selectively Differentiate towards Mesendodermal Lineage via Epithelial-Mesenchymal Transition-like Modulation. Stem Cell Res. 2016, 17, 222–227. [Google Scholar] [CrossRef]
- Couto, P.S.; Stibbs, D.J.; Rotondi, M.C.; Takeuchi, Y.; Rafiq, Q.A. Scalable Manufacturing of Gene-Modified Human Mesenchymal Stromal Cells with Microcarriers in Spinner Flasks. Appl. Microbiol. Biotechnol. 2023, 107, 5669–5685. [Google Scholar] [CrossRef] [PubMed]
- Manstein, F.; Ullmann, K.; Triebert, W.; Zweigerdt, R. Process Control and in Silico Modeling Strategies for Enabling High Density Culture of Human Pluripotent Stem Cells in Stirred Tank Bioreactors. STAR Protoc. 2021, 2, 100988. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, S.S. Engineering Challenges in High Density Cell Culture Systems. Cytotechnology 1996, 22, 3–16. [Google Scholar] [CrossRef]
- Bausch, M.; Schultheiss, C.; Sieck, J.B. Recommendations for Comparison of Productivity Between Fed-Batch and Perfusion Processes. Biotechnol. J. 2019, 14, 1700721. [Google Scholar] [CrossRef] [PubMed]
- Crook, J.M.; Kravets, L. Cell Preservation Method for Pluripotent Stem Cells. US9714412B2, 25 July 2017. [Google Scholar]
- Sullivan, S.; Stacey, G.N.; Akazawa, C.; Aoyama, N.; Baptista, R.; Bedford, P.; Bennaceur Griscelli, A.; Chandra, A.; Elwood, N.; Girard, M.; et al. Quality Control Guidelines for Clinical-Grade Human Induced Pluripotent Stem Cell Lines. Regen. Med. 2018, 13, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Jung, S.; Hashimura, Y.; Lee, M.; Borys, B.S.; Dang, T.; Kallos, M.S.; Rodrigues, C.A.V.; Silva, T.P.; Cabral, J.M.S. Cell Culture Process Scale-Up Challenges for Commercial-Scale Manufacturing of Allogeneic Pluripotent Stem Cell Products. Bioengineering 2022, 9, 92. [Google Scholar] [CrossRef]
- Badenes, S.M.; Fernandes, T.G.; Rodrigues, C.A.V.; Diogo, M.M.; Cabral, J.M.S. Microcarrier-Based Platforms for in Vitro Expansion and Differentiation of Human Pluripotent Stem Cells in Bioreactor Culture Systems. J. Biotechnol. 2016, 234, 71–82. [Google Scholar] [CrossRef]
- Dang, T.; Borys, B.S.; Kanwar, S.; Colter, J.; Worden, H.; Blatchford, A.; Croughan, M.S.; Hossan, T.; Rancourt, D.E.; Lee, B.; et al. Computational Fluid Dynamic Characterization of Vertical-Wheel Bioreactors Used for Effective Scale-up of Human Induced Pluripotent Stem Cell Aggregate Culture. Can. J. Chem. Eng. 2021, 99, 2536–2553. [Google Scholar] [CrossRef]
- Badenes, S.M.; Fernandes, T.G.; Cordeiro, C.S.M.; Boucher, S.; Kuninger, D.; Vemuri, M.C.; Diogo, M.M.; Cabral, J.M.S. Defined Essential 8TM Medium and Vitronectin Efficiently Support Scalable Xeno-Free Expansion of Human Induced Pluripotent Stem Cells in Stirred Microcarrier Culture Systems. PLoS ONE 2016, 11, e0151264. [Google Scholar] [CrossRef]
- Nogueira, D.E.S.; Rodrigues, C.A.V.; Carvalho, M.S.; Miranda, C.C.; Hashimura, Y.; Jung, S.; Lee, B.; Cabral, J.M.S. Strategies for the Expansion of Human Induced Pluripotent Stem Cells as Aggregates in Single-Use Vertical-WheelTM Bioreactors. J. Biol. Eng. 2019, 13, 74. [Google Scholar] [CrossRef]
- Marescal, O.; Cheeseman, I.M. Cellular Mechanisms and Regulation of Quiescence. Dev. Cell 2020, 55, 259–271. [Google Scholar] [CrossRef]
- Petry, F.; Salzig, D. Impact of Bioreactor Geometry on Mesenchymal Stem Cell Production in Stirred-Tank Bioreactors. Chem. Ing. Tech. 2021, 93, 1537–1554. [Google Scholar] [CrossRef]
- Ornelas-González, A.; González-González, M.; Rito-Palomares, M. Microcarrier-Based Stem Cell Bioprocessing: GMP-Grade Culture Challenges and Future Trends for Regenerative Medicine. Crit. Rev. Biotechnol. 2021, 41, 1081–1095. [Google Scholar] [CrossRef] [PubMed]
- Petry, F.; Salzig, D. Large-Scale Production of Size-Adjusted β-Cell Spheroids in a Fully Controlled Stirred-Tank Reactor. Processes 2022, 10, 861. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | T25 Control | BioBLU Control | BioBLU OSI | |
|---|---|---|---|---|
| * | (%) | 75.9 ± 6.2 a | 69.5 ± 19.0 a | 121.4 ± 18.6 b |
| * | (%) | - **** | 57.8 ± 31.3 a | 67.0 ± 14.4 a |
| ** | (h) | 16.4 ± 0.3 a | 15.0 ± 0.8 b | 13.9 ± 0.3 b |
| Max. VCDA *** | (105 cells cm−2) | 3.4 ± 0.3 a | 3.4 ± 1.6 a | 4.1 ± 1.3 a |
| Viability *** | (%) | 94.6 ± 1.5 a | 92.9 ± 0.6 a | 95.4 ± 1.4 a |
| (-) | 17.1 ± 1.5 a | 18.1 ± 8.6 a | 22.0 ± 7.1 a | |
| (pmol cell−1 d−1) | 14.1 ± 4.6 a | 14.7 ± 7.4 a | 10.1 ± 5.7 a | |
| (pmol cell−1 d−1) | 26.4 ± 7.0 a | 24.0 ± 4.5 a | 17.1 ± 2.2 a | |
| (pmol cell−1 d−1) | 2.3 ± 0.6 a | 2.0 ± 1.2 a | 1.4 ± 0.9 a | |
| (pmol cell−1 d−1) | 1.8 ± 0.6 a | 1.3 ± 0.8 a | 1.0 ± 0.6 a | |
| (mol mol−1) | 1.9 ± 0.5 a | 1.6 ± 0.0 a | 1.7 ± 0.1 a | |
| (mol mol−1) | 0.8 ± 0.2 a | 0.7 ± 0.1 a | 0.8 ± 0.1 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teale, M.A.; Schneider, S.L.; Eibl, D.; Eibl, R. Process Intensification in Human Pluripotent Stem Cell Expansion with Microcarriers. Processes 2024, 12, 426. https://doi.org/10.3390/pr12030426
Teale MA, Schneider SL, Eibl D, Eibl R. Process Intensification in Human Pluripotent Stem Cell Expansion with Microcarriers. Processes. 2024; 12(3):426. https://doi.org/10.3390/pr12030426
Chicago/Turabian StyleTeale, Misha Alexander, Samuel Lukas Schneider, Dieter Eibl, and Regine Eibl. 2024. "Process Intensification in Human Pluripotent Stem Cell Expansion with Microcarriers" Processes 12, no. 3: 426. https://doi.org/10.3390/pr12030426