

Signal Molecules Regulate the Synthesis of Secondary Metabolites in the Interaction between Endophytes and Medicinal Plants

 and
and
Abstract
:1. Introduction
2. Signaling Molecules Involved in the Interactions between Endophytic Bacteria and Medicinal Plants
2.1. Interaction of Metabolic Signaling Molecules in Endophytic Bacteria and Medicinal Plants
2.2. Receptor Signaling Molecules in the Interactions between Endophytic Bacteria and Medicinal Plants
3. Endophytes Mediate the Transduction and Integration of Major Signaling Molecules for the Accumulation of Secondary Metabolites in Medicinal Plants
3.1. Intracellular Signal Transduction System of Endophytic Bacteria Mediating the Accumulation of Secondary Metabolites in Medicinal Plants
3.2. Signal Integration of Endophytic Bacteria Mediating the Accumulation of Secondary Metabolites in Medicinal Plants
4. Molecular Mechanisms of Signaling by Endophytic Bacteria Mediating the Accumulation of Secondary Metabolites in Medicinal Plants
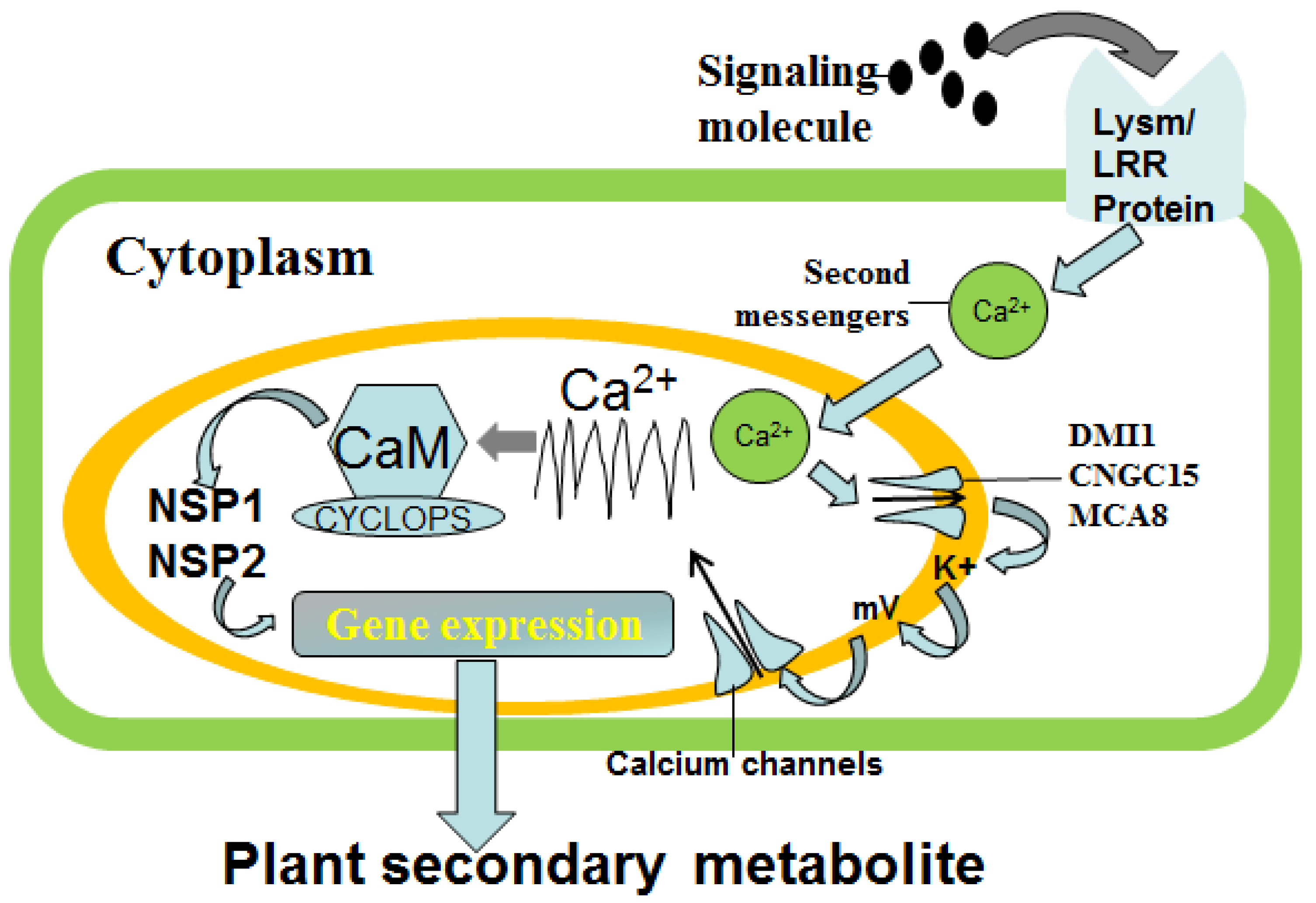
4.1. Calcium Signal Transduction Mechanism
4.2. ROS Signal Transduction Mechanism
4.3. Mechanisms of Jasmonic Acid (JA) Signaling
4.4. Salicylic Acid (SA) Signal Transduction Mechanism
4.5. Multi-Molecular Tandem Interaction Mechanism
5. Conclusions
6. Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, L.; Li, Z.-K.; Li, C.-Y.; Liang, Y.-Q.; Yang, F. Anticancer properties and pharmaceutical applications of ginsenoside compound K: A review. Chem. Biol. Drug Des. 2022, 99, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wu, H.; Yan, B.; Zhao, H.; Liu, F.; Zhang, H.; Sheng, Q.; Miao, F.; Liang, Z. Core Microbiome of Medicinal Plant Salvia miltiorrhiza Seed: A Rich Reservoir of Beneficial Microbes for Secondary Metabolism? Int. J. Mol. Sci. 2018, 19, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neha, D.; Shikha, S. Cancer chemotherapy with novel bioactive natural products. J. Chin. Pharm. Sci. 2022, 31, 589–607. [Google Scholar] [CrossRef]
- Ran, X.; Zhang, G.; Li, S.; Wang, J. Characterization and antitumor activity of camptothecin from endophytic fungus Fusarium solani isolated from Camptotheca acuminate. Afr. Health Sci. 2017, 17, 566–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumaran, R.S.; Kim, H.J.; Hur, B.K. Taxol-producing [corrected] fungal endophyte, Pestalotiopsis species isolated from Taxus cuspidata. J. Biosci. Bioeng. 2010, 110, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.-G.; Mo, Q.-Q.; Zhao, Y.; Zang, P. Microbial mediated accumulation of plant secondary metabolites and its action mechanism in medicinal plants: A review. Guangxi Agric Sci. 2019, 50, 2234–2240. [Google Scholar]
- Ding, C.; Wang, S.; Li, J.; Wang, Z. Transcriptomic analysis reveals the mechanism of host growth promotion by endophytic fungus of Rumex gmelinii Turcz. Arch. Microbiol. 2022, 204, 443. [Google Scholar] [CrossRef]
- Yipare, P.; Zulihumaer, R.; Tian, Y.-Z.; Zhu, Y.-L.; Li, Y.-T.; Ma, X.-L. Research Progress in Diversity of Endophytic Microbial Communities Isolated from Desert Plants and Their Strengthening Effect on Drought and Salt Tolerance in Crops. Biotechnol. Bull. 2022, 38, 88. [Google Scholar]
- Li, Z.; Wen, W.; Qin, M.; He, Y.; Xu, D.; Li, L. Biosynthetic Mechanisms of Secondary Metabolites Promoted by the Interaction Between Endophytes and Plant Hosts. Front. Microbiol. 2022, 13, 928967. [Google Scholar] [CrossRef]
- Mengistu, A.A. Endophytes: Colonization, Behaviour, and Their Role in Defense Mechanism. Int. J. Microbiol. 2020, 2020, 6927219. [Google Scholar] [CrossRef]
- Lu, X.; Wang, Y.-Y.; Zhang, F.-Y.; Lin, X.-Y.; Tang, K.-X. Advances in studies on transcriptional regulatory factor in secondary metabolites regulation of Chinese materia medica. Chin. Tradit. Herb. Drugs 2010, 41, 159–162. [Google Scholar]
- Dubey, A.; Malla, M.A.; Kumar, A.; Dayanandan, S.; Khan, M.L. Plants endophytes: Unveiling hidden agenda for bioprospecting toward sustainable agriculture. Crit. Rev. Biotechnol. 2020, 40, 1210–1231. [Google Scholar] [CrossRef]
- You, C.; Qin, D.; Wang, Y.; Lan, W.; Li, Y.; Yu, B.; Peng, Y.; Xu, J.; Dong, J. Plant Triterpenoids Regulate Endophyte Community to Promote Medicinal Plant Schisandra sphenanthera Growth and Metabolites Accumulation. J. Fungi 2021, 7, 788. [Google Scholar] [CrossRef] [PubMed]
- Harbort, C.J.; Hashimoto, M.; Inoue, H.; Niu, Y.; Guan, R.; Rombolà, A.D.; Kopriva, S.; Voges, M.J.E.E.E.; Sattely, E.S.; Garrido-Oter, R.; et al. Root-Secreted Coumarins and the Microbiota Interact to Improve Iron Nutrition in Arabidopsis. Cell Host Microbe. 2020, 28, 825–837. [Google Scholar] [CrossRef]
- Hauth, F.; Buck, H.; Stanoppi, M.; Hartig, J.S. Canavanine utilization via homoserine and hydroxyguanidine by a PLP-dependent γ-lyase in Pseudomonadaceae and Rhizobiales. RSC Chem. Biol. 2022, 3, 1240–1250. [Google Scholar] [CrossRef]
- Ghantasala, S.; Roy Choudhury, S. Nod factor perception: An integrative view of molecular communication during legume symbiosis. Plant Mol. Biol. 2022, 110, 485–509. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Hayashi, H. Strigolactones: Chemical signals for fungal symbionts and parasitic weeds in plant roots. Ann. Bot. 2006, 97, 925–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besserer, A.; Puech-Pagès, V.; Kiefer, P.; Gomez-Roldan, V.; Jauneau, A.; Roy, S.; Portais, J.C.; Roux, C.; Bécard, G.; Séjalon-Delmas, N. Strigolactones stimulate arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biol. 2006, 4, e226. [Google Scholar] [CrossRef]
- Liang, S.-M.; Zheng, F.-L.; Wu, Q.-S. Elucidating the dialogue between arbuscular mycorrhizal fungi and polyamines in plants. World J. Microbiol. Biotechnol. 2022, 38, 159. [Google Scholar] [CrossRef]
- Qu, Z.; Zhang, H.; Wang, Q.; Zhao, H.; Liu, X.; Fu, Y.; Lin, Y.; Xie, J.; Cheng, J.; Li, B.; et al. Exploring the Symbiotic Mechanism of a Virus-Mediated Endophytic Fungus in Its Host by Dual Unique Molecular Identifier-RNA Sequencing. mSystems 2021, 6, e0081421. [Google Scholar] [CrossRef]
- Snoeck, S.; Abramson, B.W.; Garcia, A.G.K.; Egan, A.N.; Michael, T.P.; Steinbrenner, A.D. Evolutionary gain and loss of a plant pattern-recognition receptor for HAMP recognition. Elife 2022, 11, e81050. [Google Scholar] [CrossRef] [PubMed]
- Kawaharada, Y.; Kelly, S.; Nielsen, M.W.; Hjuler, C.T.; Gysel, K.; Muszyński, A.; Carlson, R.W.; Thygesen, M.B.; Sandal, N.; Asmussen, M.H.; et al. Receptor-mediated exopolysaccharide perception controls bacterial infection. Nature 2015, 523, 308–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourassa, D.V.; Kannenberg, E.L.; Sherrier, D.J.; Buhr, R.J.; Carlson, R.W. The Lipopolysaccharide Lipid A Long-Chain Fatty Acid Is Important for Rhizobium leguminosarum Growth and Stress Adaptation in Free-Living and Nodule Environments. Mol. Plant Microbe Interact. 2017, 30, 161–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gough, C.; Cullimore, J. Lipo-chitooligosaccharide signaling in endosymbiotic plant-microbe interactions. Mol. Plant Microbe Interact. 2011, 24, 867–878. [Google Scholar] [CrossRef] [Green Version]
- Miyata, K.; Hayafune, M.; Kobae, Y.; Kaku, H.; Nishizawa, Y.; Masuda, Y.; Shibuya, N.; Nakagawa, T. Evaluation of the Role of the LysM Receptor-Like Kinase, OsNFR5/OsRLK2 for AM Symbiosis in Rice. Plant Cell Physiol. 2016, 57, 2283–2290. [Google Scholar] [CrossRef] [Green Version]
- Smit, P.; Limpens, E.; Geurts, R.; Fedorova, E.; Dolgikh, E.; Gough, C.; Bisseling, T. Medicago LYK3, an entry receptor in rhizobial nodulation factor signaling. Plant Physiol. 2007, 145, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.M.; Kobayashi, H.; Davies, B.W.; Taga, M.E.; Walker, G.C. How rhizobial symbionts invade plants: The Sinorhizobium-Medicago model. Nat. Rev. Microbiol. 2007, 5, 619–633. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.-H.; Wang, D.-G.; Pu, X.-L.; Huang, J.-H.; Zhao, X.; Qiu, W.-Q.; Jiang, S. Signal transduction of atractylodin biosynthesis in Atractylodes lancea cell induced by endophytic fungal elicitor mediated with nitric oxide followed by salicylic acid. Chin. Tradit. Herb. Drugs 2014, 45, 701–708. [Google Scholar]
- Rudrappa, T.; Czymmek, K.J.; Paré, P.W.; Bais, H.P. Root-secreted malic acid recruits beneficial soil bacteria. Plant Physiol. 2008, 148, 1547–1556. [Google Scholar] [CrossRef] [Green Version]
- Liu, C. Research progress on biological control mechanism of Pseudomonas fluorescens. Sci. Technol. Res. 2014, 23, 683. [Google Scholar]
- Fang, Z.Y. The Effects and Mechanisms of Bacillus amyloliquefaciens SQY162 on Biological Control of Bacterial Wilt of Tomato. Nanjing Agric. Univ. 2016, 4, 82. [Google Scholar]
- Voges, M.J.E.E.E.; Bai, Y.; Schulze-Lefert, P.; Sattely, E.S. Plant-derived coumarins shape the composition of an Arabidopsis synthetic root microbiome. Proc. Natl. Acad. Sci. USA 2019, 116, 12558–12565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, T.; Cai, W.; Zhang, J.; Zheng, H.; Tsou, A.M.; Xiao, L.; Zhong, Z.; Zhu, J. Host legume-exuded antimetabolites optimize the symbiotic rhizosphere. Mol. Microbiol. 2009, 73, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.-C.; Jiang, T.; Liu, Y.-X.; Bai, Y.-C.; Reed, J.; Qu, B.; Goossens, A.; Nützmann, H.-W.; Bai, Y.; Osbourn, A. A specialized metabolic network selectively modulates Arabidopsis root microbiota. Science 2019, 364, eaau6389. [Google Scholar] [CrossRef]
- Lebeis, S.L.; Paredes, S.H.; Lundberg, D.S.; Breakfield, N.; Gehring, J.; McDonald, M.; Malfatti, S.; Del Rio, T.G.; Jones, C.D.; Tringe, S.G.; et al. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-Q.; Yang, W.-Y.; Liu, J. Advances on Chemical Ecology of Plant Flavonoids. Nat. Prod. Res. Dev. 2018, 30, 2009–2016. [Google Scholar]
- Yoneyama, K.; Xie, X.; Nomura, T.; Yoneyama, K.; Bennett, T. Supra-organismal regulation of strigolactone exudation and plant development in response to rhizospheric cues in rice. Curr. Biol. 2022, 32, 3601–3608.e3. [Google Scholar] [CrossRef]
- Liu, Z.; Beskrovnaya, P.; Melnyk, R.A.; Hossain, S.S.; Khorasani, S.; O’Sullivan, L.R.; Wiesmann, C.L.; Bush, J.; Richard, J.D.; Haney, C.H. A Genome-Wide Screen Identifies Genes in Rhizosphere-Associated Pseudomonas Required to Evade Plant Defenses. mBio 2018, 9, e00433-18. [Google Scholar] [CrossRef] [Green Version]
- Sasse, J.; Martinoia, E.; Northen, T. Feed Your Friends: Do Plant Exudates Shape the Root Microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Kong, J.; Cui, D.; Zhao, H.; Niu, Y.; Xu, M.; Jiang, G.; Zhao, Y.; Wang, W. Resistance against Ralstonia solanacearum in tomato depends on the methionine cycle and the γ-aminobutyric acid metabolic pathway. Plant J. 2019, 97, 1032–1047. [Google Scholar] [CrossRef]
- Becker, A.; Fraysse, N.; Sharypova, L. Recent advances in studies on structure and symbiosis-related function of rhizobial K-antigens and lipopolysaccharides. Mol. Plant Microbe Interact. 2005, 18, 899–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D.-H.; Kong, S.H.; Guo, Z.-J. Cloning of cryptogein gene and construction of its plant expression vector. J. Zhejiang Univ. 2002, 6, 62–67. [Google Scholar]
- Gui, X.; Zhang, P.; Wang, D.; Ding, Z.; Wu, X.; Shi, J.; Shen, Q.-H.; Xu, Y.Z.; Ma, W.; Qiao, Y. Phytophthora effector PSR1 hijacks the host pre-mRNA splicing machinery to modulate small RNA biogenesis and plant immunity. Plant Cell 2022, 34, 3443–3459. [Google Scholar] [CrossRef]
- Rush, T.A.; Puech-Pages, V.; Bascaules, A.; Jargeat, P.; Maillet, F.; Haouy, A.; Maes, A.Q.; Carriel, C.C.; Khokhani, D.; Keller-Pearson, M.; et al. Lipo-Chitooligosaccharides: From Plant Symbiosis to Conserved Fungal Signals. 2020. Available online: https://www.researchgate.net/publication/342819957_Lipo-chitooligosaccharides_from_plant_symbiosis_to_conserved_fungal_signals (accessed on 10 March 2023).
- Cope, K.R.; Bascaules, A.; Irving, T.B.; Venkateshwaran, M.; Maeda, J.; Garcia, K.; Rush, T.A.; Ma, C.; Labbé, J.; Jawdy, S.; et al. The Ectomycorrhizal Fungus Laccaria bicolor Produces Lipochitooligosaccharides and Uses the Common Symbiosis Pathway to Colonize Populus Roots. Plant Cell 2019, 31, 2386–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, X.-F.; Song, Z.-F.; Ming, M.-G.; Li, Y.-J. Regulation Molecular Mechnisms or Ca2+ Singaling on Plant-Environmental Microorgainsm Interactions. Acta Bot. Boreali-Occident. Sin. 2016, 36, 2128–2136. [Google Scholar]
- Modolo, L.V.; Cunha, F.Q.; Braga, M.R.; Salgado, I. Nitric oxide synthase-mediated phytoalexin accumulation in soybean cotyledons in response to the Diaporthe phaseolorumf. sp. meridionalis elicitor. Plant Physiol. 2002, 130, 1288–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.-L.; Zhang, C.-L.; Lin, F.-C. Recent advances on physiological and molecular basis of fungal endophyteplant interactions. Acta Ecol. Sin. 2008, 28, 4430–4439. [Google Scholar]
- Lan, W.-Z.; Yu, L.-J.; Li, W.; Cai, H.-X. Selecting Preparation Methods and Isoating Components of Fungal Elicitor to Enhance Taxol Biosyn thesis. Wuhan Bot. Res. 2002, 1, 66–70. [Google Scholar]
- Xu, M.-J.; Long, J.F.; Zhu, M.-Y. NO promotes the biosynthesis of puerarin in Pueraria lobata suspension cells through salicylic acid or jasmonic acid signaling pathway mediated by fungal elicitors. Sci. Sin. 2006, 36, 10. [Google Scholar]
- Kolbert, Z.; Lindermayr, C.; Loake, G.J. The role of nitric oxide in plant biology: Current insights and future perspectives. J. Exp. Bot. 2021, 72, 777–780. [Google Scholar] [CrossRef]
- Meng, X.; Xu, J.; He, Y.; Yang, K.Y.; Mordorski, B.; Liu, Y.; Zhang, S. Phosphorylation of an ERF transcription factor by Arabidopsis MPK3/MPK6 regulates plant defense gene induction and fungal resistance. Plant Cell 2013, 25, 1126–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Li, Y.; Xie, L.; Hao, X.; Liu, H.; Qin, W.; Wang, C.; Yan, X.; Wu, Z.-K.; Yao, X.; et al. AaWRKY17, a positive regulator of artemisinin biosynthesis, is involved in resistance to Pseudomonas syringae in Artemisia annua. Hortic. Res. 2021, 8, 217. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.-X.; Li, J.-X.; Yang, C.-Q.; Hu, W.-L.; Wang, L.-J.; Chen, X.-Y. The jasmonate-responsive AP2/ERF transcription factors AaERF1 and AaERF2 positively regulate artemisinin biosynthesis in Artemisia annua L. Mol. Plant. 2012, 5, 353–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Zhang, L.; Zhang, F.; Jiang, W.; Shen, Q.; Zhang, L.; Lv, Z.; Wang, G.; Tang, K. AaORA, a trichome-specific AP2/ERF transcription factor of Artemisia annua, is a positive regulator in the artemisinin biosynthetic pathway and in disease resistance to Botrytis cinerea. New Phytol. 2013, 198, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- LI, S.-T. Cloning and Functional Study of Transcription Factors TcMYC and TcWRKY1 Regulating Taxol Synthesis. Huazhong Univ. Sci. Technol. 2012, 7, 176. [Google Scholar]
- Dai, Y.; Qin, Q.; Dai, D.; Kong, L.; Li, W.; Zha, X.; Jin, Y.; Tang, K. Isolation and characterization of a novel cDNA encoding methyl jasmonate-responsive transcription factor TcAP2 from Taxus cuspidata. Biotechnol. Lett. 2009, 31, 1801–1809. [Google Scholar] [CrossRef]
- Wei, L.; Ma, J.; Wang, S.; Wu, Y. Isolation and Functional Characterization of a Novel Gene Encoding a Dehydration Responsive Element Binding Transcription Factor from Populus euphratica. Protein Pept. Lett. 2016, 23, 459–467. [Google Scholar] [CrossRef]
- Nims, E.; Vongpaseuth, K.; Roberts, S.C.; Walker, E.L. TcJAMYC: A bHLH transcription factor that activates paclitaxel biosynthetic pathway genes in yew. J. Biol. Chem. 2015, 290, 20104. [Google Scholar] [CrossRef] [Green Version]
- Schwinn, K.; Venail, J.; Shang, Y.; Mackay, S.; Alm, V.; Butelli, E.; Oyama, R.; Bailey, P.; Davies, K.; Martin, C. A small family of MYB-regulatory genes controls floral pigmentation intensity and patterning in the genus Antirrhinum. Plant Cell 2006, 18, 831–851. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.-D. The Cause and Research Status of Flower Color Mutation of Ornamental Plants. Mod. Hortic. 2012, 16, 7–8. [Google Scholar]
- Nakatsuka, T.; Haruta, K.S.; Pitaksutheepong, C.; Abe, Y.; Kakizaki, Y.; Yamamoto, K.; Shimada, N.; Yamamura, S.; Nishihara, M. Identification and characterization of R2R3-MYB and bHLH transcription factors regulating anthocyanin biosynthesis in gentian flowers. Plant Cell Physiol. 2008, 49, 1818–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatsuka, T.; Saito, M.; Yamada, E.; Fujita, K.; Kakizaki, Y.; Nishihara, M. Isolation and characterization of GtMYBP3 and GtMYBP4, orthologues of R2R3-MYB transcription factors that regulate early flavonoid biosynthesis, in gentian flowers. J. Exp. Bot. 2012, 63, 6505–6517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MEHG, H. Cloning and transcriptional of MYB transcription factor genes from Fagopyrun tataricun. Scihuan Agric. Univ. 2011, 5, 55. [Google Scholar]
- Wu, C. Preliminary Study on the Function of Scutellaria baicalensis MYB Transcription Factor. Wuhan Univ. Technol. 2012, 6, 85. [Google Scholar]
- Chatel, G.; Montiel, G.; Pré, M.; Memelink, J.; Thiersault, M.; Saint-Pierre, B.; Doireau, P.; Gantet, P. CrMYC1, a Catharanthus roseus elicitor- and jasmonate-responsive bHLH transcription factor that binds the G-box element of the strictosidine synthase gene promoter. J. Exp. Bot. 2003, 54, 2587–2588. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Hedhili, S.; Montiel, G.; Zhang, Y.; Chatel, G.; Pré, M.; Gantet, P.; Memelink, J. The basic helix-loop-helix transcription factor CrMYC2 controls the jasmonate-responsive expression of the ORCA genes that regulate alkaloid biosynthesis in Catharanthus roseus. Plant J. 2011, 67, 61–71. [Google Scholar] [CrossRef]
- Menke, L.h.F. A novel jasmonate- and elicitor-responsive element in the periwinkle secondary metabolite biosynthetic gene Str interacts with a jasmonate- and elicitor-inducible AP2-domain transcription factor, ORCA2. EMBO J. 2014, 18, 4455–4463. [Google Scholar] [CrossRef] [Green Version]
- Memelink, J.; Gantet, P. Transcription factors involved in terpenoid indole alkaloid biosynthesis in Catharanthus roseus. Phytochemistry 2007, 6, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Sibéril, Y.; Benhamron, S.; Memelink, J.; Giglioli-Guivarc’h, N.; Thiersault, M.; Boisson, B.; Doireau, P.; Gantet, P. Catharanthus roseus G-box binding factors 1 and 2 act as repressors of strictosidine synthase gene expression in cell cultures. Plant Mol. Biol. 2001, 45, 477–488. [Google Scholar] [CrossRef]
- Suttipanta, N.; Pattanaik, S.; Kulshrestha, M.; Patra, B.; Singh, S.K.; Yuan, L. The transcription factor CrWRKY1 positively regulates the terpenoid indole alkaloid biosynthesis in Catharanthus roseus. Plant Physiol. 2011, 157, 2081–2093. [Google Scholar] [CrossRef] [Green Version]
- Pauw, B.; Hilliou, A.; Martin, V.S.; Chatel, G.; de Wolf, C.J.; Champion, A.; Pre, M.; van Duijn, B.; Kijne, J.W.; van der Fits, L.; et al. Zinc finger proteins act as tanscriptional repressors of alkaloidbiosynthesis genes in Catharanthus roseus. J. BiolChem. 2004, 279, 52940–52948. [Google Scholar]
- Yamada, Y.; Kokabu, Y.; Chaki, K.; Yoshimoto, T.; Ohgaki, M.; Yoshida, S.; Kato, N.; Koyama, T.; Sato, F. Isoquinoline alkaloid biosynthesis is regulated by a unique bHLH-type transcription factor in Coptis japonica. Plant Cell Physiol. 2011, 52, 1131–1141. [Google Scholar] [CrossRef] [Green Version]
- Kato, N.; Dubouzet, E.; Kokabu, Y.; Yoshida, S.; Taniguchi, Y.; Dubouzet, J.G.; Yazaki, K.; Sato, F. Identification of a WRKY protein as a transcriptional regulator of benzylisoquinoline alkaloid biosynthesis in Coptis japonica. Plant Cell Physiol. 2007, 48, 8–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-R.; Wang, J.; Wang, S.-Y. Construction of Artificial microRNA Expression Vector Directing Specific Silencing of the Transcription Factor Gene SmMYC and Subsequent Transfer-mation into Salvia miltiorrhiza. Plant Physiol. J. 2013, 49, 1339–1346. [Google Scholar]
- Tang, N.; Cao, Z.; Yang, C.; Ran, D.; Wu, P.; Gao, H.; He, N.; Liu, G.; Chen, Z. A R2R3-MYB transcriptional activator LmMYB15 regulates chlorogenic acid biosynthesis and phenylpropanoid metabolism in Lonicera macranthoides. Plant Sci. 2021, 308, 110924. [Google Scholar] [CrossRef]
- Zhou, Y.; Xun, Q.; Zhang, D.; Lv, M.; Ou, Y.; Li, J. TCP Transcription Factors Associate with Phytochrome Interacting Factor 4 and Cryptochrome 1 to Regulate Thermomorphogenesis in Arabidopsis thaliana. iScience 2019, 15, 600–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.; Wang, S.-M.; Wu, W.-Y.; Han, L. Inhibiting effect of Polyphyllin I on the cell growth of lung cancer by regulating Sp1/miR-542-3p/ILK signaling pathway. China J. Tradit. Chin. Med. Pharm. 2022, 37, 2805–2812. [Google Scholar]
- He, N.; Yang, Y.; Zhu, J.; Zhang, J.-M.; Chen, Q.-H.; Yang, G.-Y. Research progress in the mechanism of anti-lung cancer effect of polyphyllin. Cent. Pharm. 2022, 20, 2368–2373. [Google Scholar]
- Xu, M.-L. Optimizationof the Culturecondrtions of Transgenic Bax Hiypericum perforatun L. Suspension Cells Andinvestigationon the Signaltransduction Mechanism Ofbax-Induced Secondary Metabolitebiosynthesis. Zhejiang Gongshang Univ. 2006, 3, 72. [Google Scholar]
- Wan, S.-Z. Molecular Mechanism of Poplar PtrMYB57 Negative Regulation on Anthocyanin and Tannin Synthesis. Master’s Thesis, University of Chinese Academy of Sciences, Beijing, China, 2017. [Google Scholar]
- Jin, Z.; Jiang, W.; Luo, Y.; Huang, H.; Yi, D.; Pang, Y. Analyses on Flavonoids and Transcriptome Reveals Key MYB Gene for Proanthocyanidins Regulation in Onobrychis Viciifolia. Front. Plant Sci. 2022, 13, 941918. [Google Scholar] [CrossRef] [PubMed]
- Szymanski, D.B.; Klis, D.A.; Larkin, J.C.; Marks, M.D. cot1: A regulator of Arabidopsis trichome initiation. Genetics 1998, 149, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.-L.; Fu, S.-B.; Gao, F.; Wang, M.L. Signal mechanism of secondary metabolites accumulation in plants induced by fungus and its application in medicinal plants. Chin. Tradit. Herb. Drugs 2012, 43, 1647–1651. [Google Scholar]
- Dong, F.; Yang, F.; Liu, Y.; Jia, W.; He, X.; Chai, J.; Zhao, H.; Lv, M.; Zhao, L.; Zhou, S. Calmodulin-binding transcription activator (CAMTA)/factors in plants. In Calcium Transport Elements in Plants; Academic Press: Cambridge, MA, USA, 2021; pp. 249–266. [Google Scholar]
- Peiter, E.; Sun, J.; Heckmann, A.B.; Venkateshwaran, M.; Riely, B.K.; Otegui, M.S.; Edwards, A.; Freshour, G.; Hahn, M.G.; Cook, D.R.; et al. The Medicago truncatula DMI1 protein modulates cytosolic calcium signaling. Plant Physiol. 2007, 145, 192–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldroyd, G.E.; Downie, J.A. Coordinating nodule morphogenesis with rhizobial infection in legumes. Annu. Rev. Plant Biol. 2008, 59, 519–546. [Google Scholar] [CrossRef]
- Nasir, F.; Bahadur, A.; Lin, X.; Gao, Y.; Tian, C. Novel insights into host receptors and receptor-mediated signaling that regulate arbuscular mycorrhizal symbiosis. J. Exp. Bot. 2021, 72, 1546–1557. [Google Scholar] [CrossRef] [PubMed]
- Chabaud, M.; Genre, A.; Sieberer, B.J.; Faccio, A.; Fournier, J.; Novero, M.; Barker, D.G.; Bonfante, P. Arbuscular mycorrhizal hyphopodia and germinated spore exudates trigger Ca2+ spiking in the legume and nonlegume root epidermis. New Phytol. 2011, 189, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.-M. The regulation of the transcription factors NSP1, NSP2and IPN2 on nodule inception in Legume. Huazhong Agric. Univ. 2015, 2, 60. [Google Scholar]
- Qi, W.-L.; Sun, W.-C.; Ma, L. Research progress of reactive oxygen species involved in regulating plant growth and development and the mechanisms of stress response. Agric. Res. Arid Areas 2021, 39, 69–81,193. [Google Scholar]
- Qin, L.; Guo, L.; Zhang, X.-B.; Guo, Y.; Yang, C.; Xia, S.-T. Study Progress of ROS Signal and Its Regulations on Plant Resistance to Stresses. Acta Laser Biol. Sin. 2018, 27, 193–198. [Google Scholar]
- Wu, B.; Li, P.; Hong, X.; Xu, C.; Wang, R.; Liang, Y. The receptor-like cytosolic kinase RIPK activates NADP-malic enzyme 2 to generate NADPH for fueling ROS production. Mol. Plant. 2022, 15, 887–903. [Google Scholar] [CrossRef]
- Tian, W.; Wang, C.; Gao, Q.; Li, L.; Luan, S. Calcium spikes, waves and oscillations in plant development and biotic interactions. Nat Plants. 2020, 6, 750–759. [Google Scholar] [CrossRef]
- Bu, X.-Y.; Yang, W.-L. Research progress on antibacterial mechanism of plant endophytes and antibacterial secondary metabolites. Drugs Clin. 2021, 36, 2200–2206. [Google Scholar]
- Cheng, J.-H.; Tian, Y.-R.; Li, A.-J. Jasmonic acid signal and its research progress in woody plants. Sci. Sin. Vitae 2020, 50, 215–226. [Google Scholar]
- Mao, J.-H.; Xiong, X.-H.; Lu, Y.-C. Advances in jasmonic acid regulation of plant response to stress. Chin. J. Bioprocess Eng. 2021, 19, 413–419+462. [Google Scholar]
- Liang, X.-W.; Yang, Q.; Li, D.; Cheng, X.-X.; Tang, X.M.; Fan, L.M.; Zhang, C.-R. Regulation of Methyl Jasmonate on Secondary Metabolism of Glycyrrhiza uralensis. Guangdong Agric. Sci. 2017, 44, 57–62. [Google Scholar]
- Chang, C. Study on the Function of COI1 Gene, a Key Member of Danshen Jasmine Signaling Pathway. Shaanxi Norm. Univ. 2017, 5, 190. [Google Scholar]
- Chen, X.; Wang, W.; Cai, P.; Wang, Z.; Li, T.; Du, Y. The role of the MAP kinase-kinase protein StMKK1 in potato immunity to different pathogens. Hortic. Res. 2021, 8, 117. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Han, S.; Qi, Y. Advances in structure and function of auxin response factor in plants. Integr. Plant Biol. 2022, 65, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.-H.; Wang, X.-Y.; Liu, P.Y.; Ji, C.-G.; Wang, L.; Liu, X.Q.; Li, X.-R.; Yan, H.-C. Research progress in structure and function of NPR1. Jiangsu Agric. Sci. 2021, 49, 46–52. [Google Scholar]
- Ren, C.-G. Study on the signal regulation mechanism of endophytic bacteria promoting the accumulation of volatile oil in Atractylodes lanceolata. Nanjing Norm. Univ. 2014, 1, 119. [Google Scholar]
- Xu, M.-J.; Dong, J.F.; Zhang, X.-B. Signal interaction between NO and H2O2 in hypericin synthesis induced by thermal stress. Sci. China Ser. C Life Sci. 2008, 7, 643–653. [Google Scholar]
- Xu, M.-J.; Dong, J.-F. NO can promote the biosynthesis of puerarin in Pueraria lobata suspension cells through salicylic acid (SA) or jasmonic acid (JA) signal pathway mediated by fungal elicitors. Sci. Sin. 2006, 36, 66–75. [Google Scholar]
- Ullah, C.; Schmidt, A.; Reichelt, M.; Tsai, C.J.; Gershenzon, J. Lack of antagonism between salicylic acid and jasmonate signalling pathways in poplar. New Phytol. 2022, 235, 701–717. [Google Scholar] [CrossRef] [PubMed]
- Pelisch, F.; Blaustein, M.; Kornblihtt, A.R.; Srebrow, A. Cross-talk between signaling pathways regulates alternative splicing: A novel role for JNK. J. Biol. Chem. 2005, 280, 25461–25469. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.-J.; Dong, J.-F. Nitric oxide mediates the fungal elicitor-induced Taxol biosynthesis of Taxus chinensis suspension cells through the reactive oxygen species-dependent and-independent signal pathways. Chin. Sci. Bull. 2006, 51, 1675–1682. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Action Category | Signal Molecule | Strain (Genus) | Reference |
|---|---|---|---|---|
| Metabolic signal molecules | Nutrients | Citric acid | Rhizoctonia | [28] |
| Malic acid | Bacillus subtilis FB17 | [29] | ||
| Fumaric acid | Pseudomonas fluorescens | [30] | ||
| Succinic acid | Bacillus amylolyticus | [31] | ||
| Antibacterial substances | Coumarin | Pseudomonas | [32] | |
| Concanavaline | Nitrogenous Rhizobium | [33] | ||
| Specific products | Triterpene | Endophytic flora | [34] | |
| Salicylic acid | Endophytic flora | [35] | ||
| Metabolites | Flavonoid | Nitrogenous Rhizobium | [36] | |
| Unicornolactone | Arbuscular mycorrhiza | [37] | ||
| Polyamine | Pseudomonas | [38] | ||
| Amino acid | Nitrogenous Rhizobium | [39] | ||
| Organic acid | Nitrogenous Rhizobium | [39] | ||
| Sugar | Nitrogenous Rhizobium | [39] | ||
| Receptor Signal Molecules | Conservative MAMP | Extracellular polysaccharide | Nitrogenous Rhizobium | [40] |
| Lipopolysaccharide | Nitrogenous Rhizobium | [41] | ||
| Cell wall polysaccharide | Verticillium dahuricum | [42] | ||
| Phospholipid protein | Phytophthora camphora | [43] | ||
| Ribosomal protein | Phytophthora cryptogea | [44] | ||
| Intracellular signal molecule | Nod factor | LCO | Laccaria bicolor | [45] |
| Second Messenger | Ca2+ | Nitrogenous Rhizobium | [46] | |
| NO | Soybean Stalk Rot Pathogen | [47] | ||
| ROS | E.festucae | [48] | ||
| Hormone molecule | JA | Epichloë gansuensis | [49] | |
| SA | Penicillium citri | [50] |
| Regulate the Type of Metabolite | Botany | Category | Name | Medicinal Products | Reference |
|---|---|---|---|---|---|
| Terpenoids | Artemisia annua Linn. | WRKY | AaWRKY17 | Artemisinin | [53] |
| AP2/ERF | AaERF1, AaERF2 | Artemisinin | [54] | ||
| AaORA | Artemisinin | [55] | |||
| Taxus chinensis | WRKY | WRKY1 | Taxol | [56] | |
| AP2/ERF | TcAP2 | Taxol | [57] | ||
| TcDREB | Taxol | [58] | |||
| bHLH | TcJAMYC | Taxol | [59] | ||
| Flavonoids | Antirrhinum majus L. | R2R3MYB | Rosea1, Rosea2, Venosa | Anthocyanin | [60] |
| bHLH | Delila | Anthocyanin | [61] | ||
| Gentiana scabra Bunge | bHLH | GtbHLH1 | Anthocyanin | [62] | |
| Gentiana trichotoma Kusnez. | R2R3MYB | GtMybP3 GtMybP4 | Xanthone alcohol | [63] | |
| Tartary buckwheat | MYB | FtMyb1, FtMyb2, FtMyb3 | Rutin | [64] | |
| Scutellaria baicalensis Georgi | MYB | SbMYB | Baicalin | [65] | |
| Alkaloids | Catharanthus roseus (L.) G.Don | bHLH | CrMYC1 | Vinblastine/vincristine | [66] |
| CrMYC2 | Indole alkaloid | [67] | |||
| AP2/ERF | ORCA1, ORCA2 | Vinblastine/vincristine | [68] | ||
| ORCA3 | Vinblastine | [69] | |||
| bZIP | CrGBF1, CrGBF2 | Indole alkaloid | [70] | ||
| WRKY | CrWRKY1 | Vinblastine/vincristine | [71] | ||
| C2H2 Zinc-Finger | ZCT1, ZCT2, ZCT3 | Vinblastine/vincristine | [72] | ||
| Coptis chinensis | bHLH | CjbHLH1 | Berberine | [73] | |
| WRKY | CjWRKY1 | Isoquinoline alkaloid | [74] | ||
| Phenylpropanoids | Salvia miltiorrhiza Bge. | bHLH | SmMYC | Salb | [75] |
| Lonicera japonica Thunb. | R2R3-MYB | LmMYB15 | Chlorogenic acid | [76] | |
| Steroids | Arabidopsis thaliana | TCP | TCP1 | Brassinolide | [77] |
| Rhizoma paridis | Sp/KLF | Sp1 | Paris polyphylla saponin I | [78] | |
| NF-κB | NF-κB | Paris polyphylla saponin II | [79] | ||
| Quinones | Hypericum Linn | bcl-2 | Bax | Hypericin | [80] |
| Tannins | Populus L. | MYB | PtrMYB57 | Tannin | [81] |
| Onobrychis viciifolia Scop. | MYB | OvMYBPA2 | Condensed tannins | [82] | |
| Other | Arabidopsis thaliana | MYB-bHLH- WD40 | GL1 | Trichome | [83] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, Z.; Xu, M.; Xiao, Z.; Liu, C.; Du, B.; Xu, D.; Li, L. Signal Molecules Regulate the Synthesis of Secondary Metabolites in the Interaction between Endophytes and Medicinal Plants. Processes 2023, 11, 849. https://doi.org/10.3390/pr11030849
Wang Y, Li Z, Xu M, Xiao Z, Liu C, Du B, Xu D, Li L. Signal Molecules Regulate the Synthesis of Secondary Metabolites in the Interaction between Endophytes and Medicinal Plants. Processes. 2023; 11(3):849. https://doi.org/10.3390/pr11030849
Chicago/Turabian StyleWang, Yaxuan, Zhaogao Li, Mengwei Xu, Zhihao Xiao, Chaobo Liu, Bing Du, Delin Xu, and Lin Li. 2023. "Signal Molecules Regulate the Synthesis of Secondary Metabolites in the Interaction between Endophytes and Medicinal Plants" Processes 11, no. 3: 849. https://doi.org/10.3390/pr11030849