


Assessing Biodegradation Processes of Atrazine in Constructed Wetland Using Compound-Specific Stable Isotope Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Configuration of the Bench-Scale Two-Stage Vertical Flow Wetland System
2.2. Bioaugmentation of the Wetland System Using Atrazine-Degrading Mixed Cultures
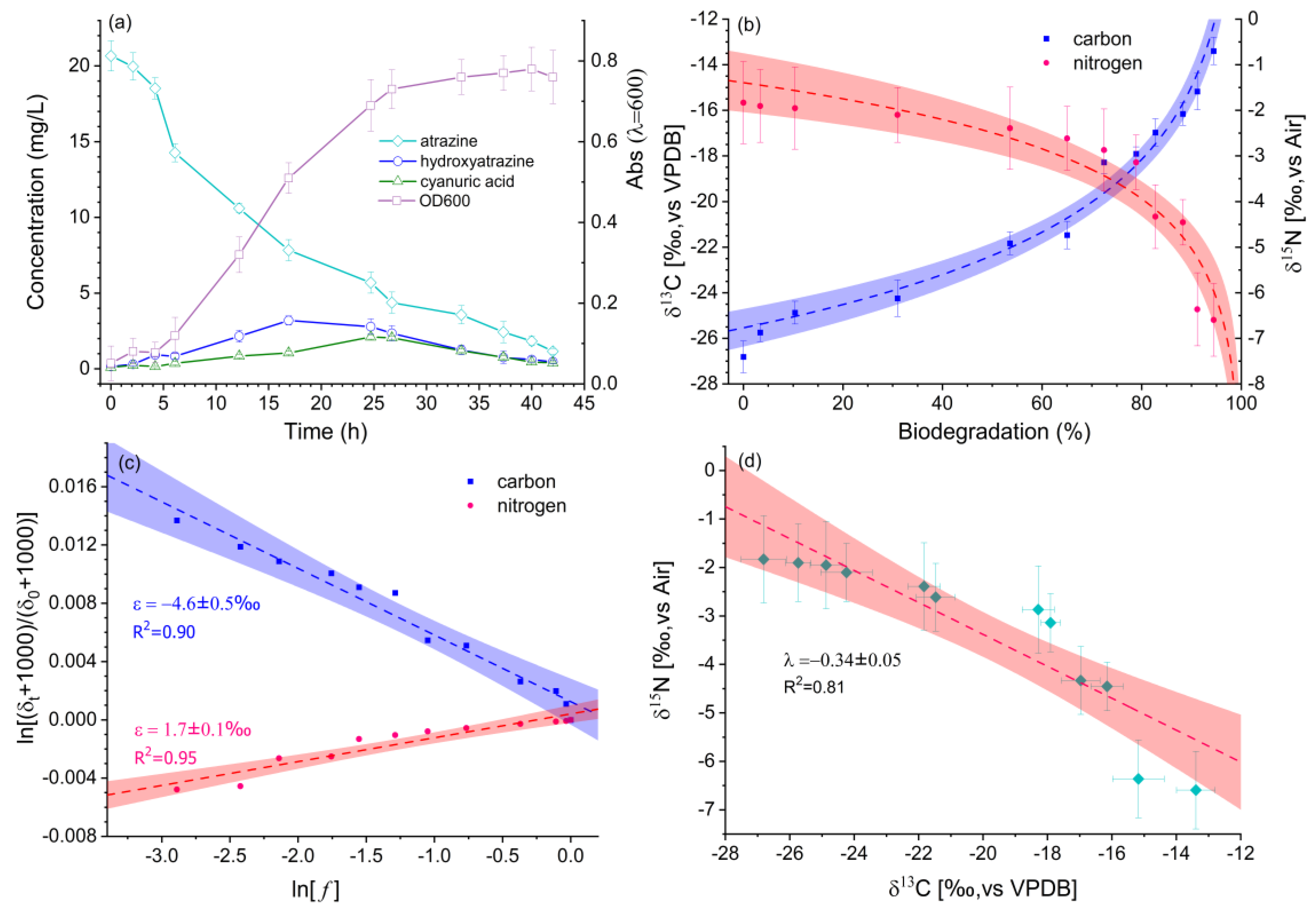
2.3. Cultivation of the Pore Water Taken from the Wetland System
2.4. Quantification of Atrazine and the Transformation Metabolites in Pore Water of the Wetland
2.5. Compound-Specific Carbon and Nitrogen Isotope Signature (δ13C, δ15N) Analysis of Atrazine
2.6. Calculation of Stable Isotope Fractionation
2.7. Quantitative Assessment of Biodegradation of Atrazine Based on CSIA
3. Results and Discussion
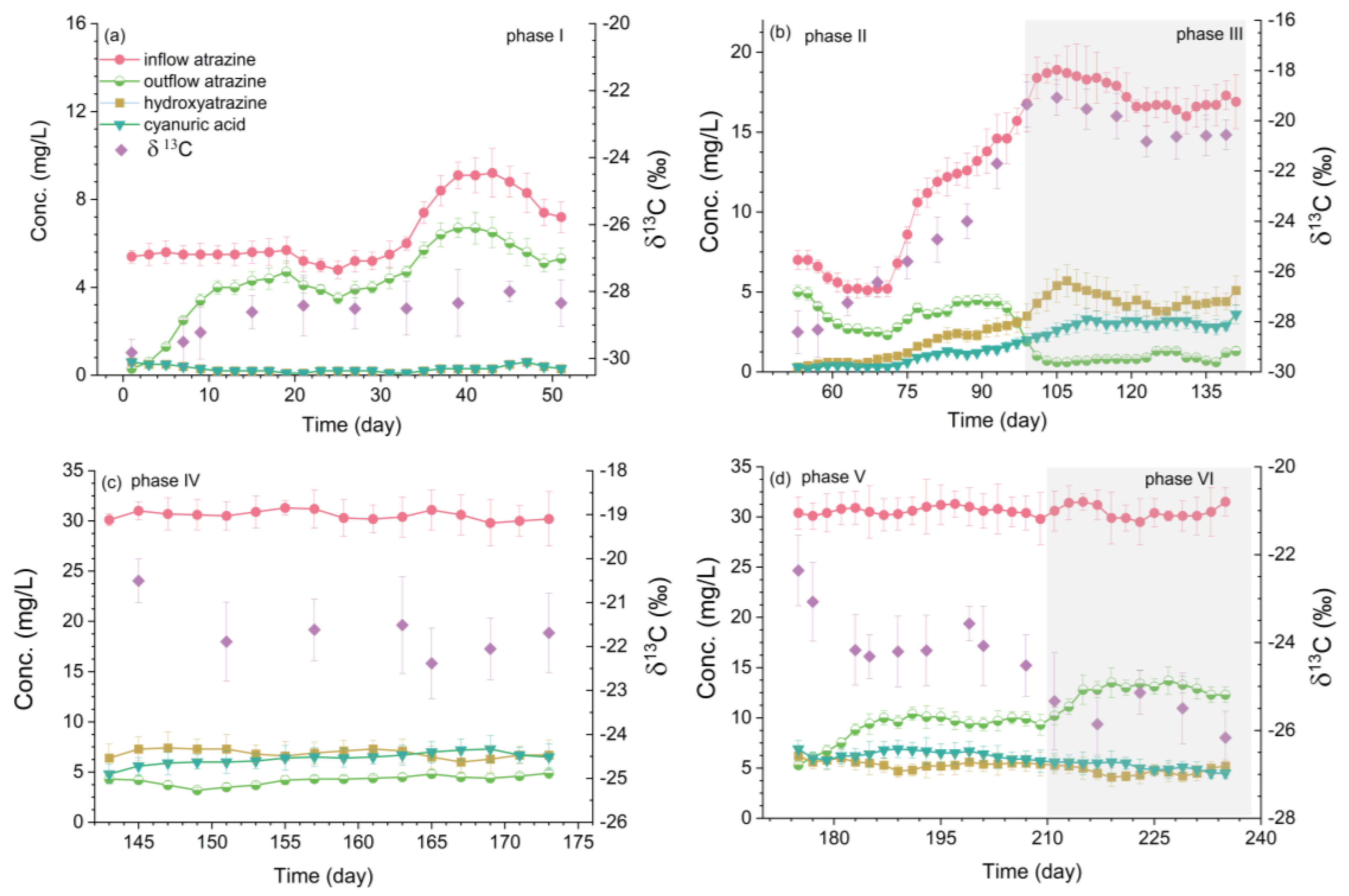
3.1. Atrazine Degradation and Carbon Isotope Signature Changes in the Wetland
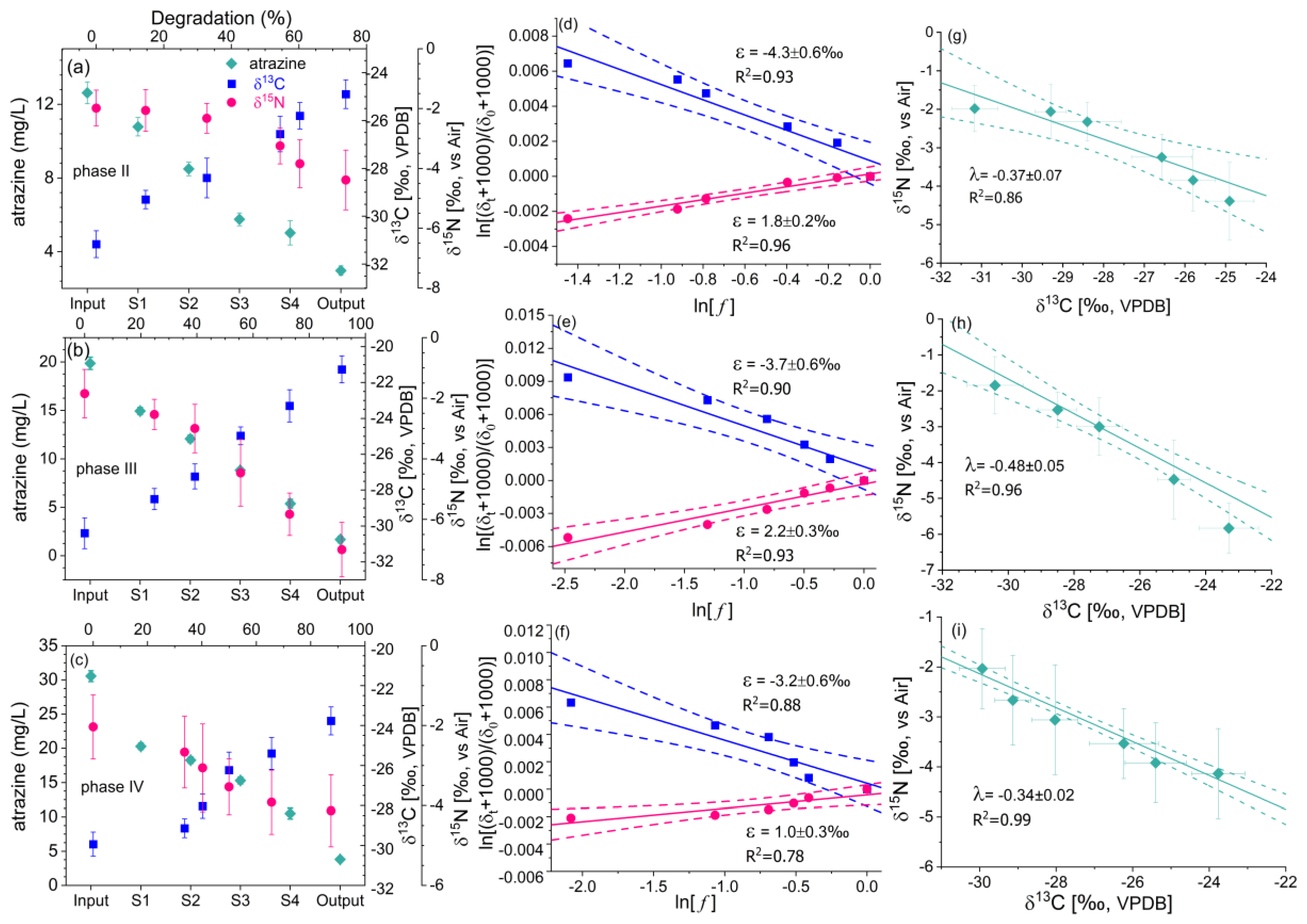
3.2. Characteristics of Carbon and Nitrogen Isotope Fractionation in Different Operational Phases
3.3. Reaction Pathway and Activation Mechanism of Atrazine Insight from Dual-Isotope Plots and Kinetic Isotope Effects
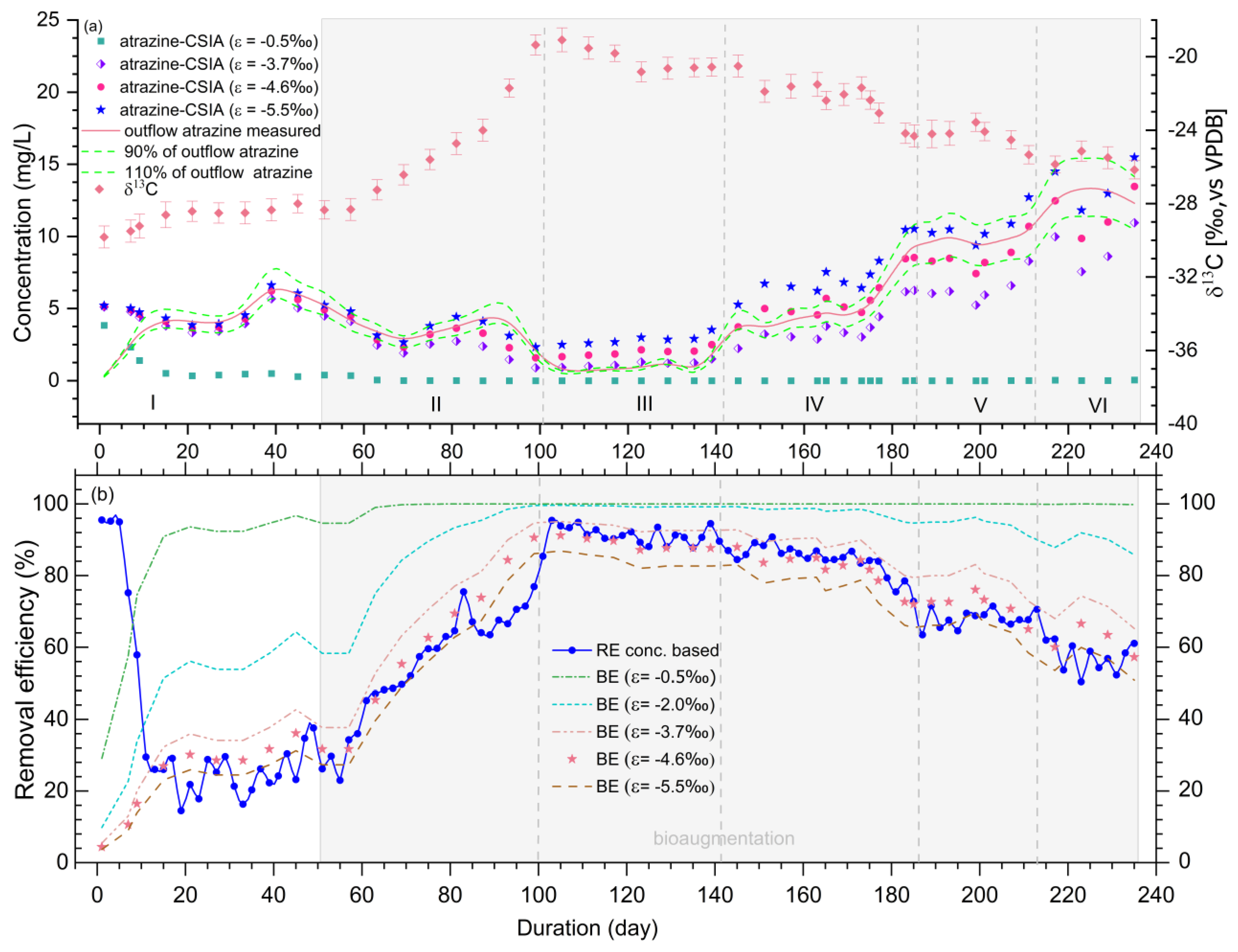
3.4. Quantification of Atrazine Biodegradation in the Wetland using Carbon Isotope Signatures
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- de Souza, R.M.; Seibert, D.; Quesada, H.B.; Bassetti, F.d.J.; Fagundes-Klen, M.R.; Bergamasco, R. Occurrence, impacts and general aspects of pesticides in surface water: A review. Process Saf. Environ. Prot. 2020, 135, 22–37. [Google Scholar] [CrossRef]
- Tang, F.H.M.; Lenzen, M.; McBratney, A.; Maggi, F. Risk of pesticide pollution at the global scale. Nat. Geosci. 2021, 14, 206–210. [Google Scholar] [CrossRef]
- Hu, Y.; Jiang, Z.; Hou, A.; Wang, X.; Zhou, Z.; Qin, B.; Cao, B.; Zhang, Y. Impact of atrazine on soil microbial properties: A meta-analysis. Environ. Pollut. 2023, 323, 121337. [Google Scholar] [CrossRef] [PubMed]
- Hennig, T.B.; Bandeira, F.O.; Puerari, R.C.; Fraceto, L.F.; Matias, W.G. A systematic review of the toxic effects of a nanopesticide on non-target organisms: Estimation of protective concentrations using a species sensitivity distribution (SSD) approach—The case of atrazine. Sci. Total Environ. 2023, 871, 162094. [Google Scholar] [CrossRef] [PubMed]
- Triassi, M.; Montuori, P.; Provvisiero, D.P.; De Rosa, E.; Di Duca, F.; Sarnacchiaro, P.; Díez, S. Occurrence and spatial-temporal distribution of atrazine and its metabolites in the aquatic environment of the Volturno River estuary, southern Italy. Sci. Total Environ. 2022, 803, 149972. [Google Scholar] [CrossRef]
- Vonberg, D.; Vanderborght, J.; Cremer, N.; Puetz, T.; Herbst, M.; Vereecken, H. 20 years of long-term atrazine monitoring in a shallow aquifer in western Germany. Water Res. 2014, 50, 294–306. [Google Scholar] [CrossRef]
- Urseler, N.; Bachetti, R.; Biole, F.; Morgante, V.; Morgante, C. Atrazine pollution in groundwater and raw bovine milk: Water quality, bioaccumulation and human risk assessment. Sci. Total Environ. 2022, 852, 158498. [Google Scholar] [CrossRef]
- Shukla, A.; Parde, D.; Gupta, V.; Vijay, R.; Kumar, R. A review on effective design processes of constructed wetlands. Int. J. Environ. Sci. Technol. 2022, 19, 12749–12774. [Google Scholar] [CrossRef]
- Palau, J.; Trueba-Santiso, A.; Yu, R.; Mortan, S.H.; Shouakar-Stash, O.; Freedman, D.L.; Wasmund, K.; Hunkeler, D.; Marco-Urrea, E.; Rosell, M. Dual C-Br Isotope Fractionation Indicates Distinct Reductive Dehalogenation Mechanisms of 1,2-Dibromoethane in Dehalococcoides- and Dehalogenimonas-Containing Cultures. Environ. Sci. Technol. 2023, 57, 1949–1958. [Google Scholar] [CrossRef]
- Ojeda, A.S.; Phillips, E.; Lollar, B.S. Multi-element (C, H, Cl, Br) stable isotope fractionation as a tool to investigate transformation processes for halogenated hydrocarbons. Environ. Ence Process. Impacts 2020, 22, 567–582. [Google Scholar] [CrossRef]
- Scott, K.M.; Lu, X.; Cavanaugh, C.M.; Liu, J.S. Optimal methods for estimating kinetic isotope effects from different forms of the Rayleigh distillation equation. Geochim. Et Cosmochim. Acta 2004, 68, 433–442. [Google Scholar] [CrossRef]
- Breukelen, B.M.V.; Prommer, H. Beyond the Rayleigh Equation: Reactive Transport Modeling of Isotope Fractionation Effects to Improve Quantification of Biodegradation. Environ. Sci. Technol. 2008, 42, 2457–2463. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.H.; Penning, H.; Elsner, M. C and N Isotope Fractionation Suggests Similar Mechanisms of Microbial Atrazine Transformation Despite Involvement of Different Enzymes (AtzA and TrzN). Environ. Sci. Technol. 2009, 43, 8079–8085. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.H.; Elsner, M. C-13/C-12 and N-15/N-14 Isotope Analysis To Characterize Degradation of Atrazine: Evidence from Parent and Daughter Compound Values. Environ. Sci. Technol. 2013, 47, 6884–6891. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liang, E.; Huang, T.; Zhao, X.; Wang, T. Insights into atrazine degradation by thermally activated persulfate: Evidence from dual C-H isotope analysis and DFT simulations. Chem. Eng. J. 2023, 454, 140207. [Google Scholar] [CrossRef]
- Liang, E.; Huang, T.; Li, J.; Wang, T. Degradation pathways of atrazine by electrochemical oxidation at different current densities: Identifications from compound-specific isotope analysis and DFT calculation. Environ. Pollut. 2023, 332, 121987. [Google Scholar] [CrossRef]
- Elsner, M.; Imfeld, G. Compound-specific isotope analysis (CSIA) of micropollutants in the environment—Current developments and future challenges. Curr. Opin. Biotechnol. 2016, 41, 60–72. [Google Scholar] [CrossRef]
- Ouyang, W.-Y.; Kümmel, S.; Adrian, L.; Zhu, Y.-G.; Richnow, H.H. Carbon and hydrogen stable isotope fractionation of sulfamethoxazole during anaerobic transformation catalyzed by Desulfovibrio vulgaris Hildenborough. Chemosphere 2023, 311, 136923. [Google Scholar] [CrossRef]
- Alvarez-Zaldívar, P.; Payraudeau, S.; Meite, F.; Masbou, J.; Imfeld, G. Pesticide degradation and export losses at the catchment scale: Insights from compound-specific isotope analysis (CSIA). Water Res. 2018, 139, 198–207. [Google Scholar] [CrossRef]
- Arar, M.; Bakkour, R.; Elsner, M.; Bernstein, A. Microbial hydrolysis of atrazine in contaminated groundwater. Chemosphere 2023, 322, 138226. [Google Scholar] [CrossRef]
- Nijenhuis, I.; Andert, J.; Beck, K.; Kästner, M.; Diekert, G.; Richnow, H.H. Stable isotope fractionation of tetrachloroethene during reductive dechlorination by Sulfurospirillum multivorans and Desulfitobacterium sp. strain PCE-S and abiotic reactions with cyanocobalamin. Appl. Environ. Microbiol. 2005, 71, 3413–3419. [Google Scholar] [CrossRef]
- Lee, P.K.H.; Conrad, M.E.; Alvarez-Cohen, L. Stable carbon isotope fractionation of chloroethenes by dehalorespiring isolates. Environ. Sci. Technol. 2007, 41, 4277–4285. [Google Scholar] [CrossRef]
- Harding, K.C.; Lee, P.K.H.; Bill, M.; Buscheck, T.E.; Conrad, M.E.; Alvarez-Cohen, L. Effects of Varying Growth Conditions on Stable Carbon Isotope Fractionation of Trichloroethene (TCE) by tceA-containing Dehalococcoides mccartyi strains. Environ. Sci. Technol. 2013, 47, 12342–12350. [Google Scholar] [CrossRef] [PubMed]
- Hermon, L.; Denonfoux, J.; Hellal, J.; Joulian, C.; Ferreira, S.; Vuilleumier, S.; Imfeld, G. Dichloromethane biodegradation in multi-contaminated groundwater: Insights from biomolecular and compound-specific isotope analyses. Water Res. 2018, 142, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Lihl, C.; Heckel, B.; Grzybkowska, A.; Dybala-Defratyka, A.; Ponsin, V.; Torrentó, C.; Hunkeler, D.; Elsner, M. Compound-specific chlorine isotope fractionation in biodegradation of atrazine. Environ. Ence Process. Impacts 2020, 2, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Ehrl, B.N.; Gharasoo, M.; Elsner, M. Isotope Fractionation Pinpoints Membrane Permeability as a Barrier to Atrazine Biodegradation in Gram-negative Polaromonas sp. Nea-C. Environ. Sci. Technol. 2018, 52, 4137–4144. [Google Scholar] [CrossRef]
- Chen, S.; Yang, P.; Rohit kumar, J.; Liu, Y.; Ma, L. Inconsistent carbon and nitrogen isotope fractionation in the biotransformation of atrazine by Ensifer sp. CX-T and Sinorihizobium sp. K. Int. Biodeterior. Biodegrad. 2017, 125, 170–176. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, K.; Jha, R.K.; Ma, L. Impact of atrazine concentration on bioavailability and apparent isotope fractionation in Gram-negative Rhizobium sp. CX-Z. Environ. Pollut. 2020, 257, 113614. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, K.; Jha, R.K.; Chen, C.; Yu, H.; Liu, Y.; Ma, L. Isotope fractionation in atrazine degradation reveals rate-limiting, energy-dependent transport across the cell membrane of gram-negative rhizobium sp. CX-Z. Environ. Pollut. 2019, 248, 857–864. [Google Scholar] [CrossRef]
- Kundu, K.; Marozava, S.; Ehrl, B.; Merl-Pham, J.; Griebler, C.; Elsner, M. Defining lower limits of biodegradation: Atrazine degradation regulated by mass transfer and maintenance demand in Arthrobacter aurescens TC1. ISME J. 2019, 13, 2236–2251. [Google Scholar] [CrossRef]
- Chen, S.; Ma, L.; Yao, G.; Wang, Y. Efficient atrazine removal in bioaugmentation constructed wetland: Insight from stable isotope fractionation analysis. Int. Biodeterior. Biodegrad. 2023, 185, 105691. [Google Scholar] [CrossRef]
- Cantu, R.; Evans, O.; Kawahara, F.K.; Shoemaker, J.A.; Dufour, A.P. An HPLC method with UV detection, pH control, and reductive ascorbic acid for cyanuric acid analysis in water. Anal. Chem. 2000, 72, 5820–5828. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.H.; Penning, H.; Lowag, H.; Elsner, M. Precise and Accurate Compound Specific Carbon and Nitrogen Isotope Analysis of Atrazine: Critical Role of Combustion Oven Conditions. Environ. Sci. Technol. 2008, 42, 7757–7763. [Google Scholar] [CrossRef] [PubMed]
- Coplen, T.B. Guidelines and recommended terms for expression of stable-isotope-ratio and gas-ratio measurement results. Rapid Commun. Mass Spectrom. 2011, 25, 2538–2560. [Google Scholar] [CrossRef] [PubMed]
- Elsner, M.; Zwank, L.; Hunkeler, D.; Schwarzenbach, R.P. A new concept linking observable stable isotope fractionation to transformation pathways of organic pollutants. Environ. Sci. Technol. 2005, 39, 6896–6916. [Google Scholar] [CrossRef]
- Runes, H.B.; Jenkins, J.J.; Moore, J.A.; Bottomley, P.J.; Wilson, B.D. Treatment of atrazine in nursery irrigation runoff by a constructed wetland. Water Res. 2003, 37, 539–550. [Google Scholar] [CrossRef]
- Hunkeler, D.; Meckenstock, R.U.; Lollar, B.S.; Schmidt, T.C.; Wilson, J.T. A Guide for Assessing Biodegradation and Source Identification of Organic Ground Water Contaminants Using Compound Specific Isotope Analysis (CSIA); U.S. Environmental Protection Agency: Washington, DC, USA, 2009; EPA/600/R-08/148.
- Braeckevelt, M.; Fischer, A.; KäStner, M. Field applicability of Compound-Specific Isotope Analysis (CSIA) for characterization and quantification of in situ contaminant degradation in aquifers. Appl. Microbiol. Biotechnol. 2012, 94, 1401–1421. [Google Scholar] [CrossRef]
- Zhao, X.; Bai, S.; Li, C.; Yang, J.; Ma, F. Bioaugmentation of atrazine removal in constructed wetland: Performance, microbial dynamics, and environmental impacts. Bioresour. Technol. 2019, 289, 121618. [Google Scholar] [CrossRef]
- Vymazal, J.; Bfezinova, T. The use of constructed wetlands for removal of pesticides from agricultural runoff and drainage: A review. Environ. Int. 2015, 75, 11–20. [Google Scholar] [CrossRef]
- Thullner, M.; Fischer, A.; Richnow, H.-H.; Wick, L.Y. Influence of mass transfer on stable isotope fractionation. Appl. Microbiol. Biotechnol. 2013, 97, 441–452. [Google Scholar] [CrossRef]
- Schuerner, H.K.V.; Seffernick, J.L.; Grzybkowska, A.; Dybala-Defratyka, A.; Wackett, L.P.; Elsner, M. Characteristic Isotope Fractionation Patterns in s-Triazine Degradation Have Their Origin in Multiple Protonation Options in the s-Triazine Hydrolase TrzN. Environ. Sci. Technol. 2015, 49, 3490–3498. [Google Scholar] [CrossRef] [PubMed]
- Grzybkowska, A.; Kaminski, R.; Dybala-Defratyka, A. Theoretical predictions of isotope effects versus their experimental values for an example of uncatalyzed hydrolysis of atrazine. Phys. Chem. Chem. Phys. 2014, 16, 15164–15172. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Makabe, A.; Yano, M.; Oshiki, M.; Kindaichi, T.; Casciotti, K.L.; Okabe, S. Dual nitrogen and oxygen isotope fractionation during anaerobic ammonium oxidation by anammox bacteria. Isme J. 2019, 13, 2426–2436. [Google Scholar] [CrossRef] [PubMed]
- Casciotti, K.L. Inverse kinetic isotope fractionation during bacterial nitrite oxidation. Geochim. Et Cosmochim. Acta 2009, 73, 2061–2076. [Google Scholar] [CrossRef]
- Peat, T.S.; Newman, J.; Balotra, S.; Lucent, D.; Warden, A.C.; Scott, C. The structure of the hexameric atrazine chlorohydrolase AtzA. Acta Crystallogr. Sect. D-Struct. Biol. 2015, 71, 710–720. [Google Scholar] [CrossRef]
- Melsbach, A.; Torrento, C.; Ponsin, V.; Bolotin, J.; Lachat, L.; Prasuhn, V.; Hofstetter, T.B.; Hunkeler, D.; Elsner, M. Dual-Element Isotope Analysis of Desphenylchloridazon to Investigate Its Environmental Fate in a Systematic Field Study: A Long-Term Lysimeter Experiment. Environ. Sci. Technol. 2020, 54, 3929–3939. [Google Scholar] [CrossRef]
- Palau, J.; Yu, R.; Hatijah Mortan, S.; Shouakar-Stash, O.; Rosell, M.; Freedman, D.L.; Sbarbati, C.; Fiorenza, S.; Aravena, R.; Marco-Urrea, E.; et al. Distinct Dual C-Cl Isotope Fractionation Patterns during Anaerobic Biodegradation of 1,2-Dichloroethane: Potential To Characterize Microbial Degradation in the Field. Env. Sci Technol 2017, 51, 2685–2694. [Google Scholar] [CrossRef]
- Fischer, A.; Gehre, M.; Breitfeld, J.; Richnow, H.H.; Vogt, C. Carbon and hydrogen isotope fractionation of benzene during biodegradation under sulfate-reducing conditions: A laboratory to field site approach. Rapid Commun. Mass Spectrom. 2009, 23, 2439–2447. [Google Scholar] [CrossRef]
- Haderlein, S.B.; Schmidt, T.C.; Elsner, M.; Zwank, L.; Berg, M.; Schwarzenbach, R.P. New Evaluation Scheme for Two-Dimensional Isotope Analysis to Decipher Biodegradation Processes: Application to Groundwater Contamination by MTBE. Environ. Ence Technol. 2005, 39, 8543–8544. [Google Scholar] [CrossRef]
- Chen, S.; Ma, L.; Wang, Y. Kinetic isotope effects of C and N indicate different transformation mechanisms between atzA- and trzN-harboring strains in dechlorination of atrazine. Biodegradation 2022, 33, 207–221. [Google Scholar] [CrossRef]
- Meyer, A.H.; Dybala-Defratyka, A.; Alaimo, P.J.; Geronimo, I.; Sanchez, A.D.; Cramer, C.J.; Elsner, M. Cytochrome P450-catalyzed dealkylation of atrazine by Rhodococcus sp. strain N186/21 involves hydrogen atom transfer rather than single electron transfer. Dalton Trans. 2014, 43, 12175–12186. [Google Scholar] [CrossRef] [PubMed]
- Dybala-Defratyka, A.; Szatkowski, L.; Kaminski, R.; Wujec, M.; Siwek, A.; Paneth, P. Kinetic Isotope Effects on Dehalogenations at an Aromatic Carbon. Environ. Sci. Technol. 2008, 42, 7744–7750. [Google Scholar] [CrossRef] [PubMed]
- Manna, R.N.; Grzybkowska, A.; Gelman, F.; Dybala-Defratyka, A. Carbon-bromine bond cleavage—A perspective from bromine and carbon kinetic isotope effects on model debromination reactions. Chemosphere 2018, 193, 17–23. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Li, L. Density functional theory calculations of nitrogen and oxygen equilibrium isotope fractionations in ––H2O aqueous system reveal inverse kinetic isotope effects during nitrite oxidation. Appl. Geochem. 2022, 139, 105265. [Google Scholar] [CrossRef]
- Hofstetter, T.B.; Berg, M. Assessing transformation processes of organic contaminants by compound-specific stable isotope analysis. Trac-Trends Anal. Chem. 2011, 30, 618–627. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phases | Dates | Duration (d) | HRT (d) | Inflow (L/d) | Inflow Atrazine Range (mg/L) | Inflow Atrazine Average (mg/L) |
|---|---|---|---|---|---|---|
| Ⅰ | 1st March to 20th April | 51 | 6 | 64 | 4.1 to 9.3 | 6.3 ± 1.4 |
| Ⅱ * | 21st April to 7th June | 46 | 6 | 64 | 4.9 to 15.6 | 9.9 ± 3.9 |
| Ⅲ | 8th June to 19th July | 42 | 6 | 64 | 15.6 to 19.3 | 17.4 ± 1.2 |
| Ⅳ | 20th July to 20th August | 32 | 6 | 64 | 28.4 to 31.9 | 30.4 ± 1.1 |
| Ⅴ | 21st August to 29th September | 40 | 4 | 96 | 29.1 to 32.0 | 30.6 ± 0.8 |
| Ⅵ | 30th September to 21st October | 29 | 2 | 192 | 28.8 to 32.8 | 30.5 ± 1.2 |
| Phase | Atz-Outflow Aver. (mg/L) | Atz-Outflow Range (mg/L) | RE Aver. (%) | RE Range (%) | δ13C-Aver. (‰) | δ13C-Range (‰) | BE-CSIA Aver. (%) (ε = −4.6‰) | BE-CSIA Range (%) |
|---|---|---|---|---|---|---|---|---|
| Ⅰ | 4.8 ± 1.1 | 2.9–7.0 | 26.9 ± 6.9 | 14.5–37.6 | −28.8 ± 0.7 | −30.0–−28.0 | 32.1 ± 10.5 | 11.6–43.8 |
| Ⅱ | 3.5 ± 0.9 | 2.0–4.6 | 60.9 ± 8.7 | 45.2–75.5 | −24.8 ± 2.6 | −27.6–−20.0 | 68.7 ± 15.1 | 44.5–90.6 |
| Ⅲ | 1.5 ± 0.4 | 0.9–2.6 | 90.6 ± 4.1 | 76.9–95.4 | −20.1 ± 1.1 | −21.6–−18.8 | 90.1 ± 2.6 | 86.5–92.8 |
| Ⅳ | 4.2 ± 0.6 | 2.8–4.9 | 86.7 ± 2.0 | 84.4–90.8 | −21.5 ± 1.1 | −22.7–−19.5 | 86.5 ± 2.9 | 82.8–91.5 |
| Ⅴ | 8.8 ± 1.9 | 4.9–11.3 | 72.0 ± 6.6 | 63.5–84.2 | −23.8 ± 0.9 | −25.0–−22.0 | 77.5 ± 4.7 | 71.4–85.3 |
| Ⅵ | 13.0 ± 0.9 | 12.0–14.3 | 58.4 ± 5.5 | 50.4–70.6 | −25.7 ± 0.7 | −26.3–−24.8 | 66.2 ± 5.0 | 61.7–72.2 |
| Phases | ε (C)-‰ | ε (N)-‰ | AKIE-C | AKIE-N | λ = δ15N vs. δ13C |
|---|---|---|---|---|---|
| Ⅱ | −4.3 ± 0.6 | 1.8 ± 0.2 | 1.036 ± 0.005 | 0.991 ± 0.001 | −0.37 ± 0.07 |
| Ⅲ | −3.7 ± 0.6 | 2.2 ± 0.3 | 1.031 ± 0.005 | 0.989 ± 0.001 | −0.48 ± 0.05 |
| Ⅳ | −3.2 ± 0.6 | 1.0 ± 0.3 | 1.026 ± 0.005 | 0.995 ± 0.001 | −0.34 ± 0.02 |
| Batch | −4.6 ± 0.5 | 1.7 ± 0.1 | 1.038 ± 0.004 | 0.992 ± 0.000 | −0.34 ± 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Wang, Y.; Ma, L. Assessing Biodegradation Processes of Atrazine in Constructed Wetland Using Compound-Specific Stable Isotope Analysis. Processes 2023, 11, 3252. https://doi.org/10.3390/pr11113252
Chen S, Wang Y, Ma L. Assessing Biodegradation Processes of Atrazine in Constructed Wetland Using Compound-Specific Stable Isotope Analysis. Processes. 2023; 11(11):3252. https://doi.org/10.3390/pr11113252
Chicago/Turabian StyleChen, Songsong, Yuncai Wang, and Limin Ma. 2023. "Assessing Biodegradation Processes of Atrazine in Constructed Wetland Using Compound-Specific Stable Isotope Analysis" Processes 11, no. 11: 3252. https://doi.org/10.3390/pr11113252